Introduction
Osteosarcoma is the most common primary bone
malignancy in children and adolescents (1), which usually occurs in the metaphyseal
region of tubular long bones (2).
Although numerous modern therapies, including surgery, chemotherapy
and radiotherapy have been used for treating osteosarcoma, the
5-year survival rate of patients with osteosarcoma is 60–70%
(3). With the improvements in
treatment techniques, the mortality rate of osteosarcoma is
decreasing at ~1.3% per year (4).
However, the etiology of osteosarcoma is unclear. Therefore,
identification of the key genes associated with osteosarcoma is
important for treatment of the disease.
As a type of plant polyphenol, curcumin is extracted
from Curcuma longa (5) and
exhibits anti-tumor, anti-inflammatory, antioxidant and
anticoagulation capacities (6–9). Previous
studies have demonstrated that curcumin serves functions in the
progression of a number of types of cancer, including osteosarcoma
(10), pancreatic cancer (11) and colon cancer (12). Fossey et al (13) revealed that FLLL32, a novel compound
obtained from curcumin, inhibited signal transducers and activators
of transcription 3 (STAT3) and induced apoptosis in
osteosarcoma. Chang et al (14) indicated that curcumin induced
apoptosis in the osteosarcoma MG63 cell line via mediating the
reactive oxygen species/cytochrome c/caspase-3 pathway. Leow et
al (15) demonstrated that
curcumin and PKF118-310 may delay tumorigenesis and metastasis of
osteosarcoma through inhibiting the Wnt/β-catenin signaling
pathway. Taken together, these studies indicate that curcumin
exhibits an effect on the pathogenesis of osteosarcoma through
different pathways. Despite these aforementioned data, the target
genes of curcumin in osteosarcoma remain unclear.
RNA sequencing (RNA-seq), which is based on
deep-sequencing technology, is a more developed approach compared
with transcriptome profiling (16).
Several developments in RNA-seq, including mapping of the
transcription start site, characterization of small RNA and
detection of strand-specific, gene fusion and alternative splicing
events, have been identified (17).
In the present study, RNA-seq data analysis was performed on human
osteosarcoma U-2 OS cells, which were treated by curcumin or
dimethyl sulfoxide (DMSO). Differentially expressed genes (DEGs)
between the curcumin-treated and control groups were identified,
and the functions of the DEGs were predicted by functional and
pathway enrichment analyses. A protein-protein interaction (PPI)
network involving the DEGs was also constructed to additionally
determine the target genes of curcumin in osteosarcoma. In
addition, the mRNA levels of the top 10 nodes in the PPI network
were confirmed, and it was identified that the mRNA levels of
clathrin heavy chain (CLTC) and inositol 1,4,5-triphosphate
receptor type 1 (ITPR1) in curcumin-treated cells were
significantly increased. In addition, the effects of curcumin and
ITPR1 on proliferation, apoptosis, migration and invasion in
human osteosarcoma U-2 OS cells were investigated.
Materials and methods
Cell cultivation and curcumin
treatment
The human osteosarcoma U-2 OS cell line was
purchased from Cell Bank of Chinese Academy of Sciences (Shanghai,
China). The U-2 OS cells were inoculated in RPMI-1640 medium
(Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.) and 1% penicillin-streptomycin (Gibco; Thermo
Fisher Scientific, Inc.) and cultured at 37°C in a 5%
CO2 incubator (Thermo Fisher Scientific, Inc.). When the
passaged cells reached 80–90% confluence, the cells were digested
using pancreatin (Gibco; Thermo Fisher Scientific, Inc.). The
digested cells were centrifuged at 1,000 × g and 4°C for 5 min, and
then supernatant was discarded. Subsequently, the cells were
preserved in frozen stock solution containing 10% DMSO
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany), 40% FBS and 50%
RPMI-1640 medium, and stored in program frozen box.
The U-2 OS cells were seeded in 6-well plates
(2×106 cell/well) and incubated in 5 ml serum-free
medium in a 5% CO2 incubator (Thermo Fisher Scientific,
Inc.) at 37°C overnight. Curcumin (Sigma-Aldrich; Merck KGaA, 15
µmol/l) was dissolved in DMSO. Subsequently, U-2 OS cells were
treated with 15 µM curcumin at 37°C in a 5% CO2
incubator for 48 h (curcumin group). Concurrently, in the control
groups, U-2 OS cells were treated with an equal volume of DMSO.
Finally, the cells were washed twice with cold PBS.
All studies were approved by the Scientific and
Ethical Committee of the 309th Hospital of Chinese People's
Liberation Army (Beijing, China) and performed in accordance with
the ethical standards.
RNA sequencing data
Total RNA was isolated from the U-2 OS cells using
TRIzol® (Takara Bio, Inc., Otsu, Japan) according to the
manufacturer's protocol and quantified by spectrophotometry. Then,
a transcriptome library was constructed using NEBNext®
Ultra™ RNA Library Prep kit for Illumina® (cat. no.
E7530; New England Biolabs, Inc., Ipswich, MA, USA) following the
manufacturer's protocol. RNA was separated into RNA fragments (~200
nt), followed by double-stranded cDNA being synthesized and
end-repaired. Then, adaptor ligation and polymerase chain reaction
(PCR) enrichment (denaturation step: 98°C for 30 sec; 12 cycles:
98°C for 10 sec and annealing temperature 65°C for 75 sec; final
extension step: 65°C for 5 min) were performed using the primers
and reagents supplied with the NEBNext® Ultra RNA
Library Prep kit. The quality of the library was analyzed using
Bioanalyzer 2100 (Agilent Technologies, Inc., Santa Clara, CA,
USA), and RNA was sequenced on Hiseq 2500 (Illumina, Inc., San
Diego, CA, USA).
Data quality control and screening of
DEGs
Subsequent to quality controls of the reads, they
were mapped to the reference human genome (hg19) using TopHat
(18) and assembled by Bowtie 1
software (19). Combined with
sequence alignment results from TopHat and hg19 annotation
information downloaded from the University of California, Santa
Cruz (UCSC) database (https://www.astro.ucsc.edu/) (20), the DEGs between curcumin and control
groups were identified using Cufflink software (21). P<0.01 and |log2 fold
change (FC)| ≥1 were set as cut-off criteria.
Functional and pathway enrichment
analysis
Gene Ontology (GO, http://geneontology.org/) analysis develops a series
of controlled and structured vocabularies for annotating functions
of genes and their products (22).
The Kyoto Encyclopedia of Genes and Genomes (KEGG; http://www.kegg.jp/) is a biological database, which
stores genomic, chemical and systemic functional information
(23). Using the cluster profiler
(24) package in R, GO functional and
KEGG pathway enrichment analyses were performed for upregulated and
downregulated genes, respectively. The raw P-value was adjusted by
the Benjamini-Hochberg method (25),
and the adjusted P-value (also known as false discovery rate, FDR)
<0.05 was used as a threshold.
Construction of PPI network
Human Protein Reference Database (HPRD) includes
curated proteomic information and describes human PPIs (26). Using HPRD Release 9 (http://www.hprd.org/query) (26), pairs of interacting human proteins
were downloaded, and the self-interacting protein pairs were
removed. The DEGs were mapped with the downloaded PPI pairs, and
those pairs involving the identified DEGs were extracted. Finally,
the PPI network was visualized using Cytoscape software (version
2.8.0; www.cytoscape.org) and named as the DEG
PPI network (27). Proteins are
represented by nodes, the degree is the number of nodes which
interact with the node in question, and the higher the degree the
more important the protein in the PPI network.
Quantitative reverse transcription
polymerase chain reaction (RT-qPCR) analysis
The cells treated with curcumin were dissolved in
TRIzol® reagent (Takara Bio, Inc.) for extraction of
total RNA. Primers for key genes are summarized in Table I. Using SYBR® Premix Ex
Taq™ (Applied Biosystems; Thermo Fisher Scientific, Inc.), RT-qPCR
amplification was performed according to the following
thermocycling conditions: 50°C for 3 min, 95°C for 3 min, followed
by 40 cycles of 95°C for 10 sec and 58°C for 30 sec. Gene
expression was quantified and calculated by the comparative
threshold (Cq) cycle method (2−ΔΔCq) (28).
 | Table I.Primer sequences for specific
genes. |
Table I.
Primer sequences for specific
genes.
| Name of primer | Primer sequences
(5′-3′) |
|---|
| EEF1A1-hf |
TGCCTGGGTCTTGGATAAAC |
| EEF1A1-hr |
GCCTGAGATGTCCCTGTAAT |
| ATF7IP-hf |
TTCCGCCCCAAAAGATTCAGA |
| ATF7IP-hr |
CTGCTTCAAGTTGCTGACGATC |
| HIF1A-hf |
ACTTCTGGATGCTGGTGATT |
| HIF1A-hr |
GTTCAAACTGAGTTAATCCC |
| SMAD7-hf |
ACCTTAGCCGACTCTGCGAACT |
| SMAD7-hr |
TTTCAGCGGAGGAAGGCACA |
| CLTC-hf |
TTGAATACGGTTGCTCTTGT |
| CLTC-hr |
ATGCCAGTCAGAAGTAACCA |
| MCM10-hf |
CTTTGAATACGGTTGCTCTT |
| MCM10-hr |
GTACGGTAATTGATAATCTGG |
| ITPR1-hf |
CCTGGTTGATGATCGTTGTGTT |
| ITPR1-hr |
GCTTTTGGGCAGAGTAGCGGTT |
| ADAM15-hf |
ATCCCTGCTGTGATTCTTTGACC |
| ADAM15-hr |
TGGGCATAGGAGGCACAACG |
| WWP2-hf |
CCCCGAATCCCAACACGACT |
| WWP2-hr |
TCCCATCCAGCAGGCAGAGC |
| ATP5C1-hf |
AAAAGCGAGGTTGCTACACT |
| ATP5C1-hr |
ATGACTGACGCATCTCCAAA |
| GAPDH-hf |
TGACAACTTTGGTATCGTGGAAGG |
| GAPDH-hr |
AGGCAGGGATGATGTTCTGGAGAG |
RNA interference assay
The cells in the siRNA group were digested by
pancreatin at 37°C for 5 min, and then complete medium was added to
neutralize the pancreatin. Subsequently, the cells were centrifuged
at 1,000 × g for 5 min at 4°C, and the supernatant was discarded.
Subsequent to being counted, the cells were seeded in 6-well plates
(8×105 cell/well) and cultured in 2 ml antibiotic-free
complete medium in a 5% CO2 incubator (Thermo Fisher
Scientific, Inc.) at 3°C overnight. The ITPR1 small interfering
(siRNA) were synthesized with the forward,
5′-AGACAGAAAACAGGAAAUUTT-3′ and reverse primer,
(5′-AAUUUCCUGUUUUCUGUCUCA-3′). The ITPR1 siRNA (final
concentration, 66 nM/l) and Lipofectamine® 2000 reagent
(1:25 vol/vol; Invitrogen; Thermo Fisher Scientific, Inc.) diluted
with Opti-MEM medium (Gibco; Thermo Fisher Scientific, Inc.) were
mixed and allowed to rest for 20 min. Subsequently, the cells were
added to the siRNA mixture (500 µl/well) and Opti-MEM medium was
replaced with complete medium following transfection for 6 h at
room temperature. In addition, the cells were cultivated in a 5%
CO2 incubator (Thermo Fisher Scientific, Inc.) at 37°C
for 48 h. The cells were then collected for RT-qPCR assay. The
cells in the negative control group were treated with scrambled
siRNA (final concentration, 66 nM/l) (Shanghai Biotend Biological
Technology, Co., Ltd., Shanghai, China) as control and cultured
like the cells in the siRNA group. The cells in the control group
were treated without siRNA but otherwise culture the same as the
cells in the siRNA group.
Cell proliferation assay
Subsequent to being digested and counted, the cells
were seeded in a 96-well plate (4×104 cell/well) and
cultured in 100 ml antibiotic-free RPMI-1640 medium supplemented
with 10% FBS (both FBS; Gibco; Thermo Fisher Scientific, Inc.) in a
5% CO2 incubator (Thermo Fisher Scientific, Inc.) at
37°C overnight. Then, the cells were separately transfected at room
temperature with the siRNA mixture, negative control siRNA and
Lipofectamine 2000 reagent (Invitrogen; ThermoFisher Scientific
Inc.) for 6 h. Subsequent to treatment with curcumin
(Sigma-Aldrich; Merck KGaA), the cells were incubated with cell
counting kit-8 at 37°C (CCK8; 10 µl/well; Dojindo Molecular
Technologies, Inc., Kumamoto, Japan) for 2 h. Finally, the
absorption values under 450 nm were measured using an
Epoch™ Microplate Spectrophotometer (BioTek Instruments,
Inc., Winooski, VT, USA) to calculate viability.
Flow cytometry
The U-2 OS cells were digested and counted, followed
by seeding in a 6-well plate (8×105 cell/well) and
cultivation in 2 ml antibiotic-free RPMI-1640 (Gibco; Thermo Fisher
Scientific, Inc.) in a 5% CO2 at 37°C overnight.
Following transfection at room temperature for 6 h, the cells were
treated with curcumin. The cells were digested by pancreatin,
washed by PBS and resuspended in 1X binding buffer (BD Biosciences,
San Jose, CA, USA). Subsequently, 100 µl of the above solution was
transferred to flow cytometry tube, and stained with 5 µl
fluorescein is isothiocyanate-Annexin V (BD Biosciences) and 5 µl
propidium iodide (50 µg/ml; BD Biosciences) in the dark for 15 min
at 25°C. Additionally, 400 µl 1X binding buffer (BD Biosciences)
was added prior to flow cytometry using FCS Express 4 (De Novo
Software, Glendale, CA).
Wound healing assay
Lines were drawn on the base of 6-well plate with an
equal interval of ~0.5–1 cm. Then, the digested and counted cells
were seeded in a 6-well plate (8×105 cell/well) and
cultured in 2 ml RPMI-1640 medium without serum (Gibco; Thermo
Fisher Scientific, Inc.) in a 5% CO2 at 37°C overnight.
Then, the cells were separately transfected at room temperature
with the siRNA mixture, negative control siRNA and Lipofectamine
2000 reagent (Invitrogen; Thermo Fisher Scientific Inc.) for 6 h.
Afterwards, the cells were wound by drawing lines using a pipetting
needle. After this, the cells were washed with PBS and then there
were several channels on the surface of cultured cells. Subsequent
to treatment with 15 µmol/l curcumin, images of movement of the
cells were captured every 12 h and measured by Image-Pro Plus 6.0
software (Media Cybernetics Inc., Rockville, MD, USA).
Transwell assay
Subsequent to dilution with PBS (1:3 dilution), 50
µl Matrigel (1 mg/ml; Corning Incorporated, Corning, NY, USA) was
solidified in a 24-well polycarbonate membrane following standing
for 1 h at 37°C and resuspended following addition of 200 µl
RPMI-1640 without serum (Gibco; Thermo Fisher Scientific, Inc.).
The cells in each well of cell petri dish were added with 600 µl
10% FBS (Gibco; Thermo Fisher Scientific, Inc.) and then were
cultured in a 6-well plate (8×105 cell/well) in 2 ml
RPMI-1640 medium with 10% FBS (Gibco; Thermo Fisher Scientific,
Inc.) in 5% CO2 at 37°C overnight. Following this, the
transferred and treated cells were digested by pancreatin (Gibco:
Thermo Fisher Scientific, Inc.) and cultivated in the upper wells
(1×105 cell/well) in a 5% CO2 incubator
(Thermo Fisher Scientific. Inc.) at 37°C for 12 h. Finally, the
liquids in upper wells were discarded, and the cells that had not
passed through the membranes were wiped off. The cells were stained
with 0.1% crystal violet staining at room temperature for 20 min
and observed under an inverted light microscope in 3 fields of view
at magnification, ×200 (Olympus Corporation, Tokyo, Japan).
Statistical analysis
Using SPSS 12.0 software (SPSS Inc., Chicago, IL,
USA), comparisons between groups were made using one-way ANOVA
followed by a least significant difference test. All results are
presented as the mean ± standard error. P<0.05 was considered to
indicate a statistically significant difference.
Results
Data quality control and analysis of
DEGs
Quality control of the reads was performed and then
assembled using Bowtie 1 software. The results are summarized in
Table II. With P<0.01 and
|log2 FC| ≥1 as thresholds, the DEGs between the
curcumin and control group were identified by Cufflink software.
Compared with the control group, a total of 201 DEGs were
identified in the curcumin-treated group, including 114 upregulated
and 87 downregulated genes.
| Table II.Results from quality control and
assembly of sequencing data. |
Table II.
Results from quality control and
assembly of sequencing data.
| ID | Group | Number of cleaned
paired-end reads | Number of aligned
pair reads |
|---|
| 14710C-1 | Control | 11,660,638 | 8,129,145 |
| 14710C-2 | Control | 12,351,701 | 8,531,536 |
| 14710C-5 | Curcumin | 13,820,514 | 8,368,710 |
| 14710C-6 | Curcumin | 11,927,581 | 8,797,461 |
Functional and pathway enrichment
analyses
The upregulated genes were significantly enriched in
3 GO terms, including ‘biological process’
(FDR=3.75×10−6), ‘cellular component organization’
(FDR=1.95×10−2) and ‘cellular component organization or
biogenesis’ (FDR=2.08×10−2; Table III). The downregulated genes
including hypoxia-inducible factor-1α (HIF1A) were significantly
enriched in signal transduction (FDR=1.03×10−2), cell
communication (FDR=1.89×10−2) and cellular process
(FDR=5.62×10−3; Table
III). In addition, there were no KEGG pathways significantly
enriched for the upregulated or downregulated genes. Importantly,
the GO term ‘biological process’ was enriched for upregulated genes
[eukaryotic translation elongation factor 1α1 (EEF1A1), activating
transcription factor 7 interacting protein (ATF7IP), SMAD family
member 7 (SMAD7), CLTC, minichromosome maintenance 10 (MCM10),
ITPR1, WW domain containing E3 ubiquitin protein ligase 2 (WWP2)
and ATP synthase, H+ transporting, mitochondrial F1
complex, gamma polypeptide 1 (ATP5C1)], and downregulated genes
[HIF1A and a disintegrin and metalloprotease 15 (ADAM15)].
| Table III.GO terms enriched for upregulated and
downregulated genes. |
Table III.
GO terms enriched for upregulated and
downregulated genes.
| Go term | Description | Gene number | FDR | Gene symbol |
|---|
| Upregulated
DEGs |
|
|
|
|
| GO:
8150 | Biological
process | 94 | 3.75×10-6 | EEF1A1, ATF7IP,
SMAD7, CLTC, MCM10 |
| GO:
16043 | Cellular component
organization | 41 | 1.95×10-2 | ATF7IP, SMAD7,
CLTC, CDH4, RTN3 |
| GO:
71840 | Cellular component
organization or biogenesis | 41 | 2.08×10-2 | ATF7IP, SMAD7,
CLTC, CDH4, RTN3 |
| Downregulated
DEGs |
|
|
|
|
| GO:
8150 | Biological
process | 64 | 8.18×10-4 | HIF1A, ADAM15,
ARFRP1, AKAP9, ABI2 |
| GO:
50896 | Response to
stimulus | 40 | 5.62×10-3 | HIF1A, ADAM15,
ARFRP1, AKAP9, MALT1 |
| GO:
51716 | Cellular response
to stimulus | 34 | 5.62×10-3 | HIF1A, ARFRP1,
AKAP9, MALT1, MGLL |
| GO:
9987 | Cellular
process | 59 | 5.62×10-3 | HIF1A, ADAM15,
ARFRP1,RBM5, PFDN6 |
| GO:
7165 | Signal
transduction | 29 | 1.03×10-2 | HIF1A, ARFRP1,
AKAP9, MALT1, RAP1GAP2 |
| GO:
23052 | Signaling | 30 | 1.79×10-2 | HIF1A, ARFRP1,
AKAP9, MALT1, RAP1GAP2 |
| GO:
44700 | Single organism
signaling | 30 | 1.79×10-2 | HIF1A, ARFRP1,
AKAP9, MALT1, RAP1GAP2 |
| GO:
7154 | Cell
communication | 30 | 1.89×10-2 | HIF1A, ARFRP1,
AKAP9, MALT1, RAP1GAP2 |
| GO:
71704 | Organic substance
metabolic process | 46 | 1.89×10-2 | HIF1A, ADAM15,
ARFRP1,GSTO2, ADK |
| GO:
44238 | Primary metabolic
process | 45 | 1.89×10-2 | HIF1A, ADAM15,
ARFRP1,GSTO2, ADK |
| GO:
50789 | Regulation of
biological process | 43 | 2.04×10-2 | HIF1A, ARFRP1,
AKAP9, ABI2, RBM5 |
| GO:
50794 | Regulation of
cellular process | 41 | 2.55×10-2 | HIF1A, ARFRP1,
AKAP9, ABI2, RBM5 |
| GO:
44699 | Single-organism
process | 53 | 2.98×10-2 | HIF1A, ADAM15,
ARFRP1, AKAP9, ABI2 |
| GO:
1568 | Blood vessel
development | 6 | 4.65×10-2 | HIF1A, ADAM15,
ANKRD17, RAPGEF1, NPRL3, NRP1 |
| GO:
43170 | Macromolecule
metabolic process | 38 | 4.98×10-2 | HIF1A, ADAM15,
ABI2, RBM5, PFDN6 |
Analysis of PPI network
A total of 39,240 PPI pairs were downloaded from
HPRD Release 9. The DEG.PPI network had 929 interactions and 913
node genes, including 73 upregulated, 43 downregulated and 797
genes with no significant change in expression (Fig. 1). In particular, EEF1A1 (degree=88),
ATF7IP (degree=64), HIF1A (degree=44), SMAD7 (degree=43), CLTC
(degree=42), MCM10 (degree=28), ITPR1 (degree=27), ADAM15
(degree=24), WWP2 (degree=22) and ATP5C1 (degree=21) exhibited
higher degrees in the DEG.PPI network.
Effects of curcumin treatment on the
mRNA levels of EEF1A1, ATF7IP, HIF1A, SMAD7, CLTC, MCM10, ITPR1,
ADAM15, WWP2 and ATP5C1 in U-2 OS cells
The mRNA levels of EEF1A1, ATF7IP, HIF1A, SMAD7,
CLTC, MCM10, ITPR1, ADAM15, WWP2 and ATP5C1, which were the top 10
nodes with higher degrees in the PPI network, were measured. The
results of RT-qPCR demonstrated that treatment with curcumin
significantly increased the mRNA levels of CLTC and ITPR1 compared
with DMSO-treated and control cells (P<0.01; Fig. 2). Additionally, the differential
expression of ITPR1in curcumin-treated cells was more marked
compared with CLTC.
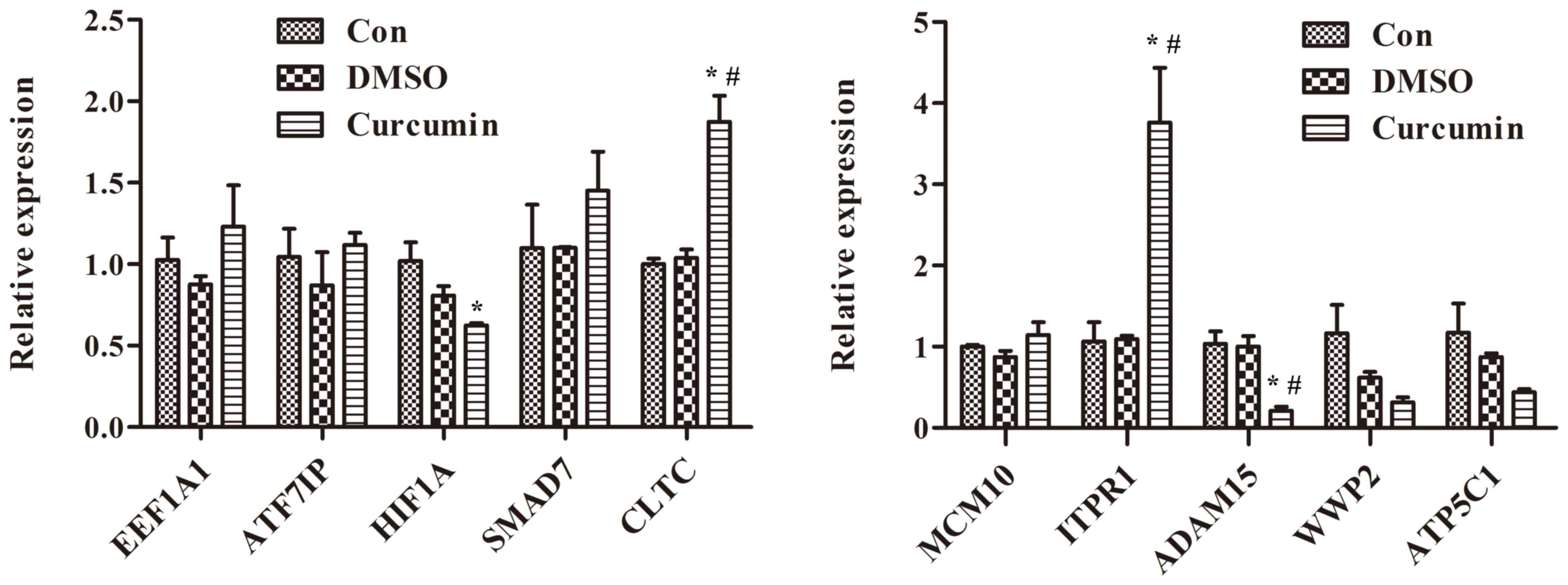 | Figure 2.Treatment with curcumin significantly
increases the mRNA levels of CLTC and ITPR1 and decreases the mRNA
levels of ADAM15 in curcumin-treated cells compared with dimethyl
sulfoxide-treated and control cells, *P<0.01 compared with
control cells, #P<0.01 compared with dimethyl
sulfoxide-treated cells. EEF1A1, eukaryotic translation elongation
factor 1α1; ATF7IP, activating transcription factor 7 interacting
protein; HIF1A, hypoxia-inducible factor-1α; SMAD7, SMAD family
member 7; CLTC, clathrin heavy chain; MCM10, minichromosome
maintenance 10 replication initiation factor; ITPR1, inositol
1,4,5-triphosphate receptor type 1; ADAM15, a disintegrin and
metalloprotease 15; WWP2, WW domain containing E3 ubiquitin protein
ligase 2; ATP5C1, ATP synthase, H+ transporting,
mitochondrial F1 complex, gamma polypeptide 1; Con, control; DMSO,
dimethyl sulfoxide. |
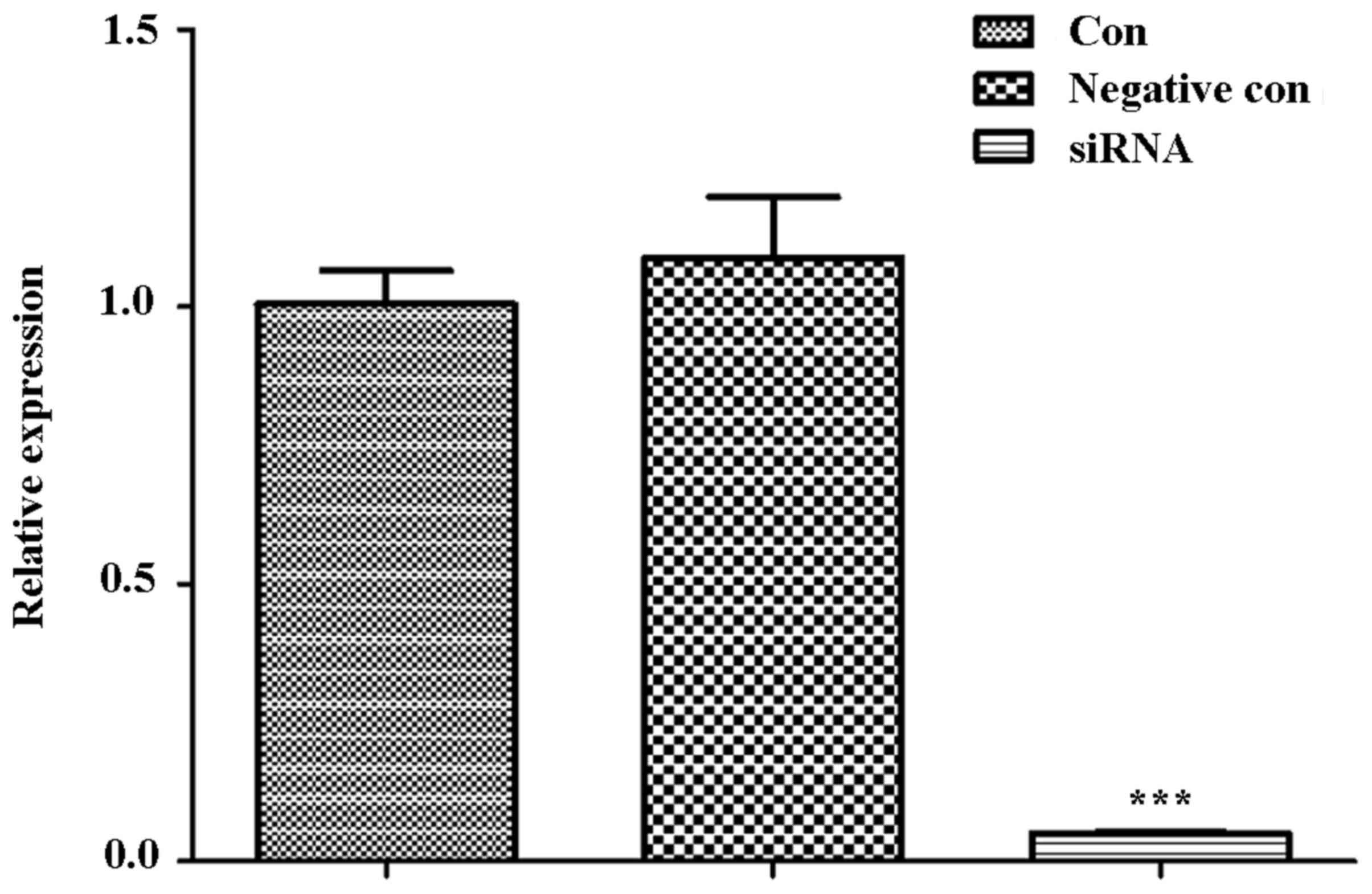
RNA interference assay of ITPR1
With ITPR1 siRNA sequences, RNA interference assay
was performed in U-2 OS cells. Then, the cells were collected for
RT-qPCR analysis. The result of RT-qPCR demonstrated that ITPR1 was
significantly decreased in cells transferred with ITPR1 siRNA
sequences compared with the negative control cells and control
cells (P<0.001; Fig. 3).
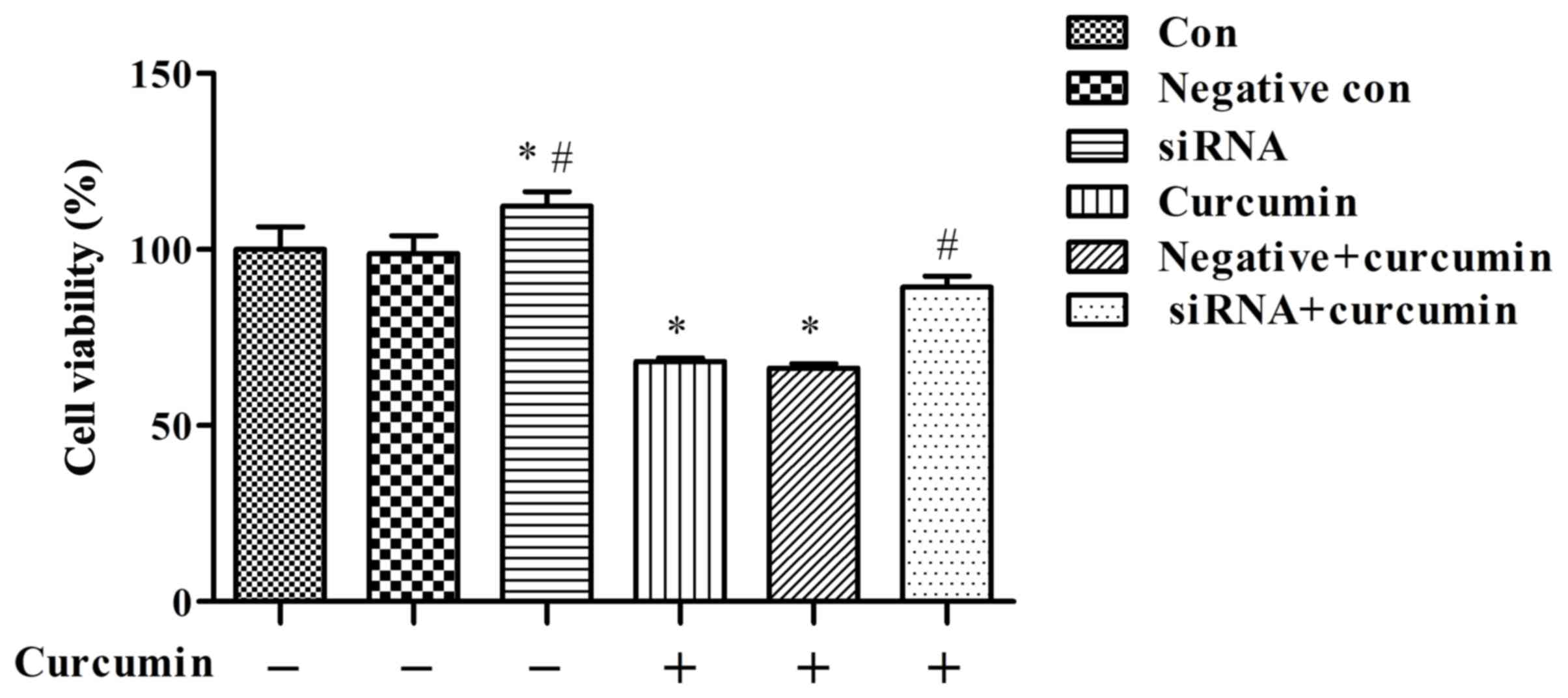
Effects of curcumin treatment and
ITPR1 expression on proliferation in U-2 OS cells
The cell proliferation assay indicated that curcumin
treatment was able to significantly suppress the proliferation of
U-2 OS cells than the control group not treated with curcumin
(P<0.05; Fig. 4), and the effect
of curcumin-mediated suppression decreased subsequent to ITPR1
knockdown (P<0.01 vs. negative + curcumin; Fig. 4).
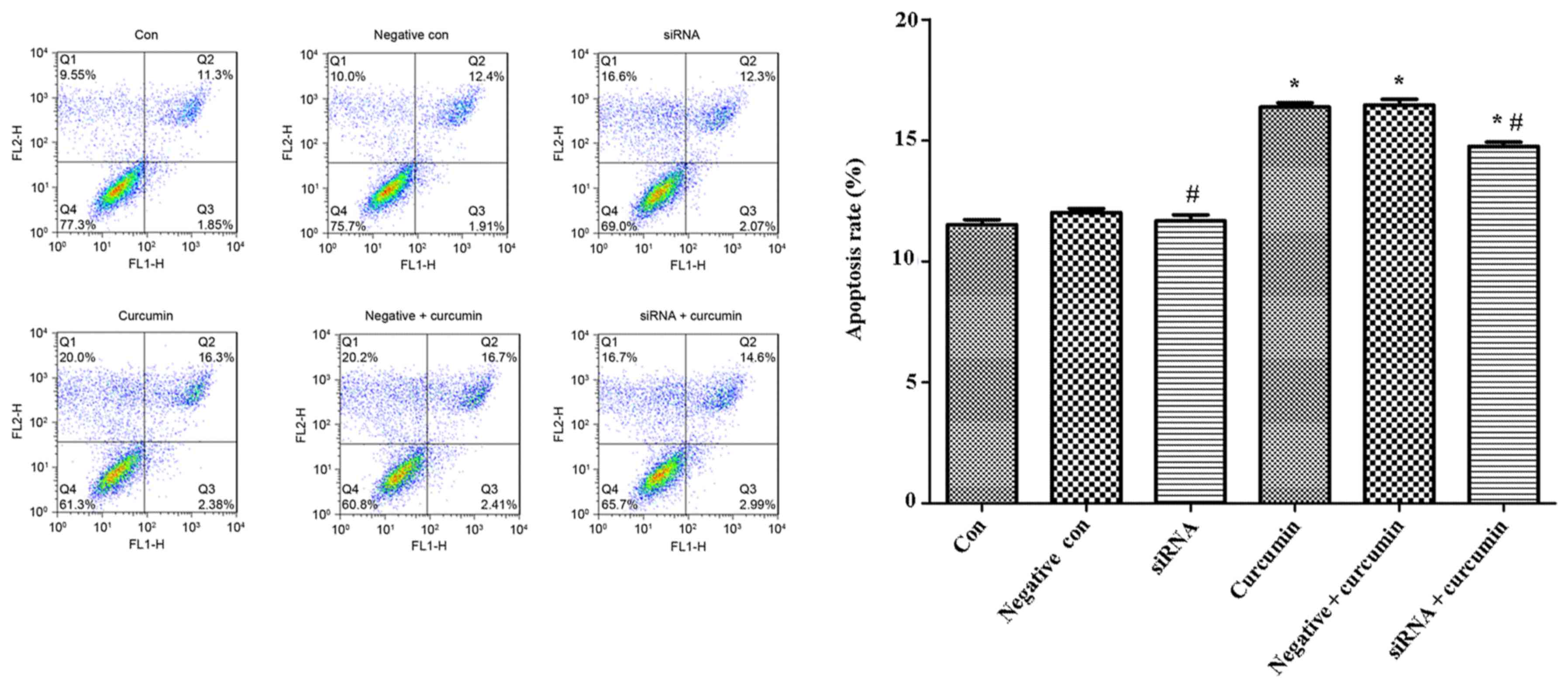
Effects of curcumin treatment and
ITPR1 expression on apoptosis of U-2 OS cells
Flow cytometry was used to analysis the apoptotic
rate of U-2 OS cells, and the results indicated that treatment with
curcumin was able to significantly promote apoptosis of U-2 OS
cells compared with the control not treated with curcumin
(P<0.05; Fig. 5), while the
apoptotic rate of U-2 OS cells transfected with ITPR1 siRNA
sequences was significantly decreased (P<0.05; Fig. 5).
Effects of curcumin treatment and
ITPR1 expression on migration of U-2 OS cells
A wound healing assay was performed to detect the
migration of U-2 OS cells. The results indicated that treatment
with curcumin significantly inhibited cell migration (P<0.01;
Fig. 6). Compared with the control
group, ITPR1 interference promoted migration of U-2 OS cells
(P<0.01; Fig. 6). Meanwhile, ITPR1
interference significantly promoted migration of U-2 OS cells
compared with curcumin-treated cells (P<0.05; Fig. 6).
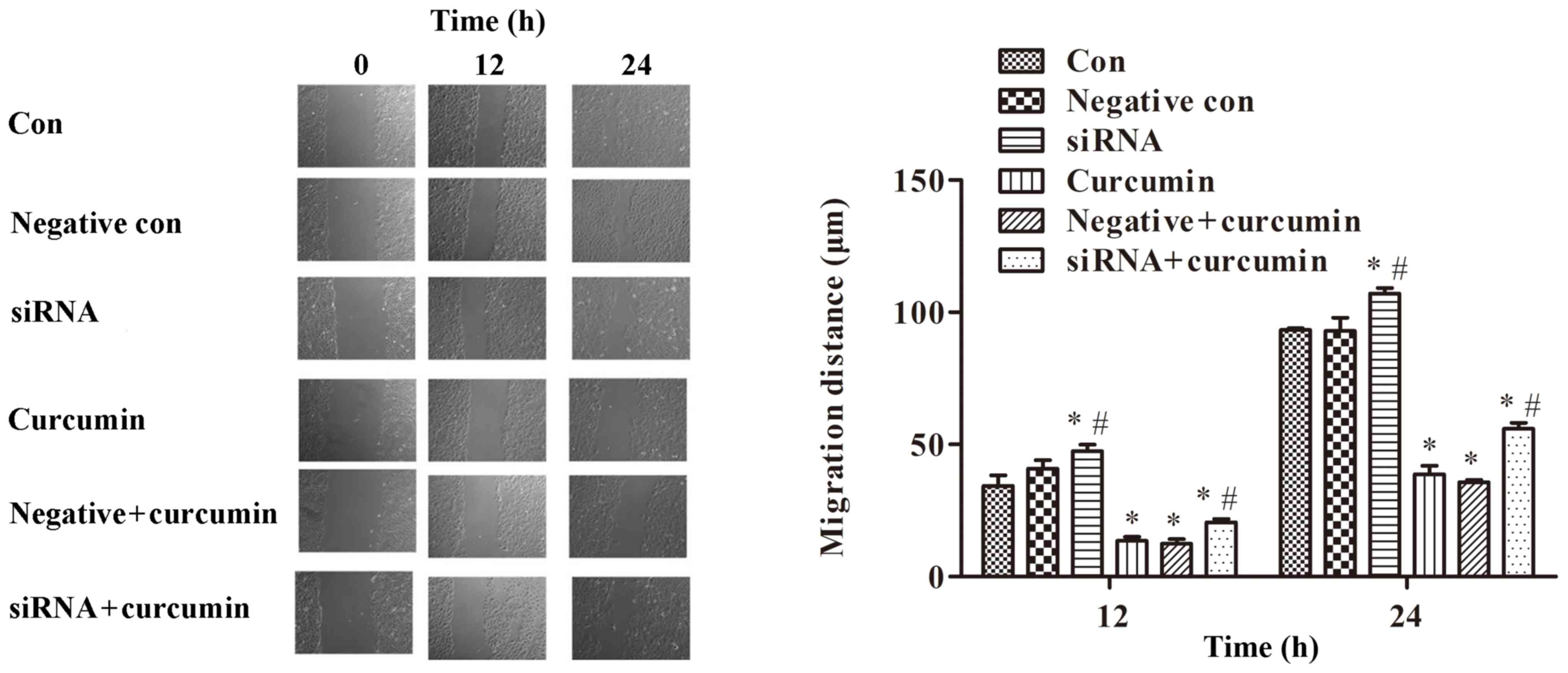 | Figure 6.Treatment with curcumin significantly
inhibits cell migration, while ITPR1 interference promotes
migration of U-2 OS cells (magnification, ×200). *P<0.01,
compared with negative control cells and control cells (Note: for
siRNA treated cells at 12 h, the *P<0.01 is compared with
control cells); #P<0.05, compared with negative and
curcumin-treated cells. si, small interfering; con, control. ITPR1,
inositol 1,4,5-triphosphate receptor type 1. |
Effects of curcumin treatment and
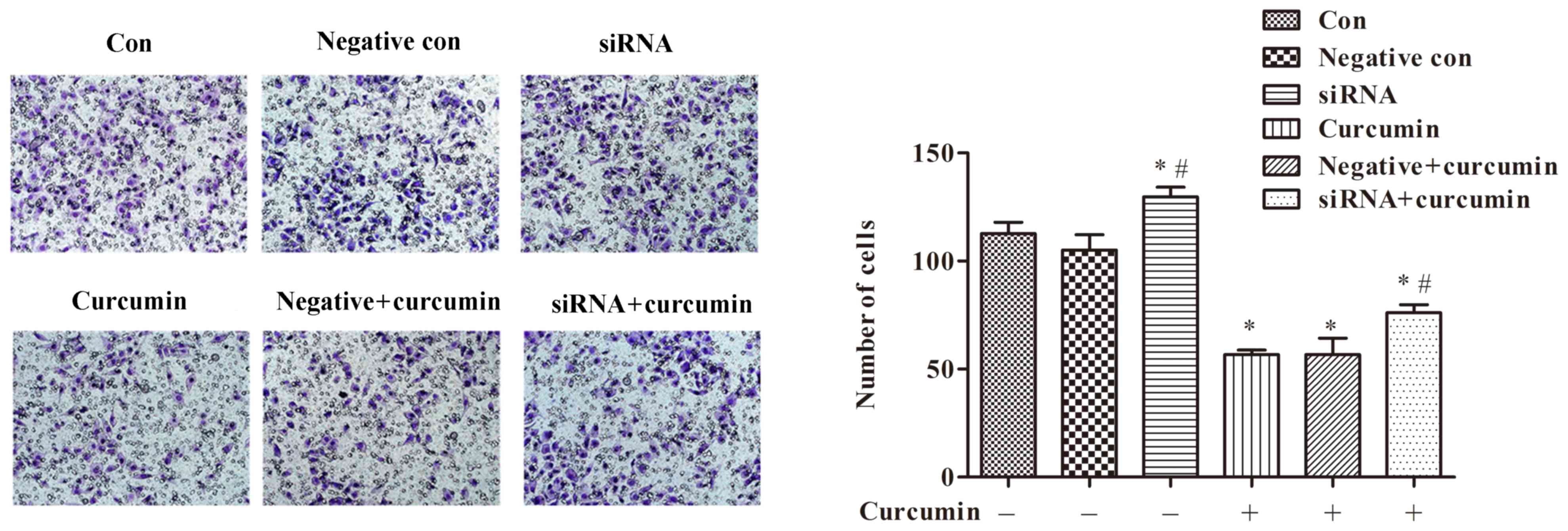
ITPR1 expression on invasion of U-2 OS cells
Additionally, a Transwell assay was utilized to
investigate the effects of curcumin treatment and ITPR1 expression
on invasion of U-2 OS cells. The results of the Transwell assay
suggested that treatment with curcumin was able to significantly
inhibit invasion of U-2 OS cells compared with the control not
treated with curcumin (P<0.05; Fig.
7). In addition, ITPR1 interference relieved the inhibitory
effect of curcumin treatment on cell invasion (P<0.05; Fig. 7).
Discussion
In the present study, RNA-sequencing was used to
perform transcriptomic analysis of human osteosarcoma U-2 OS cells
treated with curcumin or DMSO. A total of 201 DEGs were identified
in the curcumin-treated group, including 114 upregulated and 87
downregulated genes. Functional enrichment indicated that
downregulated HIF1A was involved in biological processes,
including signal transduction, cell communication and cellular
process. HIF1A serves a key role in progression, invasion
and metastasis of a number of types of human cancer, including
osteosarcoma (29). It has been
demonstrated that small hairpin RNA (shRNA)-mediated knockdown of
HIF1A was able to efficiently inhibit the hypoxia
transduction pathway and block the growth of osteosarcoma cells
(30). HIF1A functions in
osteosarcoma progression and the oxygen-dependent degradation of
HIF1A may be critical for osteosarcoma (31). HIF1A is highly relevant to the
primary occurrence or recurrence, size, clinical stage,
pathological grade and angiogenesis of the osteosarcoma of the jaw,
and may serve as a therapeutic target for the disease (32). Therefore, the present study
hypothesized that HIF1A may also be involved in
osteosarcoma.
EEF1A1 is overexpressed in
methotrexate-treated osteosarcoma Saos-2 cell line, and may promote
cell growth through the increase of protein translation (33). Concurrently, the inhibition of
EEF1A1 may protect myotubes from apoptosis (34). Blanch et al (35) revealed that EEF1A1 exhibited
effects on tumor protein (p) 53 and p73
through regulating human homologue of Mdm2 (HDM2) in a
number of types of cancer, including osteosarcoma (35). Furthermore, several studies have
demonstrated that the p53-family proteins participate in
tumorigenesis via the mediation of the genes associated with cell
cycle progression and apoptosis (36,37). In
osteosarcoma Saos2 cells, SMAD7 may suppress bone
morphogenetic protein 2 (BMP2)-induced differentiation and BMP/SMAD
signaling through interacting with nuclear factor-κB (38). Previous studies have reported that
SMAD7 overexpression may inhibit tumor progression and lung
metastasis of osteosarcoma (39,40). Taken
together, these data suggested that EEF1A1 and SMAD7
may participate in osteosarcoma. In the PPI network of the present
study, EEF1A1 (degree=88), ATF7IP (degree=64), HIF1A (degree=44),
SMAD7 (degree=43), CLTC (degree=42), MCM10 (degree=28), ITPR1
(degree=27), ADAM15 (degree=24), WWP2 (degree=22) and ATP5C1
(degree=21) exhibited the highest degrees. These genes were
involved in the GO term ‘biological process’, indicating that they
may be involved in the anticancer role of curcumin in human
osteosarcoma U-2 OS cells through biological processes.
To additionally investigate the mechanisms of action
of curcumin on human osteosarcoma U-2 OS cells, the mRNA levels of
EEF1A1, ATF7IP, HIF1A, SMAD7, CLTC, MCM10, ITPR1, ADAM15,
WWP2 and ATP5C1 in U-2 OS cells were analyzed. RT-qPCR
demonstrated that treatment with curcumin significantly increased
the mRNA levels of CLTC and ITPR1 in curcumin-treated
cells, while the mRAN levels of ADAM15 dramatically
decreased in curcumin-treated cells. Notably, the differential
expression of ITPR1 was more significant compared with
CLTC. Then, the effects of curcumin treatment and
ITPR1 expression on proliferation, apoptosis, migration and
invasion in U-2 OS cells were investigated. The results indicated
that treatment with curcumin was able to significantly promote
apoptosis and suppress proliferation, migration and invasion. It
was also indicated that ITPR1 may contribute to the effects
mediated by curcumin. A previous study had suggested that curcumin
suppressed proliferation, invasion, survival, metastasis and
angiogenesis of a number of types of cancer via interacting with
cell signaling proteins including enzymes and inflammatory
cytokines (41). Lee et al
(42) with other previous studies
demonstrated that treatment with curcumin promoted G1/S and G2/M
cell cycle arrest and that activation of the caspase-3 signaling
pathway induced apoptosis in human osteosarcoma cells (14,43). Li
et al (44) indicated that
curcumin inhibited proliferation of osteosarcoma via the
inactivation of the Notch-1 signaling pathway. The ITPR1
gene encodes the intracellular calcium release channel type 1
InsP3R that can bind sensitized cells and cytochrome c to apoptotic
stimuli (45–47). In osteoblastic cells, including G-292
osteosarcoma cells, exposure to interleukin-1β, tumor necrosis
factor α and other apoptotic stimuli increase mRNA and protein
levels of ITPR1 (48). These
finding suggest that curcumin may serve an anticancer role in human
osteosarcoma U-2 OS cells by regulating ITPR1.
In conclusion, a total of 201 DEGs were identified
in the curcumin-treated group compared with the control group.
Curcumin may serve an anticancer role by regulating apoptosis,
proliferation, migration and invasion in human osteosarcoma U-2 OS
cells. In addition, ITPR1 may be involved in the mechanism
of action of curcumin treatment of osteosarcoma.
References
|
1
|
Broadhead ML, Clark J, Myers DE, Dass CR
and Choong PF: The molecular pathogenesis of osteosarcoma: A
review. Sarcoma. 2011:9592482011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ottaviani G and Jaffe N: The epidemiology
of osteosarcoma = Pediatric and adolescent osteosarcoma. Springer;
pp. 3–13. 2010
|
|
3
|
He JP, Hao Y, Wang XL, Yang XJ, Shao JF,
Guo FJ and Feng JX: Review of the molecular pathogenesis of
osteosarcoma. Asian Pac J Cancer Prev. 15:5967–5976. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Deng BY, Hua YQ and Cai ZD: Establishing
an osteosarcoma associated protein-protein interaction network to
explore the pathogenesis of osteosarcoma. Eur J Med Res. 18:572013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen Y, Liu WH, Chen BL, Fan L, Han Y,
Wang G, Hu DL, Tan ZR, Zhou G, Cao S and Zhou HH: Plant polyphenol
curcumin significantly affects CYP1A2 and CYP2A6 activity in
healthy, male Chinese volunteers. Ann Pharmacother. 44:1038–1045.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rattigan Y and Maitra A: Metabolomic
profiling of curcumin effects on pancreatic cancer: Insights into
anti-tumor activity. Pancreatology. 13:e682013. View Article : Google Scholar
|
|
7
|
Motterlini R, Foresti R, Bassi R and Green
CJ: Curcumin, an antioxidant and anti-inflammatory agent, induces
heme oxygenase-1 and protects endothelial cells against oxidative
stress. Free Radic Biol Med. 28:1303–1312. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chattopadhyay I, Biswas K, Bandyopadhyay U
and Banerjee RK: Turmeric and curcumin: Biological actions and
medicinal applications. Curr Sci. 87:44–53. 2004.
|
|
9
|
Qiong L and Ran X: Pharmacological effects
of curcumin and bladder cancer treatment research progress.
Zhongyaoyao li yulinchuang. 3:0502012.
|
|
10
|
Walters DK, Muff R, Langsam B, Born W and
Fuchs B: Cytotoxic effects of curcumin on osteosarcoma cell lines.
Invest New Drugs. 26:289–297. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bao B, Ali S, Kong D, Sarkar SH, Wang Z,
Banerjee S, Aboukameel A, Padhye S, Philip PA and Sarkar FH:
Anti-tumor activity of a novel compound-CDF is mediated by
regulating miR-21, miR-200, and PTEN in pancreatic cancer. PLoS
One. 6:e178502011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jaiswal AS, Marlow BP, Gupta N and Narayan
S: Beta-catenin-mediated transactivation and cell-cell adhesion
pathways are important in curcumin (diferuylmethane)-induced growth
arrest and apoptosis in colon cancer cells. Oncogene. 21:8414–8427.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fossey SL, Bear MD, Lin J, Li C, Schwartz
EB, Li PK, Fuchs JR, Fenger J, Kisseberth WC and London CA: The
novel curcumin analog FLLL32 decreases STAT3 DNA binding activity
and expression and induces apoptosis in osteosarcoma cell lines.
BMC Cancer. 11:1122011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chang Z, Xing J and Yu X: Curcumin induces
osteosarcoma MG63 cells apoptosis via ROS/Cyto-C/Caspase-3 pathway.
Tumour Biol. 35:753–758. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Leow PC, Tian Q, Ong ZY, Yang Z and Ee PL:
Antitumor activity of natural compounds, curcumin and PKF118-310,
as Wnt/β-catenin antagonists against human osteosarcoma cells.
Invest New Drugs. 28:766–782. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang Z, Gerstein M and Snyder M: RNA-Seq:
A revolutionary tool for transcriptomics. Nat Rev Genet. 10:57–63.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ozsolak F and Milos PM: RNA sequencing:
Advances, challenges and opportunities. Nat Rev Genet. 12:87–98.
2011. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Trapnell C, Pachter L and Salzberg SL:
TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics.
25:1105–1111. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Langmead B: Aligning short sequencing
reads with Bowtie. Curr Protoc Bioinformatics. Dec;2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fujita PA, Rhead B, Zweig AS, Hinrichs AS,
Karolchik D, Cline MS, Goldman M, Barber GP, Clawson H, Coelho A,
et al: The UCSC genome browser database: Update 2011. Nucleic Acids
Res. 39(Database Issue): D876–D882. 2010.PubMed/NCBI
|
|
21
|
Trapnell C, Hendrickson DG, Sauvageau M,
Goff L, Rinn JL and Pachter L: Differential analysis of gene
regulation at transcript resolution with RNA-seq. Nat Biotechnol.
31:46–53. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gene Ontology Consortium: The gene
ontology (GO) project in 2006. Nucleic Acids Res. 34(Database
Issue): D322–D326. 2006.PubMed/NCBI
|
|
23
|
Kanehisa M, Araki M, Goto S, Hattori M,
Hirakawa M, Itoh M, Katayama T, Kawashima S, Okuda S, Tokimatsu T
and Yamanishi Y: KEGG for linking genomes to life and the
environment. Nucleic Acids Res. 36(Database Issue): D480–D484.
2008.PubMed/NCBI
|
|
24
|
Yu G, Wang LG, Han Y and He QY:
clusterProfiler: An R package for comparing biological themes among
gene clusters. OMICS. 16:284–287. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Benjamini Y and Hochberg Y: Controlling
the false discovery rate: A practical and powerful approach to
multiple testing. J R Stat Soc Series B Stat Methodol. 57:289–300.
1995.
|
|
26
|
Keshava Prasad TS, Goel R, Kandasamy K,
Keerthikumar S, Kumar S, Mathivanan S, Telikicherla D, Raju R,
Shafreen B, Venugopal A, et al: Human protein reference
database-2009 update. Nucleic Acids Res. 37(Database Issue):
D767–D772. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Shannon P, Markiel A, Ozier O, Baliga NS,
Wang JT, Ramage D, Amin N, Schwikowski B and Ideker T: Cytoscape: A
software environment for integrated models of biomolecular
interaction networks. Genome Res. 13:2498–2504. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chau NM, Rogers P, Aherne W, Carroll V,
Collins I, McDonald E, Workman P and Ashcroft M: Identification of
novel small molecule inhibitors of hypoxia-inducible factor-1 that
differentially block hypoxia-inducible factor-1 activity and
hypoxia-inducible factor-1alpha induction in response to hypoxic
stress and growth factors. Cancer Res. 65:4918–4928. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wu Q, Yang SH, Ye SN and Wang RY:
Therapeutic effects of RNA interference targeting HIF-1 alpha gene
on human osteosarcoma. Zhonghua Yi Xue Za Zhi. 85:409–413. 2005.(In
Chinese). PubMed/NCBI
|
|
31
|
El Naggar A, Clarkson P, Zhang F, Mathers
J, Tognon C and Sorensen PH: Expression and stability of hypoxia
inducible factor 1α in osteosarcoma. Pediatr Blood Cancer.
59:1215–1222. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen WL, Feng HJ and Li HG: Expression and
significance of hypoxemia-inducible factor-1alpha in osteosarcoma
of the jaws. Oral Surg Oral Med Oral Pathol Oral Radiol Endod.
106:254–257. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Selga E, Oleaga C, Ramírez S, de Almagro
MC, Noé V and Ciudad CJ: Networking of differentially expressed
genes in human cancer cells resistant to methotrexate. Genome Med.
1:832009. View
Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ruest LB, Marcotte R and Wang E: Peptide
elongation factor eEF1A-2/S1 expression in cultured differentiated
myotubes and its protective effect against caspase-3-mediated
apoptosis. J Biol Chem. 277:5418–5425. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Blanch A, Robinson F, Watson IR, Cheng LS
and Irwin MS: Eukaryotic translation elongation factor 1-alpha 1
inhibits p53 and p73 dependent apoptosis and chemotherapy
sensitivity. PLoS One. 8:e664362013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kastan MB, Canman CE and Leonard CJ: P53,
cell cycle control and apoptosis: Implications for cancer. Cancer
Metastasis Rev. 14:3–15. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yang A, Kaghad M, Wang Y, Gillett E,
Fleming MD, Dötsch V, Andrews NC, Caput D and McKeon F: p63, a p53
homolog at 3q27-29, encodes multiple products with transactivating,
death-inducing, and dominant-negative activities. Mol Cell.
2:305–316. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Eliseev RA, Schwarz EM, Zuscik MJ, O'Keefe
RJ, Drissi H and Rosier RN: Smad7 mediates inhibition of Saos2
osteosarcoma cell differentiation by NFkappaB. Exp Cell Res.
312:40–50. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lamora A, Talbot J, Bougras G, Amiaud J,
Leduc M, Chesneau J, Taurelle J, Stresing V, Le Deley MC, Heymann
MF, et al: Overexpression of smad7 blocks primary tumor growth and
lung metastasis development in osteosarcoma. Clin Cancer Res.
20:5097–5112. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Won KY, Kim YW and Park YK: Expression of
Smad and its signalling cascade in osteosarcoma. Pathology.
42:242–247. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kunnumakkara AB, Anand P and Aggarwal BB:
Curcumin inhibits proliferation, invasion, angiogenesis and
metastasis of different cancers through interaction with multiple
cell signaling proteins. Cancer Lett. 269:199–225. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lee DS, Lee MK and Kim JH: Curcumin
induces cell cycle arrest and apoptosis in human osteosarcoma (HOS)
cells. Anticancer Res. 29:5039–5044. 2009.PubMed/NCBI
|
|
43
|
Jin S, Xu HG, Shen JN, Chen XW, Wang H and
Zhou JG: Apoptotic effects of curcumin on human osteosarcoma U2OS
cells. Orhop Surg. 1:144–152. 2009. View Article : Google Scholar
|
|
44
|
Li Y, Zhang J, Ma D, Zhang L, Si M, Yin H
and Li J: Curcumin inhibits proliferation and invasion of
osteosarcoma cells through inactivation of Notch-1 signaling. FEBS
J. 279:2247–2259. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Li C, Wang X, Vais H, Thompson CB, Foskett
JK and White C: Apoptosis regulation by Bcl-x(L) modulation of
mammalian inositol 1,4,5-trisphosphate receptor channel isoform
gating. Proc Natl Acad Sci USA. 104:pp. 12565–12570. 2007;
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Orrenius S, Zhivotovsky B and Nicotera P:
Regulation of cell death: The calcium-apoptosis link. Nat Rev Mol
Cell Biol. 4:552–565. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Boehning D, Patterson RL and Snyder SH:
Apoptosis and calcium: New roles for cytochrome c and inositol
1,4,5-trisphosphate. Cell Cycle. 3:250–252. 2004. View Article : Google Scholar
|
|
48
|
Bradford PG, Maglich JM and Kirkwood KL:
IL-1 beta increases type 1 inositol trisphosphate receptor
expression and IL-6 secretory capacity in osteoblastic cell
cultures. Mol Cell Biol Res Commun. 3:73–75. 2000. View Article : Google Scholar : PubMed/NCBI
|