Introduction
Esophageal cancer is the eighth most common
malignant neoplasm and the sixth leading cause of cancer-related
deaths worldwide (1). Despite
advances in the treatment of esophageal cancer over recent decades,
patient prognosis has shown little improvement (2). Even with the introduction of
combination treatment regimens and early diagnostic technology, the
survival rate remains unsatisfactory, with a 5-year overall
survival rate of 17–20% in Asia (3). Lymph node metastasis is one of the
most important prognostic factors of esophageal cancer and
generally indicates a poor outcome (4). Therefore, the identification of new
prognostic biomarkers and the development of effective treatment
methods are imperative for improving the clinical outcomes of
patients with esophageal squamous cell carcinoma (ESCC).
Exosomes are a class of extracellular vesicles; that
is, lipid bilayer-enclosed carriers of proteins, nucleic acids,
lipids and metabolites, which are secreted by cells into the
extracellular environment (5). As
such, exosomes can transfer bioactive molecules from donor to
recipient cells and influence their biological activities (6). Tumor-derived exosomes are released
into peripheral blood and their quantity is a prognostic marker of
ESCC that alters cellular gene expression and contributes to tumor
progression (7). Furthermore,
tumor-derived exosomes have been shown to alter cellular gene
expression and contribute to tumor progression in ESCC (8). Therefore, a comprehensive
understanding of the mechanisms through which circulating exosomal
microRNAs (miRNAs/miRs) influence cancer progression is
warranted.
Emerging evidence has suggested that hypoxia
contributes to ESCC resistance against first-line chemotherapy,
such as cisplatin and 5-fluorouracil (9). Our previous study reported that
hypoxia-inducible factor (HIF)-1α serves an essential role in
hypoxia-induced ESCC progression by maintaining crucial mechanisms,
such as epithelial-mesenchymal transition, proliferation,
migration/invasion, apoptosis, cell cycle progression and
chemoresistance, influencing patient prognosis (10). However, the mechanisms through which
hypoxia influences miRNAs in ESCC remain poorly understood.
miRNAs are small noncoding RNAs that regulate gene
expression at the post-transcriptional level (11). A growing body of evidence has
demonstrated that miRNAs are implicated in the initiation and
progression of ESCC by regulating the expression of oncogenes and
tumor suppressors (12,13). The prognostic applications of miRNAs
in ESCC have recently attracted interest. miR-185 has been
identified as a tumor-suppressive miRNA in multiple types of
cancer, including hepatocellular carcinoma (14), osteosarcoma (15) and prostate cancer (16), in addition to being associated with
the prognosis of colon (17) and
gastric cancer (18). However,
there is limited literature on the role of exosomal miR-185 in
ESCC. The biological roles of miR-185 in cell viability, Ki-67
staining, cell migration, invasion, xenograft model, locoregional
staging and molecular mechanisms of ESCC, and the survival of
patients with ESCC have been reported (19–21).
The present study observed alterations in exosomal miR-185
expression under hypoxic conditions, as well as its association
with lymph node metastasis before chemotherapy and the sensitivity
of ESCC cells to chemotherapy. Moreover, this study investigated
the association of exosomal miR-185 levels with clinicopathological
factors and prognosis in ESCC, and explored the mechanism
underlying the role of exosomal miR-185 via in vitro
experiments using ESCC cell lines and bioinformatics analyses. The
findings highlight the importance of exosomal miR-185 as a
prognostic biomarker and therapeutic target in ESCC.
Materials and methods
Clinical samples
Plasma samples were collected from 89 patients
diagnosed with ESCC at Chiba University Hospital (Chiba, Japan;
affiliated with Chiba University Graduate School of Medicine)
between May 2011 and April 2017. Patients aged 20–85 years (median,
67 years) with histologically diagnosed ESCC were included, whereas
patients with other types of cancer were excluded; however, no
other exclusion criteria, such as history of other medical
diseases, were applied. All patients were staged according to the
Japanese Classification of Esophageal Cancer 11th edition (22). Written informed consent was obtained
from all participants. The present study was approved by the Ethics
Committee of the Chiba University Graduate School of Medicine
(approval no. 1264; Chiba, Japan). Blood examination and sampling
were performed before treatment. After receiving patient blood
samples, they were centrifuged to obtain plasma at 4°C and
1,690 × g for 10 min, and were stored at −80°C.
Cell lines and cell culture
The KYSE-960 and KYSE-410 human ESCC cell lines were
purchased from the Japanese Collection of Research Bioresources
Cell Bank, and the T.Tn, TE1, TE6, TE11 and TE14 ESCC cell lines
were provided by the Cell Resource Center at Tohoku University
(Sendai, Japan). The SCCVII mouse squamous cell carcinoma cell line
was kindly provided by Professor Yuta Shibamoto (Department of
Quantum Radiology, Nagoya City University, Nagoya, Japan).
Immortalized esophageal keratinocyte cells (R2C3), which were
established at Chiba University School of Medicine, were used as a
control cell line (23). The cells
were cultured in Dulbecco's modified Eagle's medium (DMEM; Thermo
Fisher Scientific, Inc.) supplemented with 10% fetal bovine serum
(FBS; cat. no. 10270-106; Gibco; Thermo Fisher Scientific, Inc.)
and 100 U/ml penicillin, and were maintained at 37°C in a
humidified atmosphere containing 5% CO2. For exosome
isolation under normoxic conditions, cells were incubated in DMEM
replenished with 10% exosome-free FBS and 1%
penicillin/streptomycin at 37°C in 21% O2 and 5%
CO2 for 48 h. To simulate physical hypoxia, cells were
incubated in 1% O2 and 5% CO2 in a multi-gas
incubator for 48 h (cat. no. BL-43MD; TOSC Japan Ltd.).
Exosome isolation from the cell
culture medium
A total of 5×106 cells were incubated in
medium with 10% exosome-free FBS for 48 h at 37°C and total (10 ml)
cell culture medium was harvested. Exosomes were isolated using the
total exosome isolation kit (from cell culture media) (cat. no.
4478359; Invitrogen; Thermo Fisher Scientific, Inc.), according to
manufacturer's protocol. The same method was performed that was
described in our previous study (7).
Profiling of exosomal miRNAs extracted
from normoxia/hypoxia culture medium
Microarray analysis was performed on total RNA
extracted from exosomes isolated by ultracentrifugation. Prior to
exosome isolation, KYSE-960 and T.Tn cells were cultured to 80%
confluence, and were cultured in normoxia or hypoxia, as
aforementioned. Exosomes were isolated from the cell culture media
by ultracentrifugation at 10,000 × g for 90 min at 4°C (Optima TLX
Ultracentrifuge; Beckman Coulter, Inc.), as previously reported
(8). Exosome extraction was
confirmed by transmission electron microscopy (TEM) and
nanoparticle tracking analysis (Fig.
S1). Nanoparticle tracking analysis was performed using
NanoSight NS300 and NTA2.3 software (Fujifilm), according to the
manufacturer's protocol, to define the particle size distribution
and exosome concentration. Total Exosome RNA & Protein
Isolation Kit (cat. no. 4478545; Thermo Fisher Scientific, Inc.)
was used to extract total RNA from exosomes. Exosomal RNA from
normoxic and hypoxic KYSE-960 and T.Tn cell culture media were
analyzed using Affymetrix GeneChip miRNA 4.0 (cat. no. 902412;
Affymetrix; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. Arrays were incubated in the GeneChip™
Hybridization Oven 645 (Thermo Fisher Scientific, Inc.) at 48°C for
18 h (agitation at 60 rpm) and scanned using the GeneChip Scanner
3000 7G (Thermo Fisher Scientific, Inc.) according to the
accompanying manual [GeneChip Command Console (AGCC) 4.0 User
Manual]. miRNA expression was calculated using the detection above
background algorithm and was normalized by robust multichip
analysis. Chip data analysis was performed using R software (4.2.0)
(https://www.r-project.org).
Exosome isolation from plasma
Each plasma sample (1–1.5 ml) was centrifuged at
2,000 × g for 20 min at room temperature to remove cells and
debris. Exosomes were then isolated using the total exosome
isolation kit (from plasma) (cat. no. 4484450; Invitrogen; Thermo
Fisher Scientific, Inc.), according to the manufacturer's protocol.
The same method was performed that was described in our previous
study (7).
TEM
TEM observation was performed using a carbon-coated
copper grid (Excel support film; 200 mesh; cat. no. RL26A; Nisshin
Em Co., Ltd.) and the negative staining method. Sample preparation
was performed according to the method described in a previous
report (24). The samples were then
subjected to TEM observation (H-7650; Hitachi High-Technologies
Corporation) at an acceleration voltage of 80.0 kV.
miRNA and mRNA isolation and detection
via reverse transcription-quantitative PCR (RT-qPCR)
Total RNA was extracted from the exosomes using the
Total Exosome RNA and Protein Isolation Kit (Invitrogen; Thermo
Fisher Scientific, Inc.), according to the manufacturer's protocol.
Total cellular RNA was extracted using the Mini Kit (Qiagen GmbH),
according to the manufacturer's protocol. Total RNA was then
reverse-transcribed to cDNA using a High-Capacity RNA-to-DNA™ Kit
(Thermo Fisher Scientific, Inc.), according to the manufacturer's
protocol. qPCR analysis was performed using TaqMan MicroRNA Assays
(Invitrogen; Thermo Fisher Scientific, Inc.) or the SsoFast™
EvaGreen Supermix (Bio-Rad Laboratories, Inc.). The TaqMan primer
for hsa-miR-185 (assay ID 002271) was used to detect miR-185
expression; miR-16 (assay ID 000391), which was previously used as
a control for cell-free miRNA analysis in our laboratory (25), was used as an internal control for
detection of exosomal miR-185 from plasma and cancer cell culture
medium, whereas U6 small nuclear RNA (assay ID 001973) was used as
an internal control for detection of miR-185 expression in cancer
cell lines (all from Applied Biosystems; Thermo Fisher Scientific,
Inc.). BCL-2 expression was normalized to β-actin. The
thermocycling conditions were as follows: For miRNA detection,
samples were incubated at 95°C for 10 min, followed by 40 cycles at
95°C for 15 sec and 60°C for 60 sec; for mRNA detection, samples
were incubated for 30 sec at 95°C, followed by 40 cycles at 95°C
for 5 sec and 60°C for 10 sec. Relative expression was calculated
using the 2−ΔΔCq method (26); BCL-2 and miR-185 specific primers
are detailed in Table I.
 | Table I.Primer sequences used for reverse
transcription- quantitative PCR. |
Table I.
Primer sequences used for reverse
transcription- quantitative PCR.
| Name | Sequence,
5′-3′ |
|---|
| BCL-2 | F:
GGATCCAGGATAACGGAGGC |
| BCL-2 | R:
GGCAGGCATGTTGACTTCAC |
| β-actin | F:
CATGTACGTTGCTATCCAGGC |
| β-actin | R:
CTCCTTAATGTCACGCACGAT |
| hsa-mir-185
stem-loop |
AGGGGGCGAGGGAUUGGAGA |
|
|
GAAAGGCAGUUCCUGAUGGUC |
|
|
CCCUCCCCAGGGGCUGGCUUU |
|
|
CCUCUGGUCCUUCCCUCCCA |
| hsa-mir-16-1
stem-loop |
GUCAGCAGUGCCUUAGCAGCA |
|
|
CGUAAAUAUUGGCGUUAAGAU |
|
|
UCUAAAAUUAUCUCCAGUAUU |
|
|
AACUGUGCUGCUGAAGUAAGG |
|
| UUGAC |
| U6 snRNA
stem-loop |
GTGCTCGCTTCGGCAGCACATA |
|
|
TACTAAAATTGGAACGATACAG |
|
|
AGAAGATTAGCATGGCCCCTGC |
|
|
GCAAGGATGACACGCAAATTC |
|
|
GTGAAGCGTTCCATATTTT |
miRNA transfection
KYSE-960 and T.Tn cells were seeded in 6-well
plates (2.5×105 cells/well) and, after 24 h, the cells
were transfected with miR-185 mimic or negative control (25
pmol/well) using Lipofectamine™ RNAiMAX Transfection Reagent
(Invitrogen; Thermo Fisher Scientific, Inc.). The miR-185 mimic and
negative control were purchased from Thermo Fisher Scientific, Inc.
(mirVana miRNA mimic and negative control; assay ID MC12486). The
miR-185 mimic and negative control were transfected into cells for
48 h at 37°C. After 48 h of transfection, the cells were harvested
for the subsequent experiments.
Migration, invasion, cell
proliferation and colony formation assays
Migration and invasion were detected using Transwell
assays. In a 24-well plate, 5×104 transfected KYSE-960
and T.Tn cells/well were seeded in the upper chamber (8-µm pore;
cat. no. 354480; Corning BioCoat Matrigel Invasion Chamber;
Corning, Inc.) in FBS-free medium. A medium containing 10% FBS was
added to the lower chamber. After incubation for 48 h at 37°C,
non-invading cells were removed from the upper chamber with a
cotton swab, whereas cells on the lower surface were fixed with 99%
methanol for 5 sec at room temperature and stained using Diff-Quick
Staining (Sysmex Corporation) at room temperature overnight. Images
of three random fields from triplicate wells were recorded.
Migration assays were performed in the same manner, except that the
chambers had no Matrigel coating (8-µm pore; cat. no. 662638;
Greiner Bio-One International GmbH), and the incubation time was 24
h at 37°C.
To assess cell proliferation, a total of
5×103 KYSE-960 and T.Tn transfected cells/well were
seeded in a 96-well plate. Cell proliferation was assessed using a
Cell Counting Kit-8 (CCK-8) assay (Dojindo Laboratories, Inc.). The
reagent was added into each well and incubated for a further 2 h at
37°C every 24 h and the absorbance at 450 nm was measured using a
microplate reader (Bio-Rad Laboratories, Inc.). The data were
statistically analyzed on day 4 using Student's t-test.
For the colony formation assay, 800 cells/well of
KYSE-960 and T.Tn cells were plated in a 6-well plate and
were maintained in complete culture medium. After 2 weeks, the
colonies were stained with a Diff-Quick Stain (Sysmex Corporation)
for 15 sec at room temperature. Images of the visible colonies were
captured and the number of colonies consisting of >50 cells was
counted.
Apoptosis and cell cycle analyses
The IC50 values of cisplatin in KYSE-960
and T.Tn cells were determined using CCK-8 assay. KYSE-960 and T.Tn
cells were seeded in 96-well plates (5×103 cells/well)
and were incubated at 37°C in a humidified 5% CO2
atmosphere for 24 h. Subsequently, the medium was replaced with
fresh medium with or without various concentrations (0–100 µM) of
cisplatin (cat. no. P4394; MilliporeSigma). A total of 48 h after
cisplatin administration, cell viability was measured using the
CCK-8 assay, as aforementioned, and IC50 was calculated.
Subsequently, KYSE-960 and T.Tn cell lines were treated with the
IC50 of cisplatin for 48 h at 37°C. The cells were then
harvested and washed with PBS, resuspended in 100 µl Annexin V
Binding Solution, and incubated with 5 µl Annexin V FITC and 5 µl
PI solution (Annexin V-FITC Apoptosis Detection Kit; Nacalai
Tesque, Inc.) at room temperature for 15 min. Finally, 400 µl
Annexin V Binding Solution was added prior to analysis (BD
FACSCanto™ II Flow Cytometer; BD Biosciences). The results were
analyzed using FlowJo software (FlowJo, 10.8.1; FlowJo, LLC)
For cell cycle analysis, cells were harvested after
48 h of transfection, washed with PBS, harvested and fixed with
pre-cooled 70% ethanol at 4°C overnight (18–24 h). After
washing, the cells were incubated with 100 µg/ml RNase A
(Invitrogen; Thermo Fisher Scientific, Inc.) and 0.1% Triton-100 at
37°C for 5 min at 37°C, and were then stained with 50
µg/ml PI at room temperature for 30 min in the dark. The DNA
content was measured on a BD FACSCanto II Flow Cytometer using the
MOdfitLT 5.0 software program.
Gene Set Enrichment Analysis
(GSEA)
Enrichment analysis of The Cancer Genome Atlas
Esophageal Carcinoma Collection (TCGA-ESCA) data (https://gdac.broadinstitute.org/runs/stddata__2016_01_28/data/ESCA/20160128/)
was performed using GSEA v4.0.1 (https://www.gsea-msigdb.org/gsea/index.jsp). RNA-seq
and miRNA-seq data from a total of 195 patients with esophageal
cancer were used to evaluate miRNA expression. Patients were
divided into high and low miR-185-5p expression groups based on
median miR-185-5p expression.
Statistical analysis
Statistical analysis was performed using SPSS 21
(IBM Corp.) and GraphPad Prism 7.04 (Dotmatics). Data are presented
as the mean ± standard deviation. An unpaired Student's t-test and
one-way analysis of variance with Tukey's post hoc test were
employed for quantitative variables, and χ2 test
or Fisher's exact test were employed for qualitative variables to
compare the characteristics of each group. The association between
exosomal miR-185 expression in cell lines and in cancer cell
culture media was determined using Pearson correlation analysis.
Progression-free survival (PFS) and disease specific survival (DSS)
curves were plotted using the Kaplan-Meier method and results were
compared using the log-rank test. PFS was defined as the time-frame
between the start of treatment and the date of the first
progression, and was used as an indicator of treatment efficacy.
DSS was defined as the time-frame between the start of treatment
and the date of death due to ESCC, and was used as a prognostic
indicator. Each experiment was repeated three times, except for miR
microarray analysis. P<0.05 was considered to indicate a
statistically significant difference.
Results
miRNA array expression analysis of
KYSE-960 and T.Tn cell culture media
miRNA array analysis was performed to screen
differentially expressed exosomal miRNAs in KYSE-960 and T.Tn cells
culture medium under normoxic and hypoxic conditions. The
expression of 33 exosomal miRNAs exhibited 2-fold downregulation,
whereas 12 miRNAs exhibited 2-fold upregulation in normoxic versus
hypoxic KYSE-960 cells. In the T.Tn cell line, 33 exosomal miRNAs
exhibited 2-fold downregulation, and nine exhibited 2-fold
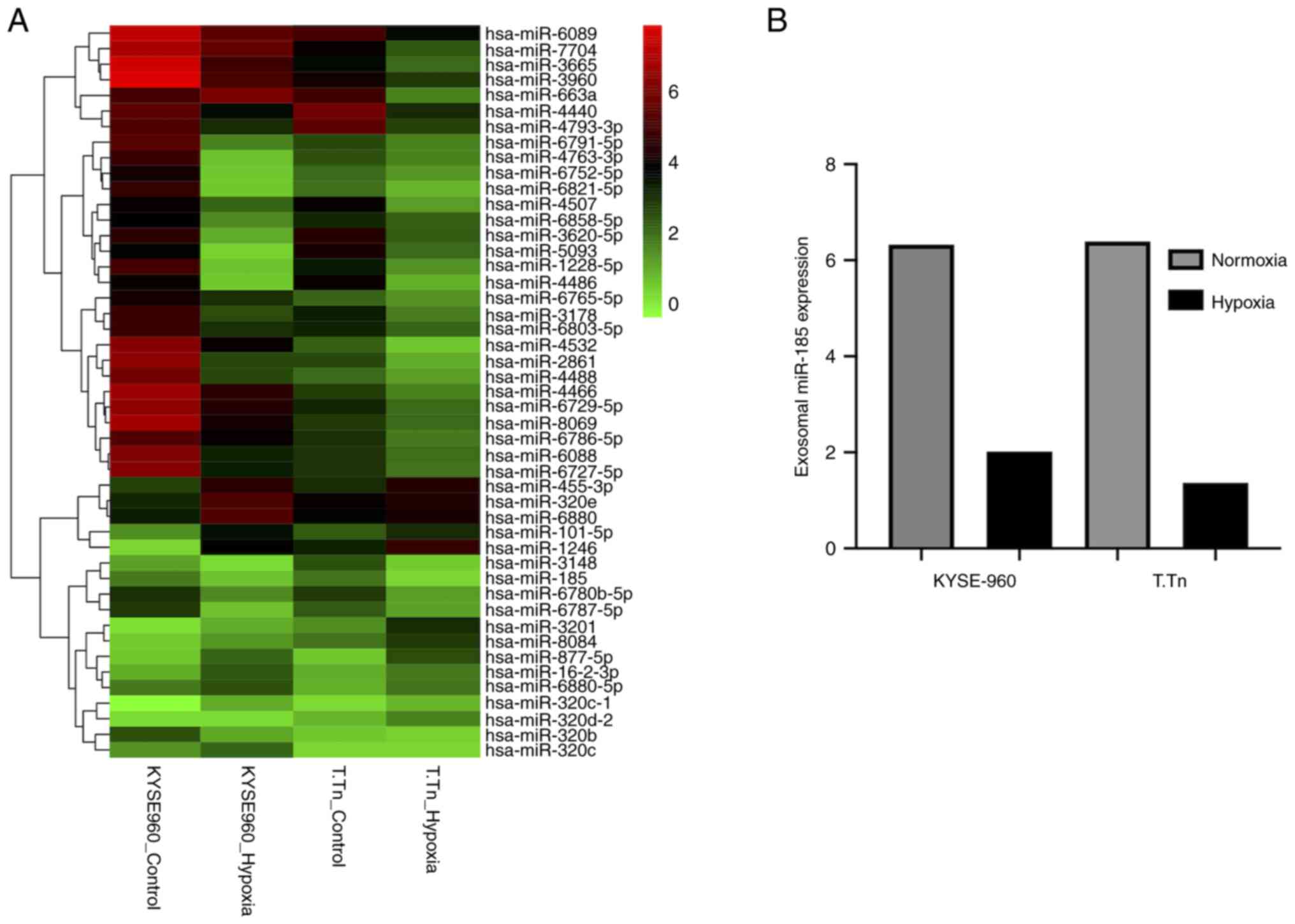
upregulation. Cluster analysis of the intersecting miRNAs is shown
in Fig. 1A. The expression levels
of exosomal miR-185 were decreased in both KYSE-960 and T.Tn cell
culture media under hypoxic conditions (fold change relative to
that in KYSE-960 cells: 0.32, fold change relative to that in T.Tn
cells: 0.22; Fig. 1B). Different
cells respond differently to external stimuli, which may be why
inconsistent fold changes occur in the expression levels of miRNAs
in KYES-960 and T.Tn cell lines.
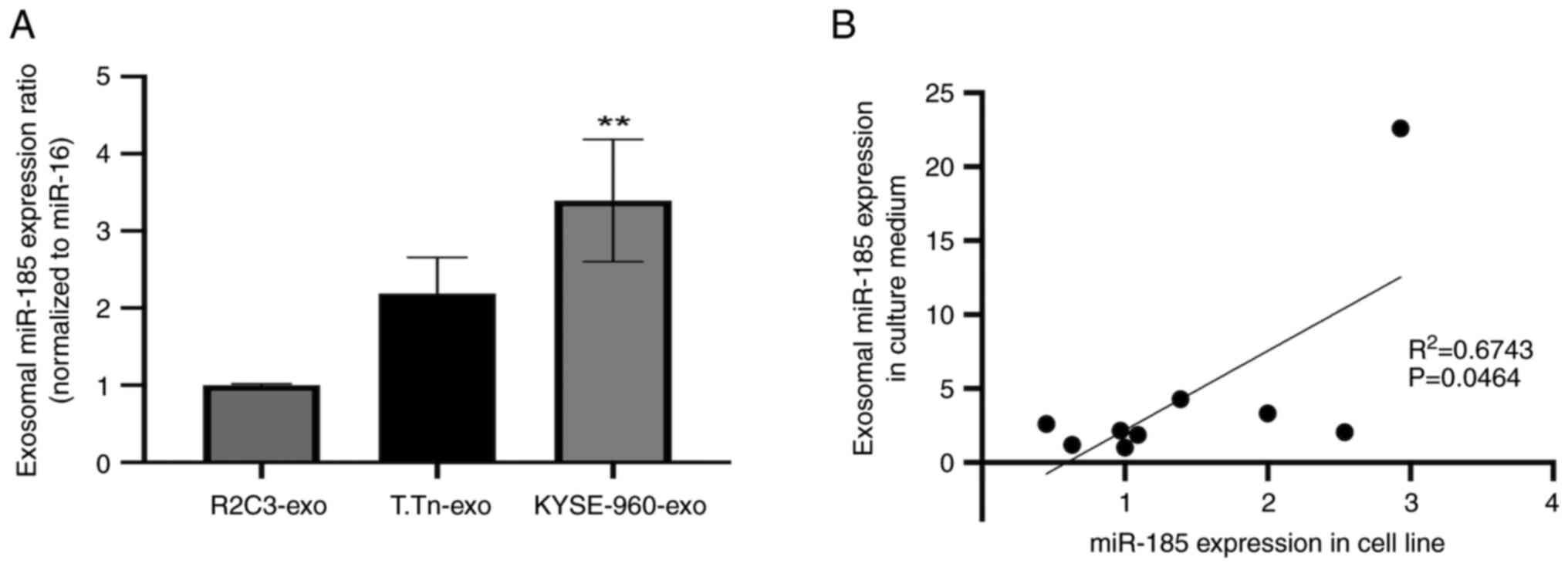
Analysis of miR-185-5p expression in
ESCC cells and exosomes
miR-185-5p expression in T.Tn cells was similar to
that detected in normal esophageal keratinocytes (R2C3), whereas
the expression levels of miR-185-5p were slightly higher in
KYSE-960 cells than those in normal esophageal keratinocytes
(R2C3), but not statistically significant (Fig. S2). miR-185-5p expression in
exosomes derived from T.Tn cell culture medium was two times higher
than that in exosomes from normal esophageal keratinocytes, but the
difference was not statistically significant. Furthermore,
miR-185-5p expression in exosomes derived from KYSE-960 culture
medium was significantly higher than that in exosomes from normal
esophageal keratinocytes (Fig. 2A).
Correlation analysis of miR-185 expression in R2C3, T.Tn, TE1, TE6,
TE11, TE14, KYSE-960, KYSE-410 and SCCVII cells, and its expression
in exosomes was conducted using data obtained from RT-qPCR
(R2=0.6743, P=0.0464; Fig.
2B). A significant positive correlation between miR-185
expression in ESCC cell lines and miR-185 expression in culture
medium-derived exosomes was observed.
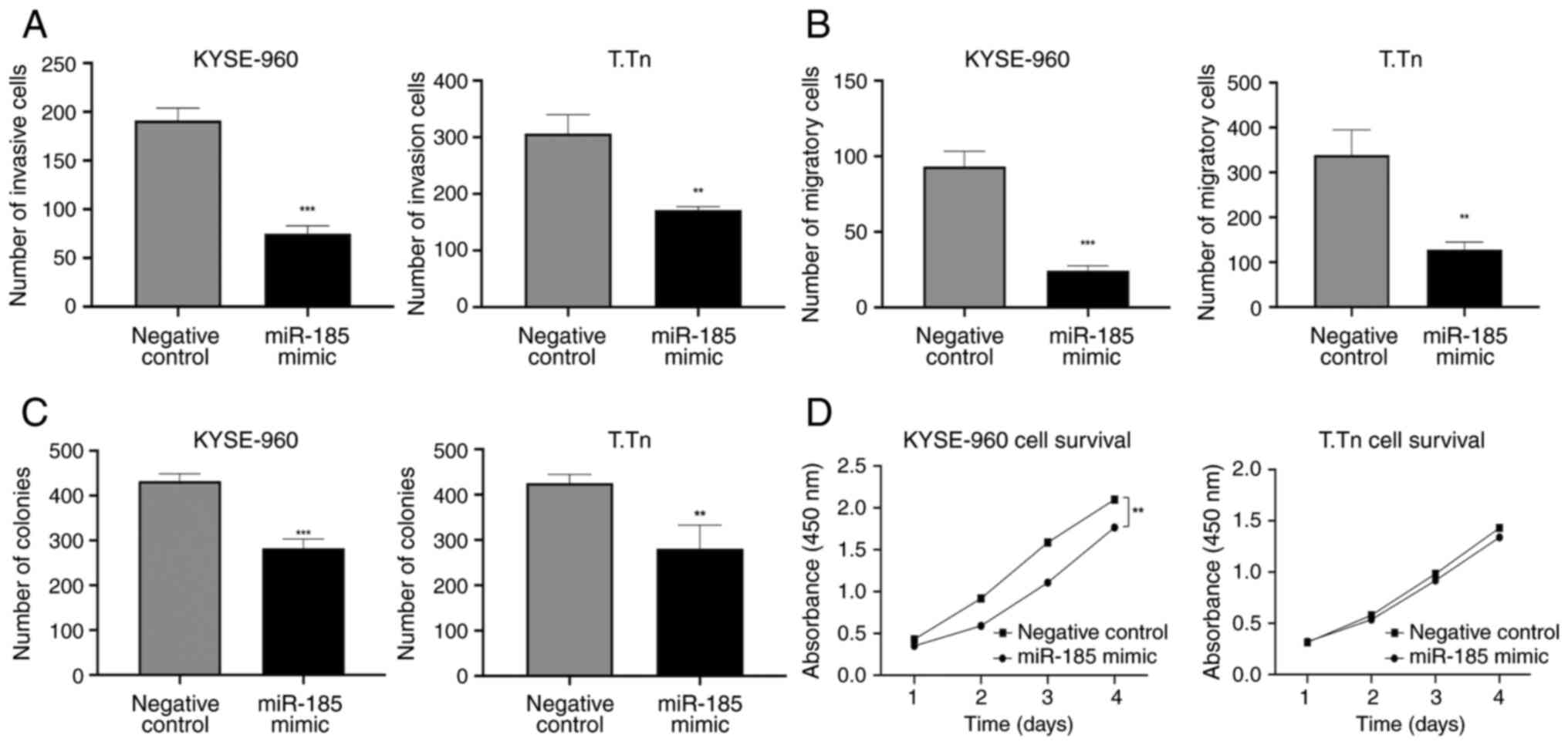
miR-185 overexpression suppresses the
invasion, migration and colony formation of ESCC cells in
vitro
The statistically significant overexpression of
miR-185 in KYSE-960 and T.Tn cells was confirmed by RT-qPCR
following miR-185 mimic transfection (>1,000-fold increase in
KYSE-960 and T.Tn cell lines, compared with in cells transfected
with the negative control mimic; Fig.
S3). The invasion and migration of miR-185-overexpressing cells
was significantly decreased compared with those in the negative
control cells [invasion: KYSE-960, P=0.0001; T.Tn, P=0.0022
(Figs. 3A and S3); migration: KYSE-960, P=0.0003; T.Tn,
P=0.0034 (Figs. 3B and S3)]. Cells transfected with the miR-185
mimic exhibited a lower survival rate than those in the negative
control group, as determined by the colony formation assay
[KYSE-960, P=0.0005; T.Tn, P=0.01 (Figs. 3C and S4)]. The results of the CCK-8 assay
indicated that transfection with the miR-185 mimic significantly
weakened the proliferative capacity of KYSE-960 cells compared with
in the control group (P=0.002); however, the miR-185 mimic was not
able to significantly inhibit the proliferative capacity of T.Tn
cells (P=0.6) (Fig. 3D).
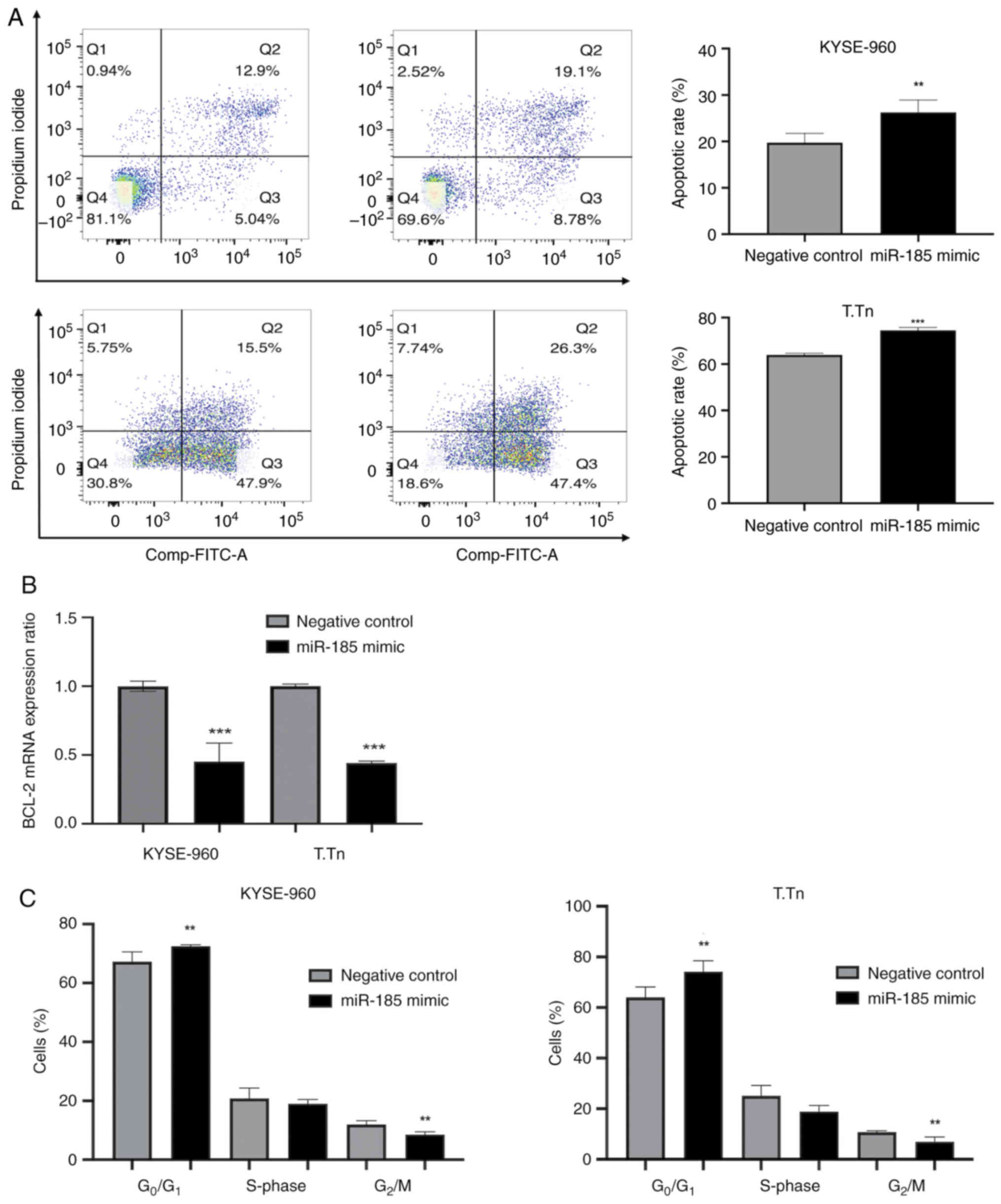
miR-185 regulates apoptosis after
cisplatin treatment and cell cycle progression in vitro
The apoptotic rate of miR-185-transfected KYSE-960
and T.Tn cells after cisplatin exposure for 48 h was significantly
higher than that of the negative control cells (KYSE-960, P=0.027;
T.Tn, P=0.0001; Fig. 4A). In
addition, the mRNA expression levels of BCL-2 were reduced in both
the KYSE-960 and T.Tn miR-185 mimic groups (Fig. 4B). Cell cycle analyses showed that
the percentages of ESCC cells in the G0/G1
phase after miR-185 transfection were significantly higher compared
with those in the negative control groups, whereas the percentages
of ESCC cells in the G2/M phase were significantly lower
(KYSE-960 G0/G1, P<0.05 and
G2/M, P<0.02; T.Tn G0/G1,
P<0.04 and G2/M, P<0.03; Figs. 4C and S5). Notably, KYSE-960 and T.Tn cells in
Fig. 4B and C were not treated with
cisplatin.
miR-185-5p-associated signaling
Enrichment analysis of TCGA data was carried out
using GSEA, and the results revealed that the high miR-185
expression group was enriched in signaling pathways, such as cell
death (apoptosis) and DNA damage (UV response up, reactive oxygen
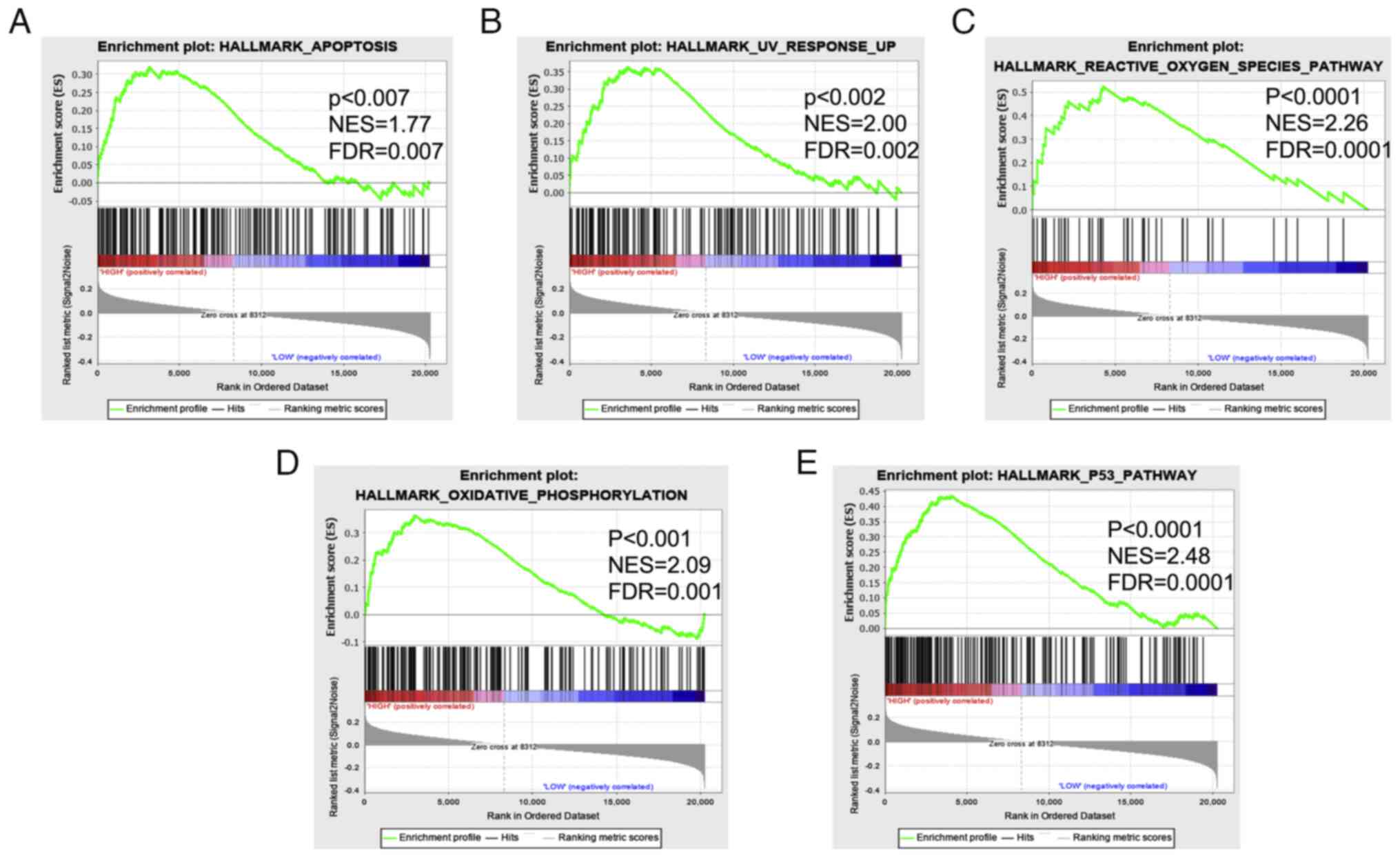
species, oxidative phosphorylation) and p53 signaling (Fig. 5A-E).
Characterization of exosomes from
patient samples
The present study isolated exosomes from the plasma
samples of 89 patients with ESCC. To confirm the successful
isolation of exosomes, TEM was employed to characterize their shape
in the supernatant (Fig. S6).
Notably, plasma exosomes exhibited an elliptical shape.
Relationship between circulating
miR-185 levels and clinicopathological characteristics of patients
with ESCC
The clinicopathological characteristics of patients
are summarized in Table II. The
total number of patients was 89, including nine endoscopically
treated cases, 55 preoperatively untreated surgical cases and 25
cases treated via neoadjuvant chemotherapy, followed by surgery.
Pretreatment staging was performed by whole body analysis,
including PET/CT. Patients were divided into high (n=28) and low
(n=61) circulating miR-185 groups. A cutoff value of 4.3 was set
for the miR-185/miR-16 ratio based on the mean value of miR-185 in
patients with esophageal cancer. In patients with high circulating
exosomal miR-185, the frequency of lymph node metastasis at
preoperative diagnosis was significantly lower (P=0.0045), and
cStage was significantly lower in the high circulating exosomal
miR-185 group (P=0.0001).
| Table II.Demographics and clinicopathological
characteristics of patients with esophageal squamous cell
carcinoma. |
Table II.
Demographics and clinicopathological
characteristics of patients with esophageal squamous cell
carcinoma.
| Variable | High exosomal
miR-185-5p group (n=28) | Low exosomal
miR-185-5p group (n=61) | P-value |
|---|
| Mean age ± SD,
years | 66.6±8.03 | 66.8±8.32 | 0.923a |
| Sex, n |
|
|
|
|
Male/Female | 24/4 | 51/10 |
>0.999b |
| Histological grade,
n |
|
|
|
|
G1/G2/G3/X | 4/15/7/2 | 11/29/13/8 | 0.853b |
| Location of the
tumor |
|
|
|
|
Ce/Ut/Mt/Lt/Ae | 0/2/15/10/1 | 0/8/27/23/3 | 0.836b |
| cT category, n |
|
|
|
| cT1a +
1b/2/3/4a + 4b | 16/7/5/0 | 25/15/21/0 | 0.235c |
| cN category, n |
|
|
|
|
cN0/1/2/3/4 | 20/2/3/0/3 | 28/11/18/4/0 | 0.0045b |
| cM category, n |
|
|
|
|
cM0/1 | 28/0 | 61/0 |
>0.999b |
| cStage, n |
|
|
|
| cStage
0/1/2/3/4a/4b | 7/6/9/3/3/0 | 6/11/19/25/0/0 | 0.0001b |
| Treatment, n |
|
| 0.0597b |
|
Endoscopic | 6 | 3 |
|
| Esophagectomy
first | 16 | 39 |
|
| NAC +
esophagectomy | 6 | 19 |
|
| pT category, n |
|
|
|
| pT0/1a
+ 1b/2/3/4a | 3/19/2/4/0 | 4/31/10/15/1 | 0.411b |
| pN category, n |
|
|
|
|
pN0/1/2/3/4/X | 16/3/4/2/0/3 | 31/7/12/8/1/2 | 0.702b |
| pStage, n |
|
|
|
| pStage
0/1/2/3/4a/X | 2/11/5/5/2/3 | 1/18/14/15/9/4 | 0.527b |
Although the sample size was small, the preoperative
patient group with high circulating exosomal miR-185 levels tended
to exhibit a slightly higher response rate to chemotherapy than
patients with low circulating exosomal miR-185; however, this was
not statistically significant (P=0.081; Table III).
| Table III.Pathological response of patients
with esophageal squamous cell carcinoma treated with neoadjuvant
chemotherapy. |
Table III.
Pathological response of patients
with esophageal squamous cell carcinoma treated with neoadjuvant
chemotherapy.
| Variable | High exosomal
miR-185-5p group (n=6) | Low exosomal
miR-185-5p group (n=19) |
P-valuea |
|---|
| Pathological
response |
|
|
|
| Grade
1a + 1b/2/3 | 3/1/2 | 16/3/0 | 0.081 |
PFS and DSS of patients with ESCC
after initial treatment
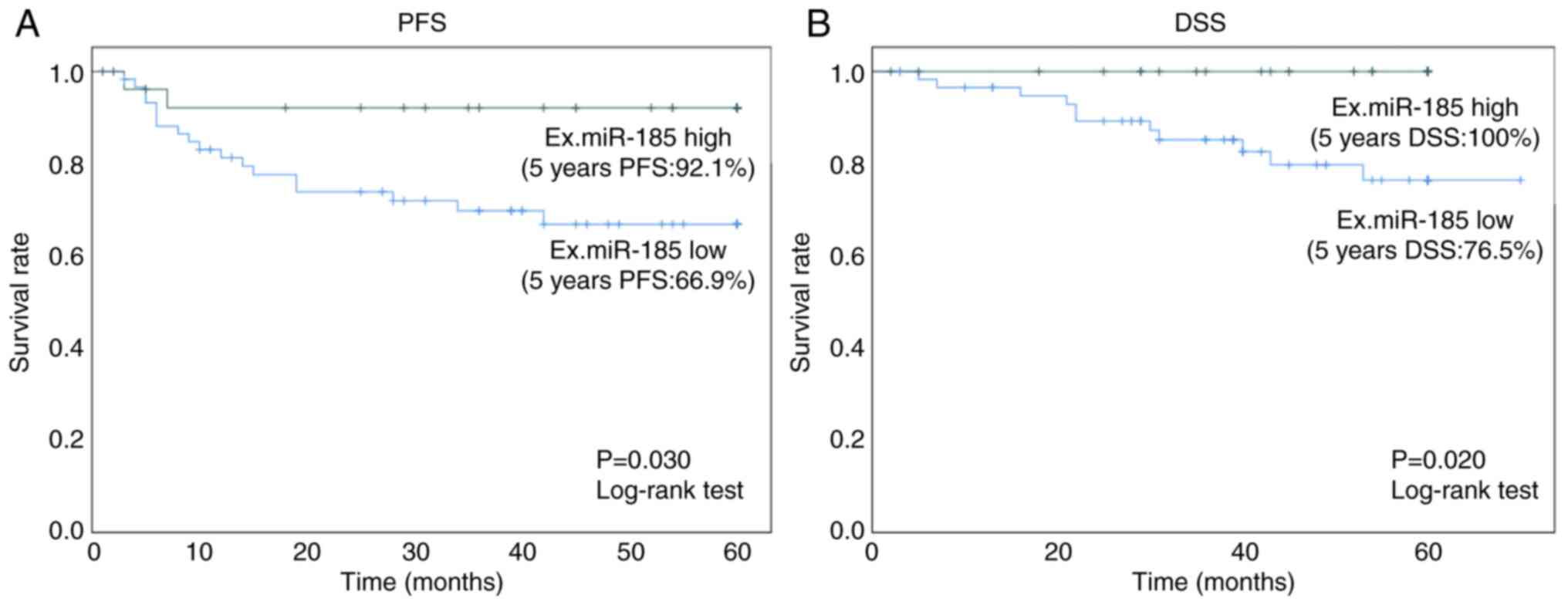
The 5-year PFS rate in the high exosomal miR-185
group was 92.1% (95% CI: 82.0–103.0%), which was significantly
higher (P=0.030, log-rank test) than that in the low exosomal
miR-185 group (66.9%; 95% CI: 54.0–80.0%) (Fig. 6A). The 5-year DSS rate in the high
exosomal miR-185 group was 100%, which was significantly higher
(P=0.020, log-rank test) than that in the low exosomal miR-185
group (76.5%; 95% CI: 64.0–89.0%; Fig.
6B).
Discussion
ESCC is one of the most lethal types of cancer and
is a public health issue of great concern worldwide (27). Despite advances in its diagnosis and
treatment, patient survival rates remain unsatisfactory (28). Therefore, the identification of
novel biomarkers and therapeutic targets in ESCC is urgently
required.
Exosomal miRNAs can be easily isolated from
peripheral blood, which makes them candidate noninvasive biomarkers
(29). Their potential to serve as
biomarkers in patients with ESCC has been previously reported,
along with a possible mechanism of exosomal trafficking (25). Exosomes influence gene expression,
and thus, various biological processes (30). However, the influence of exosomal
miR-185 levels in plasma on cancer treatment outcomes remains to be
fully elucidated. It has been reported that plasma miR-185 levels
are decreased in patients with ESCC and that tumor metastasis is
suppressed by targeting RAGE (21);
the tumor suppressive role of miR-185 in this previous report is
consistent with the results of present study. The impact of
different storage conditions and treatments on the stability and
abundance of individual and total miRNAs in human plasma and plasma
exosomes has been investigated; it has been highlighted that
exosomal miRNAs have the potential to serve as biomarkers based on
their increased stability under various conditions compared with
plasma miRNAs (31). Thus, there is
a possibility that exosomal miRNAs could serve as novel therapeutic
targets for the development of effective methods for the treatment
of ESCC. However, to the best of our knowledge, the present study
is the first to determine the different profiles of circulating
exosomal miR-185 levels in patients with ESCC. The results revealed
that exosomal miR-185 was associated with lymph node metastasis and
may act as a predictor of the prognosis of patients with ESCC. The
data from the current study suggested that exosomal miR-185 may be
important for esophageal cancer initiation and progression, and
that it could hold promise as a novel suppressor of metastasis in
esophageal cancer.
Hypoxia enhances the degree of glycolysis,
angiogenesis and other survival responses in tumors, as well as
their invasion and metastasis, by activating relevant gene
expression through HIFs (32). The
expression levels of HIF-1α are elevated in cancer cell lines under
hypoxia; notably, HIF-1α drives oncogenic expression in ESCC and is
associated with a poor prognosis (10). In the present study, the expression
of 33 exosomal miRNAs was downregulated in KYSE-960 and T.Tn cell
culture media under hypoxia, whereas certain exosomal miRNAs were
upregulated. Notably, exosomal miR-185 expression was decreased
under hypoxia in both cell line culture media.
miRNA dysregulation serves a critical role in the
initiation and progression of multiple types of human cancer
(33,34), by either promoting (35) or suppressing tumorigenesis (36). The characterization of miRNAs
involved in ESCC progression and their targets may contribute to
the identification of new prognostic markers and therapeutic
targets (37). Circulating miRNAs
have recently emerged as potential biomarkers for various types of
cancer (38). In the present study,
exosomal miR-185 was significantly associated with cN and cStage,
with high exosomal miR-185 levels in plasma being associated with a
good prognosis in patients with ESCC. Upregulation of miR-185 may
have a suppressive role in tumor malignancy, paving the way for the
development of effective treatment methods in ESCC.
miRNAs are associated with lymph node metastasis in
esophageal cancer (22,33,39),
which may provide a novel insight into the design of better
therapeutic strategies. Furthermore, albeit based on a limited
number of cases, the present findings indicated that circulating
miR-185 might not be able to influence chemotherapy sensitivity,
but that it could predict lymph node metastasis prior to
chemotherapy. To provide new insights for designing better
therapeutic strategies to treat esophageal cancer in patients with
lymph node metastasis and to predict prognosis more accurately, a
further study in a larger cohort and a detailed mechanistic
investigation are warranted.
Overcoming cisplatin resistance is a major aim in
cancer therapy. One of the most widely accepted approaches is
through combination with other agents that enhance cisplatin
toxicity (40). In the present
study, overexpression of miR-185 regulated cancer cell cycle
progression and induced apoptosis following cisplatin treatment.
The apoptotic rate in a human gastric cancer cell line has been
reported to be ~10% without any other intervention except for
miR-185 transfection (41). In
another study, the apoptotic rate in a breast cancer cell line was
reported to be ~20% without any other intervention except for
miR-185 transfection (42). In the
present study, the negative control group comprised ESCC cells
transfected with the control mimic and treated with cisplatin for
48 h, and the target group comprised ESCC cells treated with
cisplatin and transfected with the miR-185 mimic. Therefore,
considering the previously reported numbers, and the fact that all
cells that were treated with cisplatin, it is reasonable that the
apoptotic rate detected in the present study was >10%. Tumor
cells may utilize several molecular mechanisms to suppress
apoptosis and acquire resistance to cytotoxic agents, such as via
upregulation of the antiapoptotic protein BCL-2 (43). In the present study, the mRNA
expression levels of BCL-2 were suppressed in
miR-185-overexpressing cells.
The present study also observed a significant
enrichment for hallmark gene sets related to cell death
(apoptosis), DNA damage (UV response up, reactive oxygen species
and oxidative phosphorylation), and p53 in patients with high
versus low miR-185 expression. Such dysregulation is expected to
serve a significant role in esophageal cancer cell transformation.
These data suggested that miR-185 alters cancer-associated pathway
activity.
In the present study, miR-16 was used as an internal
control for the clinical evaluation of cell-free miR-185 and U6 was
used as an internal reference for evaluation of cellular miR-185;
however, a limitation is that these internal controls are not
standardized and can only be quantified in relative amounts, which
may not accurately reflect the amplification of the primary target
and ultimately lead to different Cq value results. Development of
accurate internal controls or quantification of absolute value are
thus required for further confirmation in future studies. In
addition, larger patient cohorts are required for future
confirmation of the use of exosomal miR-185 in the prediction of
prognosis.
In summary, the data from the current study
suggested that miR-185 may be important in ESCC progression and
holds promise as a novel suppressor of metastasis in ESCC. Although
there are many factors, such as target genes, that can be used as a
guide or reference when considering therapeutic efficacy,
circulating exosomal miR-185 may act as a potential prognostic
biomarker and could have potential as a novel therapeutic target in
ESCC.
Supplementary Material
Supporting Data
Acknowledgements
The authors are grateful to Ms. Keiko Iida
(Department of Frontier Surgery, Chiba University Graduate School
of Medicine) for technical advice and assistance.
Funding
This study was supported by JSPS KAKENHI (grant nos. 15K19872,
19K23880 and 21K16414).
Availability of data and materials
The data generated in the present study may be
requested from the corresponding author. Microarray analysis data
generated in the present study may be found in the Gene Expression
Omnibus database under accession number GSE263921 or at the
following URL: https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE263921.
Authors' contributions
YM and HMa conceived the theme, AM and YM designed
the study. HW, YM, KKand MK performed the comprehensive analysis of
microRNA. AM and HW developed the methodology for in vitro
analyses. TS, KM, SI, HMo, TM and YN performed clinical data
collection and validation. AM, KM, TT, RO and JH analyzed and
interpreted clinical and in vitro data. TS and KM confirm the
authenticity of all the raw data. AM and YM wrote the draft of the
manuscript under the supervision of MK and HMa. All authors were
involved in the planning, data interpretation and core revision of
the paper, and read and approved the final version of the
manuscript.
Ethics approval and consent to
participate
This study was approved by the Ethics Committee of
the Graduate School of Medicine, Chiba University (approval no.
1264). Written informed consent was obtained from all patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Author's information
Yasunori Matsumoto ORCI: https://orcid.org/0000-0002-6239-6691
Glossary
Abbreviations
Abbreviations:
|
ESCC
|
esophageal squamous cell carcinoma
|
|
RT-qPCR
|
reverse transcription-quantitative
PCR
|
|
HIF
|
hypoxia-inducible factor
|
|
GSEA
|
Gene Set Enrichment Analysis
|
|
TCGA
|
The Cancer Genome Atlas
|
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jin W, Luo W, Fang W, Wang Y, Wang L, Shen
Q, Liu W and Zhang H: miR-145 expression level in tissue predicts
prognosis of patients with esophageal squamous cell carcinoma.
Pathol Res Pract. 215:1524012019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chen JG, Chen HZ, Zhu J, Yang YL, Zhang
YH, Huang PX, Chen YS, Zhu CY, Yang LP, Shen K, et al: Cancer
survival in patients from a hospital-based cancer registry, China.
J Cancer. 9:851–860. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rice TW, Ishwaran H, Hofstetter WL,
Schipper PH, Kesler KA, Law S, Lerut EM, Denlinger CE, Salo JA,
Scott WJ, et al: Esophageal Cancer: Associations With (pN+) Lymph
Node Metastases. Ann Surg. 265:122–129. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Skotland T, Sagini K, Sandvig K and
Llorente A: An emerging focus on lipids in extracellular vesicles.
Adv Drug Deliv Rev. 159:308–321. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yang H, Fu H, Wang B, Zhang X, Mao J, Li
X, Wang M, Sun Z, Qian H and Xu W: Exosomal miR-423-5p targets SUFU
to promote cancer growth and metastasis and serves as a novel
marker for gastric cancer. Mol Carcinog. 57:1223–1236. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Matsumoto Y, Kano M, Akutsu Y, Hanari N,
Hoshino I, Murakami K, Usui A, Suito H, Takahashi M, Otsuka R, et
al: Quantification of plasma exosome is a potential prognostic
marker for esophageal squamous cell carcinoma. Oncol Rep.
36:2535–2543. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Matsumoto Y, Kano M, Murakami K, Toyozumi
T, Suito H, Takahashi M, Sekino N, Shiraishi T, Kamata T, Ryuzaki
T, et al: Tumor-derived exosomes influence the cell cycle and cell
migration of human esophageal cancer cell lines. Cancer Sci.
111:4348–4358. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fang P, Zhou J, Liang Z, Yang Y, Luan S,
Xiao X, Li X, Zhang H, Shang Q, Zeng X and Yuan Y: Immunotherapy
resistance in esophageal cancer: Possible mechanisms and clinical
implications. Front Immunol. 13:9759862022. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tang K, Toyozumi T, Murakami K, Sakata H,
Kano M, Endo S, Matsumoto Y, Suito H, Takahashi M, Sekino N, et al:
HIF-1α stimulates the progression of oesophageal squamous cell
carcinoma by activating the Wnt/β-catenin signalling pathway. Br J
Cancer. 127:474–487. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Berezikov E, Guryev V, van de Belt J,
Wienholds E, Plasterk RH and Cuppen E: Phylogenetic shadowing and
computational identification of human microRNA genes. Cell.
120:21–24. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hemmatzadeh M, Mohammadi H, Karimi M,
Musavishenas MH and Baradaran B: Differential role of microRNAs in
the pathogenesis and treatment of esophageal cancer. Biomed
Pharmacother. 82:509–519. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Harada K, Baba Y, Ishimoto T, Shigaki H,
Kosumi K, Yoshida N, Watanabe M and Baba H: The role of microRNA in
esophageal squamous cell carcinoma. J Gastroenterol. 51:520–530.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Niu Y and Tang G: miR-185-5p targets ROCK2
and inhibits cell migration and invasion of hepatocellular
carcinoma. Oncol Lett. 17:5087–5093. 2019.PubMed/NCBI
|
|
15
|
Liu C, Cai L and Li H: miR-185 regulates
the growth of osteosarcoma cells via targeting hexokinase 2. Mol
Med Rep. 20:2774–2782. 2019.PubMed/NCBI
|
|
16
|
Ostadrahimi S, Abedi Valugerdi M, Hassan
M, Haddad G, Fayaz S, Parvizhamidi M, Mahdian R and Fard Esfahani
P: miR-1266-5p and miR-185-5p promote cell apoptosis in human
prostate cancer cell lines. Asian Pac J Cancer Prev. 19:2305–2311.
2018.PubMed/NCBI
|
|
17
|
Zhang W, Sun Z, Su L, Wang F, Jiang Y, Yu
D, Zhang F, Sun Z and Liang W: miRNA-185 serves as a prognostic
factor and suppresses migration and invasion through Wnt1 in colon
cancer. Eur J Pharmacol. 825:75–84. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tan Z, Jiang H, Wu Y, Xie L, Dai W, Tang H
and Tang S: miR-185 is an independent prognosis factor and
suppresses tumor metastasis in gastric cancer. Mol Cell Biochem.
386:223–231. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li BX, Yu Q, Shi ZL, Li P and Fu S:
Circulating microRNAs in esophageal squamous cell carcinoma:
Association with locoregional staging and survival. Int J Clin Exp
Med. 8:7241–7250. 2015.PubMed/NCBI
|
|
20
|
Zhao ZT, Zhou W, Liu LY, Lan T, Zhan QM
and Song YM: Molecular mechanism and effect of microRNA185 on
proliferation, migration and invasion of esophageal squamous cell
carcinoma. Zhonghua Yi Xue Za Zhi. 93:1426–1431. 2013.(In Chinese).
PubMed/NCBI
|
|
21
|
Jing R, Chen W, Wang H, Ju S, Cong H, Sun
B, Jin Q, Chu S, Xu L and Cui M: Plasma miR-185 is decreased in
patients with esophageal squamous cell carcinoma and might suppress
tumor migration and invasion by targeting RAGE. Am J Physiol
Gastrointest Liver Physiol. 309:G719–G729. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Japan Esophageal Society: Japanese
classification of esophageal cancer, 11th edition: Part I.
Esophagus. 14:1–36. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sashiyama H, Shino Y, Kawamata Y, Tomita
Y, Ogawa N, Shimada H, Kobayashi S, Asano T, Ochiai T and Shirasawa
H: Immortalization of human esophageal keratinocytes by E6 and E7
of human papillomavirus type 16. Int J Oncol. 19:97–103.
2001.PubMed/NCBI
|
|
24
|
Jung MK and Mun JY: Sample preparation and
imaging of exosomes by transmission electron microscopy. J Vis Exp.
131:564822018.
|
|
25
|
Takeshita N, Hoshino I, Mori M, Akutsu Y,
Hanari N, Yoneyama Y, Ikeda N, Isozaki Y, Maruyama T, Akanuma N, et
al: Serum microRNA expression profile: miR-1246 as a novel
diagnostic and prognostic biomarker for oesophageal squamous cell
carcinoma. Br J Cancer. 108:644–652. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Talukdar FR, di Pietro M, Secrier M,
Moehler M, Goepfert K, Lima SSC, Pinto LFR, Hendricks D, Parker MI
and Herceg Z: Molecular landscape of esophageal cancer:
Implications for early detection and personalized therapy. Ann N Y
Acad Sci. 1434:342–359. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang Y: Epidemiology of esophageal
cancer. World J Gastroenterol. 19:5598–5606. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Manier S, Liu CJ, Avet-Loiseau H, Park J,
Shi J, Campigotto F, Salem KZ, Huynh D, Glavey SV, Rivotto B, et
al: Prognostic role of circulating exosomal miRNAs in multiple
myeloma. Blood. 129:2429–2436. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mercer TR and Mattick JS: Structure and
function of long noncoding RNAs in epigenetic regulation. Nat
Struct Mol Biol. 20:300–307. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ge Q, Zhou Y, Lu J, Bai Y, Xie X and Lu Z:
miRNA in plasma exosome is stable under different storage
conditions. Molecules. 19:1568–1575. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lu X and Kang Y: Hypoxia and
hypoxia-inducible factors: Master regulators of metastasis. Clin
Cancer Res. 16:5928–5935. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Valeri N, Braconi C, Gasparini P, Murgia
C, Lampis A, Paulus-Hock V, Hart JR, Ueno L, Grivennikov SI, Lovat
F, et al: MicroRNA-135b promotes cancer progression by acting as a
downstream effector of oncogenic pathways in colon cancer. Cancer
Cell. 25:469–483. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhou W, Fong MY, Min Y, Somlo G, Liu L,
Palomares MR, Yu Y, Chow A, O'Connor ST, Chin AR, et al:
Cancer-secreted miR-105 destroys vascular endothelial barriers to
promote metastasis. Cancer Cell. 25:501–515. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Garofalo M, Di Leva G, Romano G, Nuovo G,
Suh SS, Ngankeu A, Taccioli C, Pichiorri F, Alder H, Secchiero P,
et al: miR-221&222 regulate TRAIL resistance and enhance
tumorigenicity through PTEN and TIMP3 downregulation. Cancer Cell.
16:498–509. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhang Y, Yang P, Sun T, Li D, Xu X, Rui Y,
Li C, Chong M, Ibrahim T, Mercatali L, et al: miR-126 and miR-126*
repress recruitment of mesenchymal stem cells and inflammatory
monocytes to inhibit breast cancer metastasis. Nat Cell Biol.
15:284–294. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Mao Y, Li L, Liu J, Wang L and Zhou Y:
MiR-495 inhibits esophageal squamous cell carcinoma progression by
targeting Akt1. Oncotarget. 7:51223–51236. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhao Y, Song Y, Yao L, Song G and Teng C:
Circulating microRNAs: Promising biomarkers involved in several
cancers and other diseases. DNA Cell Biol. 36:77–94. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Iorio MV and Croce CM: MicroRNAs in
cancer: Small molecules with a huge impact. J Clin Oncol.
27:5848–5856. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Xiang Y, Ma N, Wang D, Zhang Y, Zhou J, Wu
G, Zhao R, Huang H, Wang X, Qiao Y, et al: MiR-152 and miR-185
co-contribute to ovarian cancer cells cisplatin sensitivity by
targeting DNMT1 directly: A novel epigenetic therapy independent of
decitabine. Oncogene. 33:378–386. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Fan L, Tan B, Li Y, Zhao Q, Yuan H, Liu Y,
Wang D and Zhang Z: Upregulation of miR-185 promotes apoptosis of
the human gastric cancer cell line MGC803. Mol Med Rep.
17:3115–3122. 2018.PubMed/NCBI
|
|
42
|
Değerli E, Torun V and Cansaran-Duman D:
miR-185-5p response to usnic acid suppresses proliferation and
regulating apoptosis in breast cancer cell by targeting Bcl2. Biol
Res. 53:192020. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Hassan M, Watari H, AbuAlmaaty A, Ohba Y
and Sakuragi N: Apoptosis and molecular targeting therapy in
cancer. Biomed Res Int. 2014:1508452014. View Article : Google Scholar : PubMed/NCBI
|