Hepatocellular carcinoma (HCC) is the most
frequently diagnosed cancer worldwide and the fourth leading cause
of cancer-related mortality (1).
According to the Global Cancer Burden Data Report from the Global
Cancer Observatory platform (https://gco.iarc.fr/projects), World Health
Organization, there were 905,677 new cases of liver cancer in 2020,
of which 72.5% occurred in Asia. The total deaths from liver cancer
were 830,180 in 2020. Despite the advances in surgical excision and
systematic treatment, the prognosis of patients with HCC remains
poor (2). This may be due to the
high rate of tumor recurrence and metastasis. Hypoxia is a hallmark
of the solid tumor microenvironment. Activation of the
transcription factor hypoxia-inducible factor (HIF) enables cancer
cells to adapt to the hypoxic environment via the transactivation
of downstream target genes (3).
The human genome encodes three different HIF subtypes: HIF-1α,
HIF-2α and HIF-3α (4). Recent
studies have shown that the HIF-2α expression level in HCC is
correlated with clinical progression and poor survival (5), which promotes HCC metastasis by
promoting the epithelial-mesenchymal transition (EMT) pathway,
promoting lncRNA NEAT1 activation and inducing stem cell factor
(SCF) transcription (6–8). Moreover, HIF-1α plays an important
role in HCC metastasis under hypoxic conditions (9), but the underlying mechanism remains
unclear. This review focuses on the role of HIF-1α in tumor
metastasis, including linked signaling pathways, their impact on
the resistance to current first-line therapies, and their
implication in future therapies. HIF-1α is expected to become a
critical target for the treatment of HCC in the future.
The significant sequelae of most solid tumors
include hypoxia and necrosis. Abnormal tumor microvessels, blocked
microcirculation, and impaired diffusion lead to insufficient or no
oxygen supply in the tumor microenvironment (TME) (10). HIF is an important biosensor for
the oxygen concentration in tumors. HIF is a heterodimer that
consists of an O2-sensitive HIF-1α subunit and an
O2-insensitive HIF-1β subunit (11).
Hypoxia-induced gene expression mainly depends on
the stability of the α subunit of HIF-1α, which is an
oxygen-unstable subunit whose transcriptional activity is regulated
by cellular oxygen tension (12).
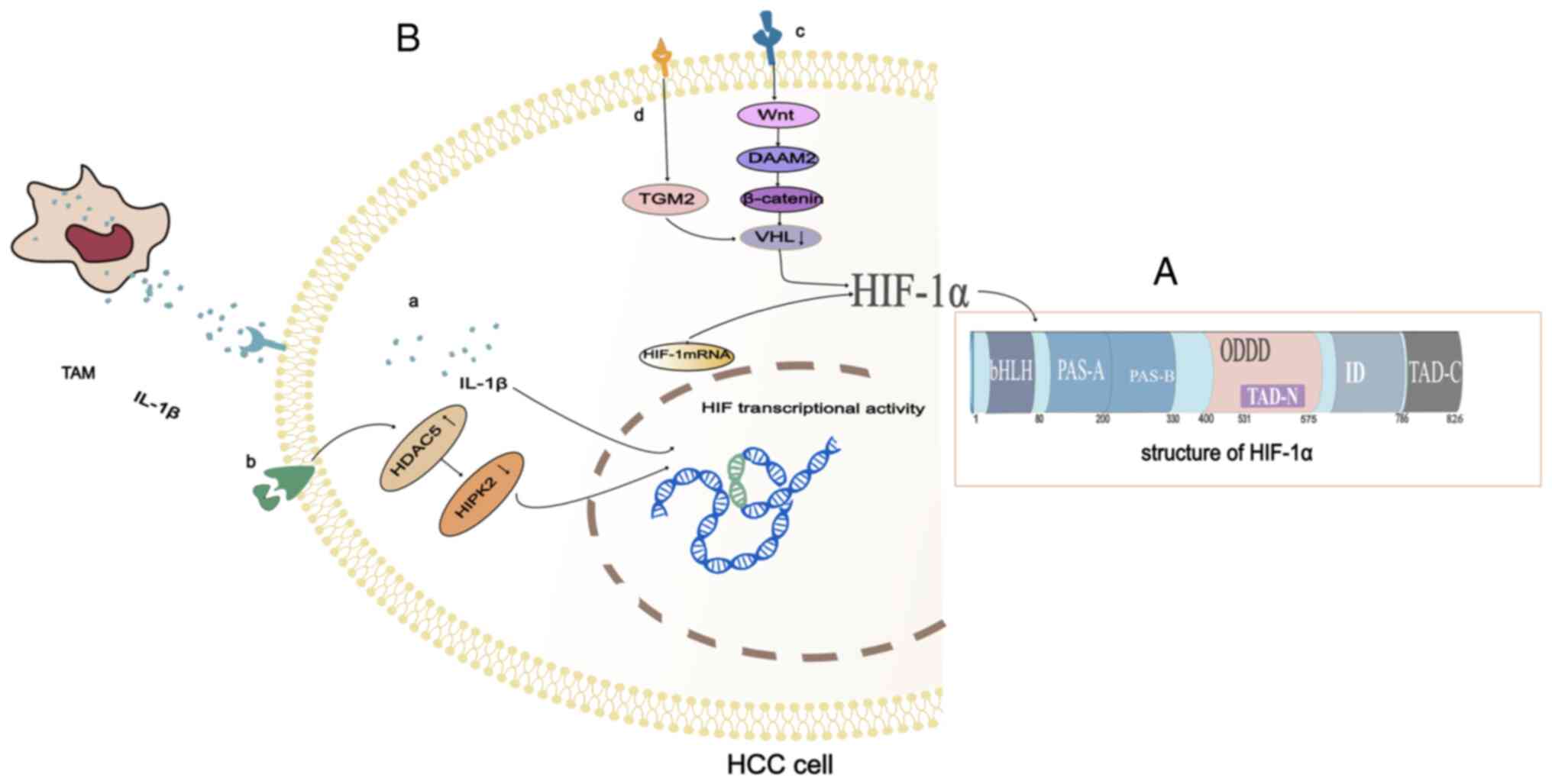
HIF-1α has two transactivation domains (TAD). The COOH terminal
transactivation domain (C-TAD) is located in amino acid residues
from 786 to 826 in humans. The C-TAD is recruited by co-activators,
and the co-activators CBP and p300 regulate the transactivation of
target genes (13). The
NH2-terminal transactivation domain (N-TAD) is located
in a region called the oxygen-dependent degradation domain (ODDD),
which is positioned in amino acid residues from 400 to 600
(Fig. 1A). The ODDD is only
recognized by von Hippel-Lindau tumor suppressor protein (pVHL)
under normoxic conditions, and pVHL is required to mediate the
degradation of HIF-1α in the ubiquitin-proteasome pathway. The ODDD
contains two key proline residues that target hydroxylation under
normoxic conditions. Under normal oxygen levels, HIF-1α undergoes
hydroxylation of the proline. It is then recognized by the pVHL E3
ligase and degraded by the proteasome (14). In a hypoxic environment, an
inadequate oxygen supply leads to impaired hydroxylation and
degradation of the HIF-1α. This causes the HIF-1α to stabilize and
the stable HIF-1α is then transferred to the nucleus. It acts as a
heterodimer trans-activator with a nuclear protein called an aryl
hydrocarbon receptor nuclear transposer (ARNT). The resulting
heterodimer (HIF-1α/ARNT) specifically interacts with the hypoxia
response element (HRE) to increase the transcription of genes
involved in angiogenesis, erythropoiesis, and glycolysis (15). The HIF-1α and HIF-1β play critical
roles in angiogenesis, proliferation, invasion, and cancer
metabolism (16,17). Therefore, the ODDD controls the
activity and stability of the α subunit, and the carboxy-terminal
region of the HIF-1α represents the stability domain of the
protein.
HIF-1α has been confirmed to be closely related to
tumor metastasis, but the role of HIF-1α in the metastasis of
primary HCC has not been systematically discussed. This review aims
to comprehensively elaborate on the role of HIF-1α in the
metastasis of HCC.
The progression of hepatocellular carcinoma (HCC)
depends on its local microenvironment. Hypoxia and inflammation are
two key factors affecting the microenvironment of HCC (24,25).
Under moderate hypoxia, the stability of HIF-1α is increased and
tumor-associated macrophages secrete more interleukin-1β (IL-1β).
In the presence of persistent and severe hypoxia, the necrotic
debris of HCC cells mediates the release of potent IL-1β from
tumor-associated macrophages with an M2 phenotype via
interferon-β/nuclear factor kappa-light-chain-enhancer containing
Toll-like receptor 4/TIR domains that activate B-cell signaling.
The IL-1β in HCC cells promotes HIF-1α synthesis through
cyclooxygenase 2 (COX-2) (26).
Inflammation and hypoxia interact to promote the development of
tumors (Fig. 1B-a).
In addition to the interaction of hypoxia and
inflammation, the increase in epigenetic regulators and the
Dishevelled-associated activator of morphogenesis (DAAM) can also
promote the high expression of HIF-1α in hepatoma cells. As an
epigenetic regulator, histone deacetylase (HDAC) is essential for
activating gene transcription under physiological conditions and is
also involved in the occurrence and development of tumors. HDAC5 is
a histone deacetylase with a high expression in HCC. An increase in
HDAC5 leads to silencing of homeodomain-interacting protein
kinase-2 (HIPK2), which induces the transcription of HIF-1α
resulting in high expression of HIF-1α (27) (Fig.
1B-b). DAAM consisting of DAAM1 and DAAM2 mainly mediates the
coordination of form-dependent actin remodeling (28). DAAM controls cell shape and
polarized cell growth by regulating the actin cytoskeleton. In HCC
cells, DAAM2 is a Wnt signal transduction effector that acts
downstream of the Wnt ligand and upstream of β-catenin. DAAM2
reduces Von Hippel-Lindau (VHL) expression and thus upregulates
HIF-1α (29) (Fig. 1B-c).
Pseudohypoxia also causes HIF-1α hyperexpression.
Activated hepatic stellate cells promote epithelial-mesenchymal
transformation (EMT) in HCC through pseudohypoxia promoted by
transglutaminase 2 (TGM2) (30).
Activated hepatic stellate cells upregulate TGM2 in HCC cells
through inflammatory signaling (31). VHL induced by TGM2 under normoxic
conditions is the key molecule of HIF-1α degradation (32). Under normoxic conditions, TGM2 can
cause VHL consumption, resulting in HIF-1α accumulation and
pseudohypoxia, thus promoting EMT in HCC cells (30) (Fig.
1B-d).
The accumulation of HIF-1α was also found to be
related to its stability. Hydroxylation of proline residues by
proline hydroxylase domain enzymes (PHDs) is a key step in the
degradation of HIF-1α (33).
Prolyl hydroxylase domain-containing protein 2 (PHD2) requires
O2 and α-ketoglutarate as substrates for hydroxylation
of HIF-1α. O-linked β-N-acetylglucosamine transferase
(OGT) can stabilize HIF-1α by reducing the level of α-ketoglutarate
(34). Recent results show that
HAUSP (USP7) is a deubiquitinase of HIF-1α (35). HAUSP can undergo hypoxia-induced
K63-linked polyubiquitination by HectH9 to enhances its ability to
deubiquitinate HIF-1α and also serve as a scaffold of
HIF-1α-induced gene transcription (36). Plasmacytoma variant translocation 1
(PVT1) is a long non-coding (lnc)RNA that has been found to play an
oncogenic role in a variety of malignant tumors. Lysine
acetyltransferase 2A (KAT2A) is a histone acetyltransferase.
Research shows that lncRNA PVT1 stabilizes HIF-1α via KAT2A
(37). Research has shown that the
STAT3 protein binds to HIF-1α through competition with pVHL, thus
stabilizing HIF-1α protein levels (38). GATA binding protein 3 interacts
with the full-length and the N-terminal part of HIF-1α (aa 1–401)
under hypoxia to inhibit ubiquitination of HIF-1α (39).
The expression of HIF-1α in HCC tissues was found to
be significantly higher than that in chronic hepatitis B and normal
liver tissues, but there was no significant difference when
compared with that in the cirrhotic liver (40,41).
A study analyzing the clinicopathological features in patients with
HCC showed that upregulation of HIF-1α mRNA expression was strongly
associated with TNM stage III and Barcelona clinical hepatocellular
carcinoma (BCLC) stage C. The survival analysis showed that HCC
patients with high HIF-1α mRNA expression had reduced overall
survival when compared with patients with low HIF-1α mRNA
expression (42). HIF-1α
expression was positively correlated with vascular invasion, TNM
stage, tumor size, and portal vein tumor thromboembolism in HCC
patients with cirrhosis. The patients with high HIF-1α expression
on the basis of liver cirrhosis had reduced overall survival (OS)
when compared with the patients with low HIF-1α expression
(40). However, a meta-analysis
involving 851 patients with HCC showed no association between
HIF-1α protein expression and HCC envelope formation, cirrhosis,
tumor size, or tumor differentiation, except for vascular invasion
(43).
As a prognostic factor, high HIF-1α expression was
found to be associated with poor disease-free survival and poor
prognosis (44). A meta-analysis
of 34 studies involving 3,578 patients showed that HIF-1α
overexpression was associated with poor OS, disease-free survival
(DFS), and relapse-free survival (RFS) in HCC patients.
Overexpression of HIF-1α was also associated with clinicopathologic
features including BCLC, intrahepatic metastasis, lymph node
metastasis, tumor-to-lymph node metastasis (TNM), tumor
differentiation, tumor number, tumor size, vascular invasion, and
angiogenesis (5). Another
meta-analysis involving 3,238 patients in 22 studies also showed
poor prognosis in HCC patients presenting with overexpression of
HIF-1α (45). However, other
studies showed that HIF-1α protein expression was associated with
tumor differentiation, and intrahepatic and extrahepatic
metastases, but not with the presence of portal vein tumor
embolism, prognosis, or hepatitis B surface antigen (HBsAg) status
(41).
Metastasis is a major challenge in the treatment of
cancers. The vast majority of cancer-related deaths are caused by
metastatic conditions in vital organs (46). In a solid tumor, the abnormal new
vascular system of the tumor and the increase in oxygen consumption
by proliferating HCC cells are not balanced, and thereby there is
always hypoxia in HCC tissue. Hypoxia promotes the invasion and
migration of HCC, and HIF-1α is also upregulated in HCC (47). Our understanding of the molecular
basis of metastasis has improved in recent years, and the
hypoxia-induced transcription factor (HIF) remains one of the best
indicators for monitoring metastasis (48).
Enhancement of HIF activity drives tumor progression
by regulating the expression of hundreds of genes. These genes are
closely associated with tumor progressions such as immune escape,
glycolysis, cancer stem cell maintenance, angiogenesis, EMT, and
cancer stem cell maintenance (Table
I) (26,47,49–61).
HIF signals affect almost every step of the cascade leading to
tumor metastasis (48,62).
Changes in the development of immunosuppressive
mechanisms by tumor cells play a crucial role in tumor development,
which allows them to escape the host's immune system, thereby
enhancing their survival and proliferative, migratory, and invasive
capabilities (63). Hypoxia, which
is associated with an imbalance in rapid tumor growth and an
insufficient blood supply, is a common change that occurs in the
microenvironment of solid tumors (64). Previous studies have found that
HIF-1α can increase the number of myeloid-derived suppressor cells
(MDSCs) (49,50) and plasmacytoid dendritic cells
(pDCs) (51), and can induce IL-1β
(26,52,65)
expression in the TME leading to immune escape.
MDSCs possess immunosuppressive activities, which
allow cancers to escape immune surveillance and become
non-responsive to immune checkpoint blockade. HIF-1α induces
ectonucleoside triphosphate diphosphohydrolase 2 (ENTPD2) in cancer
cells to increase the extracellular level of 5′-AMP, which
maintains MDSC undifferentiation in the tumor stroma (49). MDSCs enhance tumor cell stemness,
increase angiogenesis, and advance the metastatic process by
promoting EMT though IL-6 secretion (50). pDCs play immunosuppressive roles in
the TME. HIF-1α was found to transcriptionally upregulate the
expression of extracellular adenosine (eADO), and eADO was found to
significantly enhance pDC recruitment into tumors via the adenosine
A1 receptor (ADORA1) leading to immunosuppression (51). Tumor-associated macrophages, the
primary proinflammatory cells within tumors, secrete more IL-1β
under hypoxic conditions due to increased stability of HIF-1α
(26). Another study demonstrated
that IL-1β could increase the expression of solute carrier family 7
member 11, which facilitated HCC metastasis through colony
stimulating factor 1-induced tumour-associated macrophage and MDSC
infiltration in tumors (52).
The glycolytic phenotype destroys normal tissues and
promotes the secretion of proteolytic enzymes such as cathepsin B
or metalloproteinases through lactate and H+-mediated
extracellular environmental acidification, thereby promoting
metastasis and invasion (66). As
a result, glycolysis plays an important role in tumor invasion and
metastasis. Recently, it was found that HIF-1α can enhance the
process of glycolysis in HCC cells. HIF-1α directly binds to the
hypoxia response element (HRE) in the promoter region of uroplakin
1A (UPK1A), leading to UPK1A being upregulated under hypoxia,
thereby increasing the glycolysis of HCC cells (53). HIF-1α activates transcription of
lncRNA RAET1K through miR-100-5p to enhance hypoxia-induced
glycolysis in HCC cells (54). In
addition, hypoxia-inducible lncRNA NPSR1-AS1 promotes the
proliferation and glycolysis of HCC cells by regulating the
MAPK/ERK pathway (55).
Yes-associated protein (YAP) binds to HIF-1α and sustains HIF-1α
protein stability to promote HCC cell glycolysis under hypoxic
stress (47).
EMT is a highly conserved cellular program during
which epithelial cells lose their polarized organization and
acquire migratory and invasive capabilities, which has been
recognized as a pro-metastatic cellular event that promotes tumor
cell invasion and malignant tumor progression (67). HIF-1α induces EMT to promote
metastasis through many pathways. i) Lysyl oxidases like 2 (LOXL2)
is a member of the lysyl oxidase family, whose main function is to
catalyze the covalent cross-linkages of collagen and elastin in the
extracellular matrix. HIF-1α was found to induce EMT, HCC cell
migration, invasion and vasculogenic mimicry (VM) formation by
regulating LOXL2 (56). ii)
Vasodilator-stimulated phosphoprotein (VASP) is a regulator of the
actin cytoskeleton and cell migration. HIF-1α induces VASP
overexpression by directly binding two HREs in the VASP promoter
region, and promotes the expression of EMT, MMP2 and MMP9 (57). iii) Hypoxia increases the
production of reactive oxygen species (ROS) by inducing
nicotinamide adenine dinucleotide phosphate (NADPH) oxidase 4
(NOX4). ROS can induce the GLI1-dependent EMT process and promote
the invasion and metastasis of HCC (58). iv) Epithelial (E)-cadherin controls
proliferation and cell polarization through intercellular adhesion
and is essential for maintaining epithelial structure and
homeostasis (68). E-cadherin
activation inhibits metastasis at multiple stages, including CTC
accumulation in the primary tumor and extravasation of tumor cells
in the vascular system (69).
HIF-1α inhibits transcription of E-cadherin by upregulating Snai1
and SIP1, a transcriptional inhibitor of E-cadherin (70), and activates EMT in HCC cells and
promotes HCC invasion and metastasis (59). v) HIF-1α downregulates the
Wnt/β-catenin pathway and enhances EMT in HCC cells by inducing
expression of miR-1273F in exosomes (60).
Cancer stem cells (CSCs) are a small subset of tumor
cells with the capability to influence self-renewal,
differentiation, and tumorigenesis. As far as we know, the
inflammatory microenvironment in HCC leads to the proliferation of
stem cells with genetic or epigenetic alterations, facilitating
their transformation from normal liver stem cells to liver cancer
stem cells (LCSCs) (71). LCSCs
play critical roles in regulating HCC stemness, self-renewal,
tumorigenicity, metastasis (72).
Accumulating evidence has demonstrated that CSCs are
a primary cause of metastasis due to their role in new tumor
initiation at local or distant sites (73–75).
The generation of CSCs is closely related to the EMT mechanism
(76), and β-catenin is one of the
markers of EMT. Research has found that the Notch and Wnt/β-catenin
signaling pathways promote the stemness characteristics of LCSCs.
Expression of transcription factors involving EMT (such as Snail)
and stemness (such as Sox2 and Nanog) can be decreased by blocking
the function of Wnt/β-catenin and/or Notch (77). As mentioned above, stabilization of
HIF-1α in hypoxic environments induces the EMT process and thus
also modulates HCC stemness. Recent research has found that hypoxia
enhances HIF-1α stability and transcriptional activity by promoting
HIF-1α deSUMOylation of sentrin-specific protease 1 (SENP1), and
HIF-1α promotes HCC stem cell by increasing the expression levels
of CD24, CD44 and CD133 (61).
Sorafenib, a kinase inhibitor drug, exerts
anti-angiogenic and anti-metastatic effects by dose-dependent
inhibition of HIF-1α and VEGF protein expression (78). However, the anti-angiogenic
activity by continuous sorafenib therapy can lead to tumor
starvation and intratumor hypoxia, which may facilitate the
generation of resistant cell clones adapted to hypoxia and nutrient
deficiency (79). Sustained
sorafenib treatment increases intratumoral hypoxia, which
stabilizes the HIF-1α protein, thereby reducing the sensitivity to
sorafenib. Liang et al (80) suggested that hypoxia induced by
continuous sorafenib treatment could lead to sorafenib resistance
in HCC through activation of HIF-1α and nuclear factor (NF)-κB. It
was also found that the β-2-adrenergic receptor (ADRB2) signal
destroys the beclin1/VPS34/Atg14 complex in an Akt-dependent
manner, which negatively regulates autophagy and leads to stability
of HIF-1α, reprogramming of glucose metabolism in HCC cells, and
resistance to sorafenib (81). A
significant correlation between the hypoxic microenvironment and
sorafenib resistance was demonstrated, suggesting that targeting
HIF is a promising approach to improve therapeutic efficiency.
Targeting of HIF-1α can improve the efficacy of
sorafenib. The combination of sorafenib and curcumin analog EF24
inhibited the hypoxic resistance to sorafenib by promoting the
proteasome degradation of the VHL-dependent HIF-1α in HCC cells,
resulting in the inhibition of target genes MDR1 and GLUT-1, and a
decrease in the activities of VEGF and NF-κB (80). The use of natural compounds has
also shown positive effects in improving sorafenib treatment.
Genistein, a natural isoflavone, was found to enhance the antitumor
effect of sorafenib in sorafenib-resistant HCC cells and HCC
xenograft mouse models by downregulating HIF-1α, thereby
inactivating Glut-1 and HK2 to inhibit and sensitize aerobic
glycolysis in HCC cells, resulting in mitochondrial apoptosis
(82). Simvastatin that belongs to
a group of HMG-CoA reductase inhibitors, inhibits the
HIF-1α/PPAR-γ/PKM2 axis by hindering PKM2-mediated glycolysis,
resulting in decreased proliferation and increased apoptosis of HCC
cells, and re-sensitization of HCC cells to sorafenib (83).
Transarterial chemoembolization (TACE) is considered
to be one of the most effective palliative care for patients with
unresectable HCC, and the BCLC staging system has defined it as the
standard of care for patients with mid-stage HCC (84). Previous research has shown that the
overexpression of HIF-1α reduces the effectiveness of TACE against
HCC (85). The expression of the
HIF-1α protein in HCC tissues was found to be increased after TACE
surgery. To adapt to the hypoxic environment, HIF-1α stimulates the
expression of COX-2 protein in HCC cells and promotes the EMT
process, thus enhancing the invasion and metastasis of HCC, and
this may lead to poor prognosis of HCC patients after TACE
treatment (86). Therefore,
improving the hypoxic state after TACE or reducing the expression
of HIF-1α may improve the prognosis of these patients.
The innate and adaptive immune systems play an
important role in anticancer immune surveillance. Immune checkpoint
inhibitors (ICIs) have proven that an effective immune response is
able to eliminate tumor cells (87). Currently, preclinical evidence and
clinical use of immunotherapy in HCC includes the use of programmed
cell death-1 (PD-1), programmed death-ligand 1 (PDL1) and cytotoxic
T lymphocyte-associated antigen 4 (CTLA4) inhibitors, such as
nivolumab, atezolizumab and ipilimumab (88). Although immunotherapy brings hope
to the treatment of HCC, its efficacy is still limited, which may
be related to the complex TME in HCC (89). Research has revealed the increased
HIF-1α levels in the hypoxic TME causing the downregulation of
T-cell activity and induction of PD-L1 on the surface of tumor
cells, resulting in immune suppression and increased cell
proliferation of tumor cells resulting in immune escape (90). Therefore, inhibition of HIF-1α
expression represents a promising novel strategy in cancer
immunotherapy (91).
Studies have shown that HIPK2, USP22, and NF-κB can
regulate the expression of HIF-1α. Some drugs, such as metformin,
heme alkali, dandelion polysaccharide regulate HIF-1α and inhibit
the proliferation and metastasis of hepatoma cells. This provides
some novel ideas for the treatment of HCC.
Serine/threonine homologous domain interaction
protein kinase 2 (HIPK2) is often downregulated in HCC tissues.
HIPK2 directly binds to HIF-1α and stimulates HIF-1α ubiquitination
to degrade proteasomes. The downregulation of HIPK2 can enhance the
angiogenesis of HCC by activating the HIF-1α pathway and further
promote tumor growth and metastasis. The downregulation of HIPK2
protein expression in HCC patients is associated with poor overall
survival (92). Ubiquitin-specific
proteinase 22 (USP22) can stabilize HIF-1α by deubiquitination.
Promoting hypoxia can induce HCC stem cells and glycolysis
(93). As the direct target genes
of HIF-1α, USP22 and TP53 can be upregulated by HIF-1α
transcription in hypoxia. The upregulation of TP53 inhibited the
upregulation of USP22. In HCC cells with TP53 mutation, USP22 and
HIF-1α form a positive feedback loop promoting HCC stem cells. With
the loss of function mutation at TP53 and USP22 and/or HIF-1α, HCC
patients with high expression tend to have poor prognoses. The
lipoprotein complex targeting the USP22 highly induced tumor
inhibition and enhanced the sensitivity to sorafenib in
tumor-bearing mice. USP22 promotes hypoxia-induced HCC stem cells
through the TP53-inactivated HIF-1α/USP22 positive feedback loop.
USP22 is a promising target for HCC treatment (94). A study has assessed the temporal
expression of HIF-1α and NF-κB subunits in HCC under short-term and
long-term hypoxia and found that NF-κB regulates HIF-1α in hypoxic
HCC (95). The NF-κB subunits, p50
and p65 were found to enhance HIF-1α transcription, while c-Rel
downstream miRNAs (including miR-199a-5p and miR-93) and Dicer1
promoted HIF-1α degradation (96).
Targeting this regulator temporarily in the early hypoxic
microenvironment may be a new therapeutic option for early cancer
intervention.
Metformin-mediated HIF-1α inactivation leads to a
decrease in PFKFB3 expression, which further inhibits
phospho-fructose kinase-1 (PFK1) activity. Metformin significantly
inhibits HCC cell proliferation by blocking the PFK1 glycolytic
flux. The HIF-1α/PFKFB3/pfk1 regulatory axis is an important
determinant of glucose metabolism reprogramming in HCC, which may
provide an effective therapeutic target to inhibit the HCC
(97).
Heme base is a benzo phenanthridine alkaloid, which
can inhibit HIF-1α signal transduction and EMT marker expression,
Snail translocation, Smad and PI3K-AKT pathway activation. One
study found that the heme base is a promising drug candidate that
will target HIF-1α/TGF-β signal transduction to improve treatment
in patients with HCC (98).
Dandelion polysaccharide (DP) is an α-type
polysaccharide derived from the root of the dandelion. It consists
of glucose, galactose, arabinose, arabinose rhamnose, and
glucuronic acid (99,100). It was found that dandelion
polysaccharide inhibited HIF-1α expression by regulating the
PI3K/Akt signaling pathway and exhibited an anti-HCC cell
proliferation, anti-metastasis, and angiogenesis role (101).
Hypoxia is a typical microenvironmental condition in
almost all solid tumors, including HCC (102). HIF-1α is highly expressed in a
hypoxic environment and is associated with the metastasis and
prognosis of HCC (64). This
phenomenon has aroused research attention to the role of HIF-1α in
promoting tumor progression. This review summarizes the role of
HIF-1α in the metastasis and prognosis of HCC. Here, we revealed
the mechanisms of HIF-1α in promoting the development of HCC and
developing clinical drug resistance, as well as its predictive
effect on the postoperative recurrence of HCC, which has important
clinical significance for the treatment and prognosis of HCC.
Detection of the HIF-1α expression level can be considered in the
routine assessment of HCC, which is of great significance for the
treatment and prognosis of HCC.
However, researchers are still faced with challenges
when they switch from laboratory discovery to clinical application
by targeting HIF-1α to inhibit the metastasis and development of
HCC. Moreover, there are few experimental studies on inhibiting
HIF-1α clinically in the treatment of HCC. In an open study,
patients with HCC who participated in clinical trials were given
intravenous (i.v.) infusion of drugs to prove the mechanism of the
inhibition of HIF-1α by reducing HIF-1α mRNA (nct02564614). The
study results have not been published yet. One observational study
on the comparison of HIF-1α levels (nct00866957) in patients with
various HCC treatments is being carried out by Dr Laura Kulik of
the Feinberg School of Medicine at Northwestern University
(Chicago, IL, USA).
Therefore, there is an urgent need for further
studies involving multiple centers, higher sample size, and a
long-time follow-up for the treatment and control of metastatic HCC
by targeting HIF-1α. It is believed that it will become a clinical
reality in the near future.
Not applicable.
This study was supported by the Natural Science Foundation of
Beijing, China (grant nos. 7212171 and 7212172) and the Pilot
Project of Public Welfare Development and Reform of Beijing
Municipal Medical Research Institutes (2019–6).
Not applicable.
HC, WL and XL conceived and designed the review. HC,
HY and JC were involved in the collection and collation of
references. HY and JC put forward meaningful suggestions for the
revision of the article and participated in the revision of the
manuscript. HC and WL designed the figures. HC wrote the
manuscript. Data authentication is not applicable. All authors read
and approved the final manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Liu X, Zhang X, Peng Z, Li C, Wang Z, Wang
C, Deng Z, Wu B, Cui Y, Wang Z, et al: Deubiquitylase OTUD6B
Governs pVHL stability in an enzyme-independent manner and
suppresses hepatocellular carcinoma metastasis. Adv Sci (Weinh).
7:19020402020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Forner A, Reig M and Bruix J:
Hepatocellular carcinoma. Lancet. 391:1301–1314. 2018. View Article : Google Scholar
|
|
3
|
Wang X, Li L, Zhao K, Lin Q, Li H, Xue X,
Ge W, He H, Liu D, Xie H, et al: A novel LncRNA HITT forms a
regulatory loop with HIF-1α to modulate angiogenesis and tumor
growth. Cell Death Differ. 27:1431–1446. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Guo Y, Xiao Z, Yang L, Gao Y, Zhu Q, Hu L,
Huang D and Xu Q: Hypoxia-inducible factors in hepatocellular
carcinoma (Review). Oncol Rep. 43:3–15. 2020.PubMed/NCBI
|
|
5
|
Méndez-Blanco C, Fernández-Palanca P,
Fondevila F, González-Gallego J and Mauriz JL: Prognostic and
clinicopathological significance of hypoxia-inducible factors 1α
and 2α in hepatocellular carcinoma: A systematic review with
meta-analysis. Ther Adv Med Oncol. 13:17588359209870712021.
View Article : Google Scholar
|
|
6
|
Mu H, Yu G, Li H, Wang M, Cui Y, Zhang T,
Song T and Liu C: Mild chronic hypoxia-induced HIF-2α interacts
with c-MYC through competition with HIF-1α to induce hepatocellular
carcinoma cell proliferation. Cell Oncol (Dordr). 44:1151–1166.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang X, Dong J, Jia L, Zhao T, Lang M, Li
Z, Lan C, Li X, Hao J, Wang H, et al: HIF-2-dependent expression of
stem cell factor promotes metastasis in hepatocellular carcinoma.
Cancer Lett. 393:113–124. 2017. View Article : Google Scholar
|
|
8
|
Chen J, Chen J, Huang J, Li Z, Gong Y, Zou
B, Liu X, Ding L, Li P, Zhu Z, et al: HIF-2α upregulation mediated
by hypoxia promotes NAFLD-HCC progression by activating lipid
synthesis via the PI3K-AKT-mTOR pathway. Aging (Albany NY).
11:10839–10860. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tian H, Huang P, Zhao Z, Tang W and Xia J:
HIF-1α plays a role in the chemotactic migration of hepatocarcinoma
cells through the modulation of CXCL6 expression. Cell Physiol
Biochem. 34:1536–1546. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Albadari N, Deng S and Li W: The
transcriptional factors HIF-1 and HIF-2 and their novel inhibitors
in cancer therapy. Expert Opin Drug Discov. 14:667–682. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Prabhakar NR and Semenza GL: Adaptive and
maladaptive cardiorespiratory responses to continuous and
intermittent hypoxia mediated by hypoxia-inducible factors 1 and 2.
Physiol Rev. 92:967–1003. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jiang BH, Zheng JZ, Leung SW, Roe R and
Semenza GL: Transactivation and inhibitory domains of
hypoxia-inducible factor 1alpha. Modulation of transcriptional
activity by oxygen tension. J Biol Chem. 272:19253–19260. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lando D, Peet DJ, Gorman JJ, Whelan DA,
Whitelaw ML and Bruick RK: FIH-1 is an asparaginyl hydroxylase
enzyme that regulates the transcriptional activity of
hypoxia-inducible factor. Genes Dev. 16:1466–1471. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chen C and Lou T: Hypoxia inducible
factors in hepatocellular carcinoma. Oncotarget. 8:46691–46703.
2017. View Article : Google Scholar
|
|
15
|
Hur E, Kim HH, Choi SM, Kim JH, Yim S,
Kwon HJ, Choi Y, Kim DK, Lee MO and Park H: Reduction of
hypoxia-induced transcription through the repression of
hypoxia-inducible factor-1alpha/aryl hydrocarbon receptor nuclear
translocator DNA binding by the 90-kDa heat-shock protein inhibitor
radicicol. Mol Pharmacol. 62:975–982. 2002. View Article : Google Scholar
|
|
16
|
Luo D, Wang Z and Wu J, Jiang C and Wu J:
The role of hypoxia inducible factor-1 in hepatocellular carcinoma.
Biomed Res Int. 2014:4092722014. View Article : Google Scholar
|
|
17
|
Mossenta M, Busato D, Dal Bo M and Toffoli
G: Glucose metabolism and oxidative stress in hepatocellular
carcinoma: Role and possible implications in novel therapeutic
strategies. Cancers (Basel). 12:16682020. View Article : Google Scholar
|
|
18
|
Jiang BH, Rue E, Wang GL, Roe R and
Semenza GL: Dimerization, DNA binding, and transactivation
properties of hypoxia-inducible factor 1. J Biol Chem.
271:17771–17778. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Huang LE, Arany Z, Livingston DM and Bunn
HF: Activation of hypoxia-inducible transcription factor depends
primarily upon redox-sensitive stabilization of its alpha subunit.
J Biol Chem. 271:32253–32259. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fu C, An N, Liu J, A J, Zhang B, Liu M,
Zhang Z, Fu L, Tian X, Wang D and Dong JT: The transcription factor
ZFHX3 is crucial for the angiogenic function of hypoxia-inducible
factor 1α in liver cancer cells. J Biol Chem. 295:7060–7074. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wiener CM, Booth G and Semenza GL: In vivo
expression of mRNAs encoding hypoxia-inducible factor 1. Biochem
Biophys Res Commun. 225:485–488. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang GL, Jiang BH, Rue EA and Semenza GL:
Hypoxia-inducible factor 1 is a basic-helix-loop-helix-PAS
heterodimer regulated by cellular O2 tension. Proc Natl Acad Sci
USA. 92:5510–5514. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang GL and Semenza GL: Purification and
characterization of hypoxia-inducible factor 1. J Biol Chem.
270:1230–1237. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bao MH and Wong CC: Hypoxia, metabolic
reprogramming, and drug resistance in liver cancer. Cells.
10:17152021. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yang YM, Kim SY and Seki E: Inflammation
and liver cancer: Molecular mechanisms and therapeutic targets.
Semin Liver Dis. 39:26–42. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang J, Zhang Q, Lou Y, Fu Q, Chen Q, Wei
T, Yang J, Tang J, Wang J, Chen Y, et al: Hypoxia-inducible
factor-1α/interleukin-1β signaling enhances hepatoma
epithelial-mesenchymal transition through macrophages in a
hypoxic-inflammatory microenvironment. Hepatology. 67:1872–1889.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ye M, Fang Z, Gu H, Song R, Ye J, Li H, Wu
Z, Zhou S, Li P, Cai X, et al: Histone deacetylase 5 promotes the
migration and invasion of hepatocellular carcinoma via increasing
the transcription of hypoxia-inducible factor-1α under hypoxia
condition. Tumour Biol. 39:10104283177050342017. View Article : Google Scholar
|
|
28
|
Habas R, Kato Y and He X: Wnt/Frizzled
activation of Rho regulates vertebrate gastrulation and requires a
novel Formin homology protein Daam1. Cell. 107:843–854. 2001.
View Article : Google Scholar
|
|
29
|
Fang X, Zhang D, Zhao W, Gao L and Wang L:
Dishevelled associated activator of morphogenesis (DAAM)
facilitates invasion of hepatocellular carcinoma by upregulating
hypoxia-inducible factor 1α (HIF-1α) Expression. Med Sci Monit.
26:e9246702020. View Article : Google Scholar
|
|
30
|
Ma H, Xie L, Zhang L, Yin X, Jiang H, Xie
X, Chen R, Lu H and Ren Z: Activated hepatic stellate cells promote
epithelial-to-mesenchymal transition in hepatocellular carcinoma
through transglutaminase 2-induced pseudohypoxia. Commun Biol.
1:1682018. View Article : Google Scholar
|
|
31
|
Ientile R, Curro M, Caccamo D, Ferlazzo N,
Gangemi C and Gugliandolo A: Transglutaminase 2 is involved in the
inflammatory response through mechanisms linked to NF-kappa B/HIF-1
alpha pathways. Amino Acids. 27:1630–1631. 2015.
|
|
32
|
Kim DS, Choi YB, Han BG, Park SY, Jeon Y,
Kim DH, Ahn ER, Shin JE, Lee BI, Lee H, et al: Cancer cells promote
survival through depletion of the von Hippel-Lindau tumor
suppressor by protein crosslinking. Oncogene. 30:4780–4790. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chu Q, Gu X, Zheng Q and Zhu H: Regulatory
mechanism of HIF-1α and its role in liver diseases: A narrative
review. Ann Transl Med. 10:1092022. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Makwana V, Ryan P, Patel B, Dukie SA and
Rudrawar S: Essential role of O-GlcNAcylation in stabilization of
oncogenic factors. Biochim Biophys Acta Gen Subj. 1863:1302–1317.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wu HT, Kuo YC, Hung JJ, Huang CH, Chen WY,
Chou TY, Chen Y, Chen YJ, Chen YJ, Cheng WC, et al:
K63-polyubiquitinated HAUSP deubiquitinates HIF-1α and dictates
H3K56 acetylation promoting hypoxia-induced tumour progression. Nat
Commun. 7:136442016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wu KJ: The role of miRNA biogenesis and
DDX17 in tumorigenesis and cancer stemness. Biomed J. 43:107–114.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang Y, Chen W, Lian J, Zhang H, Yu B,
Zhang M, Wei F, Wu J, Jiang J, Jia Y, et al: The lncRNA PVT1
regulates nasopharyngeal carcinoma cell proliferation via
activating the KAT2A acetyltransferase and stabilizing HIF-1α. Cell
Death Differ. 27:695–710. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Jung JE, Kim HS, Lee CS, Shin YJ, Kim YN,
Kang GH, Kim TY, Juhnn YS, Kim SJ, Park JW, et al: STAT3 inhibits
the degradation of HIF-1alpha by pVHL-mediated ubiquitination. Exp
Mol Med. 40:479–485. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lin MC, Lin JJ, Hsu CL, Juan HF, Lou PJ
and Huang MC: GATA3 interacts with and stabilizes HIF-1α to enhance
cancer cell invasiveness. Oncogene. 36:4243–4252. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wang D, Zhang X, Lu Y, Wang X and Zhu L:
Hypoxia inducible factor 1α in hepatocellular carcinoma with
cirrhosis: Association with prognosis. Pathol Res Pract.
214:1987–1992. 2018. View Article : Google Scholar
|
|
41
|
Ding L, Chen XP and Wang HP: Expression
and clinical significance of HIF-1a protein in hepatocellular
carcinoma tissues. Zhonghua Gan Zang Bing Za Zhi. 12:656–659.
2004.(In Chinese).
|
|
42
|
Cheng W, Cheng Z, Yang Z, Xing D and Zhang
M: Upregulation of hypoxia-inducible factor 1α mRNA expression was
associated with poor prognosis in patients with hepatocellular
carcinoma. Onco Targets Ther. 12:6285–6296. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Cao S, Yang S, Wu C, Wang Y, Jiang J and
Lu Z: Protein expression of hypoxia-inducible factor-1 alpha and
hepatocellular carcinoma: A systematic review with meta-analysis.
Clin Res Hepatol Gastroenterol. 38:598–603. 2014. View Article : Google Scholar
|
|
44
|
Qian Y, Li Y, Ge Y, Song W and Fan H:
Elevated LncRNA TRERNA1 correlated with activation of HIF-1α
predicts poor prognosis in hepatocellular carcinoma. Pathol Res
Pract. 227:1536122021. View Article : Google Scholar
|
|
45
|
Ding ZN, Dong ZR, Chen ZQ, Yang YF, Yan
LJ, Li HC, Liu KX, Yao CY, Yan YC, Yang CC and Li T: Effects of
hypoxia-inducible factor-1α and hypoxia-inducible factor-2α
overexpression on hepatocellular carcinoma survival: A systematic
review with meta-analysis. J Gastroenterol Hepatol. 36:1487–1496.
2021. View Article : Google Scholar
|
|
46
|
Chitty JL, Filipe EC, Lucas MC, Herrmann
D, Cox TR and Timpson P: Recent advances in understanding the
complexities of metastasis. F1000Res 7: F1000 Faculty Rev-1169.
2018. View Article : Google Scholar
|
|
47
|
Zhang X, Li Y, Ma Y, Yang L, Wang T, Meng
X, Zong Z, Sun X, Hua X and Li H: Yes-associated protein (YAP)
binds to HIF-1α and sustains HIF-1α protein stability to promote
hepatocellular carcinoma cell glycolysis under hypoxic stress. J
Exp Clin Cancer Res. 37:2162018. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Schito L and Semenza GL: Hypoxia-inducible
factors: Master regulators of cancer progression. Trends Cancer.
2:758–770. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Chiu DK, Tse AP, Xu IM, Di Cui J, Lai RK,
Li LL, Koh HY, Tsang FH, Wei LL, Wong CM, et al: Hypoxia inducible
factor HIF-1 promotes myeloid-derived suppressor cells accumulation
through ENTPD2/CD39L1 in hepatocellular carcinoma. Nat Commun.
8:5172017. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Hinshaw DC and Shevde LA: The tumor
microenvironment innately modulates cancer progression. Cancer Res.
79:4557–4566. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Pang L, Ng KT, Liu J, Yeung WO, Zhu J,
Chiu TS, Liu H, Chen Z, Lo CM and Man K: Plasmacytoid dendritic
cells recruited by HIF-1α/eADO/ADORA1 signaling induce
immunosuppression in hepatocellular carcinoma. Cancer Lett.
522:80–92. 2021. View Article : Google Scholar
|
|
52
|
He Q, Liu M, Huang W, Chen X, Zhang B,
Zhang T, Wang Y, Liu D, Xie M, Ji X, et al: IL-1β-induced elevation
of solute carrier family 7 member 11 promotes hepatocellular
carcinoma metastasis through up-regulating programmed death ligand
1 and colony-stimulating factor 1. Hepatology. 74:3174–3193. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Song Y, Wang H, Zou XJ, Zhang YX, Guo ZQ,
Liu L, Wu DH and Zhang DY: Reciprocal regulation of HIF-1α and
Uroplakin 1A promotes glycolysis and proliferation in
Hepatocellular Carcinoma. J Cancer. 11:6737–6747. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Zhou Y, Huang Y, Hu K, Zhang Z, Yang J and
Wang Z: HIF1A activates the transcription of lncRNA RAET1K to
modulate hypoxia-induced glycolysis in hepatocellular carcinoma
cells via miR-100-5p. Cell Death Dis. 11:1762020. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
He H, Chen T, Mo H, Chen S, Liu Q and Guo
C: Hypoxia-inducible long noncoding RNA NPSR1-AS1 promotes the
proliferation and glycolysis of hepatocellular carcinoma cells by
regulating the MAPK/ERK pathway. Biochem Biophys Res Commun.
533:886–892. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Wang M, Zhao X, Zhu D, Liu T, Liang X, Liu
F, Zhang Y, Dong X and Sun B: HIF-1α promoted vasculogenic mimicry
formation in hepatocellular carcinoma through LOXL2 up-regulation
in hypoxic tumor microenvironment. J Exp Clin Cancer Res.
36:602017. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Liu Z, Wang Y, Dou C, Xu M, Sun L, Wang L,
Yao B, Li Q, Yang W, Tu K and Liu Q: Hypoxia-induced up-regulation
of VASP promotes invasiveness and metastasis of hepatocellular
carcinoma. Theranostics. 8:4649–4663. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Liu Z, Tu K, Wang Y, Yao B, Li Q, Wang L,
Dou C, Liu Q and Zheng X: Hypoxia accelerates aggressiveness of
hepatocellular carcinoma cells involving oxidative stress,
epithelial-mesenchymal transition and non-canonical hedgehog
signaling. Cell Physiol Biochem. 44:1856–1868. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Zhang L, Huang G, Li X, Zhang Y, Jiang Y,
Shen J, Liu J, Wang Q, Zhu J, Feng X, et al: Hypoxia induces
epithelial-mesenchymal transition via activation of SNAI1 by
hypoxia-inducible factor-1α in hepatocellular carcinoma. BMC
Cancer. 13:1082013. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Yu Y, Min Z, Zhou Zhihang, Linhong M, Tao
R, Yan L and Song H: Hypoxia-induced exosomes promote
hepatocellular carcinoma proliferation and metastasis via miR-1273f
transfer. Exp Cell Res. 385:1116492019. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Cui CP, Wong CC, Kai AK, Ho DW, Lau EY,
Tsui YM, Chan LK, Cheung TT, Chok KS, Chan ACY, et al: SENP1
promotes hypoxia-induced cancer stemness by HIF-1α deSUMOylation
and SENP1/HIF-1α positive feedback loop. Gut. 66:2149–2159. 2017.
View Article : Google Scholar
|
|
62
|
Rankin EB and Giaccia AJ: Hypoxic control
of metastasis. Science. 352:175–180. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Liu Y and Cao X: Immunosuppressive cells
in tumor immune escape and metastasis. J Mol Med (Berl).
94:509–522. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Jing X, Yang F, Shao C, Wei K, Xie M, Shen
H and Shu Y: Role of hypoxia in cancer therapy by regulating the
tumor microenvironment. Mol Cancer. 18:1572019. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Kiss M, Vande Walle L, Saavedra PHV,
Lebegge E, Van Damme H, Murgaski A, Qian J, Ehling M, Pretto S,
Bolli E, et al: IL1β promotes immune suppression in the tumor
microenvironment independent of the inflammasome and gasdermin D.
Cancer Immunol Res. 9:309–323. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Feng J, Li J, Wu L, Yu Q, Ji J, Wu J, Dai
W and Guo C: Emerging roles and the regulation of aerobic
glycolysis in hepatocellular carcinoma. J Exp Clin Cancer Res.
39:1262020. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Yuan K, Xie K, Lan T, Xu L, Chen X, Li X,
Liao M, Li J, Huang J, Zeng Y and Wu H: TXNDC12 promotes EMT and
metastasis of hepatocellular carcinoma cells via activation of
β-catenin. Cell Death Differ. 27:1355–1368. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Venhuizen JH, Jacobs FJC, Span PN and
Zegers MM: P120 and E-cadherin: Double-edged swords in tumor
metastasis. Semin Cancer Biol. 60:107–120. 2020. View Article : Google Scholar
|
|
69
|
Na TY, Schecterson L, Mendonsa AM and
Gumbiner BM: The functional activity of E-cadherin controls tumor
cell metastasis at multiple steps. Proc Natl Acad Sci USA.
117:5931–5937. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Evans AJ, Russell RC, Roche O, Burry TN,
Fish JE, Chow VW, Kim WY, Saravanan A, Maynard MA, Gervais ML, et
al: VHL promotes E2 box-dependent E-cadherin transcription by
HIF-mediated regulation of SIP1 and snail. Mol Cell Biol.
27:157–169. 2007. View Article : Google Scholar
|
|
71
|
Wu Y, Zhang J, Zhang X, Zhou H, Liu G and
Li Q: Cancer stem cells: A potential breakthrough in HCC-targeted
therapy. Front Pharmacol. 11:1982020. View Article : Google Scholar
|
|
72
|
Liu YC, Yeh CT and Lin KH: Cancer stem
cell functions in hepatocellular carcinoma and comprehensive
therapeutic strategies. Cells. 9:13312020. View Article : Google Scholar
|
|
73
|
Lambert AW, Pattabiraman DR and Weinberg
RA: Emerging biological principles of metastasis. Cell.
168:670–691. 2017. View Article : Google Scholar
|
|
74
|
Wang N, Wang S, Li MY, Hu BG, Liu LP, Yang
SL, Yang S, Gong Z, Lai PBS and Chen GG: Cancer stem cells in
hepatocellular carcinoma: An overview and promising therapeutic
strategies. Ther Adv Med Oncol. 10:17588359188162872018. View Article : Google Scholar
|
|
75
|
Lee TK, Guan XY and Ma S: Cancer stem
cells in hepatocellular carcinoma-from origin to clinical
implications. Nat Rev Gastroenterol Hepatol. 19:26–44. 2022.
View Article : Google Scholar
|
|
76
|
Du B and Shim JS: Targeting
epithelial-mesenchymal transition (EMT) to overcome drug resistance
in cancer. Molecules. 21:9652016. View Article : Google Scholar
|
|
77
|
Wang R, Sun Q, Wang P, Liu M, Xiong S, Luo
J, Huang H, Du Q, Geller DA and Cheng B: Notch and Wnt/β-catenin
signaling pathway play important roles in activating liver cancer
stem cells. Oncotarget. 7:5754–5768. 2016. View Article : Google Scholar
|
|
78
|
Liu LP, Ho RL, Chen GG and Lai PB:
Sorafenib inhibits hypoxia-inducible factor-1α synthesis:
Implications for antiangiogenic activity in hepatocellular
carcinoma. Clin Cancer Res. 18:5662–5671. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Méndez-Blanco C, Fondevila F,
García-Palomo A, González-Gallego J and Mauriz JL: Sorafenib
resistance in hepatocarcinoma: Role of hypoxia-inducible factors.
Exp Mol Med. 50:1–9. 2018. View Article : Google Scholar
|
|
80
|
Liang Y, Zheng T, Song R, Wang J, Yin D,
Wang L, Liu H, Tian L, Fang X, Meng X, et al: Hypoxia-mediated
sorafenib resistance can be overcome by EF24 through Von
Hippel-Lindau tumor suppressor-dependent HIF-1α inhibition in
hepatocellular carcinoma. Hepatology. 57:1847–1857. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Wu FQ, Fang T, Yu LX, Lv GS, Lv HW, Liang
D, Li T, Wang CZ, Tan YX, Ding J, et al: ADRB2 signaling promotes
HCC progression and sorafenib resistance by inhibiting autophagic
degradation of HIF1α. J Hepatol. 65:314–324. 2016. View Article : Google Scholar
|
|
82
|
Li S, Li J, Dai W, Zhang Q, Feng J, Wu L,
Liu T, Yu Q, Xu S, Wang W, et al: Genistein suppresses aerobic
glycolysis and induces hepatocellular carcinoma cell death. Br J
Cancer. 117:1518–1528. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Feng J, Dai W, Mao Y, Wu L, Li J, Chen K,
Yu Q, Kong R, Li S, Zhang J, et al: Simvastatin re-sensitizes
hepatocellular carcinoma cells to sorafenib by inhibiting
HIF-1α/PPAR-γ/PKM2-mediated glycolysis. J Exp Clin Cancer Res.
39:242020. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Song DS, Nam SW, Bae SH, Kim JD, Jang JW,
Song MJ, Lee SW, Kim HY, Lee YJ, Chun HJ, et al: Outcome of
transarterial chemoembolization-based multi-modal treatment in
patients with unresectable hepatocellular carcinoma. World J
Gastroenterol. 21:2395–2404. 2015. View Article : Google Scholar
|
|
85
|
Liu K, Min XL, Peng J, Yang K, Yang L and
Zhang XM: The changes of HIF-1α and VEGF expression After TACE in
patients with hepatocellular carcinoma. J Clin Med Res. 8:297–302.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Huang M, Wang L, Chen J, Bai M, Zhou C,
Liu S and Lin Q: Regulation of COX-2 expression and
epithelial-to-mesenchymal transition by hypoxia-inducible factor-1α
is associated with poor prognosis in hepatocellular carcinoma
patients post TACE surgery. Int J Oncol. 48:2144–2154. 2016.
View Article : Google Scholar
|
|
87
|
Ribas A and Wolchok JD: Cancer
immunotherapy using checkpoint blockade. Science. 359:1350–1355.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Sangro B, Sarobe P, Hervás-Stubbs S and
Melero I: Advances in immunotherapy for hepatocellular carcinoma.
Nat Rev Gastroenterol Hepatol. 18:525–543. 2021. View Article : Google Scholar
|
|
89
|
Ruiz de Galarreta M, Bresnahan E,
Molina-Sánchez P, Lindblad KE, Maier B, Sia D, Puigvehi M, Miguela
V, Casanova-Acebes M, Dhainaut M, et al: β-catenin activation
promotes immune escape and resistance to Anti-PD-1 therapy in
hepatocellular carcinoma. Cancer Discov. 9:1124–1141. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Kalantari Khandani N, Ghahremanloo A and
Hashemy SI: Role of tumor microenvironment in the regulation of
PD-L1: A novel role in resistance to cancer immunotherapy. J Cell
Physiol. 235:6496–6506. 2020. View Article : Google Scholar
|
|
91
|
Deng Z, Teng YJ, Zhou Q, Ouyang ZG, Hu YX,
Long HP, Hu MJ, Mei S, Lin FX, Dai XJ, et al: Shuyu pills inhibit
immune escape and enhance chemosensitization in hepatocellular
carcinoma. World J Gastrointest Oncol. 13:1725–1740. 2021.
View Article : Google Scholar
|
|
92
|
Chen P, Duan X, Li X, Li J, Ba Q and Wang
H: HIPK2 suppresses tumor growth and progression of hepatocellular
carcinoma through promoting the degradation of HIF-1α. Oncogene.
39:2863–2876. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Xu S, Ling S, Shan Q, Ye Q, Zhan Q, Jiang
G, Zhuo J, Pan B, Wen X, Feng T, et al: Self-activated
cascade-responsive sorafenib and USP22 shRNA Co-delivery system for
synergetic hepatocellular carcinoma therapy. Adv Sci (Weinh).
8:20030422021. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Ling S, Shan Q, Zhan Q, Ye Q, Liu P, Xu S,
He X, Ma J, Xiang J, Jiang G, et al: USP22 promotes hypoxia-induced
hepatocellular carcinoma stemness by a HIF1α/USP22 positive
feedback loop upon TP53 inactivation. Gut. 69:1322–1334. 2020.
View Article : Google Scholar
|
|
95
|
Korbecki J, Simińska D,
Gąssowska-Dobrowolska M, Listos J, Gutowska I, Chlubek D and
Baranowska-Bosiacka I: Chronic and cycling hypoxia: Drivers of
cancer chronic inflammation through HIF-1 and NF-κB Activation: A
review of the molecular mechanisms. Int J Mol Sci. 22:107012021.
View Article : Google Scholar
|
|
96
|
Jiang Y, Zhu Y, Wang X, Gong J, Hu C, Guo
B, Zhu B and Li Y: Temporal regulation of HIF-1 and NF-κB in
hypoxic hepatocarcinoma cells. Oncotarget. 6:9409–9419. 2015.
View Article : Google Scholar
|
|
97
|
Hu L, Zeng Z, Xia Q, Liu Z, Feng X, Chen
J, Huang M, Chen L, Fang Z, Liu Q, et al: Metformin attenuates
hepatoma cell proliferation by decreasing glycolytic flux through
the HIF-1α/PFKFB3/PFK1 pathway. Life Sci. 239:1169662019.
View Article : Google Scholar
|
|
98
|
Su Q, Fan M, Wang J, Ullah A, Ghauri MA,
Dai B, Zhan Y, Zhang D and Zhang Y: Sanguinarine inhibits
epithelial-mesenchymal transition via targeting HIF-1α/TGF-β
feed-forward loop in hepatocellular carcinoma. Cell Death Dis.
10:9392019. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Chen M, Wu J, Shi S, Chen Y, Wang H, Fan H
and Wang S: Structure analysis of a heteropolysaccharide from
Taraxacum mongolicum Hand.-Mazz. and anticomplementary activity of
its sulfated derivatives. Carbohydr Polym. 152:241–252. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Cai L, Wan D, Yi F and Luan L:
Purification, preliminary characterization and hepatoprotective
effects of polysaccharides from dandelion root. Molecules.
22:14092017. View Article : Google Scholar
|
|
101
|
Ren F, Wu K, Yang Y, Yang Y, Wang Y and Li
J: Dandelion polysaccharide exerts anti-angiogenesis effect on
hepatocellular carcinoma by regulating VEGF/HIF-1α expression.
Front Pharmacol. 11:4602020. View Article : Google Scholar
|
|
102
|
Shao C, Yang F, Miao S, Liu W, Wang C, Shu
Y and Shen H: Role of hypoxia-induced exosomes in tumor biology.
Mol Cancer. 17:1202018. View Article : Google Scholar : PubMed/NCBI
|