Introduction
Glioma is among the most common primary brain tumors
in adults (1) and is treated by a
combination of surgical resection, temozolomide chemotherapy and
radiotherapy (2). However, the
median survival is not more than 14.6 months (2,3) and
the 5-year survival rate after diagnosis is only 0.05–4.7%
(4). Two obstacles for
chemotherapeutic agents in glioma treatment are penetration through
the blood-brain barrier and the development of drug resistance
(5,6).
Paeoniflorin (PF) is the main component of
Paeoniae radix, which is widely used in traditional Chinese
medicine (7–12). PF has been shown to repair injury
(9), provide neuroprotection
(10), and exert anti-inflammatory
(11,12) and immunomodulatory (13,14)
effects. In addition, in vitro and in vivo studies
have reported that PF inhibits tumor growth, invasion and
metastasis (15–17). PF suppresses glioma cell
proliferation by upregulating microRNA-16 and inhibiting matrix
metalloproteinase (MMP)9 (18) and
suppresses glioma growth and proliferation by promoting the
degradation of signal transducer and activator of transcription
(STAT)3 (19). However, the precise
mechanism underlying the anticancer activity of PF in glioma cells
is unclear.
S-phase kinase-associated protein (Skp)2 is a
component of Skp2-Skp1/Cullin-1/F-box-protein (SCF) complex that
functions as an E3 ligase in protein ubiquitination and
degradation. Skp2 has been linked to tumor development (20–23),
playing an oncogenic role by promoting the degradation of target
proteins such as P21 (24), P27
(25) and P57 (26). Skp2 overexpression is correlated
with poor prognosis in many types of human cancers, including
hepatocellular carcinoma (27),
breast cancer (28), melanoma
(29) and glioma (30); moreover, Skp2 has been shown to
regulate tumor cell proliferation, invasion, migration, senescence,
glycolysis, and the Warburg effect (31,32) as
well as the self-renewal capacity and functioning of cancer stem
cells (33). Protein kinase B (AKT)
modulates the phosphorylation of Skp2, leading to enhancement of
cell proliferation and tumor progression (34), while suppression of Skp2 blocks
tumor progression by promoting cellular senescence (32). Skp2-SCF E3 ligase may also promote
the ubiquitination of AKT to induce tumorigenesis (31). These findings suggest that Skp2 is a
potential therapeutic target for glioma treatment.
To evaluate this possibility, the present study
investigated the biological effects of PF on glioma cell growth,
apoptosis, migration and invasion and examined whether Skp2
mediates the antitumor effects of PF. We found that Skp2 plays an
important role in glioma development. PF treatment inhibited Skp2
expression, leading to upregulation of P21 and downregulation of
phosphorylated (p-)AKT, which in turn blocked tumor progression.
These results indicate that PF is a potentially effective agent for
the treatment of glioma.
Materials and methods
Cell culture and reagents
The Second Affiliated Hospital of Soochow University
Institutional Animal Care and Use Committee approved this study.
U87 and U251 human glioma cell lines were obtained from the Chinese
Academy of Medical Sciences (Beijing, China). Cells were cultured
in Dulbeccos modified Eagles medium (DMEM; HyClone Laboratories,
Logan, UT, USA) supplemented with 10% fetal bovine serum (FBS) in a
5% CO2 atmosphere at 37°C. Primary antibodies against
Skp2, P21, P27, p-AKT (S473), AKT, extracellular signal-regulated
kinase (ERK), p-ERK, cyclin D, p-cyclin D, cleaved caspase-3 and
−9, MMP2, MMP9 and β-actin were purchased from Cell Signaling
Technology (Danvers, MA, USA). Secondary antibodies were from
Liankebio (Hangzhou, China). Lipofectamine 3000 was from Invitrogen
(Carlsbad, CA, USA). PF was from Tianjin Shilan Technology Co.,
Ltd., (Tianjin, China) and had a purity of 98%. PF was diluted in
DMEM at a stock concentration of 400 mM. Cell Counting Kit-8
(CCK-8) was obtained from Dojindo Laboratories (Kumamoto,
Japan).
Cell viability assay
U87 and U251 cells (5×103) were seeded in
a 96-well plate. Cells were treated with different concentrations
of PF for 24 and 48 h. At the end of the treatment period, 10 µl of
CCK-8 were added to each well. The plates were incubated at 37°C
for 1 h and the optical density at 450 nm was then determined on a
Varioskan microplate reader (Thermo Fisher Scientific, Waltham, MA,
USA).
Cell apoptosis analysis
U87 and U251 cells were seeded in a 6-well plate
(1–1.5×105/well) and treated with 15 and 20 mM PF for 24
h. The Annexin V-phycoerythrin (PE)/7-aminoactinomycin D (7-AAD)
assay was performed using a kit (BD Biosciences, Franklin Lakes,
NJ, USA) according to the manufacturers instructions. Briefly,
cells were washed twice with cold phosphate-buffered saline (PBS),
resuspended in 100 µl binding buffer with 7-AAD and PE-conjugated
anti-Annexin V antibody, and incubated for 15 min at room
temperature in the dark before analysis by flow cytometry (Beckman
Coulter Inc., Brea, CA, USA).
Cell cycle analysis
U87 and U251 cells were seeded in a 6-well plate
(1–1.5×105/well) and treated with 15 or 20 mM PF for 24
h. The cells were harvested and fixed overnight at 4°C with cold
70% ethanol. Cell pellets were resuspended in PBS at a
concentration of 1×106 cells/ml and then incubated with
propidium iodide (PI)/RNAase staining buffer (BD Biosciences) at
room temperature for 15 min. DNA content was determined by flow
cytometry.
Cell migration and invasion assay
To assess the effect of PF on cell migration, U87
and U251 cells were seeded in the upper chamber of a Boyden chamber
(Corning Inc., Corning, NY, USA) with 200 µl of DMEM supplemented
with 1% FBS; 600 µl complete medium with 20% serum were added to
the lower chamber along with indicated concentration of PF. After
24 h, cells remaining in the upper chamber were removed using
cotton swabs, and those in the lower chamber were fixed with
methanol, stained with 0.1% crystal violet and photographed under a
microscope. The invasion assay was similarly performed with a
Matrigel-coated Transwell chamber (BD Biosciences). Cells in three
randomly selected fields were counted.
Transfection
Cells were seeded in a 6-well plate at a density of
1.5–2×105/well. When they reached 80% confluence, the
cells were transfected with Skp2 cDNA or short interfering (si)RNA
(sense, 5-GGAGUGACAAAGACUUUGUTT-3 and antisense,
5-ACAAAGUCUUUGUCACUCCTT-3) or empty vector (Shanghai GenePharma,
Co., Ltd., Shanghai, China) using Lipofectamine 3000. At 48 h
post-transfection, the cells were collected for analysis of cell
proliferation, migration and invasion and for western blotting.
Quantitative real-time PCR
analysis
Total RNA was extracted with TRIzol reagent
(Invitrogen) and reverse transcribed into cDNA using the
PrimeScript II First Strand cDNA Synthesis kit (Takara Bio, Shiga
Japan). PCR was performed using the SYBR Premix Ex Taq kit (Takara
Bio) and the following forward and reverse primers: Skp2,
5-GAAGGGAGTCCCATGAAACA-3 and 5-GCTGAAGAGCAAGGGAGTG-3; and
glyceraldehyde 3-phosphate dehydrogenase, 5-ACCCAGAAGACTGTGGATGG-3
and 5-CAGTGAGCTTCCCGTTCAG-3. The relative expression level of Skp2
was determined with the 2−ΔΔCt method.
Western blotting
Harvested cells or tumor tissue samples were lysed
with radioimmunoprecipitation buffer (Beyotime Institute of
Biotechnology, Nantong, China) supplemented with protease and
phosphatase inhibitors (Beyotime Institute of Biotechnology). A
bicinchoninic acid protein assay kit (Beyotime Institute of
Biotechnology) was used to determine protein concentration, and
30–50 µg were separated by sodium dodecyl sulfate-polyacrylamide
gel electrophoresis on a 10–12% gel and transferred to a
polyvinylidene difluoride membrane (Millipore, Billerica, MA, USA)
that was blocked with 5% bovine serum albumin (BSA) for 1 h at room
temperature and then probed overnight at 4°C with primary
antibodies. The membrane was then washed with Tris-buffered
saline/0.1% Tween-20 and incubated with secondary antibodies.
Protein expression was detected by enhanced chemiluminescence
(Millipore) and the signal intensity of protein bands was analyzed
with ImageJ software (National Institutes of Health, Bethesda, MD,
USA) to determine the relative expression levels of target protein
to β-actin. The experiment was performed three times.
Xenograft model
Female nude mice (6 weeks old) were purchased from
Shanghai Slack Experimental Animal Center (Shanghai, China). U87
glioma cells (2×106) in 100 µl saline were injected into
the backs of the mice. Tumor size was measured with calipers; when
tumors reached 100 mm3, mice were randomly divided into
experimental and control groups that were treated with PF and
saline, respectively, by intragastric administration at a dose of 1
g/kg in 0.2 ml every day for 18 days. Tumor size and body weight
were measured two or three times per week. Animal experiments and
use of human cancer cell lines were approved by the Second
Affiliated Hospital of Soochow University Institutional Animal Care
and Use Committee (Suzhou, China).
Statistical analysis
Statistical analyses were carried out using Prism
v.5.0 software (GraphPad Inc., La Jolla, CA, USA). Differences
between the groups were evaluated by one-way analysis of variance
(three or more groups) or the independent t-test (two groups).
Results are presented as mean ± SD. P<0.05 was considered
statistically significant.
Results
PF inhibits glioma cell
proliferation
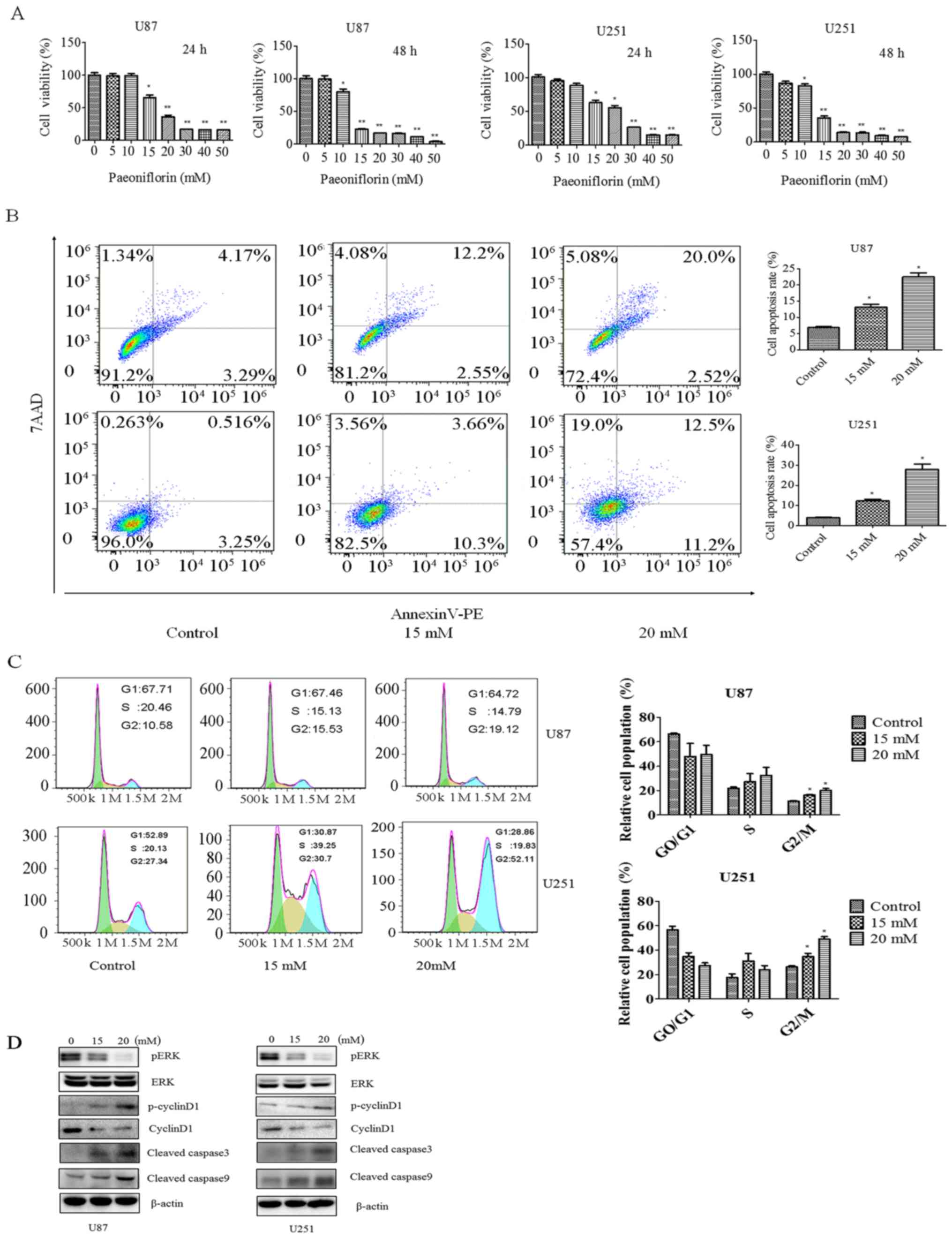
In order to investigate the effect of PF on glioma
cell growth, the viability of U87 and U251 cells treated with
different concentrations of PF for 24 and 48 h was evaluated with
the CCK-8. The cell growth was decreased in a time- and
dose-dependent manner by the PF treatment in both U87 and U251
cells (Fig. 1A). After the
treatment with PF for 24 h, the half-maximal inhibitory
concentration of U87 and U251 cells was approximately 20 mM.
PF induces apoptosis and G2/M arrest
in glioma cells
PF has been shown to inhibit cancer cell growth. We
investigated whether apoptosis is involved in this process with the
Annexin V/PE/7-AAD assay. U87 and U251 cells were treated with 15
or 20 mM PF for 24 h. The rate of apoptosis increased from 7.4 to
22.5% in U87 cells and from 3.7 to 23.7% in U251 cells upon PF
treatment (Fig. 1B), indicating
that PF induces glioma cell apoptosis as reported in previous
studies (19).
The effect of PF on cell cycle progression was
evaluated by PI staining and flow cytometry. Treatment with PF for
24 h caused cells to arrest in G2/M phase, with the fraction
increasing from 23.48 to 34.07% in U87 cells and from 27.34 to
52.11% in U251 cells (Fig. 1C).
To clarify the effects of PF on glioma cell
proliferation and apoptosis, ERK, p-cyclin D1, cyclin D, and
cleaved caspase-3 and −9 expression was evaluated by western
blotting. PF treatment decreased p-ERK and cyclin D and increased
cleaved caspase-3 and −9 levels in a dose-dependent manner
(Fig. 1D), confirming the results
of CCK-8 and flow cytometric analyses.
PF inhibits cell migration and
invasion
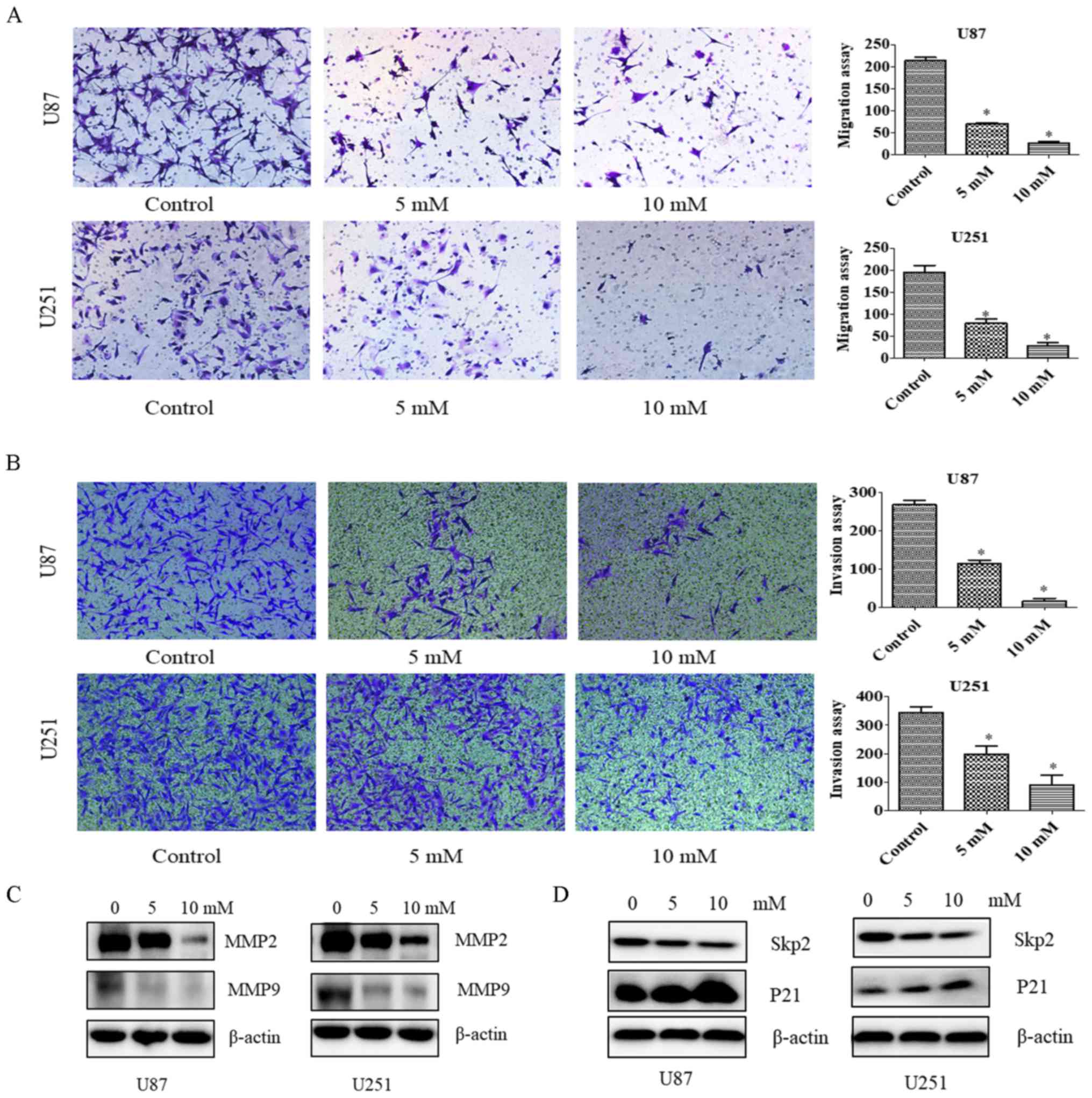
Since PF at concentrations of 5 and 10 mM did not
inhibit the proliferation of U87 and U251 cells (Fig. 1), we chose these concentrations to
study the effects of PF on glioma cell motility with the migration
and invasion assays. The number of migrating U87 and U251 cells
(Fig. 2A) and the number of glioma
cells infiltrating the membrane (Fig.
2B) were decreased by PF treatment relative to the control
group. These results demonstrate that PF inhibits glioma cell
invasion.
To confirm the effects of PF on tumor cell migration
and invasion, MMP2 and MMP9 levels were evaluated by western
blotting. We found that the expression of both proteins was
downregulated by treatment with PF (Fig. 2C); a similar trend was observed for
Skp2 in the presence of 5 and 10 mM PF (Fig. 2D). Thus, PF likely inhibits glioma
cell migration and invasion via modulation of Skp2 signaling.
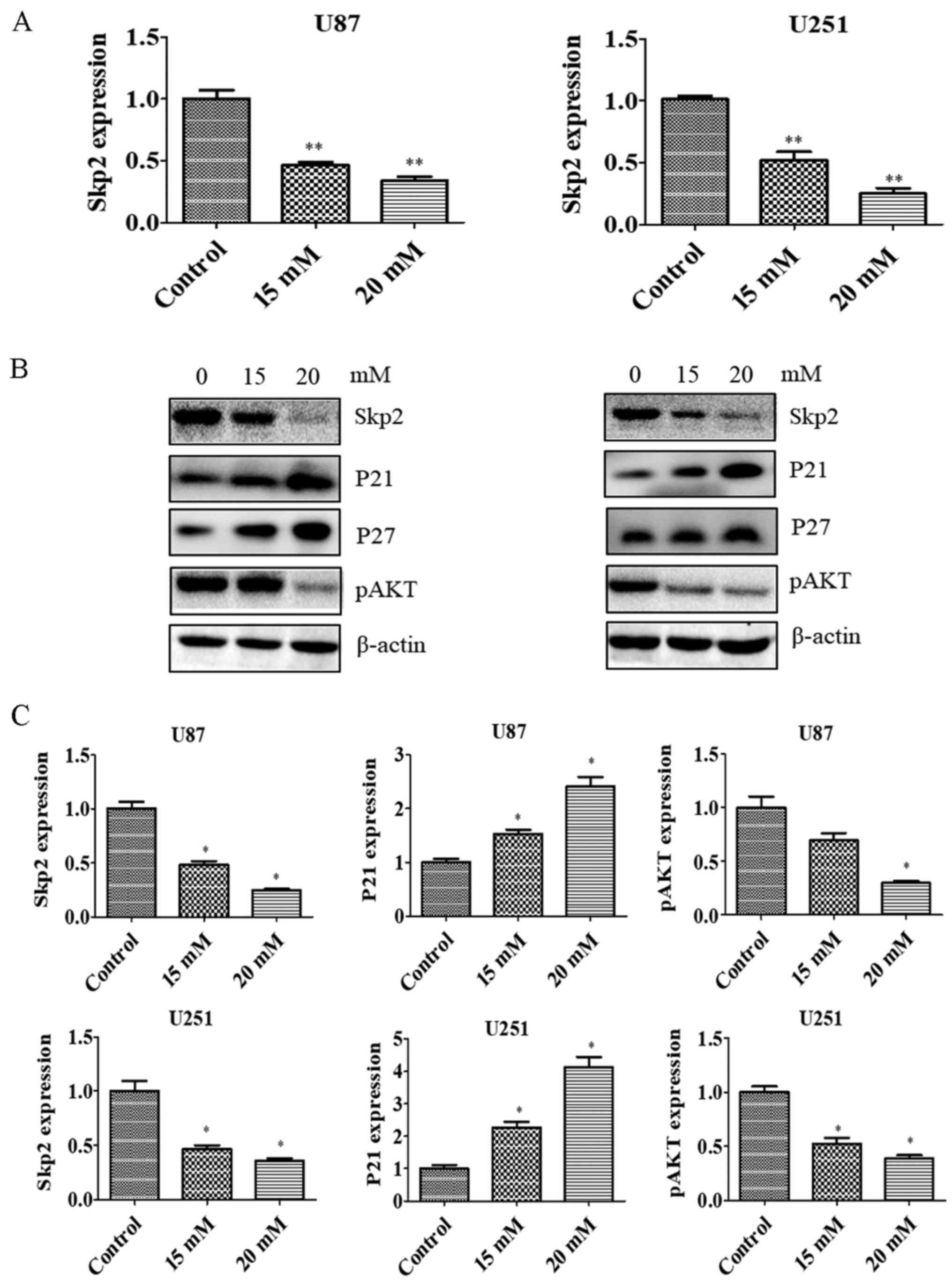
PF suppresses Skp2 expression
Skp2 is known to play an oncogenic role in
tumorigenesis. To test whether Skp2 is involved in the anti-glioma
effects of PF, we evaluated the Skp2 expression in glioma
cells treated with PF. Skp2 mRNA (Fig. 3A) and protein (Fig. 3B and C) levels were downregulated by
PF treatment in U87 and U251 cells. We then examined whether Skp2
inactivation by PF influences the expression of downstream factors
such as P21, P27 and p-AKT. PF inhibited the expression of p-AKT
and induced that of P21 and P27 in both U87 and U251 cells
(Fig. 3B and C). These results
suggest that the antitumor activities of PF are mediated by Skp2
signaling.
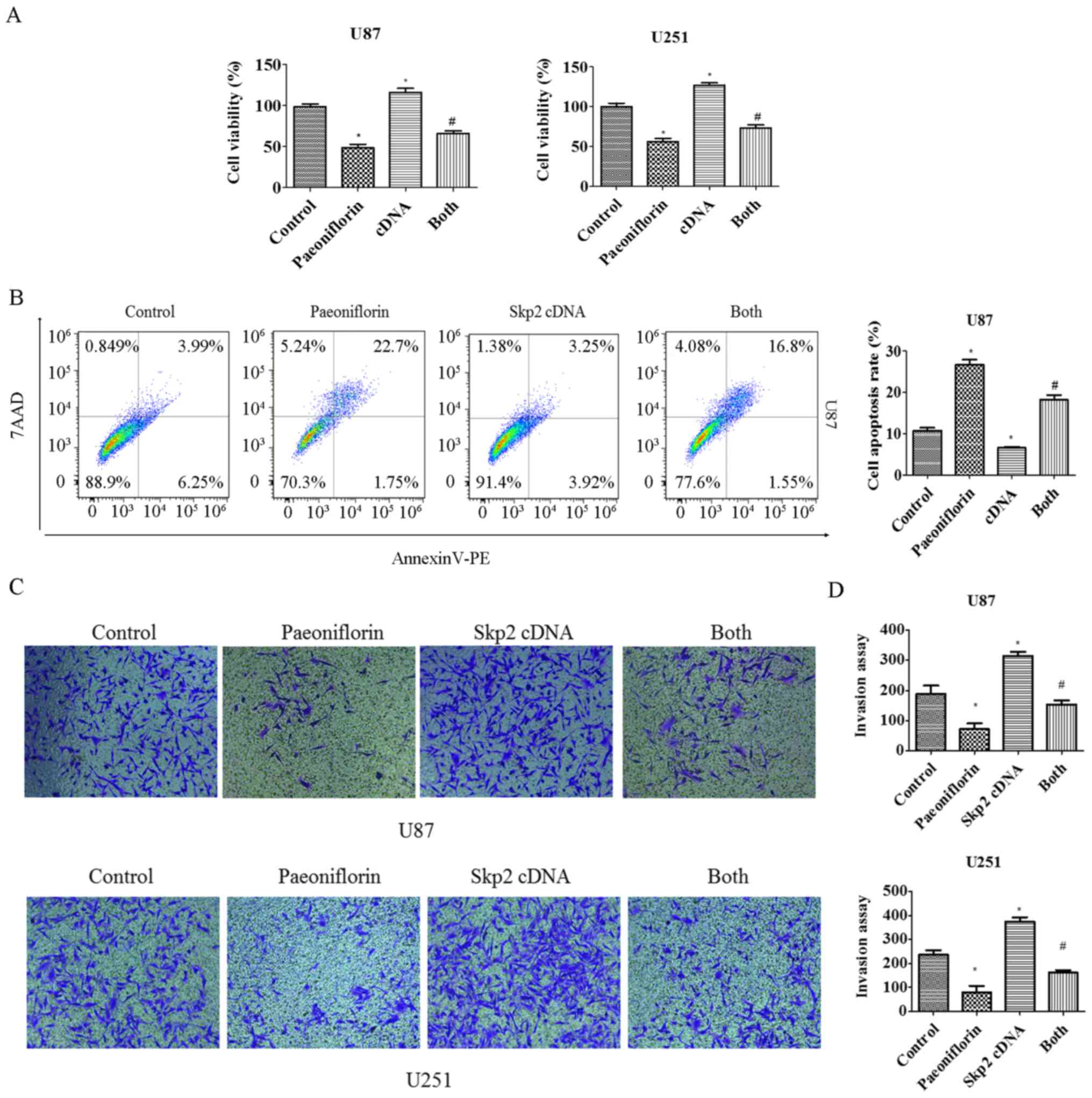
Skp2 overexpression abolishes the
antitumor effects of PF
To clarify the role of Skp2 in the antitumor
activities of PF in glioma cells, U87 and U251 cells were
transfected with Skp2 cDNA and treated with PF for 24 h, and
cell growth and apoptosis were evaluated with the CCK-8 and Annexin
V/PE/7-AAD assays, respectively. Cell proliferation was increased
in cells overexpressing Skp2 (Fig.
4A); moreover, inhibition of cell growth by PF treatment was
abolished (Fig. 4A), suggesting
that the antitumor effects of PF are exerted at least in part via
negative regulation of Skp2. This was confirmed by the finding that
overexpressing Skp2 decreased the number of apoptotic U87 glioma
cells and abrogated the pro-apoptotic effect of PF (Fig. 4B).
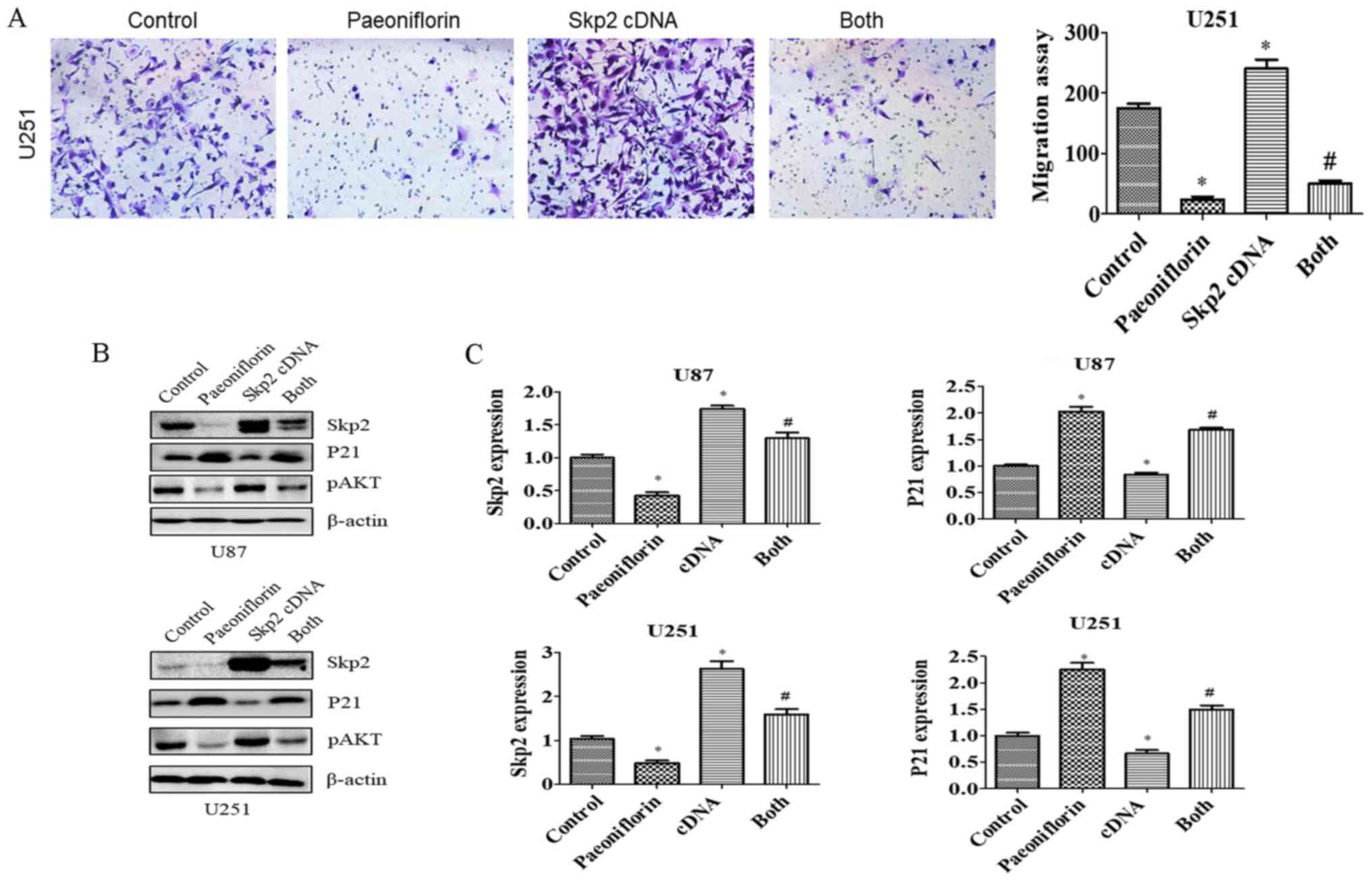
To evaluate the role of Skp2 in glioma cell
motility, U87 and U251 cells were transfected with Skp2 cDNA
and treated with PF for 24 h, and migration and invasion assays
were performed. Skp2 overexpression increased the number of cells
that infiltrated through the Transwell membrane and abrogated the
inhibitory effect of PF on glioma cell invasion (Fig. 4C and D). Similar results were
obtained for the migration assay (Fig.
5A). Skp2 overexpression increased Skp2 levels in glioma cells
and reversed the inhibition of Skp2 and p-AKT and upregulation of
P21 induced by PF (Fig. 5B and C).
These results confirm that PF exerts antitumor effects via
downregulation of Skp2 in glioma cells.
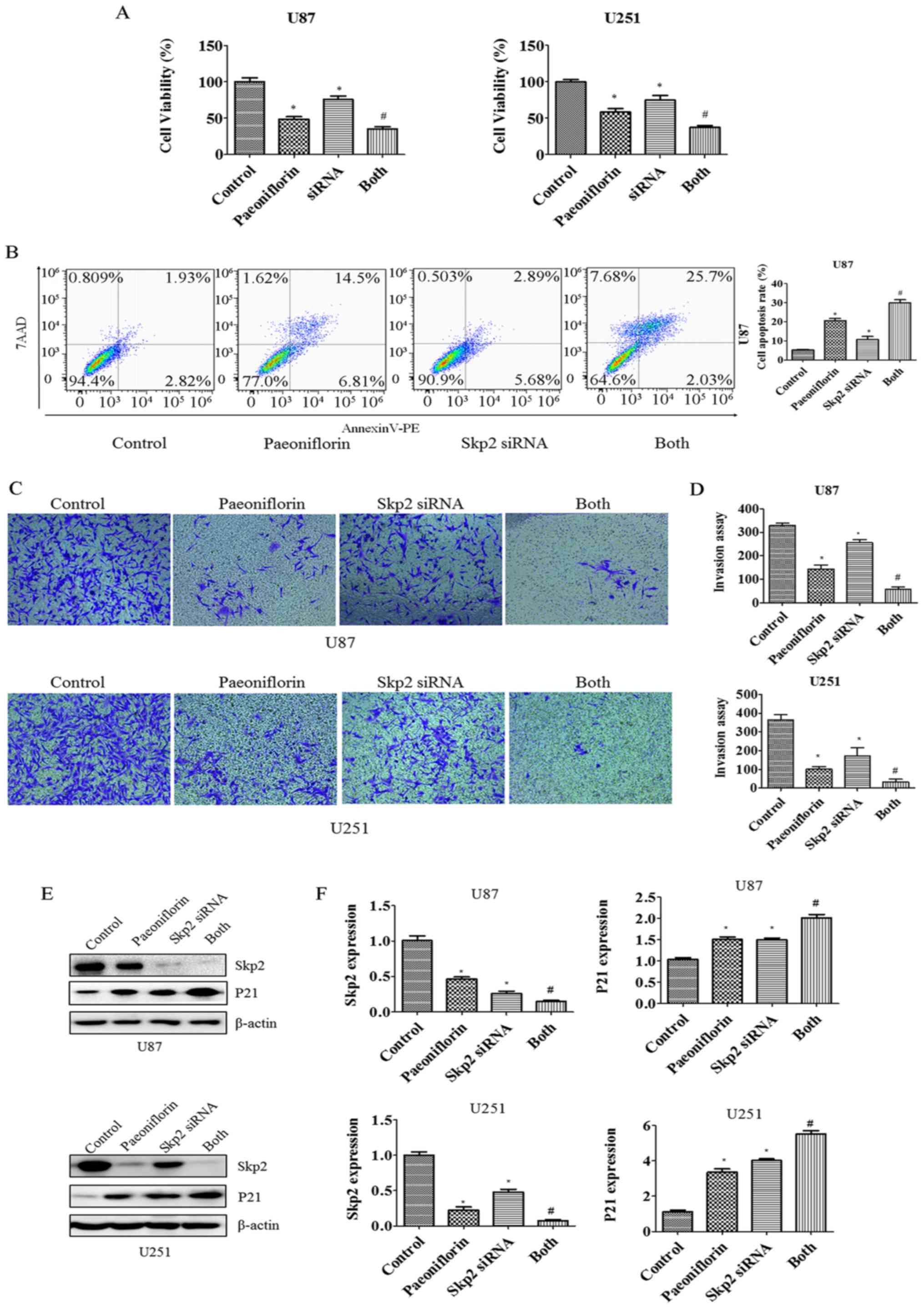
siRNA-mediated knockdown of Skp2
enhances the antitumor effects of PF
To examine the role of Skp2 in glioma in greater
detail, RNA interference was carried out to suppress Skp2
expression in U87 and U251 cells. The cells were transfected with
Skp2 siRNA and then treated with PF for 24 h. The growth
rate decreased (Fig. 6A) whereas
apoptosis was increased in U87 cells upon Skp2 knockdown
(Fig. 6B), indicating that
Skp2 deficiency increases the sensitivity of the cells to
PF. We next investigated whether Skp2 is involved in the regulation
of U87 and U251 cell invasion. The inhibitory effects of PF on
glioma cell invasion were enhanced in cells transfected with
Skp2 siRNA as compared to those treated with PF or
Skp2 siRNA only (Fig. 6C and
D). We also observed that Skp2 expression was suppressed and
P21 expression was increased to a greater degree in
Skp2-depleted cells treated with PF as compared to those
subjected to either treatment alone (Fig. 6E and F).
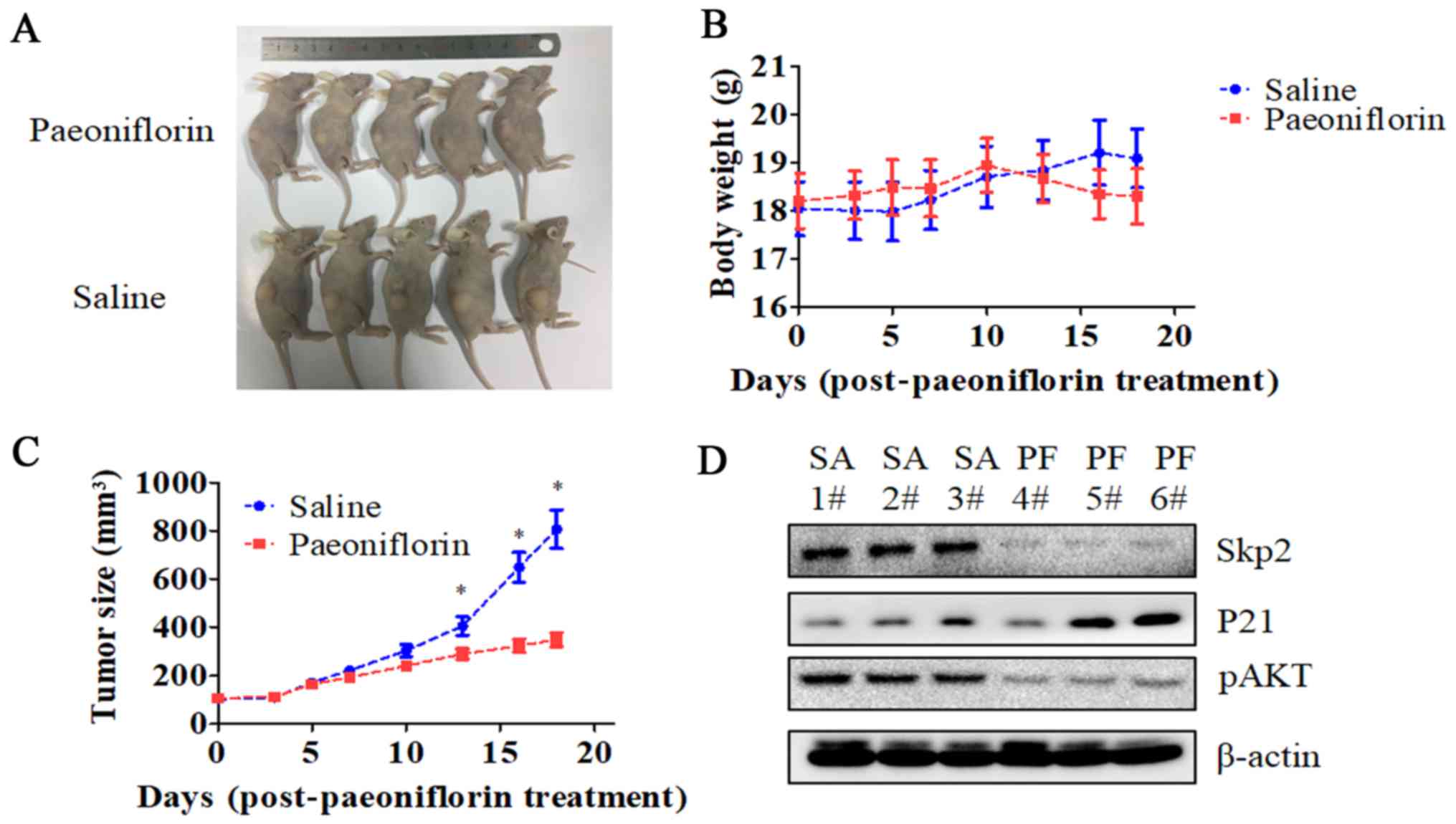
PF suppresses the growth of
glioma-derived tumors in a mouse xenograft model
We established a U87 cell mouse xenograft model to
evaluate the in vivo effects of PF. Tumor volume was lower
in PF-treated as compared to control mice (P<0.05; Fig. 7), whereas no difference in body
weight was observed between the two groups. Consistent with our
in vitro observations, Skp2 and p-AKT levels were lower
while that of P21 was higher in the tumor tissue of mice treated
with PF relative to that of control animals.
Discussion
In the present study, we found that PF induced
apoptosis and inhibited the proliferation, migration, and invasion
of glioma cells. Moreover, we showed that PF exerts its antitumor
effects in part by inhibiting Skp2 expression, since Skp2
overexpression restored whereas Skp2 knockdown enhanced these
effects. Cell proliferation was enhanced in both U87 and U251 cell
lines by overexpression of Skp2 (Fig.
4A), which also rescued the inhibition of cell growth resulting
from PF treatment. On the other hand, Skp2 knockdown
enhanced cell growth inhibition by PF (Fig. 6A). Similar effects were observed in
the cell invasion (Figs. 4C and D
and 6C and D) and migration
(Fig. 5A) assays.
There is increasing evidence to suggest that Skp2
plays a critical role in glioma development and progression
(5,30,35,36).
Skp2 expression is diffuse or focal in most cases of glioblastoma
(GBM), but is absent or expressed at a low level in
well-differentiated astrocytomas (35). Survival in primary GBM patients was
negatively correlated with Skp2 expression level in the tumor
(30); a similar relationship has
been observed in other tumor types such as nasopharyngeal carcinoma
(37). Skp2 deficiency
induces apoptosis and inhibits growth in T98G cells (36). Accordingly, we found that
Skp2 knockdown caused apoptosis and growth arrest in U87
glioma cells. Skp2 overexpression has been shown to promote
cancer progression and metastasis (38); indeed, we observed that Skp2
overexpression stimulated the growth, migration, and invasion of
glioma cells whereas downregulation of Skp2 had the opposite
effect, which is supported by a recent study (5).
Given its importance in tumor progression,
inactivation of Skp2 is a potential strategy for glioma
treatment (21,39). Skp2 knockout mice are viable
and fertile (40), suggesting that
Skp2 inhibition is safe. Inhibition of the Skp2-SCF complex
resulted in tumor regression in preclinical studies (32). Oncostatin M suppressed the
proliferation of GBM cells via downregulation of the
Skp2/Cyclin-dependent kinase regulatory subunit 1 E3 ligase complex
while increasing the expression of P21 and P27 (41). In this study, we found that PF
treatment decreased Skp2 and increased P21 and P27 levels in U87
and U251 glioma cells. Additionally, the Skp2 inhibitor Compound
#25 not only blocked Skp2-mediated Akt ubiquitination, but also
decreased the self-renewal capacity of cancer stem cell populations
(33). Since cancer stem cells are
responsible for GBM initiation and maintenance as well as
chemoresistance and tumor relapse (42–44),
we propose that Skp2 inhibitors have clinical value for treatment
of glioma. Notably, lovastatin was found to suppress the growth of
cancer cells by inducing the degradation of Skp2 (45), and a recent study reported that the
matrine derivative YF-18 inhibited the proliferation and migration
of lung cancer cells via Skp2 suppression (46). Given that these inhibitors have
side-effects, natural agents may be a safer approach for glioma
treatment. In fact, agents such as longikaurin A (47), caffeic acid phenethyl ester,
curcumin (5,48) and rottlerin (49,50)
have shown antitumor effects that are exerted via suppression of
Skp2. This study demonstrates that PF has similar clinical
potential.
To the best of our knowledge, this is the first
report demonstrating that PF exerts an antitumor effect via
downregulation of Skp2. PF inhibited the expression of Skp2 as well
as its target genes P21 and p-AKT. AKT promotes Skp2
phosphorylation at serine 72, resulting in its translocation to the
cytoplasm and activation of its oncogenic function (34,51).
Notably, Skp2 was shown to regulate AKT ubiquitination in breast
cancer and activate AKT in glioma cells (5), which is in agreement with our results.
We previously reported that PF suppressed the expression of STAT3,
which is known to regulate Skp2 expression in cancer cells
(52–54). We speculate that PF inhibits Skp2
expression via negative regulation of STAT3, although further
investigation is needed to evaluate this possibility.
Our results indicate that PF also has therapeutic
effects in a glioma U87 cell xenograft mouse model. The fact that
PF treatment had no effect on body weight suggests that it is a
safe treatment for glioma patients. Consistent with our in
vitro data, Skp2 and p-AKT levels were reduced whereas that of
p21 was increased in tumor tissue upon PF treatment. Unlike drugs
for other cancers, those used to treat glioma must also pass
through the blood-brain barrier. It was previously reported that PF
crosses the blood-brain barrier in rats (55), underscoring its potential as an
effective treatment for glioma.
Acknowledgements
The present study study was financially supported by
the following grants: the Program of Medical Innovation Team and
Leading Talent of Jiangsu Province, China (no. LJ201150); the
Science and Technology Plan Projects of Jiangsu Province, China
(no. BL2012048) and the National Natural Sciences Foundation of
China (no. 81572480).
References
|
1
|
Ahmed R, Oborski MJ, Hwang M, Lieberman FS
and Mountz JM: Malignant gliomas: Current perspectives in
diagnosis, treatment, and early response assessment using advanced
quantitative imaging methods. Cancer Manag Res. 6:149–170.
2014.PubMed/NCBI
|
|
2
|
Stupp R, Mason WP, Van den Bent MJ, Weller
M, Fisher B, Taphoorn MJ, Belanger K, Brandes AA, Marosi C, Bogdahn
U, et al European Organisation for Research and Treatment of Cancer
Brain Tumor and Radiotherapy Groups, ; National Cancer Institute of
Canada Clinical Trials Group, : Radiotherapy plus concomitant and
adjuvant temozolomide for glioblastoma. N Engl J Med. 352:987–996.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fine HA: New strategies in glioblastoma:
Exploiting the new biology. Clin Cancer Res. 21:1984–1988. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ostrom QT, Bauchet L, Davis FG, Deltour I,
Fisher JL, Langer CE, Pekmezci M, Schwartzbaum JA, Turner MC, Walsh
KM, et al: The epidemiology of glioma in adults: A ‘state of the
science’ review. Neuro Oncol. 16:896–913. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang L, Ye X, Cai X, Su J, Ma R, Yin X,
Zhou X, Li H and Wang Z: Curcumin suppresses cell growth and
invasion and induces apoptosis by down-regulation of Skp2 pathway
in glioma cells. Oncotarget. 6:18027–18037. 2015.PubMed/NCBI
|
|
6
|
Walbert T and Chasteen K: Palliative and
supportive care for glioma patients. Cancer Treat Res. 163:171–184.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ma Z, Chu L, Liu H, Wang W, Li J, Yao W,
Yi J and Gao Y: Beneficial effects of paeoniflorin on non-alcoholic
fatty liver disease induced by high-fat diet in rats. Sci Rep.
7:448192017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang H, Qi Y, Yuan Y, Cai L, Xu H, Zhang
L, Su B and Nie H: Paeoniflorin ameliorates experimental autoimmune
encephalomyelitis via inhibition of dendritic cell function and
Th17 cell differentiation. Sci Rep. 7:418872017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen YF, Wu KJ and Wood WG: Paeonia
lactiflora extract attenuating cerebral ischemia and arterial
intimal hyperplasia is mediated by paeoniflorin via modulation of
VSMC migration and Ras/MEK/ERK signaling pathway. Evid Based
Complement Alternat Med. 2013:4824282013.PubMed/NCBI
|
|
10
|
Wang K, Zhu L, Zhu X, Zhang K, Huang B,
Zhang J, Zhang Y, Zhu L, Zhou B and Zhou F: Protective effect of
paeoniflorin on Aβ25–35-induced SH-SY5Y cell injury by
preventing mitochondrial dysfunction. Cell Mol Neurobiol.
34:227–234. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen T, Fu LX, Zhang LW, Yin B, Zhou PM,
Cao N and Lu YH: Paeoniflorin suppresses inflammatory response in
imiquimod-induced psoriasis-like mice and peripheral blood
mononuclear cells (PBMCs) from psoriasis patients. Can J Physiol
Pharmacol. 94:888–894. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li J, Huang S, Huang W, Wang W, Wen G, Gao
L, Fu X, Wang M, Liang W, Kwan HY, et al: Paeoniflorin ameliorates
interferon-alpha-induced neuroinflammation and depressive-like
behaviors in mice. Oncotarget. 8:8264–8282. 2017.PubMed/NCBI
|
|
13
|
Zhai T, Sun Y, Li H, Zhang J, Huo R, Li H,
Shen B and Li N: Unique immunomodulatory effect of paeoniflorin on
type I and II macrophages activities. J Pharmacol Sci. 130:143–150.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
He DY and Dai SM: Anti-inflammatory and
immunomodulatory effects of paeonia lactiflora pall., a traditional
Chinese herbal medicine. Front Pharmacol. 2:102011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang Q, Yuan Y, Cui J, Xiao T and Jiang
D: Paeoniflorin inhibits proliferation and invasion of breast
cancer cells through suppressing Notch-1 signaling pathway. Biomed
Pharmacother. 78:197–203. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang S and Liu W: Paeoniflorin inhibits
proliferation and promotes apoptosis of multiple myeloma cells via
its effects on microRNA-29b and matrix metalloproteinase-2. Mol Med
Rep. 14:2143–2149. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lu JT, He W, Song SS and Wei W:
Paeoniflorin inhibited the tumor invasion and metastasis in human
hepatocellular carcinoma cells. Bratisl Lek Listy. 115:427–433.
2014.PubMed/NCBI
|
|
18
|
Li W, Qi Z, Wei Z, Liu S, Wang P, Chen Y
and Zhao Y: Paeoniflorin inhibits proliferation and induces
apoptosis of human glioma cells via microRNA-16 upregulation and
matrix metalloproteinase-9 downregulation. Mol Med Rep.
12:2735–2740. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Nie XH, Ou-yang J, Xing Y, Li DY, Dong XY,
Liu RE and Xu RX: Paeoniflorin inhibits human glioma cells via
STAT3 degradation by the ubiquitin-proteasome pathway. Drug Des
Devel Ther. 9:5611–5622. 2015.PubMed/NCBI
|
|
20
|
Wang Z, Inuzuka H, Zhong J, Liu P, Sarkar
FH, Sun Y and Wei W: Identification of acetylation-dependent
regulatory mechanisms that govern the oncogenic functions of Skp2.
Oncotarget. 3:1294–1300. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang Z, Fukushima H, Inuzuka H, Wan L, Liu
P, Gao D, Sarkar FH and Wei W: Skp2 is a promising therapeutic
target in breast cancer. Front Oncol. 1:572011.
|
|
22
|
Seki R, Ohshima K and Okamura T:
Prognostic significance of Skp2 and p27kip in diffuse
large B cell lymphoma. Rinsho Ketsueki. 51:1741–1747. 2010.(In
Japanese). PubMed/NCBI
|
|
23
|
Frescas D and Pagano M: Deregulated
proteolysis by the F-box proteins SKP2 and beta-TrCP: Tipping the
scales of cancer. Nat Rev Cancer. 8:438–449. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bornstein G, Bloom J, Sitry-Shevah D,
Nakayama K, Pagano M and Hershko A: Role of the SCFSkp2 ubiquitin
ligase in the degradation of p21Cip1 in S phase. J Biol Chem.
278:25752–25757. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tsvetkov LM, Yeh KH, Lee SJ, Sun H and
Zhang H: p27Kip1 ubiquitination and degradation is
regulated by the SCFSkp2 complex through phosphorylated
Thr187 in p27. Curr Biol. 9:661–664. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kamura T, Hara T, Kotoshiba S, Yada M,
Ishida N, Imaki H, Hatakeyama S, Nakayama K and Nakayama KI:
Degradation of p57Kip2 mediated by SCFSkp2-dependent
ubiquitylation. Proc Natl Acad Sci USA. 100:pp. 10231–10236. 2003;
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Shin E, Kim SH, Jeong HY, Jang JJ and Lee
K: Nuclear expression of S-phase kinase-associated protein 2
predicts poor prognosis of hepatocellular carcinoma. APMIS.
120:349–357. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liu J, Wei XL, Huang WH, Chen CF, Bai JW
and Zhang GJ: Cytoplasmic Skp2 expression is associated with p-Akt1
and predicts poor prognosis in human breast carcinomas. PLoS One.
7:e526752012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chen G, Cheng Y, Zhang Z, Martinka M and
Li G: Cytoplasmic Skp2 expression is increased in human melanoma
and correlated with patient survival. PLoS One. 6:e175782011.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Saigusa K, Hashimoto N, Tsuda H, Yokoi S,
Maruno M, Yoshimine T, Aoyagi M, Ohno K, Imoto I and Inazawa J:
Overexpressed Skp2 within 5p amplification detected by array-based
comparative genomic hybridization is associated with poor prognosis
of glioblastomas. Cancer Sci. 96:676–683. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chan CH, Li CF, Yang WL, Gao Y, Lee SW,
Feng Z, Huang HY, Tsai KK, Flores LG, Shao Y, et al: The Skp2-SCF
E3 ligase regulates Akt ubiquitination, glycolysis, herceptin
sensitivity, and tumorigenesis. Cell. 149:1098–1111. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lin HK, Chen Z, Wang G, Nardella C, Lee
SW, Chan CH, Yang WL, Wang J, Egia A, Nakayama KI, et al: Skp2
targeting suppresses tumorigenesis by Arf-p53-independent cellular
senescence. Nature. 464:374–379. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chan CH, Morrow JK, Li CF, Gao Y, Jin G,
Moten A, Stagg LJ, Ladbury JE, Cai Z, Xu D, et al: Pharmacological
inactivation of Skp2 SCF ubiquitin ligase restricts cancer stem
cell traits and cancer progression. Cell. 154:556–568. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lin HK, Wang G, Chen Z, Teruya-Feldstein
J, Liu Y, Chan CH, Yang WL, Erdjument-Bromage H, Nakayama KI, Nimer
S, et al: Phosphorylation-dependent regulation of cytosolic
localization and oncogenic function of Skp2 by Akt/PKB. Nat Cell
Biol. 11:420–432. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Schiffer D, Cavalla P, Fiano V, Ghimenti C
and Piva R: Inverse relationship between p27/Kip.1 and the F-box
protein Skp2 in human astrocytic gliomas by immunohistochemistry
and Western blot. Neurosci Lett. 328:125–128. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lee SH and McCormick F: Downregulation of
Skp2 and p27/Kip1 synergistically induces apoptosis in T98G
glioblastoma cells. J Mol Med (Berl). 83:296–307. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang J, Huang Y, Guan Z, Zhang JL, Su HK,
Zhang W, Yue CF, Yan M, Guan S and Liu QQ: E3-ligase Skp2 predicts
poor prognosis and maintains cancer stem cell pool in
nasopharyngeal carcinoma. Oncotarget. 5:5591–5601. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hung WC, Tseng WL, Shiea J and Chang HC:
Skp2 overexpression increases the expression of MMP-2 and MMP-9 and
invasion of lung cancer cells. Cancer Lett. 288:156–161. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chan CH, Morrow JK, Zhang S and Lin HK:
Skp2: A dream target in the coming age of cancer therapy. Cell
Cycle. 13:679–680. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Nakayama K, Nagahama H, Minamishima YA,
Matsumoto M, Nakamichi I, Kitagawa K, Shirane M, Tsunematsu R,
Tsukiyama T, Ishida N, et al: Targeted disruption of Skp2 results
in accumulation of cyclin E and p27Kip1, polyploidy and
centrosome overduplication. EMBO J. 19:2069–2081. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Halfter H, Friedrich M, Resch A, Kullmann
M, Stögbauer F, Ringelstein EB and Hengst L: Oncostatin M induces
growth arrest by inhibition of Skp2, Cks1, and cyclin A expression
and induced p21 expression. Cancer Res. 66:6530–6539. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Singh SK, Hawkins C, Clarke ID, Squire JA,
Bayani J, Hide T, Henkelman RM, Cusimano MD and Dirks PB:
Identification of human brain tumour initiating cells. Nature.
432:396–401. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Galli R, Binda E, Orfanelli U, Cipelletti
B, Gritti A, De Vitis S, Fiocco R, Foroni C, Dimeco F and Vescovi
A: Isolation and characterization of tumorigenic, stem-like neural
precursors from human glioblastoma. Cancer Res. 64:7011–7021. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Chen J, Li Y, Yu TS, McKay RM, Burns DK,
Kernie SG and Parada LF: A restricted cell population propagates
glioblastoma growth after chemotherapy. Nature. 488:522–526. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Vosper J, Masuccio A, Kullmann M, Ploner
C, Geley S and Hengst L: Statin-induced depletion of geranylgeranyl
pyrophosphate inhibits cell proliferation by a novel pathway of
Skp2 degradation. Oncotarget. 6:2889–2902. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Wu L, Wang G, Wei J, Huang N, Zhang S,
Yang F, Li M, Zhou G and Wang L: Matrine derivative YF-18 inhibits
lung cancer cell proliferation and migration through
down-regulating Skp2. Oncotarget. 8:11729–11738. 2017.PubMed/NCBI
|
|
47
|
Liao YJ, Bai HY, Li ZH, Zou J, Chen JW,
Zheng F, Zhang JX, Mai SJ, Zeng MS, Sun HD, et al: Longikaurin A, a
natural ent-kaurane, induces G2/M phase arrest via downregulation
of Skp2 and apoptosis induction through ROS/JNK/c-Jun pathway in
hepatocellular carcinoma cells. Cell Death Dis. 5:e11372014.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Su J, Zhou X, Wang L, Yin X and Wang Z:
Curcumin inhibits cell growth and invasion and induces apoptosis
through down-regulation of Skp2 in pancreatic cancer cells. Am J
Cancer Res. 6:1949–1962. 2016.PubMed/NCBI
|
|
49
|
Yin X, Zhang Y, Su J, Hou Y, Wang L, Ye X,
Zhao Z, Zhou X, Li Y and Wang Z: Rottlerin exerts its anti-tumor
activity through inhibition of Skp2 in breast cancer cells.
Oncotarget. 7:66512–66524. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Su J, Wang L, Yin X, Zhao Z, Hou Y, Ye X,
Zhou X and Wang Z: Rottlerin exhibits anti-cancer effect through
inactivation of S phase kinase-associated protein 2 in pancreatic
cancer cells. Am J Cancer Res. 6:2178–2191. 2016.PubMed/NCBI
|
|
51
|
Gao D, Inuzuka H, Tseng A, Chin RY, Toker
A and Wei W: Phosphorylation by Akt1 promotes cytoplasmic
localization of Skp2 and impairs APCCdh1-mediated Skp2 destruction.
Nat Cell Biol. 11:397–408. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Huang H, Zhao W and Yang D: Stat3 induces
oncogenic Skp2 expression in human cervical carcinoma cells.
Biochem Biophys Res Commun. 418:186–190. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Wei Z, Jiang X, Qiao H, Zhai B, Zhang L,
Zhang Q, Wu Y, Jiang H and Sun X: STAT3 interacts with Skp2/p27/p21
pathway to regulate the motility and invasion of gastric cancer
cells. Cell Signal. 25:931–938. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Wang ST, Ho HJ, Lin JT, Shieh JJ and Wu
CY: Simvastatin-induced cell cycle arrest through inhibition of
STAT3/SKP2 axis and activation of AMPK to promote p27 and p21
accumulation in hepatocellular carcinoma cells. Cell Death Dis.
8:e26262017. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
He X, Xing D, Ding Y, Li Y, Xiang L, Wang
W and Du L: Determination of paeoniflorin in rat hippocampus by
high-performance liquid chromatography after intravenous
administration of Paeoniae Radix extract. J Chromatogr B Analyt
Technol Biomed Life Sci. 802:277–281. 2004. View Article : Google Scholar : PubMed/NCBI
|