Introduction
Head and neck squamous cell carcinoma (HNSCC) is the
sixth most common cancer with over 640,000 cases diagnosed annually
and high mortality worldwide (1).
Lifestyles such as smoking, alcohol consumption, and betel nut
chewing are primary etiological factors of HNSCC and are linked to
the increased prevalence of HNSCC worldwide (2). Although clinical interventions such as
surgery, radiotherapy, chemotherapy, and chemo-radiotherapy have
rapidly advanced, 5-year survival and morbidity rates of patients
with HNSCC have not improved significantly in the last 30 years
(2,3). Furthermore, despite the advancement of
diagnostic techniques and therapeutics, clinical interventions
frequently result in irreversible loss of function and
disfigurement (4). Therefore, there
is an urgent need to develop chemotherapeutic agents that
effectively treat the cancer with fewer adverse effects.
Recently, natural compounds purified from herbal
plants used traditionally in folk and oriental medicine have
received considerable interest as a potential preventive or
clinical drugs due to their various pharmacological effects
combined with potential biological safety (5). Many recent studies have tested the
anticancer effects of natural compounds to develop chemotherapeutic
agents with fewer side-effects. Due to these studies, several
natural compounds with effective anticancer effects and strong
safety data have been used as clinical chemotherapeutic agents for
cancer patients, after approval by the United States Food and Drug
Administration (5).
Apoptosis is defined as programmed cell death
characterized by morphological shrinkage (6), nucleus condensation (7) and DNA degradation (8). Cancer cell-specific apoptosis is one
of current strategies for developing chemotherapeutic agents from
natural compounds (9).
Biochanin-A (5,7-dihydroxy-4-methoxy-isoflavone) is
a phytoestrogen, which is a natural estrogen analogue, derived from
edible and herbal plants such as peanuts, alfalfa sprouts, soy and
red clover (10). Recent studies
have reported that biochanin-A alleviates climacteric vasomotor
symptoms and reduces low-density lipoprotein levels in menopausal
women (11). Furthermore,
biochanin-A has been reported to possess various pharmacological
effects including antioxidant activity (12), anti-inflammation (13), neuroprotection (14) and the prevention of articular
cartilage degeneration (10).
Biochanin-A has also been reported to show anticancer activity such
as increased radiotoxicity in colon cancer (15), the increased survival in pancreatic
cancer, and induction of apoptosis in prostate cancer (16) and hepatoma (17).
Therefore, the aim of the present study was to
determine whether biochanin-A has the potential to function as a
chemotherapeutic agent to treat HNSCC. We evaluated the potential
apoptotic effects of biochanin-A on HNSCC and elucidated the
induced apoptotic signaling pathway.
Materials and methods
Cell culture
FaDu cells originating from human pharynx squamous
carcinoma were obtained from the American Type Culture Collection
(ATCC; Manassas, VA, USA) and cultured by following the instruction
provided by ATCC. Briefly, FaDu cells were maintained in minimum
essential medium (MEM; Gibco, Grand Island, NY, USA) containing 10%
fetal bovine serum (FBS; Gibco, Grand Island, NY, USA), 100 µg/ml
streptomycin and 100 Unit/ml penicillin (Gibco) in a humidified, 5%
CO2 and 37°C incubator.
Cell cytotoxicity assay
To determine the cytotoxicity of biochanin-A in FaDu
cells, the colorimetric MTT
(3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide)
assay was performed as previously described (2). Briefly, FaDu cells were cultured at a
density of 0.2×05 cells/ml in 96-well plates and allowed
to attach to the well overnight. After incubation, cultured FaDu
cells were treated with 25, 50 and 100 µM biochanin-A (Santa Cruz
Biotechnology, Dallas, TX, USA) for 24 and 48 h. Thereafter, 200 µl
of 5 mg/ml MTT (Sigma-Aldrich, St. Louis, MO, USA) was added into
cultured FaDu cells and incubated for another 4 h. Sequentially,
supernatant was removed, and MTT crystals were dissolved in 200
µl/well dimethyl sulfoxide (DMSO; Sigma-Aldrich). Finally, optical
density was measured at 570 nm by a spectrophotometer (Epoch;
BioTek Instruments, Inc., Winooski, VT, USA).
Live and dead cell assay
To visualize the live and dead FaDu cells following
treatment with biochanin-A, the cell live and dead assay was
performed with green calcein AM and ethidium homodimer-1 as
previously described (4). Briefly,
FaDu cells were cultured at a density of 0.2×105
cells/ml in an 8-well chamber slide (Electron Microscopy Sciences,
Hatfield, PA, USA) and allowed to attach to the well overnight.
After incubation, cultured FaDu cells were treated with 25 and 50
µM biochanin-A for 24 h. Thereafter, to visualize either live or
dead FaDu cells, live and dead cell assay was performed by live and
dead cell assay kit (Thermo Fisher Scientific, Rockford, IL, USA),
which is composed of green calcein AM to stain the live cells with
green fluorescence and ethidium homodimer-1 to stain the dead cells
with red fluorescence, following the instructions provided by the
manufacturer. Cells were imaged using fluorescence microscopy
(Eclipse TE2000; Nikon Instruments, Inc., Melville, NY, USA).
DAPI staining
To determine the nucleus condensation in FaDu cells
treated with biochanin-A, DAPI (4′,6-diamidino-2-phenylindole)
staining was performed by the protocol previously described
(18). Briefly, FaDu cells were
cultured at a density of 0.2×105 cells/ml in an 8-well
chamber slide (Electron Microscopy Sciences) and allowed to attach
to the well overnight. After incubation, cultured FaDu cells were
treated with 25 and 50 µM biochanin-A for 24 h. Thereafter, FaDu
cells were fixed with 4% paraformaldehyde after washing with
phosphate-buffered solution (PBS) and stained with 1 mg/ml DAPI for
20 min. The nucleus of FaDu cells were imaged using fluorescence
microscopy (Eclipse TE2000; Nikon Instruments).
Flow cytometric analysis
Flow cytometric analysis (FACS) was performed to
determine the extent of apoptosis and necrosis using Annexin
V-fluorescein isothiocyanate (FITC) and propidium iodide (PI),
respectively. The FaDu cells were cultured at a density of
1×105 cells/ml and allowed to attach to the well
overnight. After incubation, cultured FaDu cells were treated with
25 and 50 µM biochanin-A for 24 h. Thereafter, FaDu cells were
washed twice in PBS and re-suspended in a binding buffer (BD
Biosciences, San Diego, CA, USA). Annexin V-FITC and PI (BD
Biosciences) were added to the cells and incubated in the dark for
15 min. The cells were analyzed using a fluorescence-activated cell
sorting Calibur flow cytometer (Becton-Dickinson, San Jose, CA,
USA). Data analysis was performed using standard CellQuest software
(Becton-Dickinson) and WinMDI version 2.9 software (The Scripps
Research Institute, San Diego, CA, USA).
Immunoblotting
The FaDu cells were cultured at a density of
1×105 cells/ml and allowed to attach to the well
overnight. After incubation, cultured FaDu cells were treated with
25 and 50 µM biochanin-A for 24 h. Thereafter, FaDu cells were
harvested, lysed using cell lysis buffer (Cell Signaling
Technology, Danvers, MA, USA) containing protease inhibitor
(Sigma-Aldrich) and phosphatase inhibitor cocktails (Sigma-Aldrich)
according to the manufacturers protocol. Total protein
concentrations of cell lysates were determined by bicinchoninic
acid (BCA) protein assays (Pierce, Rockford, IL, USA). In addition,
conditioned media were harvested to detect the proteins secreted
from FaDu cells. Equal amounts of protein and conditioned media
were mixed with 5X loading buffer, boiled at 90°C for 10 min,
separated using sodium dodecyl sulfate polyacrylamide gene
electrophoresis (SDS-PAGE) and transferred onto nitrocellulose
membranes (BD Biosciences). After blocking with 5% bovine serum
albumin (BSA; Sigma-Aldrich) in TBS-T (Tris-buffered saline with
0.1% Tween-20 (Sigma-Aldrich) at room temperature for 1 h,
membranes were reacted with TBS-T containing 5% BSA and primary
antibodies of the corresponding proteins at 4°C for 12 h and then
incubated with horseradish peroxidase-conjugated secondary
antibody. The following antibodies were used: antibodies against
FasL, caspases (−3, −7, −8 and −9), Bcl-2, Bcl-xL, Bad, Poly(ADP
ribose) polymerase (PARP), phospho-NF-κB and total NF-κB were
purchased from Cell Signaling Technology. β-actin, phospho-ERK,
total ERK, phospho-p38, total p-38, phospho-Akt, total Akt, matrix
metalloproteinase (MMP)-2 and MMP-9 were purchased from Santa Cruz
Biotechnology. The immunoreactive bands were visualized using the
ECL system (Amersham Biosciences, Piscataway, NJ, USA) and were
exposed on radiographic film.
In vitro migration assay
To perform the migration assay, FaDu cells were
cultured onto 2×0.22 cm2 culture inserts (Ibidi,
Regensburg, Germany) at a density of 1×104 cells/well.
Wounds were introduced by removing the culture inserts after 24 h
of incubation. Thereafter, cultured FaDu cells were treated with 25
µM biochanin-A for 48 and 72 h. Wound widths were imaged using an
inverted microscope (Eclipse TE2000; Nikon Instruments).
Colony formation assay
A colony formation assay was performed according to
the previously described protocol (19). Briefly, FaDu cells were cultured at
a density of 200 cells/well in a 6-well culture plate and allowed
to attach to the well overnight. After incubation, cultured FaDu
cells were treated with 25 and 50 µM biochanin-A for 24 h and were
then incubated in culture medium without biochanin-A for 5 days.
Thereafter, the medium was removed and the cells were washed with
PBS and fixed with 4% paraformaldehyde for 10 min at 4°C.
Sequentially, the colonies were stained with 2% crystal violet for
10 min. Finally, the colonies were washed with PBS and dried at
room temperature, before being imaged by a digital camera (Nikon
Instruments).
Gelatin zymography
Gelatin zymography was performed to assess the
activity of matrix metalloproteinases (MMPs) secreted from FaDu
cells treated with biochanin-A. An equal volume of conditioned
media was mixed with non-reducing sample buffer [4% SDS, 0.15 M
Tris (pH 6.8) and 20% (v/v) glycerol containing 0.05% (w/v)
bromophenol blue] and resolved on a 10% polyacrylamide gel
containing copolymerized 0.2% (1 mg/ml) swine skin gelatin. After
electrophoresis of the conditioned media samples, gels were washed
with cold PBS containing 2.5% (v/v) Triton X-100 for 30 min and
washed twice with cold PBS for 15 min. After washing, gels were
incubated in the zymogram renaturing buffer [50 mM Tris-HCl (pH
7.6), 10 mM CaCl2, 50 mM NaCl and 0.05% Brij-35) at 37°C
for 72 h. After renaturation of MMPs, gels were stained with 0.1%
Coomassie brilliant blue R250. The gelatinolytic activity was
revealed as a clear band on a background of uniform light blue
staining.
Statistical analysis
Data are reported as the mean ± SD of three
individual experiments performed. Statistical analysis was carried
out using the Student's t-test and a P-value <0.05 was
considered to indicate a statistically significant result.
Results
Biochanin-A induces death of FaDu
cells via increased cytotoxicity
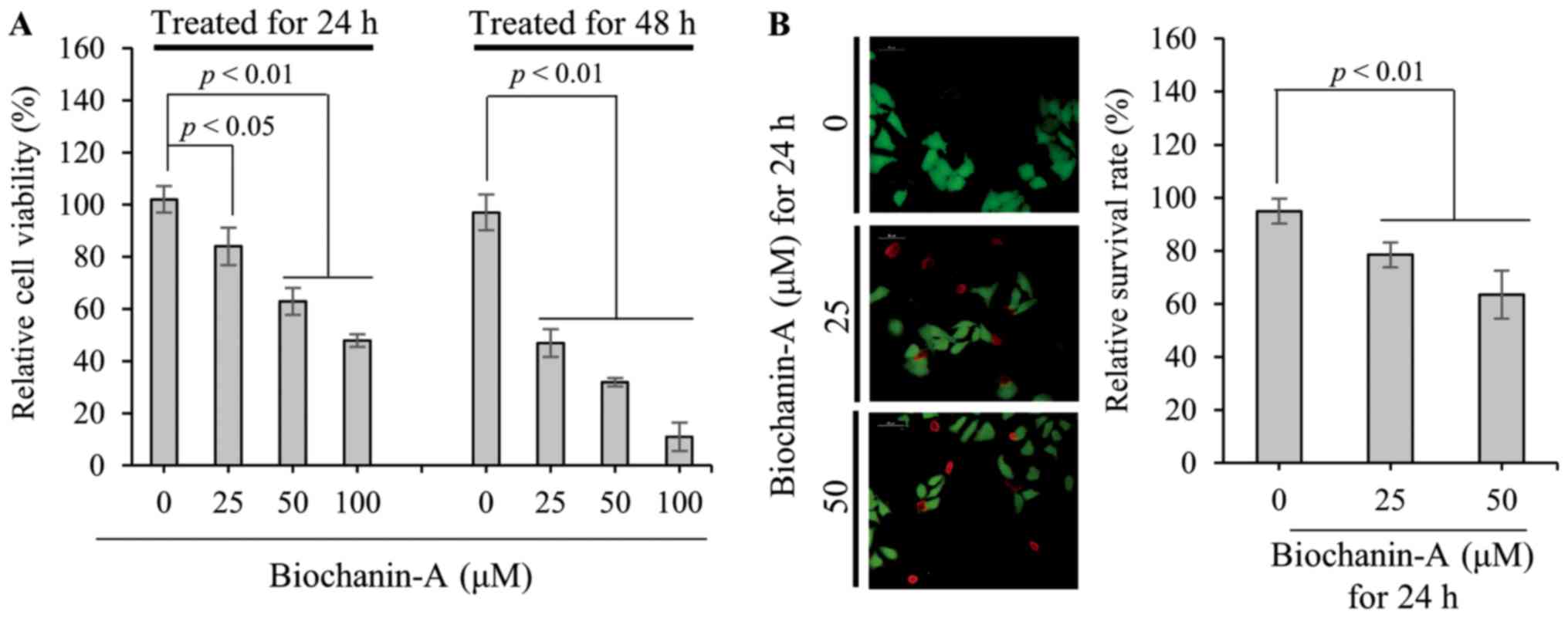
To measure the viability of FaDu cells treated with
25, 50 and 100 µM biochanin-A for 24 and 48 h, MTT assays were
performed. As shown in Fig. 1A,
relative viabilities of FaDu cells treated with 25, 50 and 100 µM
biochanin-A for 24 h are 84±7.2, 63±5.15 and 48±2.4%, respectively,
compared to non-treated control (102±5.1%). Furthermore, relative
viabilities of FaDu cells treated with the same concentrations of
biochanin-A for 48 h are 47±5.3, 32±1.6 and 11±5.5%, respectively.
Live and dead cell assays were performed to visualize the live and
dead cells after treatment with 25 and 50 µM biochanin-A for 24 h.
In the non-treated control, as shown in Fig. 1B, almost all cells are stained
fluorescent green by membrane permeable calcein AM, which is
cleaved by cytosolic esterase in living cells. In contrast, the
number of dead cells stained fluorescent red by ethidium
homodimer-1 increases with biochanin-A treatment in a
dose-dependent manner. The relative rate of live FaDu cells after
treatment with 25 and 50 µM biochanin-A were measured as 78.5±4.7
and 63.5±9%, respectively. These results consistently demonstrate
that biochanin-A suppresses the viability of FaDu cells through an
increase in cell cytotoxicity, in a dose- and time-dependent
manner.
Biochanin-A induced FaDu cell death is
mediated by apoptosis
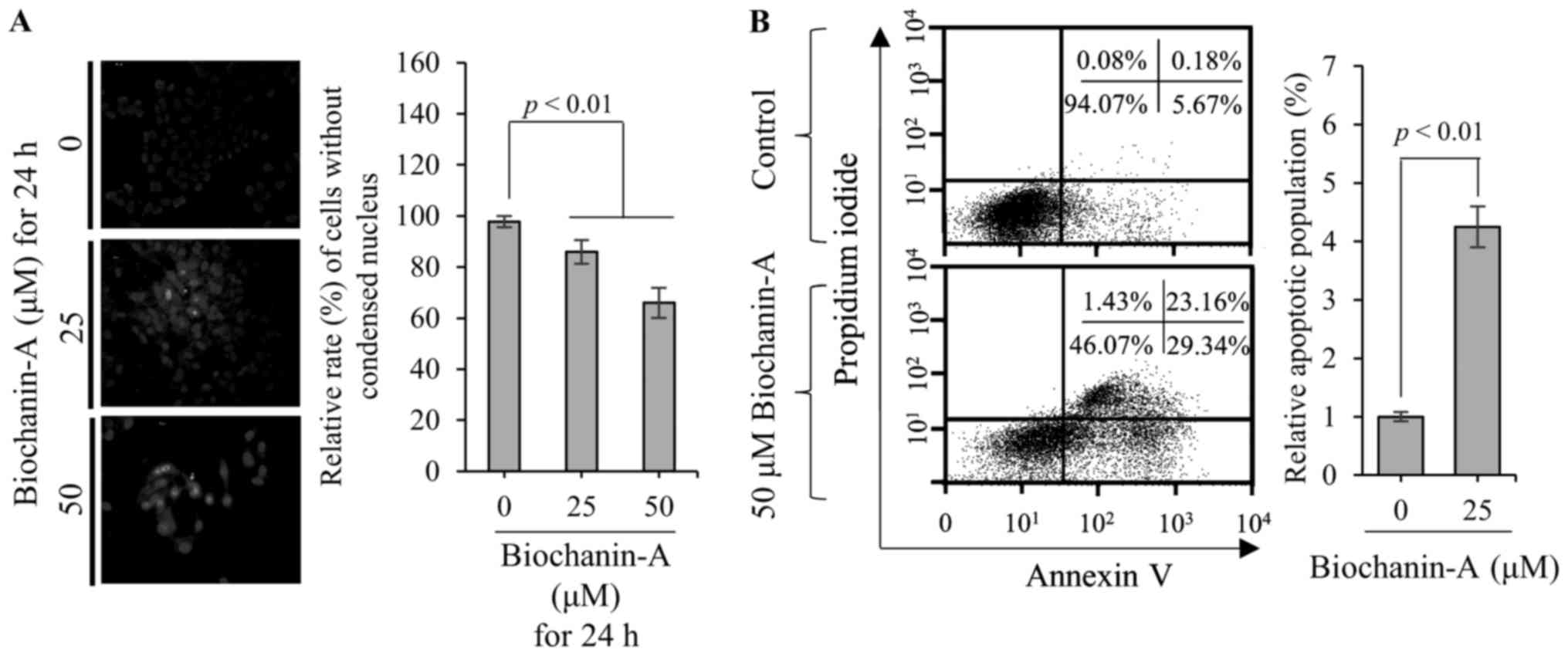
Nucleus condensation is a typical phenomenon of
apoptosis (7). Therefore, DAPI
staining was performed to observe the nucleus morphology of FaDu
cells treated with biochanin-A. As shown in Fig. 2A, the relative rate of FaDu cells
without condensed nucleus is ~85.9±4.7 and 66.0±5.9% at 25 and 50
µM biochanin-A compared with non-treated control, respectively.
These data show that biochanin-A increased the number of FaDu cells
with condensed nuclei in a dose-dependent manner. Therefore, to
verify the biochanin-A-induced apoptosis of FaDu cells, FACS
analysis was performed using Annexin V-FITC and PI. As shown in
Fig. 2B, the apoptotic population
of FaDu cells treated with 50 µM biochanin-A increases ~4.2-fold
compared with than non-treated control. Taken together, these data
consistently indicate that biochanin-A-induced FaDu cell death is
mediated by apoptosis.
Biochanin-A induces the apoptosis of
FaDu cell through extrinsic and intrinsic apoptotic pathways
Next, to verify biochanin-A-induced apoptosis in
FaDu cells, the alteration in expression of pro- and anti-apoptotic
factors were observed by western blot analysis. As shown in
Fig. 3, the expression of death
ligand FasL (48 kDa) increases in FaDu cells treated with 25 and 50
µM biochanin-A in a dose-dependent manner. Sequentially, the
expression of full length caspase-8 (57 kDa), which is a
down-stream target molecule of FasL and is part of the death
receptor mediated extrinsic apoptotic signaling pathway, gradually
decreased in the FaDu cells treated with biochanin-A. In contrast,
the expression of cleaved caspase-8 (43 kDa) increased due to the
cleavage of full length caspase-8 by FasL in the FaDu cells treated
with biochanin-A. Furthermore, the expression of cleaved caspase-3
increased due to the cleavage of full length caspase-3 (35 kDa) by
the cleaved caspase-8. Sequentially, the expression of cleaved PARP
(89 kDa; full length PARP: 116 kDa) increased by cleaved caspase-3
and then induced apoptosis. Therefore, these data indicate that the
biochanin-A induces cells death through the
FasL-caspase-8-caspase-3-PARP axis mediated extrinsic apoptosis
signaling pathway in FaDu cells.
Furthermore, the expressions of Bcl-2 (26 kDa) and
Bcl-xL (26 kDa), anti-apoptotic factors associated with the
mitochondria-dependent apoptotic signaling pathway, decreased in
the FaDu cells treated with 25 and 50 µM biochanin-A in a
dose-dependent manner. Whereas, biochanin-A increased the
expression of Bad (20 kDa) and cleaved caspase-9 (35 kDa; full
length caspase-9: 47 kDa), pro-apoptotic factors associated with
the mitochondria-dependent apoptotic signaling pathway. The cleaved
caspase-9 increased the amount of cleaved caspase-3 and cleaved
PARP and then induced the apoptosis of FaDu cells treated with
biochanin-A. Therefore, these data indicate that biochanin-A
induces cell death via the mitochondria-dependent intrinsic
apoptosis signaling pathway in FaDu cells.
Biochanin-A suppresses the migration
and proliferation of FaDu cells through the downregulation and
inactivation of MMP-2 and −9
FaDu cells were treated with 25 µM biochanin-A for
48 and 72 h to verify whether biochanin-A can suppress the
migration and invasion. As shown in Fig. 4A, biochanin-A effectively suppressed
migration compared with non-treated control. In addition, to verify
whether biochanin-A suppress the proliferation of FaDu cells, a
colony formation assay was performed as shown in Fig. 4B. The number of colonies in the
untreated control was 168±42. In contrast, the number of colonies
was 55±6.6 in the FaDu cells treated with 25 µM biochanin-A for 5
days. These data indicate that biochanin-A effectively suppressed
the migration and proliferation of FaDu cells. In order to verify
the expression alteration of implicating factors associated with
the migration and proliferation of cancer cells, western blot
analysis and gelatin zymography were performed as shown as Fig. 4C and D, respectively. The
expressions of MMP-2 and MMP-9 gradually decreased in the FaDu
cells treated with 25 and 50 µM biochanin-A in a dose-dependent
manner. Clear bands formed on the gelatin zymogram gels by active
MMPs gradually decreased in the FaDu cells treated with 25 and 50
µM biochanin-A in a dose-dependent manner. These data consistently
indicate that biochanin-A suppresses the migration and invasion of
FaDu cells through the downregulation expression and activation of
MMPs. Taken together, these data suggest that the biochanin-A may
act as a potential anti-metastatic reagent in HNSCC.
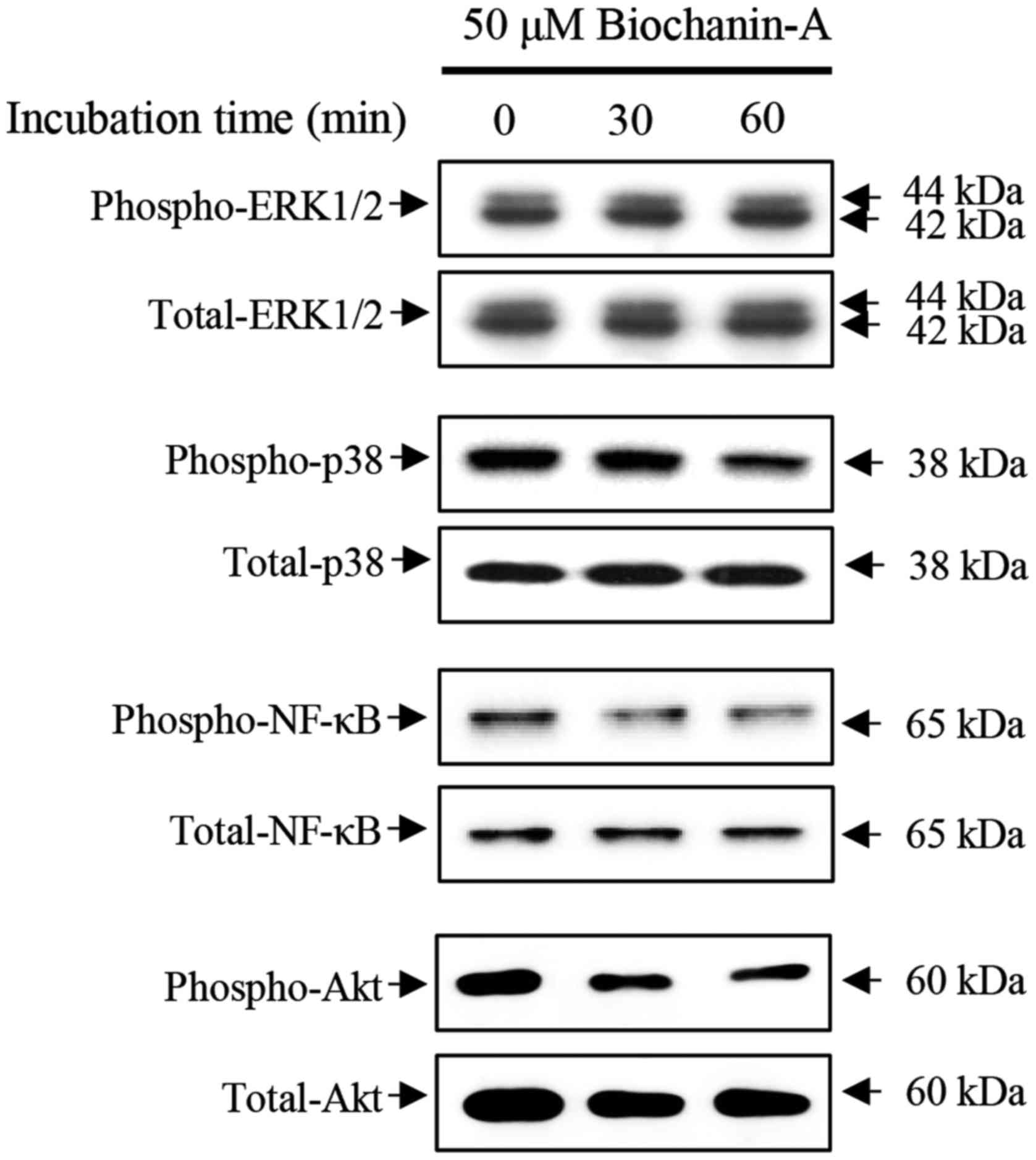
To verify the cellular signaling associated with
migration and proliferation of FaDu cells, alterations in ERK1/2,
p38 and NF-κB expression were observed by immunoblotting in FaDu
cells treated with 50 µM biochanin-A for 15, 30 and 60 min as shown
in Fig. 5. Biochanin-A did not
induce the phosphorylation of ERK1/2 in FaDu cells at the defined
treatment time, whereas, the phosphorylation of p38 and NF-κB
decreased in the FaDu cells treated with biochanin-A in a
time-dependent manner compared with non-treated control.
Furthermore, the phosphorylation of Akt, which is closely
associated with cellular signaling of cell proliferation,
significantly decreased in FaDu cells treated with biochanin-A, in
a time-dependent manner compared with non-treated control.
Therefore, these data suggest that biochanin-A induced suppression
of migration and proliferation is associated with the alteration of
p38 MAPK, NF-κB and PI3K/Akt cellular signaling pathways in FaDu
cells.
Discussion
Oral cancer is the most common type of head and neck
cancer and head and neck squamous cell carcinomas worldwide
(20). Despite therapeutic and
technological advances, the 5-year survival rate of oral cancer has
not increased (21). Furthermore,
the clinical interventions for patients with oral cancer have
multiple serious side-effects such as the loss of function of
speech, breathing, mastication and swallowing (22). Therefore, to reduce the side-effects
caused by clinical interventions to treat oral cancer, the
development of chemotherapy using natural compounds that possess
anticancer properties and excellent biological safety is
critical.
Phytoestrogens are plant-derived compounds that are
structurally and physiologically similar to estrogen. Recent
studies have reported that the phytoestrogens such as coumestrol,
kaempferol and genistein induce apoptosis in various cancers
including breast, prostate and hepatocellular carcinoma (23–25).
Biochanin-A is a phytoestrogen derived from red
clover and used as a folk medicine worldwide. In the present study,
we demonstrated that biochanin-A induced anticancer activities
including the suppression of cellular proliferation and migration,
and the increase in apoptosis through the induction of caspase
activation in FaDu head and neck squamous cell carcinoma.
In our previous study, we reported that biochanin-A
did not affect the viability of primary chondrocytes isolated from
the articular cartilage of rats for 21 days (10). However, the present study showed
that biochanin-A increased cell death through the increase of
cytotoxicity in FaDu cells, as shown in Fig. 1. Furthermore, recent studies have
reported that biochanin-A increased the cytotoxicity of pancreatic
(26) and prostate cancer (16). Therefore, these studies suggest that
biochanin-A is a phytoestrogen that induce specific cancer cell
death.
DNA fragmentation caused by nucleus condensation is
a typical phenomenon of apoptosis, which is a programmed cell
death. As shown in Fig. 2A, the
number of FaDu cells with condensed nucleus was increased by
biochanin-A in a dose-dependent manner. Therefore, these data
indicate that biochanin-A-induced FaDu cell death may involve
apoptosis. In order to verify this, FACS analysis using Annexin V
and PI staining was performed, as shown in Fig. 2B. At the early stage of apoptosis,
membrane phosphatidylserine is translocated to the cell surface
from the inner plasma membrane. Therefore, Annexin V, which is a
Ca2+-dependent phospholipid-binding protein with binding
activity to phosphatidylserine, was used as a marker to detecting
the early stage of apoptosis (21).
Furthermore, translocated membrane phosphatidylserine precedes the
loss of membrane integration. Subsequently, PI without membrane
permeability binds to DNA by intercalating between the bases
through the membranes of dead and damaged cells in the late stage
of apoptosis (21). In this study,
the results of FACS analysis showed that the biochanin-A increased
the apoptotic population of FaDu cells in both early and late stage
compared with non-treated control. Taken together, these data
consistently indicate that biochanin-A induces apoptosis in FaDu
cells.
Apoptosis is generally divided into the death
receptor mediated extrinsic pathway and mitochondrial-dependent
intrinsic pathway (27,28). The death receptor mediated extrinsic
pathways are triggered by death ligands such as FasL and
TNF-related apoptosis-inducing ligand (TRAIL), and are sequentially
mediated by the activation of caspase-8, caspase-3, and PARP to
induce DNA fragmentation (27,28).
However, the mitochondria-dependent intrinsic pathway associated
with apoptosis is triggered by growth hormone withdrawal, DNA
damage by UV- or γ-radiation, chemotherapeutic drugs, and the
activation of caspase-8 associated death receptor mediated
extrinsic apoptosis through the upregulation or activation of
pro-apoptotic factors such as Bad, Bid, Bax and caspase-9 and the
downregulation of anti-apoptotic factors such as Bcl-2 and Bcl-xL
(27,28). Finally, activated caspase-9 induces
the activation of caspase-3 and PARP to induce DNA fragmentation
(27,28). In the present study, biochanin-A
increased the expression of FasL and induced the activation of its
downstream pro-apoptotic factor caspase-8 as shown in Fig. 3. Furthermore, the expression of
anti-apoptotic factors such as Bcl-2 and Bcl-xL dose-dependently
decreased in the FaDu cells treated with biochanin-A. Whereas,
biochanin-A increased the expression of Bad and activated caspase-9
in FaDu cells. Finally, both activated caspase-8 and caspase-9
increased the activation of caspase-3 and PARP to induce the
apoptosis of FaDu cells treated with biochanin-A. Therefore, these
data indicate that biochanin-A-induced FaDu cell death is mediated
by both death receptor mediated extrinsic and mitochondria
dependent intrinsic apoptosis pathways.
Next, in the present study, we demonstrated that the
migration and proliferation of FaDu cells were significantly
decelerated by biochanin-A as shown in Fig. 4A and B. Cancer cells metastasize to
other tissues through bloodstream and lymphatic system in the
terminal stage of cancer (29).
Hence, the deceleration of cancer cell growth and the suppression
of metastasis are crucial aims of clinical treatment for metastatic
cancer (30). Furthermore, the
expressional upregulation and activation of matrix
metalloproteinase such as MMP-2 and MMP-9 are crucial factors
associated with the migration and proliferation of cancer cells
(29,31). We demonstrated that the expression
and activation of MMP-2 and MMP-9 significantly decreased in the
FaDu cells treated with biochanin-A, as shown in Fig. 4C and D. Taken together, biochanin-A
decelerates the migration and proliferation of FaDu cells through
the expression downregulation and inactivation of MMP-2 and
MMP-9.
In the present study, biochanin-A suppressed the
phosphorylation of p38 MAPK and NF-κB in FaDu cells as shown in
Fig. 5. As well as this study, Kole
et al (32), reported that
biochanin-A-induced anti-proliferation through the inhibition of
p38 MAPK and the blocking of NF-κB nuclear translocation. Recent
studies have also reported that the expression of MMP-2 and MMP-9,
associated with cell growth, proliferation and migration, are
closely regulated by MAPK cellular signaling in various cancer
cells including colon cancer (33)
and osteosarcoma (34).
Furthermore, Bindhu et al (35) and Qin et al (36) reported that the activation of MMP-2
is closely regulated by NF-κB cellular signaling in oral squamous
cell carcinoma and breast cancer. These consistently indicate that
biochanin-A-induced anti-proliferation and anti-migration are
mediated by the expressional downregulation and inactivation of
MMP-2 and MMP-9 through the suppression of p38 MAPK and NF-κB
cellular signaling pathways in FaDu cells. In addition, PI3K/Akt
signaling is closely associated with cell proliferation, survival,
growth and metastasis (37,38). Recent studies have reported that the
suppression of PI3K/Akt signaling enhances cytotoxicity through the
inhibition of the mTOR signaling pathway in various cancers such as
breast and colorectal cancer (39–41).
In the present study, we demonstrated that biochanin-A suppressed
the phosphorylation of Akt in FaDu cells. Taken together, these
studies indicate that biochanin-A-induced anticancer activities
including apoptosis, anti-proliferation, and anti-migration may be
mediated by the suppression of PI3K/Akt cellular signaling pathway
in FaDu cells.
In conclusion, we demonstrated that
biochanin-A-induced anti-apoptotic effects such as death receptor
mediated extrinsic and mitochondria-dependent intrinsic apoptosis,
anti-metastasis and anti-proliferation in FaDu cells. These
findings suggest that biochanin-A may be a promising natural
chemotherapy candidate for HNSCC treatment.
Acknowledgements
The present study was supported by a research fund
from the Chosun University Dental Hospital, 2016.
References
|
1
|
Lacko M, Braakhuis BJ, Sturgis EM,
Boedeker CC, Suárez C, Rinaldo A, Ferlito A and Takes RP: Genetic
susceptibility to head and neck squamous cell carcinoma. Int J
Radiat Oncol Biol Phys. 89:38–48. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Park MR, Kim SG, Cho IA, Oh D, Kang KR,
Lee SY, Moon SM, Cho SS, Yoon G, Kim CS, et al: Licochalcone-A
induces intrinsic and extrinsic apoptosis via ERK1/2 and p38
phosphorylation-mediated TRAIL expression in head and neck squamous
carcinoma FaDu cells. Food Chem Toxicol. 77:34–43. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Vigneswaran N, Wu J, Song A, Annapragada A
and Zacharias W: Hypoxia-induced autophagic response is associated
with aggressive phenotype and elevated incidence of metastasis in
orthotopic immunocompetent murine models of head and neck squamous
cell carcinomas (HNSCC). Exp Mol Pathol. 90:215–225. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kim JS, Oh D, Yim MJ, Park JJ, Kang KR,
Cho IA, Moon SM, Oh JS, You JS, Kim CS, et al: Berberine induces
FasL-related apoptosis through p38 activation in KB human oral
cancer cells. Oncol Rep. 33:1775–1782. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kinghorn AD, Pan L, Fletcher JN and Chai
H: The relevance of higher plants in lead compound discovery
programs. J Nat Prod. 74:1539–1555. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
McCarthy JV and Cotter TG: Cell shrinkage
and apoptosis: A role for potassium and sodium ion efflux. Cell
Death Differ. 4:756–770. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fang M, Zhang HQ and Xue SB: Apoptosis of
HL-60 cells induced by Harringtonine: Membrane blebs, nucleus blebs
and chromatin condensation. Shi Yan Sheng Wu Xue Bao. 29:221–233.
1996.(In Chinese). PubMed/NCBI
|
|
8
|
Nagata S, Nagase H, Kawane K, Mukae N and
Fukuyama H: Degradation of chromosomal DNA during apoptosis. Cell
Death Differ. 10:108–116. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fesik SW: Promoting apoptosis as a
strategy for cancer drug discovery. Nat Rev Cancer. 5:876–885.
2005. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Oh JS, Cho IA, Kang KR, You JS, Yu SJ, Lee
GJ, Seo YS, Kim CS, Kim K, Kim SG, et al: Biochanin-A antagonizes
the interleukin-1β-induced catabolic inflammation through the
modulation of NF-κB cellular signaling in primary rat chondrocytes.
Biochem Biophys Res Commun. 477:723–730. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Booth NL, Piersen CE, Banuvar S, Geller
SE, Shulman LP and Farnsworth NR: Clinical studies of red clover
(Trifolium pratense) dietary supplements in menopause: A literature
review. Menopause. 13:251–264. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Guo Q, Rimbach G, Moini H, Weber S and
Packer L: ESR and cell culture studies on free radical-scavenging
and antioxidant activities of isoflavonoids. Toxicology.
179:171–180. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ming X, Ding M, Zhai B, Xiao L, Piao T and
Liu M: Biochanin A inhibits lipopolysaccharide-induced inflammation
in human umbilical vein endothelial cells. Life Sci. 136:36–41.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wu WY, Wu YY, Huang H, He C, Li WZ, Wang
HL, Chen HQ and Yin YY: Biochanin A attenuates LPS-induced
pro-inflammatory responses and inhibits the activation of the MAPK
pathway in BV2 microglial cells. Int J Mol Med. 35:391–398. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Puthli A, Tiwari R and Mishra KP:
Biochanin A enhances the radiotoxicity in colon tumor cells in
vitro. J Environ Pathol Toxicol Oncol. 32:189–203. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Szliszka E, Czuba ZP, Mertas A, Paradysz A
and Krol W: The dietary isoflavone biochanin-A sensitizes prostate
cancer cells to TRAIL-induced apoptosis. Urol Oncol. 31:331–342.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Su SJ, Chow NH, Kung ML, Hung TC and Chang
KL: Effects of soy isoflavones on apoptosis induction and G2-M
arrest in human hepatoma cells involvement of caspase-3 activation,
Bcl-2 and Bcl-XL downregulation, and Cdc2 kinase activity. Nutr
Cancer. 45:113–123. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kim JS, Park MR, Lee SY, Kim DK, Moon SM,
Kim CS, Cho SS, Yoon G, Im HJ, You JS, et al: Licochalcone A
induces apoptosis in KB human oral cancer cells via a
caspase-dependent FasL signaling pathway. Oncol Rep. 31:755–762.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang Z, Li HM, Zhou C, Li Q, Ma L, Zhang
Z, Sun Y, Wang L, Zhang X, Zhu B, et al: Non-benzoquinone
geldanamycin analogs trigger various forms of death in human breast
cancer cells. J Exp Clin Cancer Res. 35:1492016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kim SM: Human papilloma virus in oral
cancer. J Korean Assoc Oral Maxillofac Surg. 42:327–336. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Seo YS, Yim MJ, Kim BH, Kang KR, Lee SY,
Oh JS, You JS, Kim SG, Yu SJ, Lee GJ, et al: Berberine-induced
anticancer activities in FaDu head and neck squamous cell carcinoma
cells. Oncol Rep. 34:3025–3034. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Howren MB, Christensen AJ, Karnell L
Hynds, Van Liew JR and Funk GF: Influence of pretreatment social
support on health-related quality of life in head and neck cancer
survivors: Results from a prospective study. Head Neck. 35:779–787.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zafar A, Singh S and Naseem I: Cytotoxic
activity of soy phytoestrogen coumestrol against human breast
cancer MCF-7 cells: Insights into the molecular mechanism. Food
Chem Toxicol. 99:149–161. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lee GA, Choi KC and Hwang KA: Kaempferol,
a phytoestrogen, suppressed triclosan-induced
epithelial-mesenchymal transition and metastatic-related behaviors
of MCF-7 breast cancer cells. Environ Toxicol Pharmacol. 49:48–57.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Roh T, Kim SW, Moon SH and Nam MJ:
Genistein induces apoptosis by down-regulating thioredoxin-1 in
human hepatocellular carcinoma SNU-449 cells. Food Chem Toxicol.
97:127–134. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bhardwaj V, Tadinada SM, Jain A, Sehdev V,
Daniels CK, Lai JC and Bhushan A: Biochanin A reduces pancreatic
cancer survival and progression. Anticancer Drugs. 25:296–302.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tsuruo T, Naito M, Tomida A, Fujita N,
Mashima T, Sakamoto H and Haga N: Molecular targeting therapy of
cancer: Drug resistance, apoptosis and survival signal. Cancer Sci.
94:15–21. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hassan M, Watari H, AbuAlmaaty A, Ohba Y
and Sakuragi N: Apoptosis and molecular targeting therapy in
cancer. BioMed Res Int. 2014:1508452014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Stec R, Bodnar L, Smoter M, Mączewski M
and Szczylik C: Metastatic colorectal cancer in the elderly: An
overview of the systemic treatment modalities (Review). Oncol Lett.
2:3–11. 2011.PubMed/NCBI
|
|
30
|
Hsieh MJ, Chen JC, Yang WE, Chien SY, Chen
MK, Lo YS, Hsi YT, Chuang YC, Lin CC and Yang SF:
Dehydroandrographolide inhibits oral cancer cell migration and
invasion through NF-κB-, AP-1-, and SP-1-modulated matrix
metalloproteinase-2 inhibition. Biochem Pharmacol. 130:10–20. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Huang C, Jacobson K and Schaller MD: MAP
kinases and cell migration. J Cell Sci. 117:4619–4628. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kole L, Giri B, Manna SK, Pal B and Ghosh
S: Biochanin-A, an isoflavon, showed anti-proliferative and
anti-inflammatory activities through the inhibition of iNOS
expression, p38-MAPK and ATF-2 phosphorylation and blocking NF-κB
nuclear translocation. Eur J Pharmacol. 653:8–15. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hsu HH, Liu CJ, Shen CY, Chen YJ, Chen LM,
Kuo WH, Lin YM, Chen RJ, Tsai CH, Tsai FJ, et al: p38α MAPK
mediates 17β-estradiol inhibition of MMP-2 and −9 expression and
cell migration in human lovo colon cancer cells. J Cell Physiol.
227:3648–3660. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Fromigué O, Hamidouche Z and Marie PJ:
Blockade of the RhoA-JNK-c-Jun-MMP2 cascade by atorvastatin reduces
osteosarcoma cell invasion. J Biol Chem. 283:30549–30556. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bindhu OS, Ramadas K, Sebastian P and
Pillai MR: High expression levels of nuclear factor kappa B and
gelatinases in the tumorigenesis of oral squamous cell carcinoma.
Head Neck. 28:916–925. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Qin L, Liao L, Redmond A, Young L, Yuan Y,
Chen H, OMalley BW and Xu J: The AIB1 oncogene promotes breast
cancer metastasis by activation of PEA3-mediated matrix
metalloproteinase 2 (MMP2) and MMP9 expression. Mol Cell Biol.
28:5937–5950. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tang H and Xue G: Major physiological
signaling pathways in the regulation of cell proliferation and
survival. Handb Exp Pharmacol. Feb 24–2017.https://doi.org/10.1007/164_2017_4 View Article : Google Scholar
|
|
38
|
Xu Q, Xu HX, Li JP, Wang S, Fu Z, Jia J,
Wang L, Zhu ZF, Lu R and Yao Z: Growth differentiation factor 15
induces growth and metastasis of human liver cancer stem-like cells
via AKT/GSK-3β/β-catenin signaling. Oncotarget. 8:16972–16987.
2017.PubMed/NCBI
|
|
39
|
Tseng HS, Wang YF, Tzeng YM, Chen DR, Liao
YF, Chiu HY and Hsieh WT: Aloe-emodin enhances tamoxifen
cytotoxicity by suppressing Ras/ERK and PI3K/mTOR in breast cancer
cells. Am J Chin Med. 45:337–350. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Bahrami A, Khazaei M, Hasanzadeh M, Sales
S Shahid, Mashhad M Joudi, Farazestanian M, Sadeghnia HR, Rezayi M,
Maftouh M, Hassanian SM, et al: Therapeutic potential of targeting
PI3K/AKT pathway in treatment of colorectal cancer: Rational and
progress. J Cell Biochem. Feb 23–2017.(Epub ahead of print). doi:
10.1002/jcb.25950. View Article : Google Scholar
|
|
41
|
Lux MP, Fasching PA, Schrauder MG, Hein A,
Jud SM, Rauh C and Beckmann MW: The PI3K Pathway: Background and
treatment approaches. Breast Care (Basel). 11:398–404. 2016.
View Article : Google Scholar : PubMed/NCBI
|