Introduction
Colorectal carcinoma is a common malignant tumor of
the digestive system. In North America and Western Europe,
colorectal carcinoma is the third leading cause of cancer death
(1). Recently, the incidence and
mortality of colorectal cancer in China are showing a rising trend
(2). However, the effective
therapeutic method for colorectal carcinoma is still lacking.
Notch signaling pathway is a highly-conserved
intercellular signal transduction pathway, which is composed of
Notch receptor, Notch ligand (DSL protein) and transcription factor
CSL (DNA binding protein). Notch signaling pathway regulates cell
differentiation, proliferation and apoptosis, and plays important
roles in embryonic development and cell fate determination
(3). Notch signaling pathway not
only plays an important role in normal cell differentiation, but
also participates in the occurrence and development of tumors
(4). The expression of Notch
receptors and ligands, and the aberrant activation of Notch
signaling pathway have been found in a variety of malignant tumors
(5,6). However, Notch signaling pathway and
its role in tumorigenesis are extremely complex. Notch signaling
pathway plays a role in promoting tumorigenesis in most cases
(7,8), but functions as tumor suppressor in a
few cases. The discrepancy is likely caused by that Notch-1 which
was detected in different tumor stages (9,10).
Additionally, Notch expression may be associated with the cell
microenvironment, tissue type and determines cell proliferation,
differentiation, self-renewal or apoptosis (11,12).
Notch signaling pathway has been extensively investigated in tumor
therapy (13). DAPT, a γ-secretase
inhibitor can block Notch receptor/ligand binding enzyme cleavage,
thereby blocking the Notch signaling. Therefore, DAPT has been used
in clinical trials to treat cancers (14).
There are a variety of Notch receptors and ligands
expressed in the gut tissue (15,16).
Ngn3, HES1 and Math1, three regulators of Notch signaling pathway
have intestinal phenotype. Transgenic methods further confirmed
that Notch directly regulated the fate of intestinal epithelial
cells (17). van Es et al
(18) inhibited the Notch signal by
using the γ-secretase inhibitor, and the differentiation of colon
adenoma cells in mice recovered. Nevertheless, the relationship
between Notch and colorectal cancer is not clear. In this study, we
screened the expression of Notch-1 in colorectal cancer tissue and
cancer cell lines, and investigated the functions of Notch-1 in
colorectal biological activities.
Materials and methods
Colorectal cancer tissues and cell
lines
Colorectal carcinoma, colorectal adenoma and
paracancerous tissues and normal colorectal tissues were obtained
from the First Affiliated Hospital of Nanchang University. This
study was approved by the Ethics Committee of Nanchang University.
Colorectal cancer cell lines (COLO 205, HT29, SW480 and SW1116)
were gifted by Digestion Institute of Nanfang Hospital. LoVo cells
were obtained from Institute of Biochemistry and Cell Biology,
Chinese Academy of Sciences (China).
Cell culture and transfection
Colorectal cancer cell lines (COLO 205, HT29, SW480,
SW1116 and LoVo) were cultured in Dulbecco's minimum essential
medium (DMEM) (Gibco, Grand Island, NY, USA) supplemented with 10%
fetal bovine serum (FBS) and 100 U/ml penicillin-streptomycin
(Sigma, Ronkonkoma, NY, USA) in 5% CO2 at 37°C. Cell
confluence at 50–70% was applied in the following experiments. The
cells were divided into three groups: non-RNAi group (NR), negative
control group (NC) and RNAi group (R). pSiRNA-Notch-1 and empty
vector pSilencer 5.1-H1 Retro (Shanghai GenePharma, Shanghai,
China) were transfected by Lipofectamine 2000 and packaged into
viruses.
DAPT treatment
HT29 cells were treated by DAPT (6.25–50 µM) (Sigma)
for 1, 2, 3 and 4 days, respectively. After treatments, the cell
proliferation and apoptosis were detected. DAPT was dissolved in
0.2% (v/v) DMSO and a similar concentration of DMSO was applied as
negative control. Proliferation was detected by
3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide
(MTT) assay. The cell cycle and apoptosis were detected by flow
cytometry and terminal deoxynucleotidyl transferase dUTP nick end
labeling (TUNEL) assay.
MTT assay
Cells were seeded in 96-well plates. When cell
confluence reached 50–70%, 100 µl virus supernatant was added to
knock down Notch-1 expression. After transfection for 1, 2, 3 and 4
days, MTT assay was applied to detect the cell proliferation as
previously described (15). The
optical density (OD) was determined by Microplate Reader (BioTek,
Winooski, VT, USA) at 570 nm.
Flow cytometry
When cell confluence reached 50–70%, 100 µl virus
supernatant was added to knock down Notch-1 expression. After
transfection for 48 h, the cells were collected for Annexin
V-FITC/PI staining (Beyotime, Ningbo, China) and apoptosis was
detected within 1 h by FACSCalibur (BD Biosciences, Franklin Lakes,
NJ, USA).
After transfection for 48 h, the cells were
collected for PI staining and cell cycle distribution was assessed
by FACSCalibur (BD Biosciences) within 1 h after staining.
TUNEL assay
TUNEL assay was conducted according to the
instruction of DeadEnd™ Colorimetric TUNEL system (Promega,
Madison, WI, USA).
Immunohistochemical and
immunocytochemical staining
Cancer tissues were fixed in 10% formaldehyde and
embedded in paraffin. Sections (3–5 µm) were continuously sliced.
After dewaxing by xylene, the tissues were dehydrated in 70, 75,
80, 85 and 95% gradient alcohol. Hydrogen peroxide (3%) was applied
to repair the antigen. The mounted cells were fixed in acetone. The
non-specific staining was blocked by goat serum at 4°C overnight.
Immunostaining of histological sections was performed using
monoclonal antibodies against Notch-1 (1:200, ab52627; Abcam,
Cambridge, MA, USA) and Jagged1 (1:200, ab7771; Abcam) overnight at
4°C followed by a 30-min incubation with secondary antibody (Dako,
Carpinteria, CA, USA) and visualization with 3,3′-diaminobenzidine
(DAB) for 3 min. PBS was employed as the negative control for the
primary antibody. The positive staining was analyzed by ImageJ
software.
Reverse-transcript PCR
Total RNA was extracted from tissues or cell lines
using TRIzol reagent (Thermo Fisher Scientific, Waltham, MA, USA).
RNA concentrations were determined spectrophotometrically, and 1 µg
total RNA was reverse transcribed using an avian myeloblastosis
virus reverse-transcriptase kit (Promega). The expression of
Notch-1, p53 upregulated modulator of apoptosis (PUMA), p21 and
β-actin in mRNA was detected. PCR primers used were as follows:
Notch-1-F, CTCCCCGTTCCAGCAGTCTC and Notch-1-R,
CAGCCACTCGCATTGACCAT; PUMA-F, GATGGCGGACGACCTCAAC and PUMA-R,
ACGGGCA CCAGCACAACA; p21-F, TTGATTAGCAGCGGAACA and p21-R,
TACAGTCTAGGTGGAGAAACG; β-actin-F, AAGG AAGGCTGGAAGAGTGC and
β-actin-R, CTACAATGAG CTGCGTGTGG.
The amplification reactions were carried out within
a PCR system (Applied Biosystems), with initial hold step (95°C for
4 min) and 40 cycles of a PCR (95°C for 40 sec, 54°C for 30 sec and
72°C for 30 sec). The expression of target genes was normalized to
β-actin.
Western blotting
Protein was extracted from cell lines for western
blotting as previously described (19). The anti-Notch-1 antibody (1:200,
ab52627; Abcam) was incubated overnight at 4°C. After washing with
0.1% PBST (0.1% Tween-20), membranes were probed with anti-mouse or
anti-rabbit horseradish peroxidase secondary antibodies (1:100;
cat. nos. ab131368 and ab191866; Abcam) at room temperature for 2
h. Enhanced Chemiluminescent reagent kit (GE Healthcare Life
Sciences, Chalfont, UK) was applied to assist the staining. The
blots were scanned by ChemiDoc™ XRS (Bio-Rad Laboratories,
Hercules, CA, USA) and grey density was analyzed by Quantity One
v4.62.
Statistical analysis
Data are presented as means ± standard deviations.
One-way analysis of variance with post hoc Bonferroni test for
multiple comparisons was performed. Differences were considered
significant at P<0.05.
Results
Expression of Notch-1 and Jagged1
Initially, we detected the expression of Notch-1 and
its ligand-Jagged1 in colorectal carcinoma, colorectal adenoma and
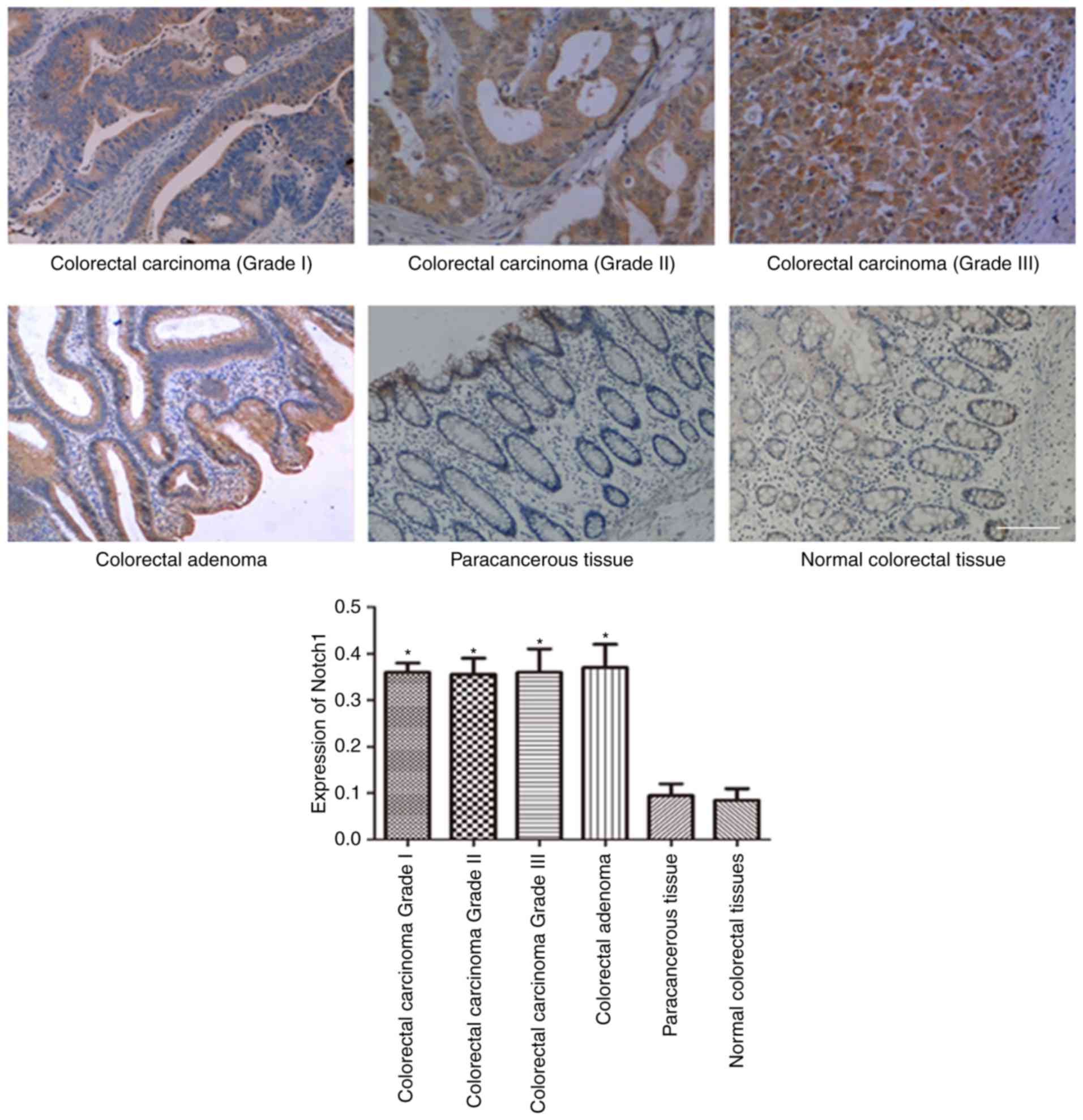
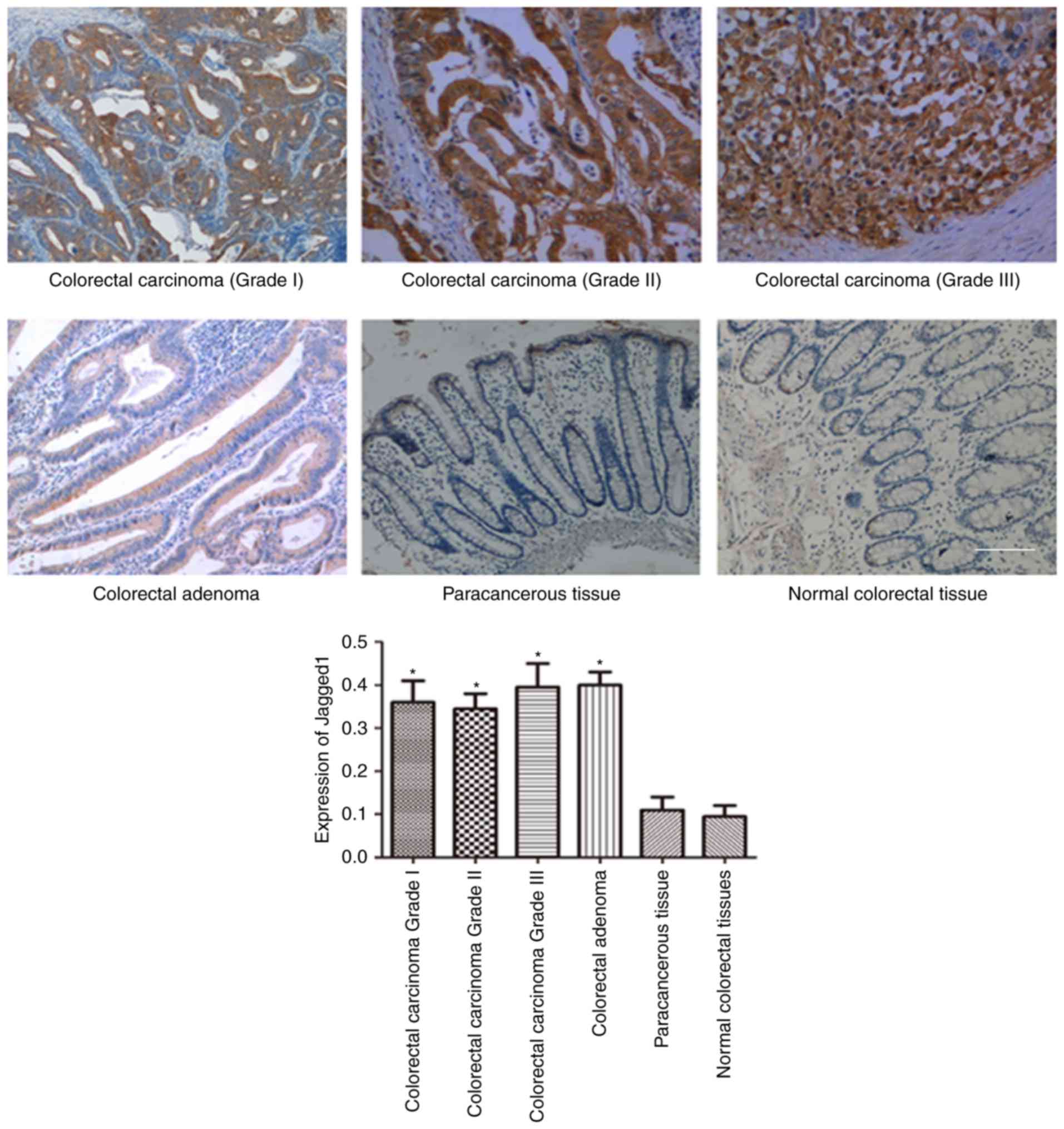
carcinoma-adjacent tissues and normal colorectal tissues. As shown
in Figs. 1 and 2, the expression of Notch-1 (Fig. 1) and Jagged1 (Fig. 2) in colorectal carcinoma and
colorectal adenoma was obviously higher than those in
carcinoma-adjacent tissues and normal colorectal tissues. However,
there was no significant difference between carcinoma-adjacent
tissues and normal colorectal tissues regarding Notch-1 and Jagged1
expression.
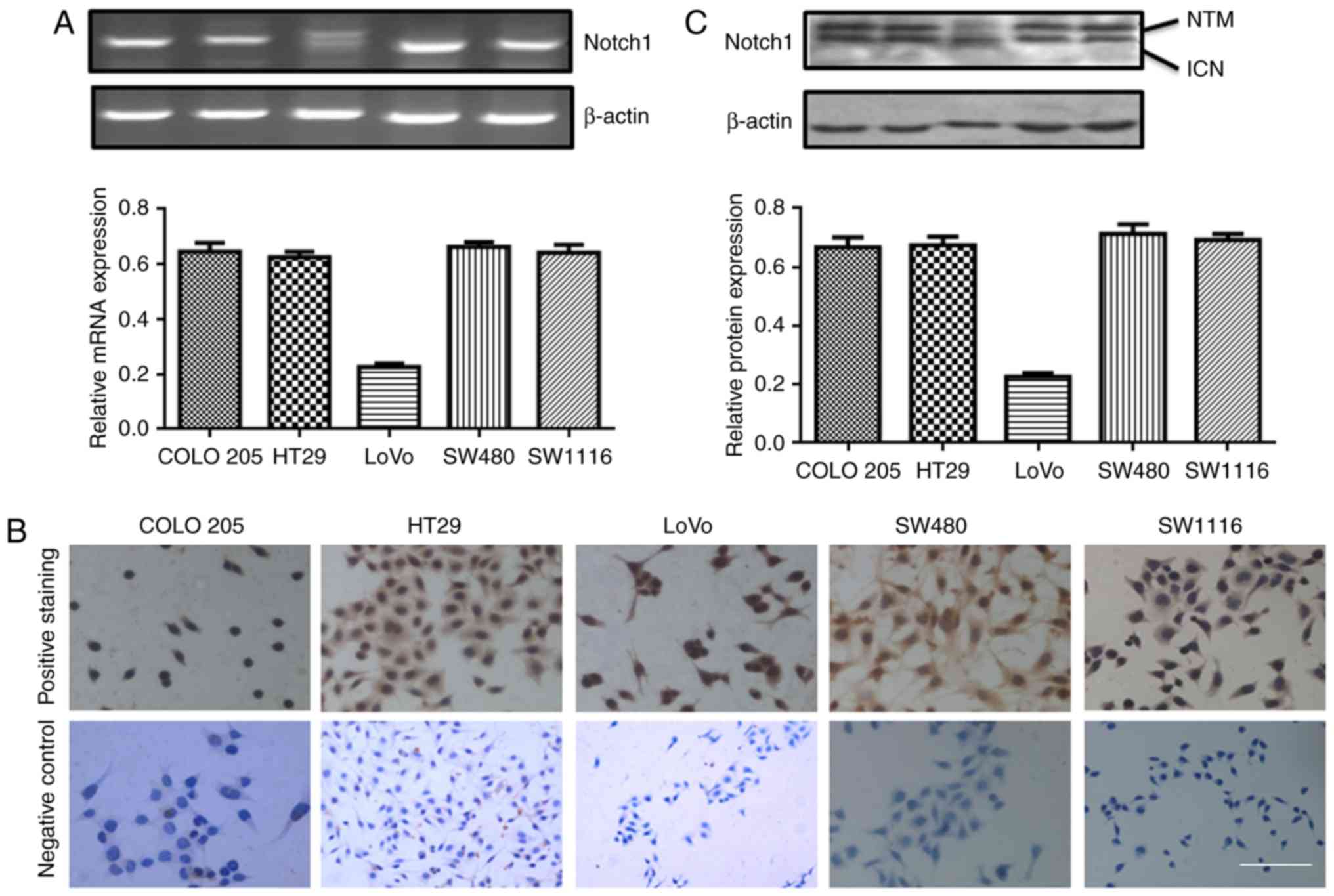
We also detected Notch-1 mRNA expression in COLO205,
HT29, LoVo, SW480 and SW1116 cells by RT-PCR. As shown in Fig. 3A, Notch-1 expression was relatively
higher in COLO 205, HT29, SW480 and SW1116 cells, compared with
LoVo. Immunohistochemical staining showed that Notch-1 was
expressed in all five colorectal carcinoma cells (Fig. 3B). Two sub-types of Notch-1-4 exist:
Notch transmembrane fragment (NTM) and intracellular domain of
Notch (ICN). We found two bands of Notch-1 in all the five cell
lines. Especially, Notch-1 was highly expressed in COLO 205, HT29,
SW480 and SW1116 cells (Fig.
3C).
pSiRNA-Notch-1 inhibits Notch-1
expression and cell proliferation in HT29 cells
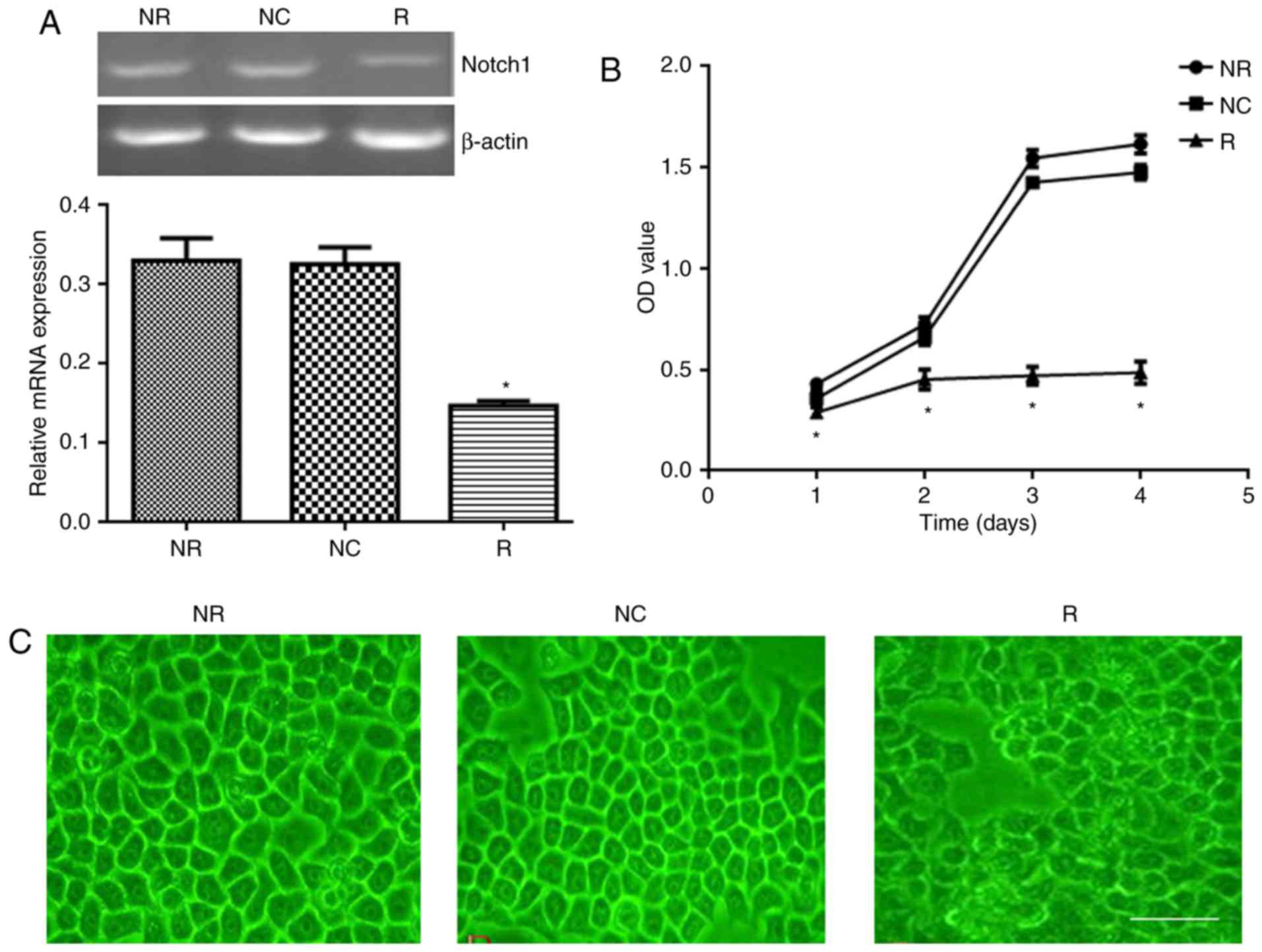
In the next part of experiments, we designed
pSiRNA-Notch-1 and transferred the vector into viruses. The viral
supernatant was collected to transfect HT29 cells. Our data showed
that viral pSiRNA-Notch-1 significantly reduced Notch-1 expression
in HT29 cells compared with NC group (Fig. 4A). The cell proliferation after
viral transfection was detected. As shown in Fig. 4B, viral pSiRNA-Notch-1 significantly
inhibited the cell proliferation compared with NC group. The
morphology of the cells was also observed. As shown in Fig. 4C, the cells in NR and NC groups
showed clear boundary, while the cells in viral pSiRNA-Notch-1
group showed diffused cell boundary. Abundant dead cells were
observed after Notch-1 knockdown.
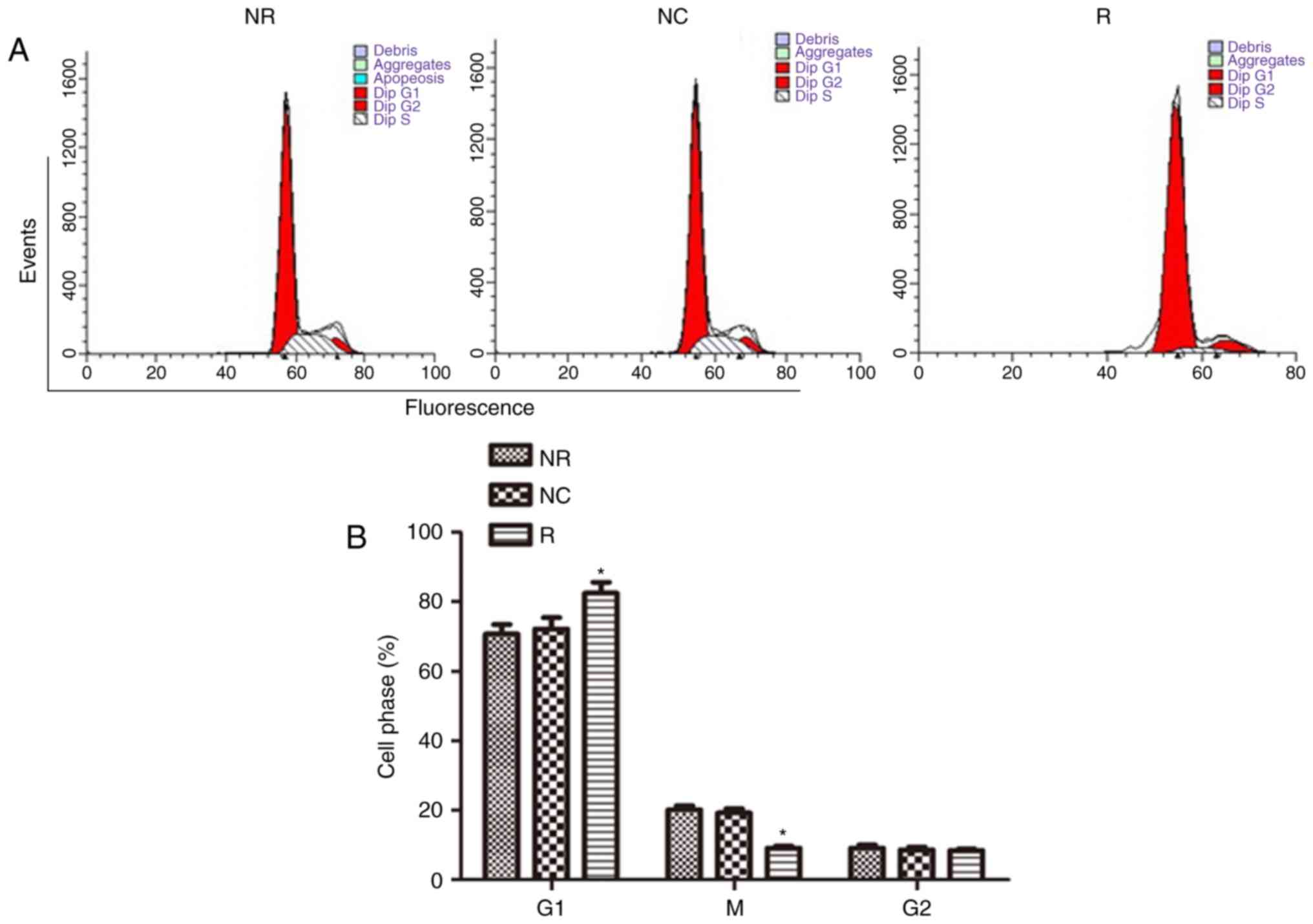
pSiRNA-Notch-1 arrests the cells at G1
phase and promotes apoptosis
Cell cycle was detected by flow cytometry after PI
staining. As shown in Fig. 5A and
B, Notch-1 knockdown significantly arrested the cell cycle at
G1 phase (vs NC, P<0.05) (NR: 70.75%; NC: 72.18%; R: 82.43%).
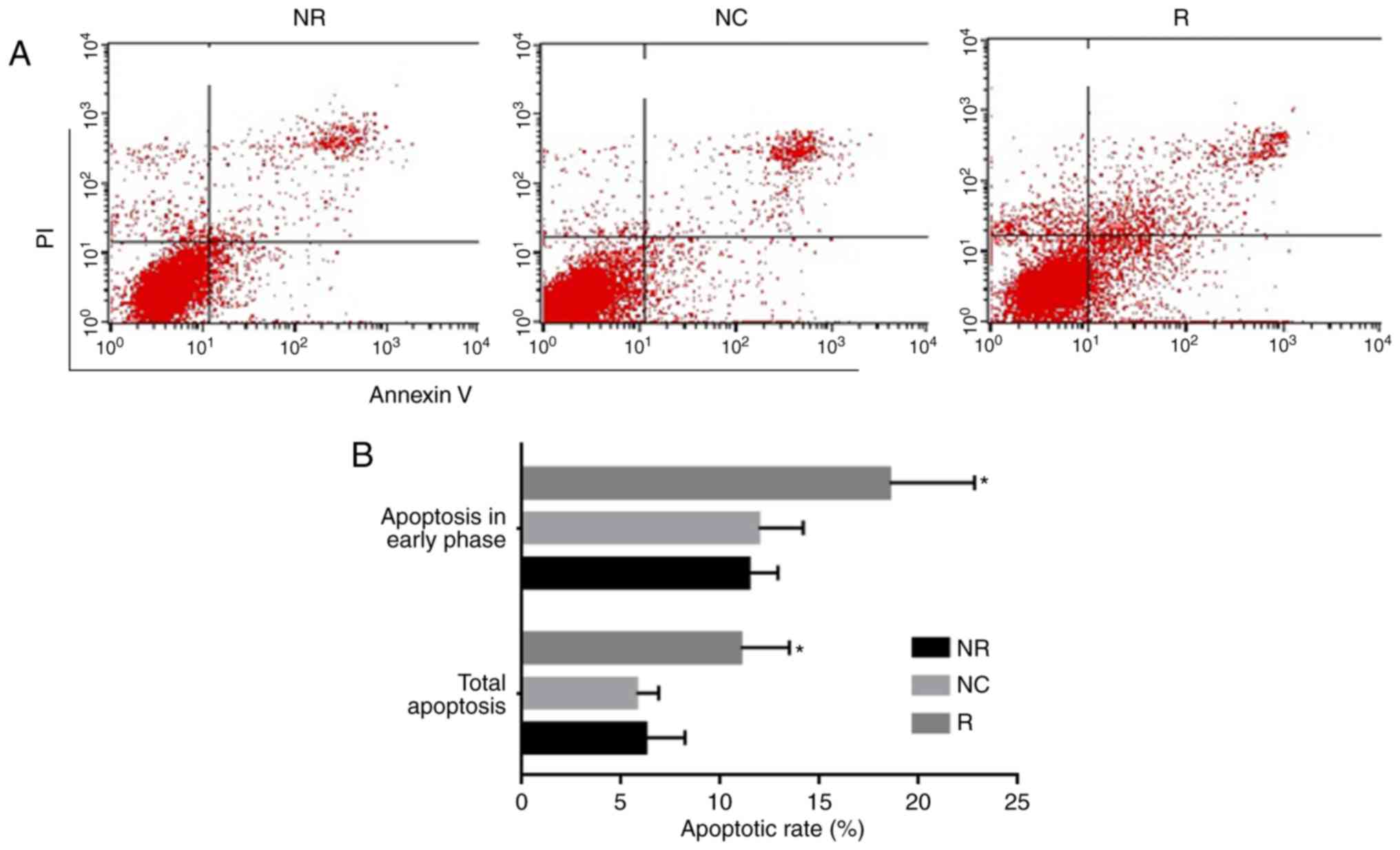
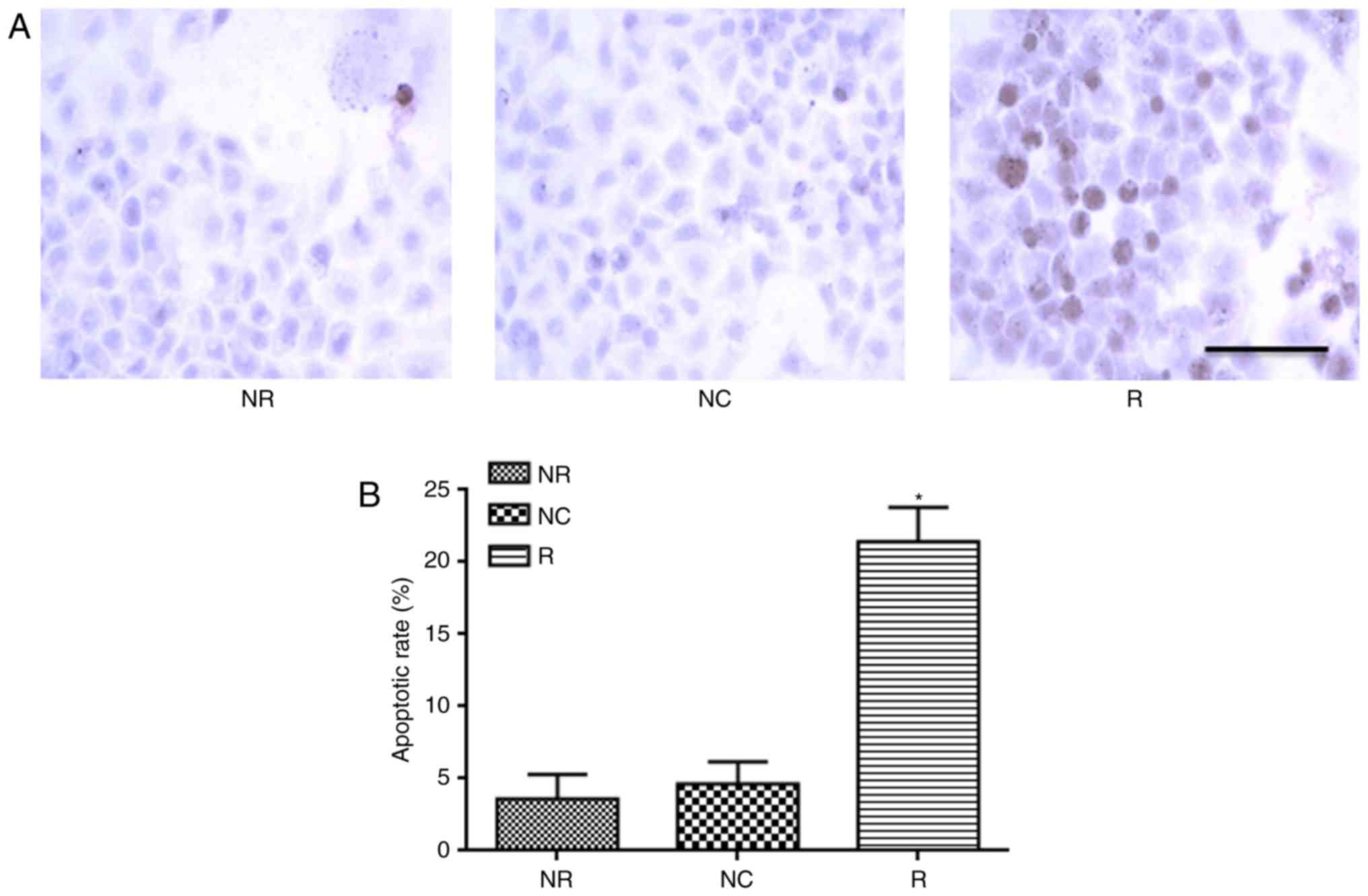
Apoptosis was detected by TUNEL assay and flow cytometry. As shown
in Fig. 6, Notch-1 knockdown not
only promoted apoptosis in early phase, but also total apoptosis.
These data were further confirmed by TUNEL assay (Fig. 7).
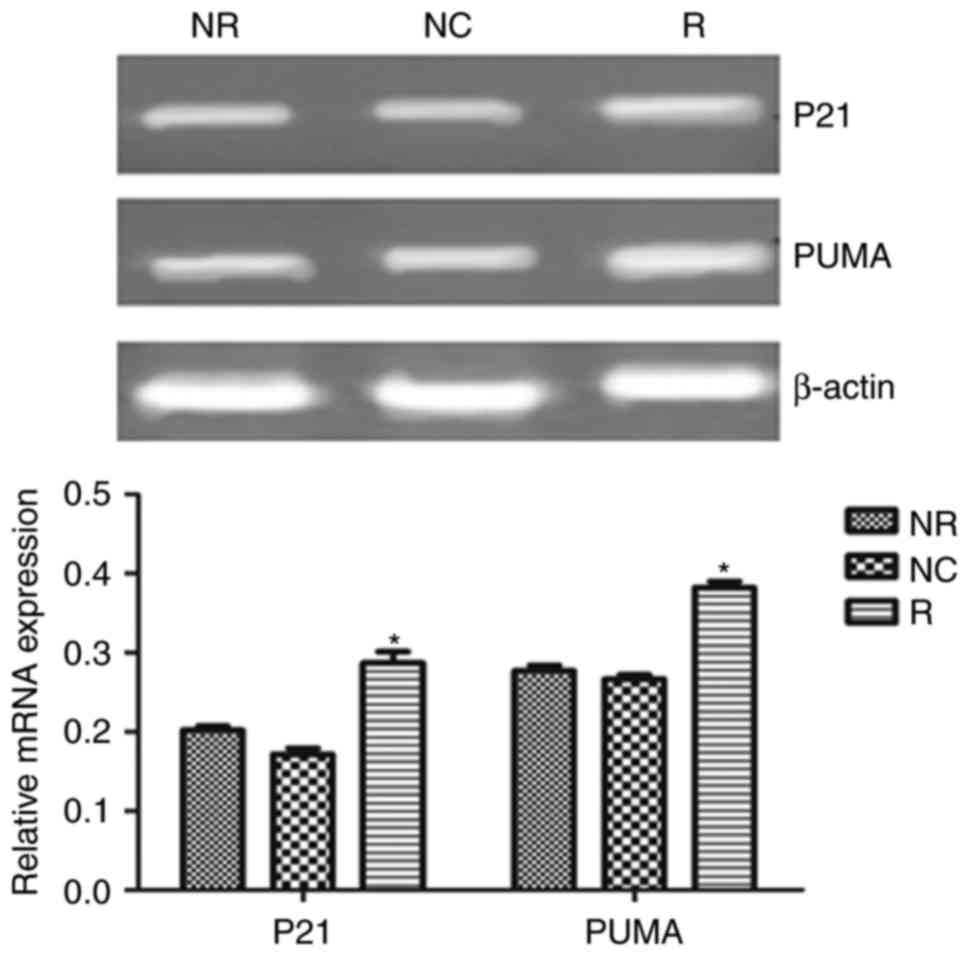
pSiRNA-Notch-1 promotes p21 and PUMA
expression
We also detected apoptosis-related gene expression.
As shown in Fig. 8, Notch-1
knockdown significantly increased p21 and PUMA expression in mRNA
level.
DAPT inhibits cell proliferation,
arrests cell cycle at G1 phase and promotes apoptosis
We also applied different concentrations of DAPT in
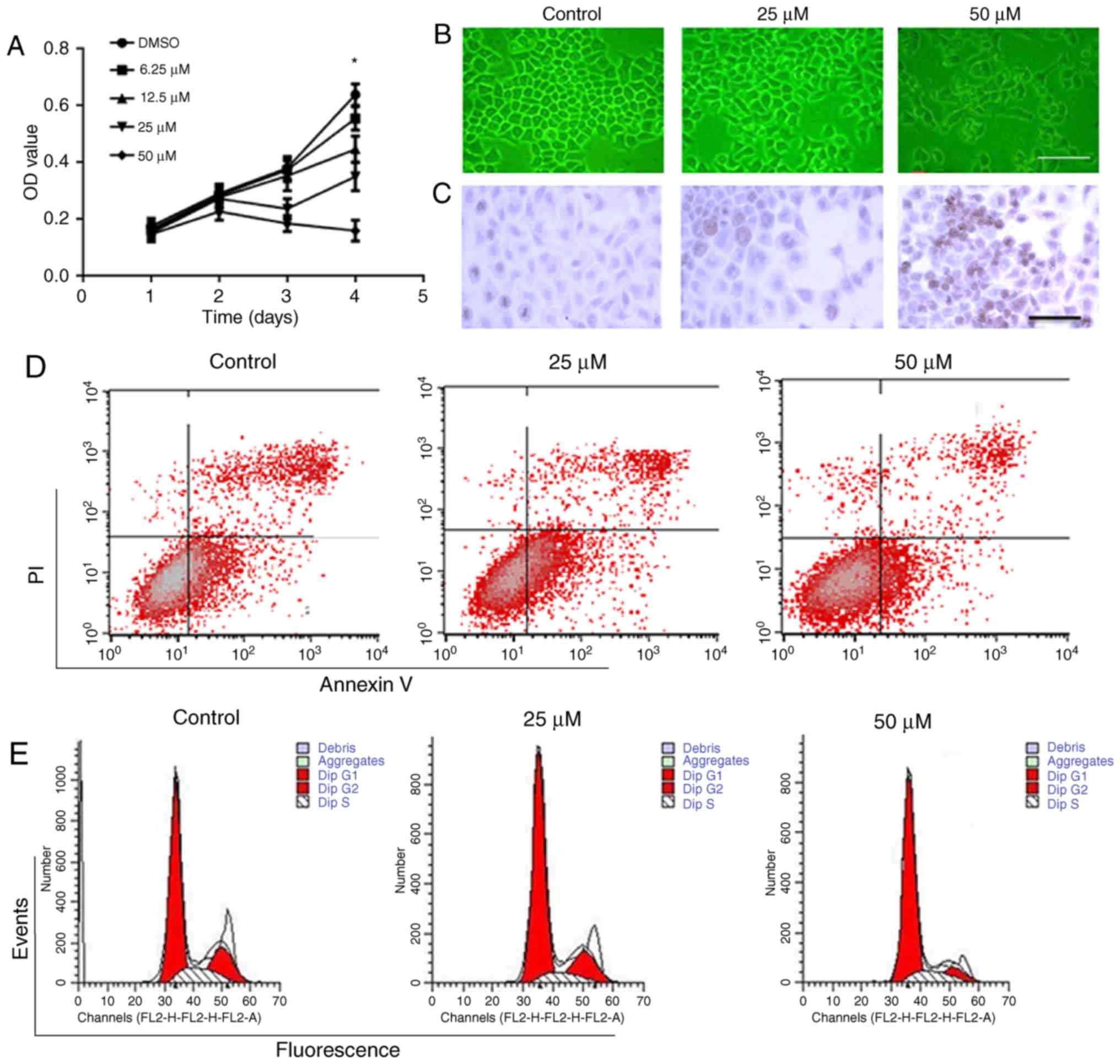
HT29 cells. As shown in Fig. 9A,
DAPT at the concentration range of 6.25–50 µM inhibited the cell
proliferation in a concentration-dependent manner. After incubation
for 4 days, DAPT at 12.5–50 µM significantly inhibited cell
proliferation compared with control. DAPT treatment (50 µM) for 48
h impaired cell morphology and structure (Fig. 9B). TUNEL assay showed that DAPT
promoted apoptosis at the concentrations of 25 and 50 µM (Fig. 9C and Table I). Compared with control, there was
significant difference regarding apoptotic rate in 25 and 50 µM
DAPT groups. DAPT-induced apoptosis in HT29 cells were also
confirmed by flow cytometry after Annexin V-FITC/PI double staining
(Fig. 9D). DAPT treatment (25 and
50 µM) for 48 h not only promoted apoptotic rate in early phase,
but also total apoptotic rate compared with control (Fig. 9D and Table II). Additionally, we found DAPT
arrested the cell cycle at G1 phase after 48-h treatment (Fig. 9E and Table III).
 | Table I.DAPT triggers apoptosis in HT29 cells
after administration for 48 h. |
Table I.
DAPT triggers apoptosis in HT29 cells
after administration for 48 h.
| Groups | Control | 25 µM DAPT | 50 µM DAPT |
|---|
| Apoptosis (%) |
4.22±2.51 |
17.61±2.82a |
25.34±5.28a |
| Table II.DAPT triggers apoptosis in HT29 cells
after administration for 48 h. |
Table II.
DAPT triggers apoptosis in HT29 cells
after administration for 48 h.
| Groups | Control | 25 µM DAPT | 50 µM DAPT |
|---|
| Apoptosis in early
phase (%) |
12.44±0.99 |
25.30±0.74a |
32.05±0.80a |
| Total apoptosis
(%) |
20.48±1.57 |
36.43±1.42a |
43.24±1.67a |
| Table III.DAPT causes cell cycle arrest in HT29
cells after administration for 48 h. |
Table III.
DAPT causes cell cycle arrest in HT29
cells after administration for 48 h.
| Groups | Control | 25 µM DAPT | 50 µM DAPT |
|---|
| G1 phase (%) |
64.813±2.525 |
71.517±1.562a |
75.797±1.251a |
| S phase (%) |
18.456±1.364 |
12.369±1.014 |
10.089±0.897 |
| G2 phase (%) |
16.731±1.145 |
16.114±1.125 |
14.114±0.974 |
Discussion
Notch signaling pathway plays key roles in
determining cell fate in multicellular organisms (20,21).
Transgenic animal model showed that the fate of intestinal
epithelial cells was directly regulated by Notch (22). However, the relationship between the
abnormal expression of Notch-1 and colorectal cancer is not clear.
Our study showed that Notch-1 expression in colorectal cancer was
significantly higher than those of paracancerous tissues and normal
colorectal tissues, which was consistent with a previous study
(23). In this study, we also found
for the first time that the expression of Jagged1 in colorectal
cancer was increased compared with paracancerous tissues and normal
colorectal tissues. These results suggest that Notch-1 signaling
pathway is involved in the pathogenesis of colorectal cancer.
However, the expression was not related to the cancer grades,
suggesting that Notch signaling is involved in carcinogenesis of
colorectal cancer even at the early phase. A large number of
studies have shown that Notch signaling pathway, especially
Notch-1, is closely related to tumorigenesis (24–29).
Immunocytochemical staining showed that Notch-1 was expressed in
all five colorectal cancer cells. These results were confirmed by
RT-PCR and western blotting methods. Interestingly, Notch-1 is
highly expressed in HT29, SW480, COLO205 and SW1116, while LoVo
cells showed relatively low expression. The in vitro data
further confirmed that Notch signaling is related to the
development of colorectal cancer.
Gene therapy is a hot topic in the research of tumor
therapy. RNA interference (RNAi) is a promising method in the study
of gene function (30). RNAi
technology is simple, fast and effective and could specifically
inhibit the expression of target genes. Therefore, this technique
is now a powerful tool to study gene function, but also provides
new technology for the treatment of cancers (31). In our study, PSilencer 5.1-H1 Retro
source was derived from murine stem cell virus (MSCV), a retroviral
vector carrying siRNA fragment. The total length was 6253 bp, and
the promoter is started by H1, with BamHI and HindIII
restriction sites. Therefore, the viruses can effectively transfect
mammalian cells. Due to the resistance to puromycin, it is
convenient to use the cells to express the recombinant DNA. In
order to clarify the role of Notch signaling pathway performed in
the proliferation of colorectal cancer, we selected HT29 cells and
knocked down Notch-1 expression. Additionally, Notch signaling
pathway inhibitor DAPT was also utilized to confirm the effects of
Notch signaling pathway on proliferation of colorectal cancer
cells. MTT assay showed that the growth and proliferation of HT29
cells were suppressed after Notch-1 knockdown, accompanied with
cell morphological changes of cell death. These results suggest
that Notch-1 signaling can promote the growth and proliferation of
colorectal cancer cells. DAPT is a chemical molecule that
specifically inhibits the activity of γ-secretase. Therefore, DAPT
can block the cleavage of Notch receptor/ligand binding and inhibit
all Notch signals. We treated HT29 cells with DAPT and results
showed that DAPT treatment could slow down HT29 cell proliferation
in a concentration- and time-dependent manner. Interestingly, a
small dose of DAPT could promote chemotherapy in colon cancer cells
(32). Mechanically, Notch-1
knockdown or inhibition of Notch signaling pathway is able to
inhibit cell cycle and arrest the cell cycle at G0/G1 phase, and
induce apoptosis in other tumors such as malignant melanoma and
pancreatic cancer. In our study, we also revealed that Notch
signaling pathway inhibition might suppress the growth and
proliferation of colorectal cancer cells by affecting cell cycle
and inhibiting cell apoptosis.
p21 protein has function in arresting cell cycle by
inhibiting Cyclin-dependent kinase (CDK) complexes, such as cyclin
A-CDK2, cyclin D-CDK4/CDK6 and cyclin E/CDK2. p21 expression can
block cell cycle at G1 phase, G2 phase or S phase (33). We used RT-PCR method to detect the
effects of Notch-1 interference on the expression of p21 mRNA in
HT29 cells. The results showed that Notch-1 silence increased
expression of p21. Therefore, we hypothesized that Notch-1
knockdown may lead to cell cycle arrest by inhibiting the
expression of p21, thereby inhibiting the growth of colorectal
cancer cells. Sjölund et al (34) also found that Notch-1 knockout can
inhibit the growth of renal cell carcinoma in vitro and
in vivo, and arrest the cell cycle at G0/G1 phase. p21 not
only inhibits the cell cycle, but also promotes apoptosis in some
conditions. For example, trichostatin A can upregulate p21 to
induce apoptosis of osteoclasts (35). Zoli et al found that the
expression of p21 was upregulated and was positively correlated
with apoptosis of breast cancer treated with adriamycin paclitaxel
combined with 5-fluorouracil (36).
Therefore, we assumed that Notch-1 signaling pathway activation in
colorectal cancer cells may suppress apoptosis of HT29 cells by
downregulating the expression of p21.
PUMA is a pro-apoptotic gene and one member of
apoptosis-related proteins in the Bcl-2 family, through its
interaction with Bcl-2/ Bcl-XL (37). Previous results showed that
transfection of PUMA into human melanoma cell line can lead to cell
apoptosis and death in p53-dependent or -independent pathways
(38). Our study found that the
expression of PUMA was upregulated after Notch-1 interference in
HT29 cells. These results implicated that the interference of
Notch-1 could promote apoptosis by upregulating the expression of
PUMA. Stylianou et al found that chemotherapy by
mitoxantrone was related to Notch-regulated PUMA and apoptosis in
breast cancer cell lines (39).
These results suggest that Notch signaling can regulate the cell
cycle and cell apoptosis by regulating the p21 and PUMA genes, and
thus plays a role in promoting cancer in colorectal cancer.
In our study, we verified Notch-1 expression in
cancer tissue and cell lines. In the next part of experiments, HT29
cells were selected because of its high expression of Notch-1.
Several siRNAs were designed to knock down Notch-1 and one optimal
siRNA was packaged in the virus. Moreover, the role of Notch-1 was
further confirmed by DAPT. In future studies, more colorectal
cancer cell lines will be selected to clarify the mechanisms.
In conclusion, Notch-1 is an oncogene in colorectal
cancer cells. Its overexpression is related to the cancer
development. Moreover, genetic method to knock down Notch-1 or
Notch signaling pathway inhibitor can effectively suppress the
growth and proliferation of colorectal cancer cells, and promote
cell apoptosis. These results indicate that Notch-1 could serve as
a therapy target for colorectal cancer.
Acknowledgements
This study was supported by Natural Science
Foundation of Jiangxi Province (20122BAB205018) and Foundation of
Jiang'xi Educational Committee (GJJ11328).
References
|
1
|
Eadens MJ and Grothey A: Curable
metastatic colorectal cancer. Curr Oncol Rep. 13:168–176. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Li L, Li YM, Zhou P, Wang XS, Wang GY,
Zhao XH, Cui BB, Ren YL, Dong XS and Chen ZQ: Abnormal expression
of p190RhoGAP in colorectal cancer patients with poor survival. Am
J Transl Res. 8:4405–4414. 2016.PubMed/NCBI
|
|
3
|
Huang T, Zhou Y, Cheng AS, Yu J, To KF and
Kang W: NOTCH receptors in gastric and other gastrointestinal
cancers: Oncogenes or tumor suppressors? Mol Cancer. 15:802016.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhang M, Biswas S, Qin X, Gong W, Deng W
and Yu H: Does Notch play a tumor suppressor role across diverse
squamous cell carcinomas? Cancer Med. 5:2048–2060. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tarulli GA, Butler LM, Tilley WD and
Hickey TE: Bringing androgens up a NOTCH in breast cancer. Endocr
Relat Cancer. 21:T183–T202. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Leong KG and Karsan A: Recent insights
into the role of Notch signaling in tumorigenesis. Blood.
107:2223–2233. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Koch U and Radtke F: Notch and cancer: A
double-edged sword. Cell Mol Life Sci. 64:2746–2762. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Radtke F and Raj K: The role of Notch in
tumorigenesis: Oncogene or tumour suppressor? Nat Rev Cancer.
3:756–767. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Talora C, Sgroi DC, Crum CP and Dotto GP:
Specific down-modulation of Notch1 signaling in cervical cancer
cells is required for sustained HPV-E6/E7 expression and late steps
of malignant transformation. Genes Dev. 16:2252–2263. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Collins BJ, Kleeberger W and Ball DW:
Notch in lung development and lung cancer. Semin Cancer Biol.
14:357–364. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li S, Hu H, He Z, Liang D, Sun R and Lan
K: Fine-tuning of the Kaposi's sarcoma-associated herpesvirus life
cycle in neighboring cells through the RTA-JAG1-Notch pathway. PLoS
Pathog. 12:e10059002016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li L, Grausam KB, Wang J, Lun MP, Ohli J,
Lidov HG, Calicchio ML, Zeng E, Salisbury JL, Wechsler-Reya RJ, et
al: Sonic Hedgehog promotes proliferation of Notch-dependent
monociliated choroid plexus tumour cells. Nat Cell Biol.
18:418–430. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cigliano A, Wang J, Chen X and Calvisi DF:
Role of the Notch signaling in cholangiocarcinoma. Expert Opin Ther
Targets. 21:471–483. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Feng Z, Xu W, Zhang C, Liu M and Wen H:
Inhibition of gamma-secretase in Notch1 signaling pathway as a
novel treatment for ovarian cancer. Oncotarget. 8:8215–8225.
2017.PubMed/NCBI
|
|
15
|
Schröder N and Gossler A: Expression of
Notch pathway components in fetal and adult mouse small intestine.
Gene Expr Patterns. 2:247–250. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sander GR and Powell BC: Expression of
notch receptors and ligands in the adult gut. J Histochem Cytochem.
52:509–516. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Stanger BZ, Datar R, Murtaugh LC and
Melton DA: Direct regulation of intestinal fate by Notch. Proc Natl
Acad Sci USA. 102:pp. 12443–12448. 2005; View Article : Google Scholar : PubMed/NCBI
|
|
18
|
van Es JH, van Gijn ME, Riccio O, Van den
Born M, Vooijs M, Begthel H, Cozijnsen M, Robine S, Winton DJ,
Radtke F, et al: Notch/gamma-secretase inhibition turns
proliferative cells in intestinal crypts and adenomas into goblet
cells. Nature. 435:959–963. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li J, Chen H, Wu S, Cheng Y, Li Q, Wang J
and Zhu G: MPP(+) inhibits mGluR1/5-mediated long-term depression
in mouse hippocampus by calpain activation. Eur J Pharmacol.
795:22–27. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Artavanis-Tsakonas S, Rand MD and Lake RJ:
Notch signaling: Cell fate control and signal integration in
development. Science. 284:770–776. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ulvklo C, MacDonald R, Bivik C, Baumgardt
M, Karlsson D and Thor S: Control of neuronal cell fate and number
by integration of distinct daughter cell proliferation modes with
temporal progression. Development. 139:678–689. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fre S, Huyghe M, Mourikis P, Robine S,
Louvard D and Artavanis-Tsakonas S: Notch signals control the fate
of immature progenitor cells in the intestine. Nature. 435:964–968.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chu D, Wang W, Xie H, Li Y, Dong G, Xu C,
Chen D, Zheng J, Li M, Lu Z, et al: Notch-1 expression in
colorectal carcinoma determines tumor differentiation status. J
Gastrointest Surg. 13:253–260. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fabbri G, Holmes AB, Viganotti M, Scuoppo
C, Belver L, Herranz D, Yan XJ, Kieso Y, Rossi D, Gaidano G, et al:
Common nonmutational NOTCH1 activation in chronic lymphocytic
leukemia. Proc Natl Acad Sci USA. 114:pp. E2911–E2919. 2017;
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Fujiki K, Inamura H, Miyayama T and
Matsuoka M: Involvement of Notch1 signaling in malignant
progression of A549 cells subjected to prolonged cadmium exposure.
J Biol Chem. 292:7942–7953. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Feng HB, Wang J, Jiang HR, Mei X, Zhao YY,
Chen FR, Qu Y, Sai K, Guo CC, Yang QY, et al: β-Elemene selectively
inhibits the proliferation of glioma stem-like cells through the
downregulation of Notch1. Stem Cells Transl Med. 6:830–839. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wieland E, Rodriguez-Vita J, Liebler SS,
Mogler C, Moll I, Herberich SE, Espinet E, Herpel E, Menuchin A,
Chang-Claude J, et al: Endothelial Notch1 activityf acilitates
metastasis. Cancer Cell. 31:355–367. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhou YF, Sun Q, Zhang YJ, Wang GM, He B,
Qi T, Zhou Y, Li XW, Li S and He L: Targeted inhibition of Notch1
gene enhances the killing effects of paclitaxel on triple negative
breast cancer cells. Asian Pac J Trop Med. 10:179–183. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kaka AS, Nowacki NB, Kumar B, Zhao S, Old
MO, Agrawal A, Ozer E, Carrau RL, Schuller DE, Kumar P, et al:
Notch-1 overexpression correlates to improved survival in cancer of
the oropharynx. Otolaryngol Head Neck Surg. 156:652–659. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Moore CB, Guthrie EH, Huang MT and Taxman
DJ: Short hairpin RNA (shRNA): Design, delivery, and assessment of
gene knockdown. Methods Mol Biol. 629:141–158. 2010.PubMed/NCBI
|
|
31
|
Han P, Fan J, Liu Y, Cuthbertson AG, Yan
S, Qiu BL and Ren S: RNAi-mediated knockdown of serine protease
inhibitor genes increases the mortality of Plutella xylostella
challenged by destruxin A. PLoS One. 9:e978632014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Aleksic T and Feller SM: Gamma-secretase
inhibition combined with platinum compounds enhances cell death in
a large subset of colorectal cancer cells. Cell Commun Signal.
6:82008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Radhakrishnan SK, Feliciano CS, Najmabadi
F, Haegebarth A, Kandel ES, Tyner AL and Gartel AL: Constitutive
expression of E2F-1 leads to p21-dependent cell cycle arrest in S
phase of the cell cycle. Oncogene. 23:4173–4176. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sjölund J, Johansson M, Manna S, Norin C,
Pietras A, Beckman S, Nilsson E, Ljungberg B and Axelson H:
Suppression of renal cell carcinoma growth by inhibition of Notch
signaling in vitro and in vivo. J Clin Invest. 118:217–228. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yi T, Baek JH, Kim HJ, Choi MH, Seo SB,
Ryoo HM, Kim GS and Woo KM: Trichostatin A-mediated upregulation of
p21(WAF1) contributes to osteoclast apoptosis. Exp Mol Med.
39:213–221. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zoli W, Ulivi P, Tesei A, Fabbri F,
Rosetti M, Maltoni R, Giunchi DC, Ricotti L, Brigliadori G, Vannini
I, et al: Addition of 5-fluorouracil to doxorubicin-paclitaxel
sequence increases caspase-dependent apoptosis in breast cancer
cell lines. Breast Cancer Res. 7:R681–R689. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hasenjäger A, Gillissen B, Müller A,
Normand G, Hemmati PG, Schuler M, Dörken B and Daniel PT: Smac
induces cytochrome c release and apoptosis independently from
Bax/Bcl-x(L) in a strictly caspase-3-dependent manner in human
carcinoma cells. Oncogene. 23:4523–4535. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Karst AM, Dai DL, Martinka M and Li G:
PUMA expression is significantly reduced in human cutaneous
melanomas. Oncogene. 24:1111–1116. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Stylianou S, Clarke RB and Brennan K:
Aberrant activation of notch signaling in human breast cancer.
Cancer Res. 66:1517–1525. 2006. View Article : Google Scholar : PubMed/NCBI
|