Introduction
Radiotherapy is an effective tool for malignant
tumor treatments. It is particularly beneficial for rapidly growing
tumors due to their high susceptibility to radiation (1,2).
Ionizing radiation can produce high amounts of reactive oxygen
species (ROS) and free radicals, which further induce cellular DNA
single-strand breaks (SSBs) and double-strand breaks (DSBs),
eventually resulting in the death of cancer cells (3,4).
However, local failure after irradiation remains a challenge due to
intrinsic and acquired resistance of tumor cells to radiation
treatment (5).
Cellular antioxidants are important for the
protection of cells against ROS and free radicals generated through
endogenous or exogenous oxidative stress (6–8).
Glutathione (GSH), the tripeptide thiol
L-γ-glutamyl-L-cysteinyl-glycine, is one of the most abundant
antioxidants in the cells with concentrations in the low millimolar
range. It plays a key role in maintaining intracellular redox
equilibrium and in augmenting cellular defenses to harmful factors
(9,10). In general, GSH is synthesized from
its constituent amino acids, glutamic acid, cysteine and glycine,
via two sequential ATP-consuming steps, which are catalyzed by
γ-glutamylcysteine synthetase (γ-GCS) and GSH synthase (GSS). The
synthesis of the di-peptide γ-glutamylcysteine by γ-GCS is the
rate-limiting step in de novo GSH synthesis (11,12).
The level of GSH is reported to be increased in various types of
tumor cells, for example in bone marrow, breast, glioma, colon,
larynx and lung cancers (13–18).
Owing to its protection, an increased level of GSH is often
associated with a refractory response to ionizing radiation via GSH
scavenging ROS and free radicals. Conversely, depletion of
intracellular GSH favorably increases the radiosensitivity of
cancer cells (19,20).
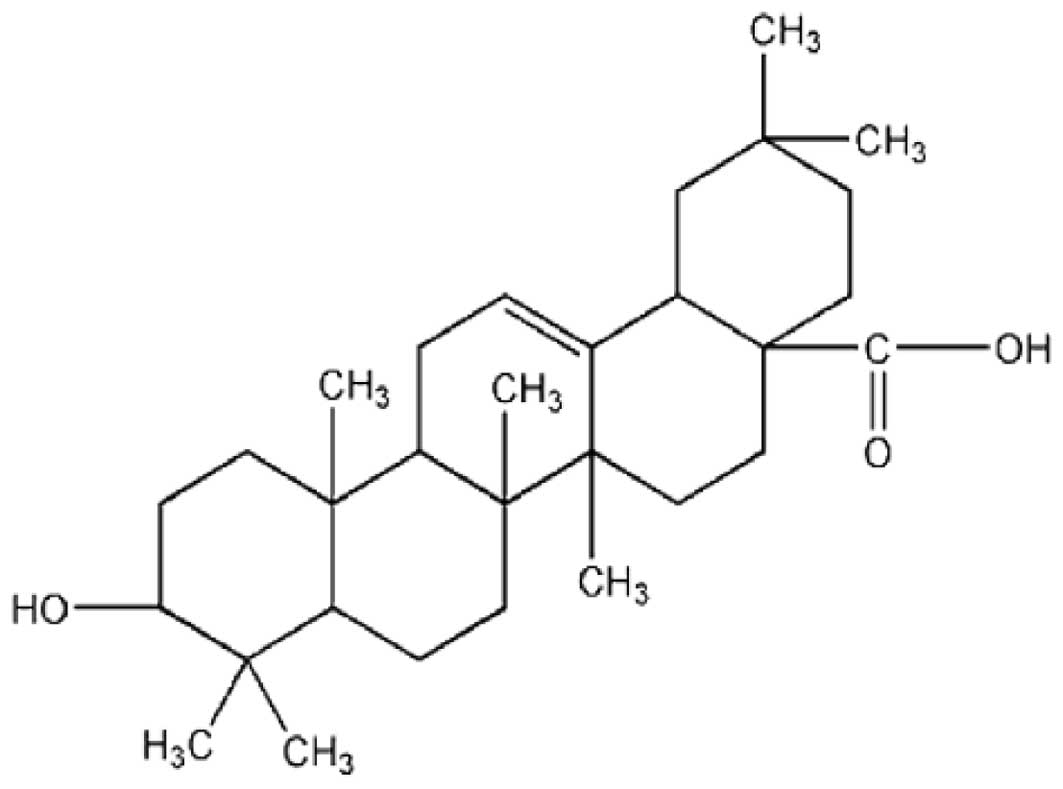
Oleanolic acid (3b-hydroxy-olea-12-en-28-oic acid;
OA) belongs to the triterpenoid family (Fig. 1), which is widely distributed in the
plant kingdom (21). The
pentacyclic triterpenoid with a wide variety of functional groups
is commonly used to treat various diseases. OA demonstrates
anti-inflammatory activity (22),
protection against hepatotoxicity (23,24),
and recovery of the hematopoietic system after irradiation
(25). Meanwhile, OA was found to
induce differentiation of malignant cells, to inhibit proliferation
of carcinoma cells and to promote apoptosis (26,27).
It has been well established that plant extracts and their
derivatives have high potential capability as new anticancer
therapeutic agents, either alone or in combination with
conventional chemotherapeutic agents for cancers that exhibit
resistance to standard drugs (28,29).
However, whether OA increases the radiotherapeutic effect on cancer
cells or not remains unclear. Therefore, in the present study, we
investigated the influence of the monomer OA on the
radiosensitivity of cancer cells and the alteration in
intracellular GSH level. Our results demonstrated that OA enhances
the radiosensitivity of C6 and A549 cells through inhibition of
intracellular GSH synthesis.
Materials and methods
Cell culture and treatment
The rat glioma C6 and human lung cancer A549 cell
lines (Cell Bank, Chinese Academy of Sciences) were cultured in
RPMI-1640 medium supplemented with 10% fetal bovine serum (FBS)
(both from Gibco, Carlsbad, CA, USA), penicillin (100 U/ml) and
streptomycin (100 μg/ml) (Sigma-Aldrich, St. Louis, MO, USA) at
37°C in an incubator containing a humid atmosphere of 95% air and
5% CO2 and propagated according to the protocol supplied
by the American Type Culture Collection. OA was purchased from
NanJing-Zelang (China) and dissolved in dimethyl sulfoxide (DMSO;
Sigma-Aldrich) at a stock concentration of 250 μg/ml and stored at
−20°C. All cells were treated with OA at different concentrations
for 24 h prior to exposure to irradiation. In addition, 5 mM
N-acetylcysteine (NAC) (Sigma-Aldrich), an antioxidant and GSH
precursor, was added to the cultured cells 12 h prior to
irradiation in order to further confirm the mechanism of OA
modulation of the radiosensitivity of tumor cells by influencing
the GSH level.
Cell viability assay
The influence of OA on cell growth was determined
using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium
bromide (MTT, Sigma-Aldrich) method. C6 and A549 cells were seeded
in 96-well plates at a density of 5×103 cells/well. They
were then treated with different concentrations of OA for 24 h.
Furthermore, the medium was replaced with fresh medium allowing
cells to be continuously grown up to 72 h. MTT dye was added to a
final concentration of 0.5 mg/ml, and cells were subsequently
incubated for an additional 4 h at 37°C. The medium containing
residual MTT dye was carefully aspirated from each of the wells,
and 200 μl DMSO was added to each well to dissolve the reduced
formazan dye. The fraction of viable cells was calculated by
comparing the optical absorbance of the culture exposed to OA
treatment with that of the untreated control.
Irradiation
Irradiation was emitted using a 6-MV X-ray linear
accelerator (Varian Inc., Palo Alto, CA, USA) at a dose rate of 250
cGy/min.
Clonogenic assay
The radiosensitivity of tumor cells was determined
using a clonogenic assay. Both tumor cell lines were seeded and
cultured overnight at an appropriate density in T25 flasks, and
subsequently different drugs were added into the medium for 24 h.
After being pretreated with control, OA, or OA and NAC, cells were
subjected to 0, 1, 2, 3, 5, or 7 Gy X-ray irradiation. The medium
was then replaced with fresh medium allowing cells to continuously
grow for colony formation for 9 to 12 days. Cell colonies were
fixed by absolute methanol and stained with Giemsa for counting.
Clonogenic survival fraction (SF) was calculated as the number of
colonies/(the number of seeded cells × plating efficiency). Plating
efficiency was defined as the number of colonies/the number of
seeded cells of the untreated control. Survival curve was fitted
with the single target multi-model of the equation: S =
1−1(1−e−D/D0)N.
Micronucleus assay
The number of micronuclei (MN) were measured with
the cytokinesis-block technique as a biological end point for the
response of mimetic hypoxia to irradiation. Briefly, the cells were
exposed to 0.83 μg/ml cytochalasin B (Sigma-Aldrich) for 19–20 h
followed by 75 mM KCl hypotonic treatment for 1–3 min and then
fixed in situ with methanol:acetic acid (9:1 v/v) for 30
min. Air-dried cells were stained with 5% Giemsa (Sigma-Aldrich)
for 10 min. Micronuclei were scored in binucleated cells, and the
formation of binucleated cells was measured as the percentage of
the total number of cells scored. For each sample, at least 1,000
binucleated cells were counted. The MN yield, YMN, was
the ratio of the number of micronuclei to the number of binucleated
cells scored.
Intracellular GSH assay
After triplicate samples of 106 cells
were treated with different reagents, the intracellular levels of
GSH were measured with the glutathione
reductase/5,5′-dithiobis-(2-nitrobenzoic acid) (DTNB) recycling
assay kit (Beyotime Biotechnology, Jiangsu, China) under the
methods recommended by the manufacturer. Briefly, GSH was
determined by using a reaction mixture, containing 50 μl of cell
lysates, 50 μl of 2.4 mM DTNB and 50 μl of 10.64 mU/μl glutathione
reductase in the assay buffer (pH 7.5) containing 153 mM sodium
phosphate and 8.4 mM EDTA. After a 5-min incubation at 25°C, the
reaction was started by the addition of 50 μl NADPH solution (0.16
mg/ml) in the assay buffer. The standard sample and checking sample
cuvettes were placed into a dual-beam spectrophotometer, and the
increases in absorbance at 412 nm were followed as a function of
time.
Intracellular γ-GCS activity assay
Cells (106) were homogenized in 50 mM
potassium phosphate (pH 7.5) containing TES/SB buffer (20 mM Tris,
1 mM EDTA, 250 mM sucrose, 20 mM sodium borate, 2 mM serine) for
the γ-GCS assay. Homogenates were centrifuged at 12,000 rpm (15
min, 4°C), and the supernatants were maintained on ice for
determination of enzyme activity. The protein concentration of cell
supernatants was measured using the Bio-Rad DC Protein Assay kit
(Bio-Rad, Herts, UK), and the enzyme activity was reported as U/mg
protein, where a unit of activity was the amount of enzyme required
to convert one micromole of substrate to product per min at 25°C.
The γ-GCS assay was an adaptation of a previously described method,
in which γ-GCS in cell extracts synthesized γ-glutamylcysteine
which was then reacted with 2,3-naphthalenedicarboxaldehyde (NDA)
to form a highly fluorescent product that was able to be measured
fluorimetrically at 520 nm (30).
Intracellular GSS activity assay
Tumor cells were plated in 60-mm culture dishes at a
density of 106 cells/dish and then divided into
different groups with various methods of pretreatment. The
intracellular GSS activities were measured using GSS assay kit
(Lanxu Biotech Co. Ltd., China). Briefly, the cells were subjected
to repeated freeze-thaw cycles to lyse the cells for release of
intracellular components. Cell lysates were centrifuged at 3,000
rpm (20 min, 4°C), and the supernatants were maintained on ice for
determination of enzyme activity. All procedures were performed
according to the protocol of the kit. After the reaction was
terminated, the absorbance was measured at 450 nm on an
enzyme-linked immunosorbent assay (ELISA) reader. The activity of
GSS in the sample was then determined by comparing the OD of the
samples to the standard curve.
Statistical analysis
Data are reported as the means ± SEM of 3 separate
experiments. Statistical analysis was measured by the independent
sample t-test and analysis of variance. P<0.05 was considered to
indicate a statistically significant result.
Results
Selection of the experimental
concentration of OA
In order to select the experimental concentration of
OA for this study, a 24 h dose-response study was conducted by
exposing the cells lines to different concentrations of OA. The
results of the MTT assay showed that the half inhibition
concentration (IC50) of OA for C6 cells was 35 μg/ml and
for the A549 cells this value was 27 μg/ml, respectively (Fig. 2). 10% IC50, 20%
IC50 and 30% IC50 of drug concentrations were
used to determine the influence of OA on the radiosensitivities of
both tumor cell lines.
Influence of OA on the radiosensitivities
of the tumor cells
No statistically significant changes were noted in
the formation of C6 and A549 clonogenic cells following treatment
with different concentrations of OA. Based on the single target
multi-model, the fitting curves of the irradiated C6 and A549 cells
had a gradually declining tendency with increasing doses. The SFs
of irradiated cells, after OA pretreatment for 24 h, were further
decreased when compared to those of the irradiated cells without OA
treatment (Fig. 3). By calculating
the D0 value and the sensitive enhancement ratio (SER),
the D0 and SERs of irradiated cells were reduced in a OA
concentration-dependent manner. When the concentration of OA
reached 10% IC50, 20% IC50 and 30%
IC50, the SERs of C6 and A549 cell were 1.16, 1.81, 2.23
and 1.13, 1.26, 1.55, respectively. In contrast, the treatment of
NAC partially reduced the sensitizing effect of OA at 30%
IC50 on the irradiated cells. Therefore, OA obviously
enhanced the radiosensitivity of both tumor cell lines.
Changes in the intracellular micronucleus
rate by OA pretreatment
The results from the MN assay showed there was no
obvious influence of the various concentrations of OA on the
formation of the numbered MN in both cell lines unexposed to
X-rays. Subsequently, the numbers of intracellular MN were
significantly increased with concomitant irradiation doses. In both
irradiated cell lines pretreatment with various concentrations of
OA, further enhanced the numbers of intracellular MN. Meanwhile, it
was found that, compared with the irradiated cells without OA
treatment, the irradiated cells treated with 20% IC50
and 30% IC50 of OA displayed a statistically significant
increase in cellular MN formation. Additionally, it was obvious
that the capability of 30% IC50 OA-induced MN generation
in irradiated cells was reduced after NAC was added into the
culture medium. These results indicate that OA prompts the
generation of MN in irradiated tumor cells (Fig. 4).
Effect of OA on the GSH level
To further study the mechanism of the influence of
OA on the radiosensitivity of tumor cells, intracellular GSH levels
were measured after pretreatment with different concentrations of
OA for 24 h. As shown in Fig. 5,
when compared with the cells in the absence of OA, significant
decreases in the GSH levels of C6 cells in the presence of OA were
noted. Furthermore, the same phenomenon was noted in A549 cells,
where intracellular GSH levels showed a gradually declining
tendency concomitant with increases in OA concentrations. In
contrast, supplementation of NAC significantly restored the
inhibition of GSH synthesis by OA. The results revealed that OA
effectively inhibits the synthesis of GSH in tumor cells.
Inhibition of γ-GCS activity by OA
Since γ-GCS is the rate-limiting enzyme in the
synthesis of intracellular GSH, its activity was further measured.
As showed in Fig. 6, the activity
of γ-GCS in C6 cells was inhibited by the treatment with 20%
IC50 and 30% IC50 of OA, while no statistical
reduction in the activity of γ-GCS was noted following 10%
IC50 of OA treatment. Unlike C6 cells, all
concentrations of OA treatment inhibited the enzyme activity in
A549 cells. The results indicated that NAC could not reverse the
OA-inhibited activity of γ-GCS in both tumor cell lines. It was
shown that OA has the capacity of inhibiting the activity of γ-GCS
in tumor cells.
Influence of OA on the activity of
GSS
GSS is another synthetic enzyme of GSH. Thus, the
activity of GSS in both cell lines following OA treatment was
determined. As shown in Fig. 7, we
found that OA pretreatment failed to affect the intracellular
activity of GSS in the C6 cells. A similar phenomenon was also
observed in A549 cells when OA concentration was 10%
IC50 or 20% IC50, even 30% IC50 of
OA could statistically enhance the activity of GSS. Meanwhile,
compared to the cells treated with 30% IC50 of OA, both
C6 and A549 cells treated with 30% IC50 of OA and NAC
did not show an obvious alteration in GSS activity. The results
thus revealed that OA does not decrease the activity of GSS in
tumor cells.
Discussion
Ionizing radiation continues to be a frontline
therapy for local control of glioma and lung cancer where surgery
is either not possible or undesirable (31,32).
However, regarding the radiotherapeutics of lung cancer, radiation
pneumonia is one serious complication after patient exposure to
high-dose irradiation (33). Thus,
it is favorable to decrease the incidence of radiation pneumonia by
enhancement of the radiosensitivity of cancer cells with
pretreatment of a radiosensitizing agent. Regarding glioma, an
appropriate radiosensitizing drug needs to have the capability for
permeating the blood-brain barrier for achieving effective plasma
concentrations in the region of the tumor. There are numerous
natural compounds from the extracts of plants, which have the
potential to increase the radiosensitivity of tumor cells (34–36).
In the present study, we investigated the radiosensitizing effect
of OA on glioma and lung cancer cell lines. The natural OA extract
in the form of free acid has the ability of infiltrating the
blood-brain barrier (37). The
rationale for combined treatment of non-toxic concentrations of OA
with radiation is to observe whether it leads to a greater extent
of tumor cell death. We thus selected three different
concentrations of OA, which had no obvious influence on cell
viability, to carry out the radiosensitizing experiment. Different
doses of radiation combined with OA greatly inhibited the cell
growth and achieved an additive effect. According to the
calculation of D0 and SERs, the sensitivity of tumor
cells to radiation was significantly enhanced by OA treatment. In a
previous study, the radiosensitizing effect of ursolic acid, an
isomer of OA, was demonstrated in diverse cell types and in
vivo(38). Additionally,
experimental data indicate that betulinic acid, another pentacyclic
triterpenoid, may be a useful agent for tumors that are resistant
to irradiation such as head and neck cancer and melanoma (39,40).
The results from our research also support that OA has
radiosensitizing capacities in tumor cells as confirmed by the
effects of combined treatment of other pentacyclic triterpenoids
and irradiation.
The protective effect of GSH is important for the
resistance of cancer cells against radiotherapeutics. Many
compounds of natural origin are capable of regulating intracellular
GSH levels. For instance, gelomulide K, a natural diterpene
extract, induces a decrease in cellular GSH in cancer cells
(41). It was also observed that
ferulic acid potentiates the efficacy of radiosensitization in
human cervical carcinoma cells by the attenuation of intracellular
GSH (42). Similarly, we further
observed the alteration of intracellular GSH levels in tumor cells
after treatment with different concentrations of OA. The results
showed that OA obviously reduces GSH levels in a
concentration-dependent manner. Because γ-GCS is the key
rate-limiting enzyme synthesizing intracellular GSH, the
intracellular GSH level was decreased by inhibition of γ-GCS
(12,43). Our study demonstrated that the
activity of γ-GCS in tumor cells could be downregulated following
OA treatment. Although GSS is another synthetic enzyme of GSH,
which is rarely considered in the field of radiation biology, we
still observed its activity for the sake of clarifying the target
of the radiosensitizing effect of OA. The experimental results
found that OA does not evidently inhibit the activity of GSS. On
the contrary, its activity was upregulated with 20% IC50
of OA. Consequently, the synthesis of GSH in tumor cells was
decreased via the downregulation of the activity of γ-GCS by OA,
but not by the inhibition of GSS.
Data indicate that MN are the result of small
chromosome fragments that are not incorporated into the daughter
nuclei during cell division, which arise from exposure to
nonrepaired or misrepaired DSBs by various clastogenic agents
(44,45). It has been suggested that DNA damage
by ROS induction leads to formation of (SSBs) and DSBs (46,47).
Meanwhile, the number of radiation-induced MN was strongly
correlated with radiation damage. The MN assay is an appropriate
biological tool for evaluating the radiosensitivity of cells in
vitro due to the good reliability and reproducibility of the
assay (48). In the present study,
treatment of OA alone did not statistically enhance the ratio of MN
in tumor cells. Our results clearly showed that the ratio of MN was
conspicuously increased when tumor cells were exposed to X-rays.
Subsequently, it was found that the combination of OA at different
concentrations and radiation led to higher ratios of MN compared to
radiation alone. According to previous reports, the depletion of
intracellular GSH results in the generation of MN. In order to
verify this theory, NAC, an antioxidant and GSH precursor, was
further used in our study. The outcome demonstrated that the level
of intracellular GSH was enhanced after NAC was added to the
culture medium, but no significant changes in γ-GCS and GSS
activity were noted. The phenomenon was contributed to the
enhancement of this enzymatic reaction substrate, but not the
increasing activity of the enzymes by themselves (49). Moreover, concomitant with the
enhancement of reduced GSH levels, the high ratios of MN with OA at
30% IC50 concentration and irradiation treatment was
decreased by NAC. Based on the high level of intracellular GSH, the
following phenomenon was found. Compared to the cells without NAC
pretreatment, both tumor cell lines treated with NAC showed a
weaker damage after exposure to irradiation.
Importantly, previous data also indicate that OA
attenuates hepatotoxicity and nephrotoxicity of various toxic
agents by increasing the level of GSH, further protecting normal
tissue and cells (50,51). Conversely, our results and other
studies indicate that high levels of GSH in tumor cells are
markedly suppressed by pretreatment of OA and its derivatives
(52,53). Regarding this difference, we presume
that the following factors possibly contributed to the
controversial findings. The protective effect of OA and its
derivatives are via the stimulation of the synthesis of GSH in
normal tissues damaged by the depletion of GSH, while in contrast
the effect of OA is through inhibition of GSH in malignant tumors
with high levels of GSH. Since extremely high or low levels of GSH
may disorder the redox equilibrium of the intracellular
microenvironment (54), OA sustains
the balance via regulation of GSH.
Taken together, our experimental results suggest for
the first time that OA sensitizes rat glioma C6 cells and human
lung cancer A549 cells to radiation in vitro. The mechanism
of this sensitization may involve the inhibition of reduced GSH
synthesis via the downregulation of γ-GCS activity, which in turn
may explain the reduction in clonogenic survival and the increase
in cellular MN. The effect of the natural medicine OA on regulating
the radiation response may provide novel benefit for the treatment
of tumors.
Acknowledgements
This study was financially supported by the China
Postdoctoral Science Foundation (grant no. 201000470822) and Anhui
Provincial Natural Science Foundation, China (grant no.
11040606M210).
References
|
1
|
Shah C, Grills IS, Kestin LL, McGrath S,
Ye H, Martin SK and Yan D: Intrafraction variation of mean tumor
position during image-guided hypofractionated stereotactic body
radiotherapy for lung cancer. Int J Radiat Oncol Biol Phys.
82:1636–1641. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lagadec C, Vlashi E, Della Donna L, Meng
Y, Dekmezian C, Kim K and Pajonk F: Survival and self-renewing
capacity of breast cancer initiating cells during fractionated
radiation treatment. Breast Cancer Res. 12:R132010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jamal M, Rath BH, Williams ES, Camphausen
K and Tofilon PJ: Microenvironmental regulation of glioblastoma
radioresponse. Clin Cancer Res. 16:6049–6059. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ayouaz A, Raynaud C, Heride C, Revaud D
and Sabatier L: Telomeres: hallmarks of radiosensitivity.
Biochimie. 90:60–72. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dumont F, Altmeyer A and Bischoff P:
Radiosensitising agents for the radiotherapy of cancer: novel
molecularly targeted approaches. Expert Opin Ther Pat. 19:775–799.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bondza-Kibangou P, Millot C, El Khoury V
and Millot JM: Antioxidants and doxorubicin supplementation to
modulate CD14 expression and oxidative stress induced by vitamin
D3 and seocalcitol in HL60 cells. Oncol Rep.
18:1513–1519. 2007.PubMed/NCBI
|
|
7
|
Pani G, Galeotti T and Chiarugi P:
Metastasis: cancer cell’s escape from oxidative stress. Cancer
Metastasis Rev. 29:351–378. 2010.
|
|
8
|
Forkink M, Smeitink JA, Brock R, Willems
PH and Koopman WJ: Detection and manipulation of mitochondrial
reactive oxygen species in mammalian cells. Biochim Biophys Acta.
1797:1034–1044. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Forman HJ, Zhang H and Rinna A:
Glutathione: overview of its protective roles, measurement, and
biosynthesis. Mol Aspects Med. 30:1–12. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Circu ML and Aw TY: Glutathione and
apoptosis. Free Radic Res. 42:689–706. 2008. View Article : Google Scholar
|
|
11
|
Wu G, Fang YZ, Yang S, Lupton JR and
Turner ND: Glutathione metabolism and its implications for health.
J Nutr. 134:489–492. 2004.PubMed/NCBI
|
|
12
|
Botta D, White CC, Vliet-Gregg P, et al:
Modulating GSH synthesis using glutamate cysteine ligase transgenic
and gene-targeted mice. Drug Metab Rev. 40:465–477. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang W, Trachootham D, Liu J, et al:
Stromal control of cystine metabolism promotes cancer cell survival
in chronic lymphocytic leukaemia. Nat Cell Biol. 14:276–286. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lewis-Wambi JS, Kim HR, Wambi C, Patel R,
Pyle JR, Klein-Szanto AJ and Jordan VC: Buthionine sulfoximine
sensitizes antihormone-resistant human breast cancer cells to
estrogen-induced apoptosis. Breast Cancer Res. 10:R1042008.
View Article : Google Scholar
|
|
15
|
Ogunrinu TA and Sontheimer H: Hypoxia
increases the dependence of glioma cells on glutathione. J Biol
Chem. 285:37716–37724. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ruiz-Gómez MJ, Souviron A,
Martínez-Morillo M and Gil L: P-glycoprotein, glutathione and
glutathione S-transferase increase in a colon carcinoma cell line
by colchicine. J Physiol Biochem. 56:307–312. 2000.PubMed/NCBI
|
|
17
|
Inci E, Civelek S, Seven A, Inci F, Korkut
N and Burçax G: Laryngeal cancer: in relation to oxidative stress.
Tohoku J Exp Med. 200:17–23. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Honda T, Coppola S, Ghibelli L, et al: GSH
depletion enhances adenoviral bax-induced apoptosis in lung cancer
cells. Cancer Gene Ther. 11:249–255. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Simons AL, Parsons AD, Foster KA, Orcutt
KP, Fath MA and Spitz DR: Inhibition of glutathione and thioredoxin
metabolism enhances sensitivity to perifosine in head and neck
cancer cells. J Oncol. 2009:5195632009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Boivin A, Hanot M, Malesys C, Maalouf M,
Rousson R, Rodriguez-Lafrasse C and Ardail D: Transient alteration
of cellular redox buffering before irradiation triggers apoptosis
in head and neck carcinoma stem and non-stem cells. PLoS One.
6:e145582011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Pollier J and Goossens A: Oleanolic acid.
Phytochemistry. 77:10–15. 2012. View Article : Google Scholar
|
|
22
|
Yang EJ, Lee W, Ku SK, Song KS and Bae JS:
Anti-inflammatory activities of oleanolic acid on HMGB1 activated
HUVECs. Food Chem Toxicol. 50:1288–1294. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Reisman SA, Aleksunes LM and Klaassen CD:
Oleanolic acid activates Nrf2 and protects from acetaminophen
hepatotoxicity via Nrf2-dependent and Nrf2-independent processes.
Biochem Pharmacol. 77:1273–1282. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yim TK, Wu WK, Pak WF and Ko KM:
Hepatoprotective action of an oleanolic acid-enriched extract of
Ligustrum lucidum fruits is mediated through an enhancement
on hepatic glutathione regeneration capacity in mice. Phytother
Res. 15:589–592. 2001.PubMed/NCBI
|
|
25
|
Hsu HY, Yang JJ and Lin CC: Effects of
oleanolic acid and ursolic acid on inhibiting tumor growth and
enhancing the recovery of hematopoietic system postirradiation in
mice. Cancer Lett. 111:7–13. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Fujiwara Y, Komohara Y, Kudo R, Tsurushima
K, Ohnishi K, Ikeda T and Takeya M: Oleanolic acid inhibits
macrophage differentiation into the M2 phenotype and glioblastoma
cell proliferation by suppressing the activation of STAT3. Oncol
Rep. 26:1533–1537. 2011.PubMed/NCBI
|
|
27
|
Wei J, Liu H, Liu M, et al: Oleanolic acid
potentiates the antitumor activity of 5-fluorouracil in pancreatic
cancer cells. Oncol Rep. 28:1339–1345. 2012.PubMed/NCBI
|
|
28
|
Bishayee A, Ahmed S, Brankov N and Perloff
M: Triterpenoids as potential agents for the chemoprevention and
therapy of breast cancer. Front Biosci. 16:980–996. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yamai H, Sawada N, Yoshida T, et al:
Triterpenes augment the inhibitory effects of anticancer drugs on
growth of human esophageal carcinoma cells in vitro and suppress
experimental metastasis in vivo. Int J Cancer. 125:952–960. 2009.
View Article : Google Scholar
|
|
30
|
White CC, Viernes H, Krejsa CM, Botta D
and Kavanagh TJ: Fluorescence-based microtiter plate assay for
glutamate-cysteine ligase activity. Anal Biochem. 318:175–180.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Frosina G: DNA repair and resistance of
gliomas to chemotherapy and radiotherapy. Mol Cancer Res.
7:989–999. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chang JY and Cox JD: Improving radiation
conformality in the treatment of non-small cell lung cancer. Semin
Radiat Oncol. 20:171–177. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kim M, Lee J, Ha B, Lee R, Lee KJ and Suh
HS: Factors predicting radiation pneumonitis in locally advanced
non-small cell lung cancer. Radiat Oncol J. 29:181–190. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zerp SF, Stoter R, Kuipers G, et al:
AT-101, a small molecule inhibitor of anti-apoptotic Bcl-2 family
members, activates the SAPK/JNK pathway and enhances
radiation-induced apoptosis. Radiat Oncol. 4:472009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Rao SK, Rao PS and Rao BN: Preliminary
investigation of the radiosensitizing activity of guduchi
(Tinospora cordifolia) in tumor-bearing mice. Phytother Res.
22:1482–1489. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Dai Y, DeSano JT, Meng Y, Ji Q, Ljungman
M, Lawrence TS and Xu L: Celastrol potentiates radiotherapy by
impairment of DNA damage processing in human prostate cancer. Int J
Radiat Oncol Biol Phys. 74:1217–1225. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tsai SJ and Yin MC: Anti-oxidative,
anti-glycative and anti-apoptotic effects of oleanolic acid in
brain of mice treated by D-galactose. Eur J Pharmacol. 689:81–88.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Koh SJ, Tak JK, Kim ST, Nam WS, Kim SY,
Park KM and Park JW: Sensitization of ionizing radiation-induced
apoptosis by ursolic acid. Free Radic Res. 46:339–345. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Eder-Czembirek C, Erovic BM, Czembirek C,
Brunner M, Selzer E, Pötter R and Thurnher D: Betulinic acid a
radiosensitizer in head and neck squamous cell carcinoma cell
lines. Strahlenther Onkol. 186:143–148. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Selzer E, Pimentel E, Wacheck V, Schlegel
W, Pehamberger H, Jansen B and Kodym R: Effects of betulinic acid
alone and in combination with irradiation in human melanoma cells.
J Invest Dermatol. 114:935–940. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yang JC, Lu MC, Lee CL, Chen GY, Lin YY,
Chang FR and Wu YC: Selective targeting of breast cancer cells
through ROS-mediated mechanisms potentiates the lethality of
paclitaxel by a novel diterpene, gelomulide K. Free Radic Biol Med.
51:641–657. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Karthikeyan S, Kanimozhi G, Prasad NR and
Mahalakshmi R: Radiosensitizing effect of ferulic acid on human
cervical carcinoma cells in vitro. Toxicol In Vitro. 25:1366–1375.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Benassi B, Zupi G and Biroccio A:
Gamma-glutamylcysteine synthetase mediates the c-Myc-dependent
response to antineoplastic agents in melanoma cells. Mol Pharmacol.
72:1015–1023. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Oliveira NG, Castro M, Rodrigues AS, et
al: Wortmannin enhances the induction of micronuclei by low and
high LET radiation. Mutagenesis. 18:37–44. 2003. View Article : Google Scholar
|
|
45
|
Sedelnikova OA, Nakamura A, Kovalchuk O,
et al: DNA double-strand breaks form in bystander cells after
microbeam irradiation of three-dimensional human tissue models.
Cancer Res. 67:4295–4302. 2007. View Article : Google Scholar
|
|
46
|
Kashino G, Prise KM, Suzuki K, et al:
Effective suppression of bystander effects by DMSO treatment of
irradiated CHO cells. J Radiat Res. 48:327–333. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Shao C, Furusawa Y, Kobayashi Y, Funayama
T and Wada S: Bystander effect induced by counted high-LET
particles in confluent human fibroblasts: a mechanistic study.
FASEB J. 17:1422–1427. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Thierens H and Vral A: The micronucleus
assay in radiation accidents. Ann Ist Super Sanita. 45:260–264.
2009.PubMed/NCBI
|
|
49
|
McConnachie LA, Mohar I, Hudson FN, et al:
Glutamate cysteine ligase modifier subunit deficiency and gender as
determinants of acetaminophen-induced hepatotoxicity in mice.
Toxicol Sci. 99:628–636. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Bai X, Qiu A, Guan J and Shi Z:
Antioxidant and protective effect of an oleanolic acid-enriched
extract of A. deliciosa root on carbon tetrachloride induced
rat liver injury. Asia Pac J Clin Nutr. 16(Suppl 1): 169–173.
2007.PubMed/NCBI
|
|
51
|
Abdel-Zaher AO, Abdel-Rahman MM, Hafez MM
and Omran FM: Role of nitric oxide and reduced glutathione in the
protective effects of aminoguanidine, gadolinium chloride and
oleanolic acid against acetaminophen-induced hepatic and renal
damage. Toxicology. 234:124–134. 2007. View Article : Google Scholar
|
|
52
|
Petronelli A, Saulle E, Pasquini L, et al:
High sensitivity of ovarian cancer cells to the synthetic
triterpenoid CDDO-imidazolide. Cancer Lett. 282:214–228. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Ikeda T, Sporn M, Honda T, Gribble GW and
Kufe D: The novel triterpenoid CDDO and its derivatives induce
apoptosis by disruption of intracellular redox balance. Cancer Res.
63:5551–5558. 2003.PubMed/NCBI
|
|
54
|
Haddad JJ, Olver RE and Land SC:
Antioxidant/pro-oxidant equilibrium regulates HIF-1alpha and
NF-kappa B redox sensitivity. Evidence for inhibition by
glutathione oxidation in alveolar epithelial cells. J Biol Chem.
275:21130–21139. 2000. View Article : Google Scholar
|