Introduction
Melanoma is malignant form of skin cancer, due to
its high metastatic potential. BRAF mutants are well-known
oncogenic drivers in malignant melanoma, and small molecules have
been developed to target BRAF, including vemurafenib and dabrafenib
(1–3). In recent years, monoclonal antibodies
against programmed cell death protein 1 (PD-1) and cytotoxic
T-lymphocyte protein 4 (CTLA-4) have been administered to melanoma
patients as immune checkpoint inhibitors (4,5).
These drugs suppress the progression of melanoma, but
melanoma-specific mechanisms of tumorigenesis are incompletely
understood.
Melanocytes, the cells from which melanomas
originate, express tyrosinase (TYR) to produce melanin. TYR is a
type I membrane glycoprotein that catalyzes the hydroxylation of
L-tyrosine and the oxidation of L-3,4-dihydroxyphenylalanine-the
rate-limiting reactions in the synthesis of melanin (6,7). The
reaction intermediates act as not only substrates for melanin
synthesis but also promotors of melanogenesis (8). In melanocytic cells, melanin
synthesis is regulated by secreted hormones.
α-melanocyte-stimulating hormone (α-MSH), a representative
melanogenesis-stimulating hormone, initially binds to
melanocortin-1 receptor (MC1R) on the cell surface. MC1R is a
seven-transmembrane G protein-coupled receptor and upregulates cAMP
synthesis in an adenylyl cyclase-dependent manner (9,10).
cAMP functions as a second messenger that activates protein kinase
A (PKA), effecting the phosphorylation of cAMP response
element-binding protein (CREB) (11). Phosphorylated CREB then upregulates
the transcription of microphthalmia-associated transcription factor
(MITF), resulting in the expression of several melanogenic genes,
including TYR (12).
MITF-amplified melanoma is malignant, and
melanin deposition is often considered to be a hallmark of
malignant melanoma (13). However,
melanogenesis is not essential for tumorigenesis in melanocytes
because amelanotic melanomas exist. By contrast, Hendrix and
colleagues have suggested that the expression of TYR is low in
aggressive melanoma (14).
Moreover, the loss of TYR correlates with poor survival in melanoma
(15). Although these reports
suggest that TYR suppresses the progression of melanoma, there is
no direct evidence that TYR functions as a tumor suppressor.
Vasculogenic mimicry (VM) is one means by which
blood is supplied for tumor growth. During VM, vascular-like
networks are formed by tumor cells, instead of by vascular
endothelial cells (16). VM was
first described in uveal melanoma by Maniotis et al in 1999
(17) and has been observed in
several aggressive cancers, such as breast, ovarian, prostate, and
lung cancer and sarcoma (18–22).
VM is associated with an extremely poor prognosis in melanoma
patients (23–26) and thus is a crucial factor in
aggressive melanoma. In addition, VM is linked to metastasis in
tumor cells (27). Given that
melanoma cells have high metastatic potential, certain
melanoma-specific proteins might regulate the onset of VM and cell
motility. Important regulators of VM, including VE-cadherin, have
previously been identified (28),
but the tissue-specific mediators of VM remain unknown.
In this study, we found that stimulators of
melanogenesis inhibit VM in MNT-1 human melanoma cells. Because TYR
is central to the melanin synthesis pathway, we focused on TYR and
determined its effects on VM. We observed that TYR negatively
regulates VM in human pigmented and amelanotic melanoma cell lines.
Further, a loss-of-function TYR mutant did not downregulate the
development of VM. Our findings constitute evidence that the
enzymatic activity of TYR is crucial for the suppression of VM in
melanomas.
Materials and methods
Cell culture
The MNT-1 human pigmented melanoma cell line, kindly
gifted by Profs. Michael S. Marks (Children's Hospital of
Philadelphia and University of Pennsylvania, PA) and Cheah Shiau
Chuen (UCSI University, Kuala Lumpur, Malaysia), was cultured with
Dulbecco's Modified Eagle Medium (Nissui Pharmaceutical, Tokyo,
Japan) that was supplemented with 7% (v/v) FBS, 10% (v/v) AIM-V
Medium liquid (Thermo Fisher Scientific, Inc. Waltham, MA), 100
U/ml penicillin G, 100 mg/l kanamycin, 2.25 g/l NaHCO3,
and 600 mg/l L-glutamine at 37°C in a humidified incubator with 5%
CO2. The SK-MEL-28 human amelanotic melanoma (RIKEN
BioResource Center, Tsukuba, Japan), WM266-4 (American Type Culture
Collection, Manassas, VA), 293T human embryonic kidney (RIKEN
BioResource Research Center), and HT1080 human fibrosarcoma
(Japanese Collection of Research Bioresources Cell Bank, Osaka,
Japan) cell lines were cultured in Dulbecco's Modified Eagle Medium
that was supplemented with 7% (v/v) FBS, 100 U/ml penicillin G, 100
mg/l kanamycin, 2.25 g/l NaHCO3, and 600 mg/l
L-glutamine at 37°C in a humidified incubator with 5%
CO2.
Reagents
Arbutin (Merck KGaA, Darmstadt, Germany) and
α-melanocyte-stimulating hormone (α-MSH; Peptide Institute, Inc.,
Osaka, Japan) were dissolved in sterilized water.
3-isobutyl-1-methylxanthine (IBMX; FUJIFILM Wako Pure Chemical
Corporation, Osaka, Japan) was dissolved in dimethyl sulfoxide
(DMSO).
Melanin content assay
Cell pellets were dissolved with 10% (v/v) DMSO that
contained 1 N NaOH at 70°C for 1 h. The melanin content was
quantified by measuring the absorbance at 405 nm. Each absorbance
value was normalized by the amount of total protein.
TYR activity assay
The enzymatic activity of cellular TYR was
quantified using the Tyrosinase Activity Assay Kit (Abcam #
ab252899; Cambridge, UK) according to the manufacturer's
instructions.
VM assay
The in vitro VM assay was conducted as
described (29–31). Initially, 96-well plates were
coated with 40 µl/well of Matrigel® Growth Factor
Reduced (Corning, Corning, NY) and incubated for 30 min at 37°C.
Cells were suspended and added to the Matrigel-coated wells at
2.0×104 cells/well and cultured at 37°C in a humidified
incubator with 5% CO2. In each well, images of five
independent, randomly selected fields were captured using
phase-contrast microscopy (Leica DMi1, Leica, Wetzlar, Germany),
and the number of tubes was counted. A tube was defined as an area
that was surrounded by cells.
MTT assay
Cells were seeded at 2.0×103 cells/well
in the presence of vehicle, IBMX, or α-MSH in 96-well plates and
cultured for 48 h. Thiazolyl blue tetrazolium bromide was added to
each well, after which the cells were cultured for 4 h at 37°C. The
media was removed, and the MTT formazan product was dissolved in
100 µl DMSO. The absorbance at 570 nm was measured to quantify the
number of living cells.
Establishment of TYR knockout
cells
Knockout (KO) of TYR was performed using the
CRISPR/Cas9 system as described (31,32).
We used the D10A Cas9 mutant to avoid off-target effects (called
the Nickase system). Thus, we designed 2 nearby targets in exon 1
of TYR; the sequences of the oligonucleotides for generating
the guide RNAs were as follows: target 1,
5′-CACCGGGCTCTAGGGAAATGGCCAG-3′ (forward) and
5′-AAACCTGGCCATTTCCCTAGAGCCC-3′ (reverse); and target 2,
5′-CACCGTGTCTCCTCTAAGAACCTGA-3′ (forward) and
5′-AAACTCAGGTTCTTAGAGGAGACAC-3′ (reverse). Each pair of
oligonucleotides was annealed and inserted into the BbsI
restriction site of pSpCas9n(BB)-2A-Puro (PX462) V2.0 (gifted by
Feng Zhang, Addgene, Cambridge, MA). MNT-1 and SK-MEL-28 cells were
cotransfected with these plasmids using Lipofectamine 3000™ (Thermo
Fisher Scientific, Inc.) and then treated with 1.25 µg/ml puromycin
dihydrochloride (Merck KGaA) to select transfectants. Clonal TYR-KO
cells were established by limiting dilution method.
Construction of TYR expression
vectors
TYR cDNA was amplified from the
pcDNA4(TO)-tyrosinase plasmid (33), a kind gift of Prof. Takafumi
Hasegawa (Tohoku University, Sendai, Japan), by polymerase chain
reaction (PCR) using the following primers:
5′-TTTTCTCGAGATGCTCCTGGCTGTTTTGTACTGC-3′ (forward) and
5′-TTTTGCGGCCGCTTATAAATGGCTCTGATACAAGCTGTGG-3′ (reverse). To avoid
recognition by Cas9, we constructed Cas9-resistant TYR cDNA
by overlap extension PCR with the following primers:
5′-GCGTGAGCAGCAAAAATCTCATGGAAAAGGAATGCTGTCCACCGTG-3′ (forward) and
5′-AAGCCCGTGGAAAGTGTCCGGCGCTGGTCTGGAAACTCCACAGCAG-3′ (reverse). The
T373K point mutation was generated by overlap extension PCR with
the following primers: 5′-AATGGAAAAATGTCCCAGGTACAGGGATCTG-3′
(forward) and 5′-TTTGGATGAAATAAAGAAATCACCATTTCTG-3′ (reverse). The
resulting amplicons were inserted into the XhoI/NotI
restriction site of CSII-CMV-MCS-IRES2-Bsd (RIKEN BioResource
Center). The CSII-CMV-MCS-IRES2-Bsd-GFP plasmid (34) was used as a control.
293T cells were transfected with these plasmids
using Lentivirus High-Titer Packaging Mix (Takara Bio Inc.) and
cultured for 6 h. The cells were washed with phosphate-buffered
saline and cultured with fresh medium for 42 h to facilitate the
production of virus particles. TYR-KO MNT-1 and WM266-4 cells were
then treated with the lentivirus-containing conditioned media.
After infection, cells were selected with 12.5 µg/ml blasticidin S
(FUJIFILM Wako Pure Chemical Corporation).
Western blot
Western blot was performed as described (31,35).
Cells were cultured and lysed in lysis buffer [50 mM Tris-HCl pH
7.5, 150 mM NaCl, 0.1% (w/v) SDS, 1% (v/v) Triton X-100, 1% (w/v)
sodium deoxycholate, and 1 mM phenylmethylsulfonyl fluoride] with
PhosSTOP phosphatase inhibitor cocktail (Merck KGaA) on ice with
sonication. The lysate was centrifuged at 15,300 × g for 10 min,
and the supernatant was collected. The amount of protein in each
cell lysate was measured by Coomassie Brilliant Blue G-250 staining
(Bio-Rad Laboratories, Inc., Hercules, CA).
Loading buffer [350 mM Tris-HCl pH 6.8, 30% (v/v)
glycerol, 0.012% (w/v) bromophenol blue, 6% (w/v) SDS, and 30%
(v/v) 2-mercaptoethanol] was added to each lysate and boiled for 3
min. The samples were electrophoresed on 9% SDS-polyacrylamide
gels, after which the proteins were transferred to polyvinylidene
fluoride membranes. The membranes were blocked with 5% skim milk at
room temperature for 30 min and immunoblotted with monoclonal
anti-TYR (Abcam # ab170905), monoclonal anti-α-tubulin (Merck KGaA
#T5168), monoclonal anti-CREB (Cell Signaling Technology #9197;
Danvers, MA), and anti-phospho-CREB (Cell Signaling Technology
#9191) at room temperature for 1 h. HRP-linked anti-rabbit IgG
(Cytiva #NA934; Marlborough, MA) and HRP-linked anti-mouse IgG
(Cytiva #NA931) were added to the membranes for 1 h at room
temperature. Signals were detected by enhanced chemiluminescence
using Western Lightning Plus-ECL (PerkinElmer, Inc., Waltham, MA)
or Immobilon Western Chemiluminescent HRP substrate (Merck KGaA)
and exposed to RX-U films (FUJIFILM, Tokyo, Japan) in a dark
room.
Statistical analysis
Differences between 2 groups were analyzed by
two-tailed student's t-test (unpaired). Datasets with 3 groups or
over were analyzed using one-way ANOVA with Tukey's test using SPSS
(version 27; IBM, Armonk, NY). The results were expressed as mean ±
SD. P<0.05 was considered to indicate a statistically
significant difference.
Results
Stimulation with IBMX and α-MSH
inhibits VM in MNT-1 cells
To determine the significance of cAMP/CREB/TYR
signaling in VM, we treated MNT-1 cells with the cAMP signaling
activator IBMX and confirmed that IBMX induces phosphorylation of
CREB and upregulates TYR (Fig. 1A and
B). IBMX also inhibited VM in MNT-1 cells (Fig. 1C). Further, α-MSH, a potent
activator of cAMP signaling, increased CREB phosphorylation and TYR
levels (Fig. 1A and B). Consistent
with this result, α-MSH impeded VM in MNT-1 cells (Fig. 1D).
 | Figure 1.Stimulation with IBMX and α-MSH
inhibits VM in MNT-1 cells. (A and B) MNT-1 cells were treated with
1 µM α-MSH, 100 µM IBMX or 1 µM α-MSH + 100 µM IBMX for (A) 30 min
or (B) 48 h. The cells were lysed, and western blotting was
performed. (C) MNT-1 cells were pretreated with vehicle or 100 µM
IBMX for 48 h, and VM assay was performed. (left) Images of VM were
captured 5 h after cell seeding; representative images are
presented (scale bars, 200 µm). (right) VM was quantified by
counting tube numbers. (D and E) MNT-1 cells were pretreated with
vehicle, (D) 1 µM α-MSH or (E) 100 µM IBMX + 1 µM α-MSH for 48 h,
and VM assay was performed. (upper) Images of VM were captured 5 h
after cell seeding; representative images are presented (scale
bars, 200 µm). (lower) VM was quantified by counting tube numbers.
Data shown are the means ± SD (n=5). *P<0.05, **P<0.01,
***P<0.001. ND, not detected; p-, phosphorylated; Ctrl, control;
IBMX, 3-Isobutyl 1-methylxanthine; a-MSH, α-melanocyte-stimulating
hormone; VM, vasculogenic mimicry; TYR, tyrosinase. |
Cotreatment with IBMX and α-MSH enhanced the
expression of TYR and inhibited VM in MNT-1 cells (Fig. 1B and E). IBMX and α-MSH did not
affect cell viability individually or in combination (Fig. S1), confirming that their
suppressive activities on VM were not attributed to cell death.
These data suggest that IBMX and α-MSH inhibit VM, consistent with
the activation of the cAMP/CREB/TYR axis.
TYR inhibitor promotes VM in MNT-1
cells
Because α-MSH and IBMX upregulated TYR (Fig. 1B), we examined the function of TYR
in VM. Arbutin is a well-known TYR inhibitor and has inhibitory
effects on melanin synthesis (36), and we confirmed the reduction of
melanin content in arbutin-treated MNT-1 cells (Fig. 2A). By contrast, arbutin promoted VM
(Fig. 2B and C), prompting us to
study the effects of arbutin on VM in non-melanoma cell lines.
HT1080 is a TYR-non-expressing tumor cell line (Fig. S2A). By VM assay, arbutin did not
increase tube numbers in HT1080 cells (Fig. S2B), indicating that arbutin
suppresses VM by inhibiting TYR in tumor cell lines.
TYR suppresses VM in melanoma cell
lines
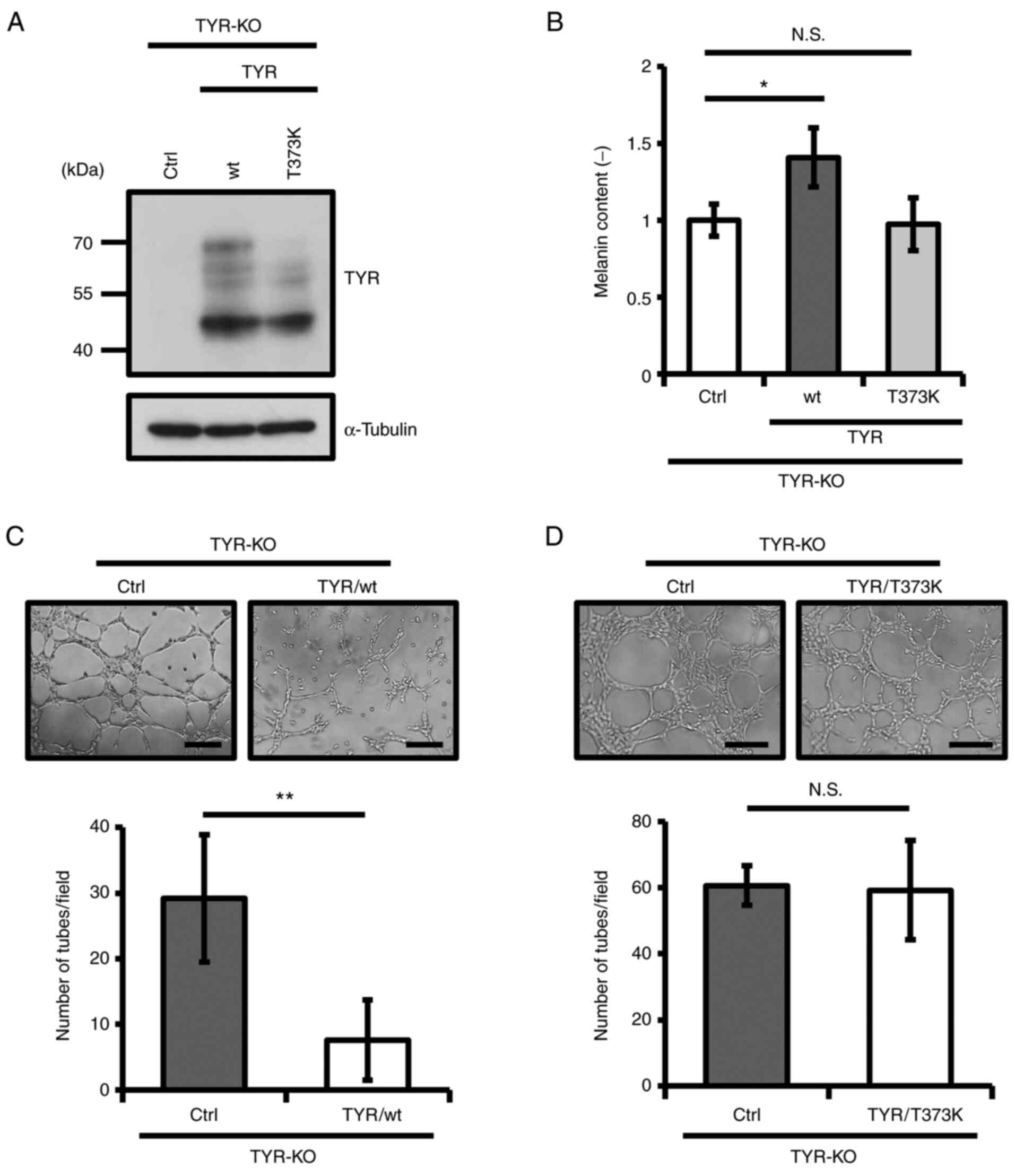
To verify the function of TYR in VM, we established
a TYR-KO MNT-1 cell line using the CRISPR/Cas9 system (Fig. 3A). As expected, the enzymatic
activity of TYR decreased significantly, and thus, melanin content
was diminished in TYR-KO MNT-1 cells (Fig. S3A and SB). Consistent with the
results after treatment with arbutin, VM was promoted in TYR-KO
MNT-1 cells (Fig. 3B). Given that
amelanotic melanomas also express TYR endogenously, we examined
whether TYR has suppressive activity against VM even in amelanotic
melanoma cells. To test this, we deleted TYR in SK-MEL-28
human amelanotic melanoma cells by CRISPR/Cas9 and confirmed its
enzymatic activity (Figs. 3C and
S4A). As shown in Fig. 3D, depletion of TYR promoted VM in
SK-MEL-28, as well as pigmented MNT-1 cells. We also confirmed that
overexpression of TYR increases its enzymatic activity and
attenuates VM in WM266-4 human amelanotic melanoma cells (Fig. S4B-SD). In addition, KO of TYR
attenuated inhibitory effect of α-MSH on VM in MNT-1 cells
(Figs. 1D and S5). These results demonstrate that TYR
suppresses VM in pigmented and amelanotic melanoma cells.
Enzymatic activity of TYR is critical
for TYR-mediated inhibition of VM
Human TYR often carries mutations, some of
which cause albinism (37). The
T373K mutation is frequently observed in albinos, attenuating the
enzymatic activity of TYR (38).
Thus, we re-expressed wild-type (wt) or T373K TYR in TYR-KO MNT-1
cells to establish TYR-rescued MNT-1 cell lines (Fig. 4A). Whereas re-expression of wt TYR
rescued its enzymatic activity and melanin production,
re-expression of T373K TYR did not, as expected (Figs. 4B and S6). Notably, rescue with wt TYR
decreased tube numbers, but T373K TYR did not affect VM in TYR-KO
MNT-1 cells (Fig. 4C and D). These
results suggest that the enzymatic activity of TYR is required for
regulating VM.
Discussion
Advanced cancer is difficult to prevent using
surgical and pharmaceutical approaches, necessitating the
identification of clear hallmarks of aggressiveness in tumors to
treat patients. In the past 2 decades, VM has garnered interest as
an indicator of tumor malignancy (27,28),
but the mechanisms by which it develops are poorly understood.
Melanoma is an aggressive and metastatic tumor, and numerous
reports have demonstrated that VM causes a poor prognosis in
melanoma patients (23–26). In this study, we aimed to determine
the melanoma-specific molecular mechanisms of VM.
Pigmentation is a unique property of melanomas. cAMP
facilitates melanin synthesis through downstream signaling; thus,
we treated MNT-1 human pigmented melanoma cells with IBMX and
α-MSH, which enhance the activity of the cAMP/PKA axis (39). As a result, these compounds
significantly inhibited VM, and the inhibition of TYR promoted it,
indicating that the activation of TYR and the consequent synthesis
of melanin correlate negatively with the potential for VM. However,
TYR regulated VM even in SK-MEL-28 and WM266-4 human amelanotic
melanoma cells. Thus, TYR itself might be a negative regulator of
VM without melanin synthesis. It has been suggested that the
enzymatic activity of TYR regulates some biological events
(40,41). Our data reinforce this concept,
because enzymatically inactive TYR did not affect VM. On the other
hand, numerous reports have indicated that melanin production
affects various cellular behaviors in normal and malignant
melanocytes (42–45). Therefore, future work is warranted
to determine whether the presence of melanin affects VM.
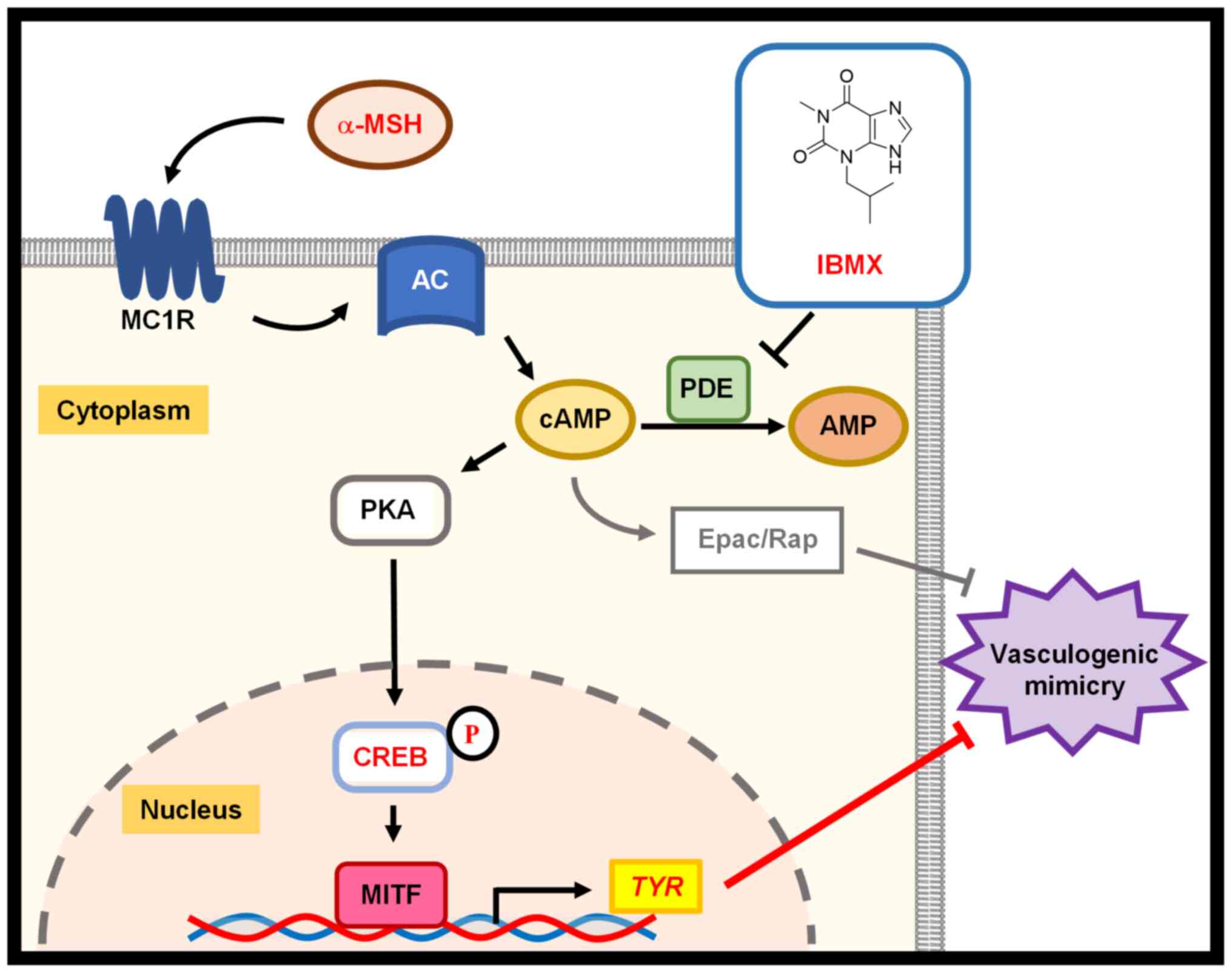
cAMP activates several signaling pathways and
suppresses VM in melanoma cells through cAMP/Epac/Rap1 signaling
(46). However, whether other
pathways that are stimulated by cAMP affect VM is unknown (46,47).
In the current study, we focused on the cAMP/PKA/CREB/TYR axis,
because this pathway is an important cascade in melanogenesis. Our
results demonstrated that IBMX and α-MSH suppress VM with the
upregulation of phosphorylated CREB and TYR in MNT-1 cells. α-MSH
decreased tube numbers in TYR-KO MNT-1 cells, albeit to a lesser
extent than in parental MNT-1 cells. Thus, TYR is critical for
CREB-mediated regulation of VM (Fig.
5).
Epidemiological data suggest that
melanoma-associated hypopigmentation after immunological therapy
for metastatic melanoma correlates with an improved prognosis
(48–50). Furthermore, inhibition of
melanogenesis leads to favorable results in the treatment of
melanoma (51–53). However, several reports indicate
that depigmentation of melanoma constitutes a sign of tumor
progression that accompanies greater metastasis (54–57).
Because VM is closely related to the high metastatic potential of
tumors, TYR expression might be a salient marker of the low
potential for metastasis and VM in melanomas. Because
antigen-specific T cells recognize TYR and are involved in tumor
rejection (48,58), a loss of TYR might affect immune
escape from CD8+ T cells (49). Further, TYR per se downregulates
cell migration, cell survival, epithelial mesenchymal transition,
and tumorigenesis in melanoma (54,59).
Thus, the loss of TYR might allow melanoma cells to escape the
immune system and tumor-suppressive activity, accelerating tumor
progression.
We have unveiled a novel function for
TYR-suppression of VM in human melanoma cells, independent of its
melanogenic activity. Our findings provide new insights into
melanoma-specific mechanisms of tumorigenesis, guiding the
development of therapeutic approaches for melanoma patients in whom
VM arises.
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to thank Dr Takafumi Hasegawa
and Mr Shun Ishiyama (Tohoku University, Sendai, Japan) for
providing the pcDNA4(TO)-tyrosinase plasmid. The authors also thank
Professor Midori A. Arai and Dr Shun Saito (Keio University,
Yokohama, Japan) for their advice.
Funding
This work was supported by JSPS KAKENHI (grant no.
JP20J11197).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HK, RK and SS designed the study. HK performed all
experiments and analyzed the data. HK and RK confirmed the
authenticity of all the raw data. HK, RK and SS wrote the original
draft. All authors have read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
VM
|
vasculogenic mimicry
|
|
α-MSH
|
α-melanocyte-stimulating hormone
|
|
IBMX
|
3-Isobutyl 1-methylxanthine
|
|
cAMP
|
cyclic adenosine monophosphate
|
|
TYR
|
tyrosinase
|
References
|
1
|
Davies H, Bignell GR, Cox C, Stephens P,
Edkins S, Clegg S, Teague J, Woffendin H, Garnett MJ, Bottomley W,
et al: Mutations of the BRAF gene in human cancer. Nature.
417:949–954. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chapman PB, Hauschild A, Robert C, Haanen
JB, Ascierto P, Larkin J, Dummer R, Garbe C, Testori A, Maio M, et
al: Improved survival with vemurafenib in melanoma with BRAF V600E
mutation. N Engl J Med. 364:2507–2516. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hauschild A, Grob JJ, Demidov LV, Jouary
T, Gutzmer R, Millward M, Rutkowski P, Blank CU, Miller WH Jr,
Kaempgen E, et al: Dabrafenib in BRAF-mutated metastatic melanoma:
A multicentre, open-label, phase 3 randomised controlled trial.
Lancet. 380:358–365. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Robert C, Long GV, Brady B, Dutriaux C,
Maio M, Mortier L, Hassel JC, Rutkowski P, McNeil C,
Kalinka-Warzocha E, et al: Nivolumab in previously untreated
melanoma without BRAF mutation. N Engl J Med. 372:320–330. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hodi FS, O'Day SJ, McDermott DF, Weber RW,
Sosman JA, Haanen JB, Gonzalez R, Robert C, Schadendorf D, Hassel
JC, et al: Improved survival with ipilimumab in patients with
metastatic melanoma. N Engl J Med. 363:711–723. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kwon BS, Haq AK, Pomerantz SH and Halaban
R: Isolation and sequence of a cDNA clone for human tyrosinase the
maps at the mouse c-albino locus. Proc Natl Acad Sci USA.
84:7473–7477. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ujvari A, Aron R, Eisenhaure T, Cheng E,
Parag HA, Smicun Y, Halaban R and Hebert DN: Translation rate of
human tyrosinase determines its N-linked glycosylation level. J
Biol Chem. 276:5924–5931. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Slominski A, Zmijewski MA and Pawelek J:
L-tyrosine and L-dihydroxyphenylalanine as hormone-like regulators
of melanocyte functions. Pigment Cell Melanoma Res. 25:14–27. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Herraiz C, Martínez-Vicente I and Maresca
V: The α-melanocyte-stimulating hormone/melanocortin-1 receptor
interaction: A driver of pleiotropic effects beyond pigmentation.
Pigment Cell Melanoma Res. 34:748–761. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Slominski A, Tobin DJ, Shibahara S and
Wortsman J: Melanin pigmentation in mammalian skin and its hormonal
regulation. Physiol Rev. 84:1155–1228. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Misra UK and Pizzo SV: Coordinate
regulation of forskolin-induced cellular proliferation in
macrophages by protein kinase A/cAMP-response element-binding
protein (CREB) and Epac1-Rap1 signaling: Effects of silencing CREB
gene expression on Akt activation. J Biol Chem. 280:38276–38289.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Price ER, Horstmann MA, Wells AG,
Weilbaecher KN, Takemoto CM, Landis MW and Fisher DE:
a-Melanocyte-stimulating hormone signaling regulates expression of
microphthalmia, a gene deficient in Waardenburg syndrome. J Biol
Chem. 273:33042–33047. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Garraway LA, Widlund HR, Rubin MA, Getz G,
Berger AJ, Ramaswamy S, Beroukhim R, Milner DA, Granter SR, Du J,
et al: Integrative genomic analyses identify MITF as a lineage
survival oncogene amplified in malignant melanoma. Nature.
436:117–122. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hendrix MJ, Seftor EA, Hess AR and Seftor
RE: Vasculogenic mimicry and tumour-cell plasticity: Lessons from
melanoma. Nat Rev Cancer. 3:411–421. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Takeuchi H, Kuo C, Morton DL, Wang HJ and
Hoon DS: Expression of differentiation melanoma-associated antigen
genes is associated with favorable disease outcome in
advanced-stage melanomas. Cancer Res. 63:441–448. 2003.PubMed/NCBI
|
|
16
|
Folberg R, Hendrix MJ and Maniotis AJ:
Vasculogenic mimicry and tumor angiogenesis. Am J Pathol.
156:361–381. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Maniotis AJ, Folberg R, Hess A, Seftor EA,
Gardner LM, Pe'er J, Trent JM, Meltzer PS and Hendrix MJ: Vascular
channel formation by human melanoma cells in vivo and in vitro:
Vasculogenic mimicry. Am J Pathol. 155:739–752. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shirakawa K, Tsuda H, Heike Y, Kato K,
Asada R, Inomata M, Sasaki H, Kasumi F, Yoshimoto M, Iwanaga T, et
al: Absence of endothelial cells, central necrosis, and fibrosis
are associated with aggressive inflammatory breast cancer. Cancer
Res. 61:445–451. 2001.PubMed/NCBI
|
|
19
|
Sood AK, Seftor EA, Fletcher MS, Gardner
LM, Heidger PM, Buller RE, Seftor RE and Hendrix MJ: Molecular
determinants of ovarian cancer plasticity. Am J Pathol.
158:1279–1288. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sharma N, Seftor RE, Seftor EA, Gruman LM,
Heidger PM Jr, Cohen MB, Lubaroff DM and Hendrix MJ: Prostatic
tumor cell plasticity involves cooperative interactions of distinct
phenotypic subpopulations: Role in vasculogenic mimicry. Prostate.
50:189–201. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Passalidou E, Trivella M, Singh N,
Ferguson M, Hu J, Cesario A, Granone P, Nicholson AG, Goldstraw P,
Ratcliffe C, et al: Vascular phenotype in angiogenic and
non-angiogenic lung non-small cell carcinomas. Br J Cancer.
86:244–249. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
van der Schaft DW, Hillen F, Pauwels P,
Kirschmann DA, Castermans K, Egbrink MG, Tran MG, Sciot R, Hauben
E, Hogendoorn PC, et al: Tumor cell plasticity in Ewing sarcoma, an
alternative circulatory system stimulated by hypoxia. Cancer Res.
65:11520–11528. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Seftor RE, Seftor EA, Koshikawa N, Meltzer
PS, Gardner LM, Bilban M, Stetler-Stevenson WG, Quaranta V and
Hendrix MJ: Cooperative interactions of laminin 5 g2 chain, matrix
metalloproteinase-2, and membrane type-1-matrix/metalloproteinase
are required for mimicry of embryonic vasculogenesis by aggressive
melanoma. Cancer Res. 61:6322–6327. 2001.PubMed/NCBI
|
|
24
|
Clarijs R, Otte-Höller I, Ruiter DJ and de
Waal RM: Presence of a fluid-conducting meshwork in xenografted
cutaneous and primary human uveal melanoma. Invest Ophthalmol Vis
Sci. 43:912–918. 2002.PubMed/NCBI
|
|
25
|
Mueller AJ, Maniotis AJ, Freeman WR,
Bartsch DU, Schaller UC, Bergeron-Lynn G, Cheng L, Taskintuna I,
Chen X, Kan-Mitchell J and Folberg R: An orthotopic model for human
uveal melanoma in SCID mice. Microvasc Res. 64:207–213. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Thies A, Mangold U, Moll I and Schumacher
U: PAS-positive loops and networks as a prognostic indicator in
cutaneous malignant melanoma. J Pathol. 195:537–542. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wei X, Chen Y, Jiang X, Peng M, Liu Y, Mo
Y, Ren D, Hua Y, Yu B, Zhou Y, et al: Mechanisms of vasculogenic
mimicry in hypoxic tumor microenvironments. Mol Cancer. 20:72021.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Delgado-Bellido D, Serrano-Saenz S,
Fernández-Cortés M and Oliver FJ: Vasculogenic mimicry signaling
revisited: Focus on non-vascular VE-cadherin. Mol Cancer.
16:652017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Schnegg CI, Yang MH, Ghosh SK and Hsu MY:
Induction of vasculogenic mimicry overrides VEGF-A silencing and
enriches stem-like cancer cells in melanoma. Cancer Res.
75:1682–1690. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Williamson SC, Metcalf RL, Trapani F,
Mohan S, Antonello J, Abbott B, Leong HS, Chester CP, Simms N,
Polanski R, et al: Vasculogenic mimicry in small cell lung cancer.
Nat Commun. 7:133222016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kawahara R, Niwa Y and Simizu S: Integrin
β1 is an essential factor in vasculogenic mimicry of human cancer
cells. Cancer Sci. 109:2490–2496. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ran FA, Hsu PD, Wright J, Agarwala V,
Scott DA and Zhang F: Genome engineering using the CRISPR-Cas9
system. Nat Protoc. 8:2281–2308. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hasegawa T: Tyrosinase-expressing neuronal
cell line as in vitro model of Parkinson's disease. Int J Mol Sci.
11:1082–1089. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mizuta H, Kuga K, Suzuki T, Niwa Y, Dohmae
N and Simizu S: C-mannosylation of R-spondin2 activates
Wnt/β-catenin signaling and migration activity in human tumor
cells. Int J Oncol. 54:2127–2138. 2019.PubMed/NCBI
|
|
35
|
Tamura Y, Simizu S, Muroi M, Takagi S,
Kawatani M, Watanabe N and Osada H: Polo-like kinase 1
phosphorylates and regulates Bcl-xL during pironetin-induced
apoptosis. Oncogene. 28:107–116. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Akiu S, Suzuki Y, Asahara T, Fujinuma Y
and Fukuda M: Inhibitory effect of arbutin on
melanogenesis-biochemical study using cultured B16 melanoma cells.
Nihon Hifuka Gakkai Zasshi. 101:609–613. 1991.(In Japanese).
PubMed/NCBI
|
|
37
|
Opitz S, Käsmann-Kellner B, Kaufmann M,
Schwinger E and Zühlke C: Detection of 53 novel DNA variations
within the tyrosinase gene and accumulation of mutations in 17
patients with albinism. Hum Mutat. 23:630–631. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Halaban R, Svedine S, Cheng E, Smicun Y,
Aron R and Hebert DN: Endoplasmic reticulum retention is a common
defect associated with tyrosinase-negative albinism. Proc Natl Acad
Sci USA. 97:5889–5894. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Busca R and Ballotti R: Cyclic AMP a key
messenger in the regulation of skin pigmentation. Pigment Cell Res.
13:60–69. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Slominski A, Moellmann G and Kuklinska E:
L-tyrosine, L-dopa, and tyrosinase as positive regulators of the
subcellular apparatus of melanogenesis in Bomirski Ab amelanotic
melanoma cells. Pigment Cell Res. 2:109–116. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Slominski A and Paus R: Towards defining
receptors for L-tyrosine and L-dopa. Mol Cell Endocrinol.
99:C7–C11. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Slominski A, Kim TK, Brożyna AA,
Janjetovic Z, Brooks DL, Schwab LP, Skobowiat C, Jóźwicki W and
Seagroves TN: The role of melanogenesis in regulation of melanoma
behavior: Melanogenesis leads to stimulation of HIF-1a expression
and HIF-dependent attendant pathways. Arch Biochem Biophys.
563:79–93. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Slominski RM, Zmijewski MA and Slominski
AT: The role of melanin pigment in melanoma. Exp Dermatol.
24:258–259. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Slominski A, Paus R and Schadendorf D:
Melanocytes as ‘sensory’ and regulatory cells in the epidermis. J
Theor Biol. 164:103–120. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Slominski A: Neuroendocrine activity of
the melanocyte. Exp Dermatol. 18:760–763. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Lissitzky JC, Parriaux D, Ristorcelli E,
Vérine A, Lombardo D and Verrando P: Cyclic AMP signaling as a
mediator of vasculogenic mimicry in aggressive human melanoma cells
in vitro. Cancer Res. 69:802–809. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wang S, Zhang Z, Qian W, Ji D, Wang Q, Ji
B, Zhang Y, Zhang C and Sun Y, Zhu C and Sun Y: Angiogenesis and
vasculogenic mimicry are inhibited by 8-Br-cAMP through activation
of the cAMP/PKA pathway in colorectal cancer. Onco Targets Ther.
11:3765–3774. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Robbins PF, El-Gamil M, Kawakami Y,
Stevens E, Yannelli JR and Rosenberg SA: Recognition of tyrosinase
by tumor-infiltrating lymphocytes from a patient responding to
immunotherapy. Cancer Res. 54:3124–3126. 1994.PubMed/NCBI
|
|
49
|
Sanchez-Perez L, Kottke T, Diaz RM, Ahmed
A, Thompson J, Chong H, Melcher A, Holmen S, Daniels G and Vile RG:
Potent selection of antigen loss variants of B16 melanoma following
inflammatory killing of melanocytes in vivo. Cancer Res.
65:2009–2017. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Vavricka CJ, Christensen BM and Li J:
Melanization in living organisms: A perspective of species
evolution. Protein Cell. 1:830–841. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Brożyna AA, Jóźwicki W, Roszkowski K,
Filipiak J and Slominski AT: Melanin content in melanoma metastases
affects the outcome of radiotherapy. Oncotarget. 7:17844–17853.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Slominski A, Zbytek B and Slominski R:
Inhibitors of melanogenesis increase toxicity of cyclophosphamide
and lymphocytes against melanoma cells. Int J Cancer.
124:1470–1477. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Slominski A, Paus R and Mihm MC:
Inhibition of melanogenesis as an adjuvant strategy in the
treatment of melanotic melanomas: Selective review and hypothesis.
Anticancer Res. 18:3709–3715. 1998.PubMed/NCBI
|
|
54
|
Fürst K, Steder M, Logotheti S, Angerilli
A, Spitschak A, Marquardt S, Schumacher T, Engelmann D,
Herchenröder O, Rupp RAW and Pützer BM: DNp73-induced degradation
of tyrosinase links depigmentation with EMT-driven melanoma
progression. Cancer Lett. 442:299–309. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Tas F: Melanoma-associated
hypopigmentation in association with locoregional relapse of
melanoma depigmentation. Surgery. 150:1011–1012. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Bennett DC: Differentiation in mouse
melanoma cells: Initial reversibility and an on-off stochastic
model. Cell. 34:445–453. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Pinner S, Jordan P, Sharrock K, Bazley L,
Collinson L, Marais R, Bonvin E, Goding C and Sahai E: Intravital
imaging reveals transient changes in pigment production and Brn2
expression during metastatic melanoma dissemination. Cancer Res.
69:7969–7977. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Hearing VJ: Biochemical control of
melanogenesis and melanosomal organization. J Investig Dermatol
Symp Proc. 4:24–28. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Sekine Y, Togi S, Muromoto R, Kon S, Kitai
Y, Yoshimura A, Oritani K and Matsuda T: STAP-2 protein expression
in B16F10 melanoma cells positively regulates protein levels of
tyrosinase, which determines organs to infiltrate in the body. J
Biol Chem. 290:17462–17473. 2015. View Article : Google Scholar : PubMed/NCBI
|