Introduction
Breast cancer is the leading cause of cancer-related
deaths in women worldwide (1).
Breast cancer risk is increased by prolonged estrogen exposure,
such as early menarche, late menopause, having no childbirth
experience, or increased estrogen levels in adipocytes due to
obesity (2,3). In addition, mutated BRCA1 and
BRCA2 genes are associated with breast cancer risk (4). Management strategies for breast cancer
include surgery, radiotherapy, chemotherapy, endocrine treatment,
and targeted treatment (5). Among
these treatments, anticancer drugs have been widely used for breast
cancer treatment. For example, tamoxifen acts on the estrogen
pathway and is applied in estrogen receptor-positive breast cancer
patients (6). Patients who have high
levels of metastatic human epidermal growth factor receptor 2
(HER2)/neu are prescribed trastuzumab, which blocks the HER2 signal
and improves patient outcomes (7).
However, these chemotherapy drugs have side effects including early
menopause, depression, rash, and renal toxicity.
Epithelial-mesenchymal transition (EMT) is a process
associated with the disruption of cell junctions and the loss of
cell polarity, which increases cell mobility and allows them to
acquire stem cell-like properties (8). As its name suggests, EMT causes the
loss of epithelial markers (E-cadherin and occludin) and
acquisition of mesenchymal markers (fibronectin, vimentin and
N-cadherin) (9). EMT is induced by
numerous growth factors and related signaling pathways, including
transforming growth factor-β1 (TGF-β1) (10). TGF-β1 plays a prominent role in
breast cancer progression and bone metastasis (11).
Hispidulin (4′,5,7-trihydroxyl-6-methoxyflavone) is
a phenolic flavonoid compound widely used in traditional medicine
(12). Several studies have shown
that hispidulin has anti-obesity, antioxidant, anti-mutagenic,
anti-inflammatory, and anti-tumor effects (13–17).
Hispidulin also has anticonvulsant, neuroprotective, and
anti-osteoporosis effects (18–21).
However, the role of hispidulin in EMT has not yet been studied in
breast cancer. In this study, we used two breast cancer cell lines,
luminal type cells (MCF-7) with strong cell-to-cell adhesion and an
aggregated structure, and claudin-low type cells (HCC38) with an
invasive form and high metastatic properties due to weak cohesion
between cells. We aimed to investigate the effect of hispidulin on
EMT and cell migration induced by TGF-β1 treatment in breast cancer
cells.
Materials and methods
Reagents and antibodies
Hispidulin was purchased from Sigma-Aldrich; Merck
KGaA and was dissolved in dimethyl sulfoxide (DMSO) and mixed with
fresh medium to achieve the desired final concentrations. TGF-β1
was purchased from R&D Systems. The following primary
antibodies were used in the experiment: anti-β-actin and
anti-total-Smad2/3 (Santa Cruz Biotechnology, Inc.),
anti-phospho-Smad2/3 (Cell Signaling Technology, Inc.),
anti-vimentin (Sigma-Aldrich; Merck KGaA), anti-E-cadherin, and
anti-occludin (BD Biosciences). The secondary antibodies used were
horseradish peroxidase (HRP-conjugated anti-mouse; Santa Cruz
Biotechnology, Inc.) and HRP-conjugated anti-rabbit (Cell Signaling
Technology, Inc.).
Cell culture
The two breast cancer cell lines used in this study
(MCF-7 and HCC38) were obtained from the Korean Cell Line Bank
(Seoul, Korea) and cultured in RPMI-1640 medium (Gibco; Thermo
Fisher Scientific, Inc.) containing 1% penicillin, streptomycin,
and 10% fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific,
Inc.) at 37°C in a 5% CO2 atmosphere.
Cell viability assay
Breast cancer cell viability was assessed using the
3-(4,5-Dimethylthiazol-2-yl)-
5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium (MTS)
assay, as previously described (22). Briefly, cells were plated
(2×103 cells/well) in 96-well plates and allowed to grow
overnight. Cells were treated with various concentrations of
hispidulin and/or TGF-β1 for 24 h. Next, MTS reagent was added to
each well according to the manufacturer's protocol.
RNA isolation and reverse
transcription-quantitative polymerase chain reaction
Total RNA was extracted from hispidulin-treated
breast cancer cells using the RNeasy mini kit (Qiagen) and cDNA was
obtained using AccuPower® RT PreMix (Bioneer).
Quantitative PCR was performed as described previously (22). All experiments were performed in
triplicate. The relative change of each gene was normalized to the
expression of glyceraldehyde-3-phosphate dehydrogenase (GAPDH). The
fold-changes in mRNA expression were calculated using the
2−∆∆Cq method (23). The
primers were as follows: E-cadherin, forward
5′-AAAGGCCCATTTCCTAAAAACCT-3′ and reverse
5′-TGCGTTCTCTATCCAGAGGCT-3′; occludin, forward
5′-CTTCAGGCAGCCTCGTTACA-3′ and reverse 5′-TACCTGATCCAGTCCTCCTC-3;
vimentin, forward 5′-CTCTTCCAAACTTTTCCTCCC-3′; and reverse
5′-AGTTTCGTTGATAACCTGTCC-3′; GAPDH, forward
5′-GGACCTGACCTGCCGTCTAGAA-3′ and reverse
5′-GGTGTCGCTGTTGAAGTCAGAG-3′.
Western blot
After treatment with hispidulin and/or TGF-β1 for 24
h, cell lysates were collected and prepared. Protein concentration
was quantified using the Bradford assay (Bio-Rad Laboratories,
Inc.). After SDS-PAGE, the proteins were transferred onto
polyvinylidene difluoride (PVDF) membranes (EMD Millipore). After
blocking with 5% non-fat dry milk, the membranes were probed with
different primary antibodies. Membranes were incubated with an
HRP-conjugated secondary antibody for 1 h, and protein detection
was performed using enhanced chemiluminescence (ECL) solution
(PerkinElmer).
Confocal microscopy
Fluorescent images were examined as previously
described (22). Briefly, the
treated breast cancer cells were fixed in 4% paraformaldehyde for 1
h. Then, the cells were permeabilized with 0.5% Triton X-100,
blocked in 1% bovine serum albumin (BSA), and stained with the
primary antibody overnight at 4°C. Cells were treated with
secondary antibody for 1 h and then mounted using
4′,6-diamidino-2-phenylindole (DAPI). Cells were observed using
confocal microscopy (LSM-700; Carl Zeiss).
Wound healing assay
Each breast cancer cell line (4×105
cells) was seeded in a 12-well plate and allowed to grow until
approximately 90% confluence. Then, the media was removed and the
monolayer was scratched with a pipette tip. 10 ng/ml TGF-β1 and
hispidulin (1.25 and 2.5 µM)-treated cells were maintained in 1%
FBS-containing media for 24 h. The media was then removed and the
cells were washed with phosphate-buffered saline (PBS). After
fixing with methanol, the cells were stained with Giemsa stain
solution (Sigma-Aldrich; Merck KGaA) for 1 h. Cells were then
observed and photographed using an optical microscope (CKX41;
Olympus Corporation) and analyzed using Image J software (NIH).
Transwell migration assay
Breast cancer cells were plated in the upper chamber
of transwell plates containing TGF-β1 (10 ng/ml) and hispidulin
(1.25 and 2.5 µM) (Costar). The lower chamber was supplemented with
serum-free media containing 5% FBS. After 24 h of incubation, the
cells were fixed with methanol. Cells were stained with hematoxylin
(Sigma-Aldrich; Merck KGaA) and eosin (Sigma-Aldrich; Merck KGaA).
Cells were then photographed using an optical microscope (CKX41;
Olympus Corporation).
Statistical analysis
Graph Pad Prism 6 Software (GraphPad Software, Inc.)
was used for statistical analysis. All experiments were performed
three times, independently. One-way ANOVA with Bonferroni post-hoc
test was used for statistical analysis of the data. P<0.05 was
considered to indicate a statistically significant difference.
Results
Hispidulin inhibits breast cancer cell
viability
The chemical structure of hispidulin is shown in
Fig. 1A. We first determined the
inhibitory effect of hispidulin on the viability of breast cancer
cells. MCF-7 and HCC38 cancer cells were treated with various
concentrations of hispidulin for 24 h. We observed that the 50%
inhibitory concentration (IC50) values of hispidulin in
MCF-7 and HCC38 cells were 25.44±0.23 and 65.42±0.31 µM,
respectively. The inhibition rate of hispidulin in MCF-7 and HCC38
breast cell lines increased with increasing concentration. Next, we
determined the TGF-β1-induced inhibitory effect of hispidulin on
cell viability. As shown in Fig. 1B,
MCF-7 and HCC38 cells showed a significant increase in growth after
treatment with only TGF-β1 compared to treatment with DMSO control
for 24 h. However, treatment with both TGF-β1 and hispidulin caused
a significant growth reduction compared to the TGF-β1-treated group
in both cell lines.
Hispidulin treatment increases the
expression of epithelial markers in breast cancer cells
We studied the effect of hispidulin on EMT markers
in MCF-7 and HCC38 cells. Interestingly, 2.5 µM of hispidulin
significantly increased the mRNA expression of epithelial markers
such as E-cadherin and occludin in MCF-7 and HCC38 cells (Fig. 2A). Moreover, hispidulin decreased the
expression of vimentin mRNA in HCC38 cells. As shown in Fig. 2B, hispidulin increased the protein
expression of E-cadherin and occludin in MCF-7 cells, and similar
results were obtained in HCC38 cells. Vimentin protein expression
was significantly decreased after hispidulin treatment in HCC38
cells.
Hispidulin suppresses EMT through
TGF-β1 in breast cancer cells
TGF-β1 plays a key role in maintaining the balance
between cell differentiation and regeneration in normal epithelial
cells (24). However, TGF-β1
promotes EMT, invasion, and immunosuppression in advanced cancers
(25). We investigated the effect of
hispidulin treatment on TGF-β1-induced EMT in breast cancer cells.
As shown in Fig. 3, both breast
cancer cells treated with both TGF-β1 and hispidulin had increased
protein expression of E-cadherin compared to the cells treated with
TGF-β1 alone. After TGF-β1 and hispidulin co-treatment, the
expression of occludin increased compared to TGF-β1 treatment alone
in HCC38 cells. Occludin protein expression was assessed in MCF-7
cells, but no significant changes were found among the treatment
groups (Fig. 3). TGF-β1 and
hispidulin co-treatment decreased vimentin protein expression in
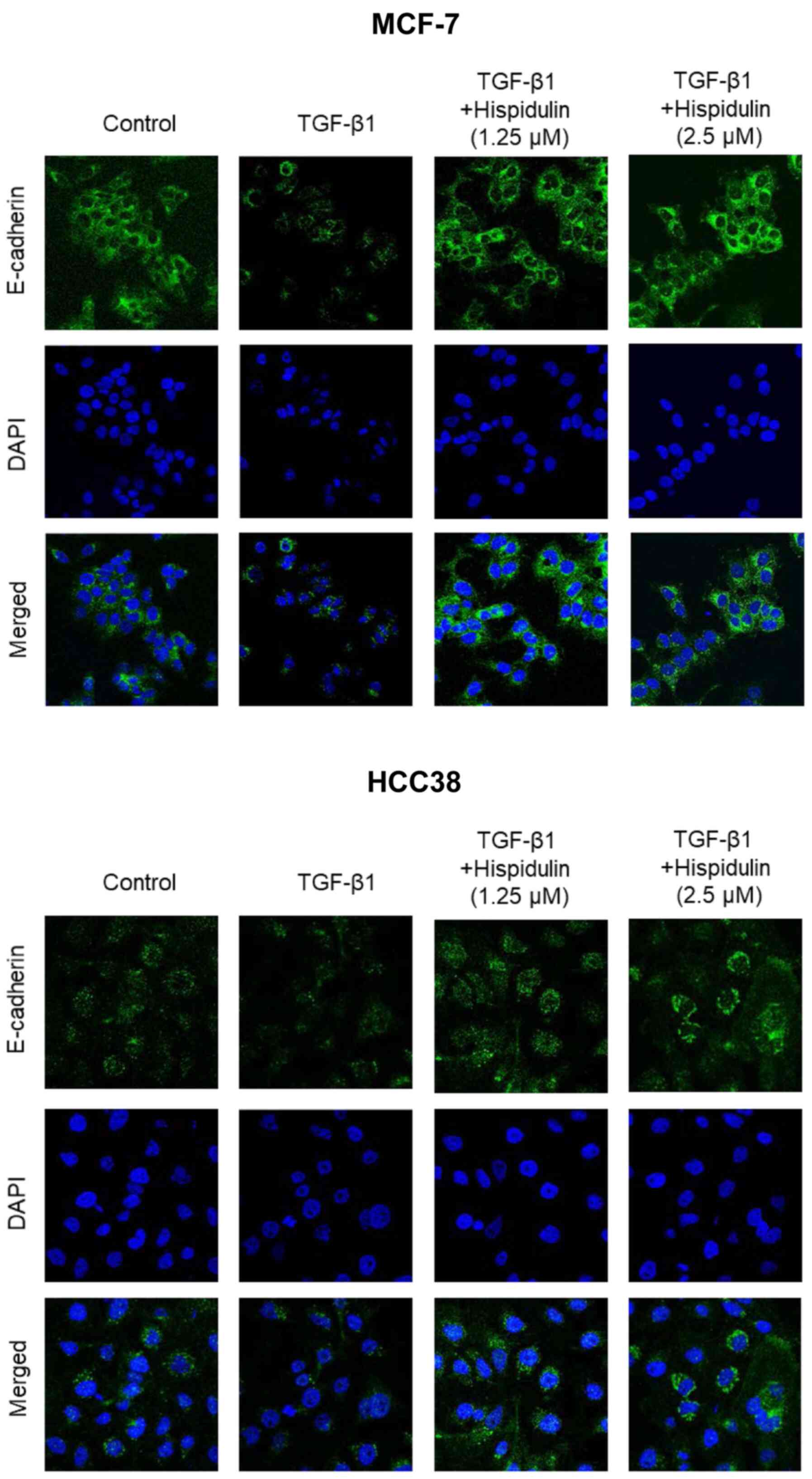
HCC38 cells. To confirm these results, immunofluorescence was
performed. As shown in Fig. 4,
TGF-β1 and hispidulin co-treatment increased the expression of
E-cadherin compared to TGF-β1 treatment alone in both MCF-7 and
HCC38 cells. These data suggest that hispidulin inhibits
TGF-β1-induced EMT.
Hispidulin downregulates
TGF-β1-induced Smad signaling
TGF-β1 induces EMT via Smad signaling pathways
(26,27). TGF-β signaling is initiated upon the
interaction of TGF-β1 with transmembrane kinase receptors, causing
the activation of downstream signaling pathways involving Smad2
and/or 3 (Smad2/3) and Smad4. Our data showed that phospho-Smad2/3
protein expression was significantly increased after TGF-β1
treatment compared to the DMSO (control) treatment, but the protein
expression of total Smad2/3 did not change in either cancer cell
line (Fig. 5). However, TGF-β1 and
hispidulin co-treatment significantly inhibited the phosphorylation
of Smad2/3 protein in breast cancer cells.
Hispidulin inhibits TGF-β1-induced
cell migration in breast cancer cells
To assess whether hispidulin influences cell
migration, we performed migration assays in breast cancer cells.
The wound area in cells treated with TGF-β1 was significantly
reduced compared to that in cells treated with DMSO (control)
(Fig. 6A). After TGF-β1 and
hispidulin co-treatment, the wound area was significantly increased
compared to that in the TGF-β1 treatment group. In addition, the
percentage of cell migration in cells treated with TGF-β1 and
hispidulin was significantly suppressed compared to the TGF-β1
treatment according to the Transwell assay in both breast cancer
cell lines (Fig. 6B).
Discussion
Recent studies have shown that bioactive compounds
in natural products have anticancer activities, and these
substances can be effective in preventing and treating cancer
(28). The natural compound
hispidulin is a phenolic flavonoid found in Lamiaceae plants
and has shown various effects against cancers (29–33).
However, studies on the effect of hispidulin on EMT in breast
cancer are limited. In this study, we studied the effect of
hispidulin on two breast cancer cell lines: MCF-7 of the luminal A
type [estrogen receptor (ER)+, progesterone receptor
(PR)+/−, human epidermal growth factor receptor 2
(HER2)−], and HCC38 of the claudin-low type
(ER−, PR−, HER2−) (34). Our results, for the first time,
demonstrated that hispidulin inhibits EMT and cell migration
through E-cadherin-mediated reorganization after TGF-β1 treatment
in breast cancer cells.
EMT is an essential process in the early stages of
embryonic development, but uncontrolled EMT is associated with
tumorigenesis (35). During EMT,
cancer cells lose the properties of epithelial cells and acquire
the properties of mesenchymal cells, allowing them to migrate to
other organs through blood vessels or lymph nodes, causing
metastasis and recurrence (36).
Most cancer-related deaths are due to metastasis, which includes
cell discharge from the primary tumor to the circulatory system,
survival in the circulatory system, adaptation in new organs,
initiation and maintenance of growth, and angiogenesis of
metastasized tumors (7,9,37). Low
expression of E-cadherin plays an important role in cancer
metastasis and is associated with a reduced survival rate in breast
cancer patients (38,39). Occludin is an integral membrane
protein located in tight junctions that physically creates a
barrier between cells (40). Loss of
occludin affects bone metastasis in cancer patients (41). It has been shown that vimentin, a
type III intermediate fibrous protein, is extensively expressed in
triple-negative breast cancer subtypes and is correlated with tumor
invasiveness and resistance to chemotherapy (42,43). In
our study, hispidulin treatment increased the expression of
E-cadherin and occludin and downregulated vimentin, with or without
TGF-β1 treatment (Fig. 5). These
results showed that hispidulin modulated EMT in breast cancers.
TGF-β1 acts as a tumor suppressor in normal
epithelial cells, but promotes tumorigenesis, metastasis, cancer
stem cell formation, and immune suppression in cancer cells
(44). High levels of TGF-β1 are
found in triple-negative breast carcinomas and are associated with
metastasis and tumor progression (24,45,46).
TGF-β1 also stimulates metastatic progression, leading to the
development of chemoresistance in breast cancer stem cells
(47). Moreover, TGF-β1 can induce
EMT and increase cell motility (48). Our study revealed that TGF-β1
decreased the expression of E-cadherin and occludin proteins and
increased the expression of vimentin in breast cancer cells
(Fig. 5). However, hispidulin
reversed the TGF-β1-induced EMT phenomenon. Therefore, hispidulin
may be effective in inhibiting TGF-β1-induced EMT in breast cancer
cells.
TGF-β1 induces EMT via the activation of
Smad-dependent and Smad-independent signaling (46). TGF-β1 binds to TGF-β1 type II
receptors (TβRII) and TGF-β1 type I receptors (TβRI) at serine and
threonine residues. Smad2/3 is then phosphorylated and activates
the expression of EMT-related transcription factors (49–52). Our
data demonstrates that hispidulin inhibits the expression of
phosphorylated-Smad2/3 induced by TGF-β1 in MCF-7 and HCC38 cell
lines (Fig. 5). Some clinical
studies have shown that TGF-β1-targeting anticancer compounds have
therapeutic effects in breast cancer patients (53). Thus, we suggest that hispidulin may
be a chemo-therapeutic via targeting TGF-β1 signaling in breast
cancers. However, further experiments are needed to investigate
non-Smad signaling in breast cancers.
Loosely connected mesenchymal cells developed
through EMT can migrate and invade other tissues (54). The present study demonstrated that
the motility of cells increases after TGF-β1 treatment; however,
TGF-β1 and hispidulin co-treatment decreases cell migration in
MCF-7 and HCC38 cells, as assessed through wound healing and
Transwell migration assays (Fig. 5).
EMT stimulates tumor cells to acquire stem cell-like properties and
increases their resistance to standard chemotherapeutic drugs as
well as conventional chemotherapy and radiotherapy (55,56). Our
study demonstrated that the inhibitory effects of hispidulin on EMT
can be modulated against both MCF-7 (moderately invasive) and HCC38
(highly invasive) breast cancers. Most studies using hispidulin
have focused on anti-cancer effects, including apoptosis signaling
including mitochondrial ROS, cell cycle mediated apoptosis using
various cancer cells (57). However,
the role of hispidulin in TGF-β1-induced EMT in human breast cells
has not been elucidated. The concentration of hispidulin used in
other cancer studies was over 10 µM (58). Our study showed that hispidulin was
effective at concentrations below 5 µM for EMT inhibition.
Therefore, this is the first study to show that the suppression of
EMT using hispidulin is a strategy for preventing and treating
breast cancers. Furthermore, several studies have shown that
hispidulin, in various drug combinations, has synergistic effects
against cancers (59). Hispidulin
enhances the anticancer effect of chemotherapeutic drugs including
gemcitabine, 5-fluorouracil, mitoxantrone, sunitinib, temozolomide,
and tumor necrosis factor-related apoptosis-inducing ligand (TRAIL)
(17,30,59–62).
However, there is a need for more research on the
synergistic effects of hispidulin and breast cancer drugs. We found
that hispidulin is rapidly absorbed in the stomach and intestines,
with an absolute bioavailability of 4.02% after oral administration
(63). Therefore, hispidulin
requires additional strategies to enhance its efficacy for
practical clinical use in chemoprevention and chemotherapy.
In conclusion, we show that hispidulin can block EMT
and that this effect may be associated with a decrease in
TGF-β1-induced signaling. Additionally, hispidulin inhibited breast
cancer cell migration after TGF-β1 treatment. Thus, hispidulin may
represent a novel anticancer agent for the treatment of early and
late stage breast cancers.
Acknowledgements
Not applicable.
Funding
The present study was supported by a research fund
from Chosun University (2020).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JL conceived and designed the study. HAK performed
the experiments. JL and HAK analyzed the data and wrote the
manuscript. JL and HAK confirm the authenticity of all the raw
data. All authors read and approved the final manuscript and agreed
to be accountable for all aspects of the research.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Torre LA, Islami F, Siegel RL, Ward EM and
Jemal A: Global Cancer in Women: Burden and Trends. Cancer
Epidemiol Biomarkers Prev. 26:444–457. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yoo KY, Kang D, Park SK, Kim SU, Kim SU,
Shin A, Yoon H, Ahn SH, Noh DY and Choe KJ: Epidemiology of breast
cancer in Korea: Occurrence, high-risk groups, and prevention. J
Korean Med Sci. 17:1–6. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Park B, Park S, Shin HR, Shin A, Yeo Y,
Choi JY, Jung KW, Kim BG, Kim YM, Noh DY, et al: Erratum to:
Population attributable risks of modifiable reproductive factors
for breast and ovarian cancers in Korea. BMC Cancer.
16:1812016.Erratum for: BMC Cancer 16: 5, 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Irving M, Elmslie F and Berg J: 18.
Genetics of breast cancer. Int J Clin Pract. 56:677–682.
2002.PubMed/NCBI
|
|
5
|
Radice D and Redaelli A: Breast cancer
management: Quality-of-life and cost considerations.
Pharmacoeconomics. 21:383–396. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Osborne CK: Tamoxifen in the treatment of
breast cancer. N Engl J Med. 339:1609–1618. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zeeshan R and Mutahir Z: Cancer metastasis
- tricks of the trade. Bosn J Basic Med Sci. 17:172–182.
2017.PubMed/NCBI
|
|
8
|
Liao TT and Yang MH: Revisiting
epithelial-mesenchymal transition in cancer metastasis: The
connection between epithelial plasticity and stemness. Mol Oncol.
11:792–804. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yeung KT and Yang J:
Epithelial-mesenchymal transition in tumor metastasis. Mol Oncol.
11:28–39. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gloushankova NA, Zhitnyak IY and Rubtsova
SN: Role of epithelial-mesenchymal transition in tumor progression.
Biochemistry (Mosc). 83:1469–1476. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yu H, Shen Y, Hong J, Xia Q, Zhou F and
Liu X: The contribution of TGF-β in Epithelial-Mesenchymal
Transition (EMT): Down-regulation of E-cadherin via snail.
Neoplasma. 62:1–15. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Patel K and Patel DK: Medicinal
importance, pharmacological activities, and analytical aspects of
hispidulin: A concise report. J Tradit Complement Med. 7:360–366.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lee SG, Kim JS, Min K, Kwon TK and Nam JO:
Hispidulin inhibits adipogenesis in 3T3-L1 adipocytes through PPARγ
pathway. Chem Biol Interact. 293:89–93. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dabaghi-Barbosa P, Mariante Rocha A,
Franco da Cruz Lima A, Heleno de Oliveira B, Benigna Martinelli de
Oliveira M, Gunilla Skare Carnieri E, Cadena SM and Eliane Merlin
Rocha M: Hispidulin: Antioxidant properties and effect on
mitochondrial energy metabolism. Free Radic Res. 39:1305–1315.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chulasiri M, Bunyapraphatsara N and
Moongkarndi P: Mutagenicity and antimutagenicity of hispidulin and
hortensin, the flavonoids from Millingtonia hortensis L.
Environ Mol Mutagen. 20:307–312. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Clavin M, Gorzalczany S, Macho A, Muñoz E,
Ferraro G, Acevedo C and Martino V: Anti-inflammatory activity of
flavonoids from Eupatorium arnottianum. J Ethnopharmacol.
112:585–589. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gao H, Xie J, Peng J, Han Y, Jiang Q, Han
M and Wang C: Hispidulin inhibits proliferation and enhances
chemosensitivity of gallbladder cancer cells by targeting HIF-1α.
Exp Cell Res. 332:236–246. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Walesiuk A, Nazaruk J and Braszko JJ:
Pro-cognitive effects of Cirsium rivulare extracts in rats. J
Ethnopharmacol. 129:261–266. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Niu X, Chen J, Wang P, Zhou H, Li S and
Zhang M: The effects of hispidulin on bupivacaine-induced
neurotoxicity: Role of AMPK signaling pathway. Cell Biochem
Biophys. 70:241–249. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhou R, Wang Z and Ma C: Hispidulin exerts
anti-osteoporotic activity in ovariectomized mice via activating
AMPK signaling pathway. Cell Biochem Biophys. 69:311–317. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yang L, Yu Z, Qu H and Li M: Comparative
effects of hispidulin, genistein, and icariin with estrogen on bone
tissue in ovariectomized rats. Cell Biochem Biophys. 70:485–490.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lee J: 3,3′-Diindolylmethane inhibits
TNF-α- and TGF-β-induced epithelial-mesenchymal transition in
breast cancer cells. Nutr Cancer. 71:992–1006. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2-ΔΔCT method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Massagué J and Chen YG: Controlling
TGF-beta signaling. Genes Dev. 14:627–644. 2000.PubMed/NCBI
|
|
25
|
Gorsch SM, Memoli VA, Stukel TA, Gold LI
and Arrick BA: Immunohistochemical staining for transforming growth
factor beta 1 associates with disease progression in human breast
cancer. Cancer Res. 52:6949–6952. 1992.PubMed/NCBI
|
|
26
|
Mani SA, Guo W, Liao MJ, Eaton EN, Ayyanan
A, Zhou AY, Brooks M, Reinhard F, Zhang CC, Shipitsin M, et al: The
epithelial-mesenchymal transition generates cells with properties
of stem cells. Cell. 133:704–715. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fuxe J, Vincent T and Garcia de Herreros
A: Transcriptional crosstalk between TGF-β and stem cell pathways
in tumor cell invasion: Role of EMT promoting Smad complexes. Cell
Cycle. 9:2363–2374. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lee KW, Bode AM and Dong Z: Molecular
targets of phytochemicals for cancer prevention. Nat Rev Cancer.
11:211–218. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lv L, Zhang W, Li T, Jiang L, Lu X and Lin
J: Hispidulin exhibits potent anticancer activity in vitro
and in vivo through activating ER stress in non-small-cell
lung cancer cells. Oncol Rep. 43:1995–2003. 2020.PubMed/NCBI
|
|
30
|
Woo SM, Seo SU, Kim SH, Nam JO, Kim S,
Park JW, Min KJ and Kwon TK: Hispidulin enhances TRAIL-mediated
apoptosis via CaMKKβ/AMPK/USP51 axis-mediated bim stabilization.
Cancers (Basel). 11:19602019. View Article : Google Scholar
|
|
31
|
Jang HJ, Lee SJ, Kim CY, Hwang JT, Choi
JH, Park JH, Lee SW and Rho MC: Effect of sunlight radiation on the
growth and chemical constituents of Salvia plebeia R.Br.
Molecules. 22:12792017. View Article : Google Scholar
|
|
32
|
Gao H, Wang H and Peng J: Hispidulin
induces apoptosis through mitochondrial dysfunction and inhibition
of P13k/Akt signalling pathway in HepG2 cancer cells. Cell Biochem
Biophys. 69:27–34. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Han M, Gao H, Xie J, Yuan YP, Yuan Q, Gao
MQ, Liu KL, Chen XH, Han YT and Han ZW: Hispidulin induces ER
stress-mediated apoptosis in human hepatocellular carcinoma cells
in vitro and in vivo by activating AMPK signaling pathway. Acta
Pharmacol Sin. 40:666–676. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Holliday DL and Speirs V: Choosing the
right cell line for breast cancer research. Breast Cancer Res.
13:2152011. View
Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lou Y, Preobrazhenska O, auf dem Keller U,
Sutcliffe M, Barclay L, McDonald PC, Roskelley C, Overall CM and
Dedhar S: Epithelial-mesenchymal transition (EMT) is not sufficient
for spontaneous murine breast cancer metastasis. Dev Dyn.
237:2755–2768. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wu Y and Zhou BP: New insights of
epithelial-mesenchymal transition in cancer metastasis. Acta
Biochim Biophys Sin (Shanghai). 40:643–650. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Chambers AF, Groom AC and MacDonald IC:
Dissemination and growth of cancer cells in metastatic sites. Nat
Rev Cancer. 2:563–572. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
38
|
Heerboth S, Housman G, Leary M, Longacre
M, Byler S, Lapinska K, Willbanks A and Sarkar S: EMT and tumor
metastasis. Clin Transl Med. 4:62015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ricciardi GR, Adamo B, Ieni A, Licata L,
Cardia R, Ferraro G, Franchina T, Tuccari G and Adamo V:
Correction: androgen receptor (AR), E-cadherin, and Ki-67 as
emerging targets and novel prognostic markers in triple-negative
breast cancer (TNBC) patients. PLoS One. 10:e01326472015.Erratum
for: PLoS One 10: e0128368, 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Nishimura N and Sasaki T: Cell-surface
biotinylation to study endocytosis and recycling of occludin.
Methods Mol Biol. 440:89–96. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Martin TA, Jordan N, Davies EL and Jiang
WG: Metastasis to Bone in human cancer is associated with loss of
occludin expression. Anticancer Res. 36:1287–1293. 2016.PubMed/NCBI
|
|
42
|
Eriksson JE, Dechat T, Grin B, Helfand B,
Mendez M, Pallari HM and Goldman RD: Introducing intermediate
filaments: From discovery to disease. J Clin Invest. 119:1763–1771.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Peuhu E, Virtakoivu R, Mai A, Wärri A and
Ivaska J: Epithelial vimentin plays a functional role in mammary
gland development. Development. 144:4103–4113. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ungefroren H: TGF-β Signaling in cancer:
control by negative regulators and crosstalk with proinflammatory
and fibrogenic pathways. Cancers (Basel). 11:3842019. View Article : Google Scholar
|
|
45
|
Walker RA and Dearing SJ: Transforming
growth factor beta 1 in ductal carcinoma in situ and invasive
carcinomas of the breast. Eur J Cancer. 28:641–644. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Grau AM, Wen W, Ramroopsingh DS, Gao YT,
Zi J, Cai Q, Shu XO and Zheng W: Circulating transforming growth
factor-beta-1 and breast cancer prognosis: Results from the
Shanghai Breast Cancer Study. Breast Cancer Res Treat. 112:335–341.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Parvani JG, Taylor MA and Schiemann WP:
Noncanonical TGF-β signaling during mammary tumorigenesis. J
Mammary Gland Biol Neoplasia. 16:127–146. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Xu J, Lamouille S and Derynck R:
TGF-beta-induced epithelial to mesenchymal transition. Cell Res.
19:156–172. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Oft M, Peli J, Rudaz C, Schwarz H, Beug H
and Reichmann E: TGF-beta1 and Ha-Ras collaborate in modulating the
phenotypic plasticity and invasiveness of epithelial tumor cells.
Genes Dev. 10:2462–2477. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Okada H, Ban S, Nagao S, Takahashi H,
Suzuki H and Neilson EG: Progressive renal fibrosis in murine
polycystic kidney disease: An immunohistochemical observation.
Kidney Int. 58:587–597. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Oldfield MD, Bach LA, Forbes JM,
Nikolic-Paterson D, McRobert A, Thallas V, Atkins RC, Osicka T,
Jerums G and Cooper ME: Advanced glycation end products cause
epithelial-myofibroblast transdifferentiation via the receptor for
advanced glycation end products (RAGE). J Clin Invest.
108:1853–1863. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Oloumi A, McPhee T and Dedhar S:
Regulation of E-cadherin expression and beta-catenin/Tcf
transcriptional activity by the integrin-linked kinase. Biochim
Biophys Acta. 1691:1–15. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Colak S and Ten Dijke P: Targeting TGF-β
Signaling in Cancer. Trends Cancer. 3:56–71. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Kennecke H, Yerushalmi R, Woods R, Cheang
MC, Voduc D, Speers CH, Nielsen TO and Gelmon K: Metastatic
behavior of breast cancer subtypes. J Clin Oncol. 28:3271–3277.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Shibue T and Weinberg RA: EMT, CSCs, and
drug resistance: The mechanistic link and clinical implications.
Nat Rev Clin Oncol. 14:611–629. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Al-Hajj M, Becker MW, Wicha M, Weissman I
and Clarke MF: Therapeutic implications of cancer stem cells. Curr
Opin Genet Dev. 14:43–47. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Ashaq A, Maqbool MF, Maryam A, Khan M,
Shakir HA, Irfan M, Qazi JI, Li Y and Ma T: Hispidulin: A novel
natural compound with therapeutic potential against human cancers.
Phytother Res (In press).
|
|
58
|
Liu K, Zhao F, Yan J, Xia Z, Jiang D and
Ma P: Hispidulin: A promising flavonoid with diverse anti-cancer
properties. Life Sci. 259:1183952020.in press. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Scoparo CT, Valdameri G, Worfel PR,
Guterres FA, Martinez GR, Winnischofer SM, Di Pietro A and Rocha
ME: Dual properties of hispidulin: Antiproliferative effects on
HepG2 cancer cells and selective inhibition of ABCG2 transport
activity. Mol Cell Biochem. 409:123–133. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Gao H, Jiang Q, Han Y, Peng J and Wang C:
Hispidulin potentiates the antitumor effect of sunitinib against
human renal cell carcinoma in laboratory models. Cell Biochem
Biophys. 71:757–764. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Wang Y, Liu W, He X and Fei Z: Hispidulin
enhances the anti-tumor effects of temozolomide in glioblastoma by
activating AMPK. Cell Biochem Biophys. 71:701–706. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Yang JM, Hung CM, Fu CN, Lee JC, Huang CH,
Yang MH, Lin CL, Kao JY and Way TD: Hispidulin sensitizes human
ovarian cancer cells to TRAIL-induced apoptosis by AMPK activation
leading to Mcl-1 block in translation. J Agric Food Chem.
58:10020–10026. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Cong Y, Wu S, Han J, Chen J, Liu H, Sun Q,
Wu Y and Fang Y: Pharmacokinetics of homoplantaginin in rats
following intravenous, peritoneal injection and oral
administration. J Pharm Biomed Anal. 129:405–409. 2016. View Article : Google Scholar : PubMed/NCBI
|