Introduction
Thyroid cancer is a common endocrine malignancy with
continuously increasing incidence in the past decade (1). Globally in 2012, there were estimated
to be ~230,000 novel cases of thyroid cancer among women and 70,000
among men (2). Thyroid cancer may
be divided into parafollicular C cell-derived medullary thyroid
cancer (MTC) and follicular epithelial cell-derived tumor types,
including papillary thyroid cancer (PTC), follicular thyroid cancer
(FTC), anaplastic thyroid cancer (ATC) and poorly differentiated
thyroid cancer (PDTC). These subtypes of thyroid cancer exhibit
different phenotypes, clinical behaviors and molecular mechanisms
(3). The constitutive activity of
the RET proto-oncogene is a well-reported mechanism for MTC
(4), while the molecular
pathogenesis of follicular epithelial cell-derived thyroid cancer
includes genetic and epigenetic alterations, including mutation,
gene copy-number gain, aberrant gene expression and
post-translational modification (3,5).
Investigating these molecular alterations may provide novel insight
into potential treatment strategies for this cancer type.
Kruppel-like factor 5 (KLF5) is a critical member of
the KLF family (6). The KLF5 gene
is located at chromosome 13q21, which was frequently deleted in
human cancer types (7,8). KLF5 protein contains a proline rich
transactivation domain and three zinc-finger domains (9,10). A
number of studies have demonstrated the association between KLF5
and human malignancies. Du et al (11) reported that KLF5 promoted cell
migration and increased the lamellipodia formation of bladder
cancer. In hepatocellular carcinoma, KLF5 was upregulated in
CD44C/CD133C cells and was essential for the regulation of cancer
stem cells (12). However, evidence
in prostate cancer demonstrated that KLF5 loss promoted tumor
angiogenesis through inhibiting phosphoinositide 3-kinase
(PI3K)/protein kinase B (AKT) signaling (13). Thus, the biological function of KLF5
remains controversial and may be tumor-type dependent.
The involvement of KLF5 in thyroid cancer remains
largely unknown. The present study aimed to clarify the biological
roles and mechanisms of KLF5 in thyroid cancer.
Materials and methods
Clinical data
The Ethics Committee of Zhengzhou University
(Zhengzhou, China) ethically approved the study protocol, and
written informed consent was obtained from each participant prior
to enrollment in the study. Between January 2015 and December 2016,
98 paired thyroid cancer tissues (24 males and 74 females; age
range, 23–54 years; median age, 37 years) were collected from The
First Affiliated Hospital of Zhengzhou University. Patients, who
received preoperative treatment including radiation or
chemotherapy, or had tumor types of other organs, were excluded
from the cohort used in the present study. All cases were
clinically and histologically diagnosed.
Immunohistochemistry (IHC) assay
The samples were fixed with 4% buffered formaldehyde
for 48 h at room temperature and embedded in paraffin. Then a
tissue microarray containing 98 paired thyroid cancer tissues and
adjacent non-cancerous tissues were constructed. For the IHC assay,
the sections (4 µm) were deparaffinized, rehydrated and heated in
citric buffer at 100°C for antigen retrieval. Endogenous peroxidase
activity was inactivated by adding 3% hydrogen peroxide for 5 min
at room temperature. Then the sections were incubated with
anti-KLF5 (1:200; cat. no. 21017-1-AP; ProteinTech Group, Inc.,
Chicago, IL, USA) at 4°C overnight. The next day, sections were
washed three times with PBS, incubated with goat anti-rabbit
immunoglobulin G (IgG) second antibodies (dilution 1:1,000; cat.
no. ab6720; Abcam, Cambridge, MA, USA) at 37°C for 30 min and
visualized using the Metal Enhanced DAB Substrate kit (Thermo
Fisher Scientific, Inc., Waltham, MA, USA) according to the
manufacturer's protocol. Finally, the sections were counterstained
with hematoxylin (0.5% for 1 min at room temperature) and
dehydrated with graded concentrations of ethanol (70% ethanol for 2
min, 95% ethanol for 2 min and absolute ethanol for 2 min) and
dimethyl benzene.
Scoring was performed by two investigators of The
First Affiliated Hospital of Zhengzhou University (Dr Qingzhu Wang
and Dr Fei Liu) independently, and discrepancies were resolved by
consensus with another researcher (Dr Lina Wu). The proportion of
positive cells was scored as 0 (<10%), 1 (10–25%), 2 (26–50%) or
3 (>50 %); staining intensity was scored as 0 (no staining), 1
(weak staining), 2 (moderate staining) or 3 (strong staining). The
final score of each case was determined by multiplying the
proportion and intensity scores. For statistical analysis, cases
were divided into KLF5 low expression (0–3) or high expression
(4–9) groups.
Cell culture
Human thyroid cancer cell lines SW579 and B-CPAP
were obtained from the Type Culture Collection of the Chinese
Academy of Sciences (Shanghai, China). B-CPAP was originally
classified as a PTC cell line but is now considered to be a PDTC
cell line (14). SW579 was a
squamous cell carcinoma cell line of human thyroid cancer. Cells
were cultured in Dulbecco's modified Eagle's medium (DMEM; Hyclone;
GE Healthcare Life Sciences, Logan, UT, USA) supplemented with 10%
fetal bovine serum (FBS; Hyclone; GE Healthcare Life Sciences) in a
humidified incubator at 37°C with 5% CO2.
Transient transfection
Small interfering RNAs (siRNAs) targeting KLF5 and
the scrambled (scrRNA) siRNAs were synthesized by Shanghai
GenePharma Co., Ltd. (Shanghai, China). KLF5 or F-box/WD
repeat-containing protein 7 (FBW7) expressing plasmids and the
empty vector were obtained from Shanghai GeneChem Co., Ltd.
(Shanghai, China). For transient transfection, 3×105
SW579/B-CPAP cells were plated into 6-well plates until they
reached 70% confluence. Then cells were transfected (48 h at 37°C)
with 8 μl siRNA/4 μg DNA and 10 µl Lipofectamine 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
protocol. RNA/protein extraction and all the functional studies
were conducted 48 h after transfection. The siRNA target sequences
were: siRNA#1, 5′-CGAUUACCCUGGUUGCACA-3′; siRNA#2,
5′-AAGCUCACCUGAGGACUCA-3′. To stably silence KLF5,
pLVshRNA-EGFP(2A)Puro vectors (Cyagen US Inc., Santa Clara, CA,
USA) expressing short hairpin RNA (shRNA/sh) negative control
(shNC), shKLF5#1 (target sequence: 5′-CGATTACCCTGGTTGCACA-3′) or
shKLF5#2 (target sequence: 5′-AAGCTCACCTGAGGACTCA-3′) were
established. Then, 3×105 SW579 cells were seeded in to a
6-well plate and transient transfection (48 h at 37°C) was
performed using Lipofectamine 2000 when the cells reached 70%
confluence. Stable cells were selected with culture medium
containing puromycin (5 µg/ml; Beyotime Institute of Biotechnology,
Haimen, China). Stable B-CPAP/NC and B-CPAP/KLF5 colonies were
obtained by G418 selection (400 µg/ml for 10 days; Beyotime
Institute of Biotechnology) following transient transfection. The
knockdown/overexpression efficacy of KLF5 was verified using a
western blot assay.
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA of the clinical samples was isolated with
TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.) and
cDNA was synthesized using a RT kit (Promega Corporation, Madison,
WI, USA) according to the manufacturer's protocol. qPCR was
conducted using an Applied Biosystems 7900 System and the SYBR
Green Reagent kit (Applied Biosystems; Thermo Fisher Scientific,
Inc.) to quantitatively determine the mRNA levels of KLF5 according
to the manufacturer's protocol. The PCR primers used in the present
study were as follows: GAPDH forward, 5′-GGTGAAGGTCGGAGTCAACG-3′
and reverse, 5′-TGGGTGGAATCATATTGGAACA-3′; KLF5 forward,
5′-ACACCAGACCGCAGCTCCA-3′ and reverse,
5′-TCCATTGCTGCTGTCTGATTTGTAG-3′. The thermocycling conditions were
as follows: Precubation at 95°C for 10 min; amplification at 95°C
for 10 sec, 60°C for 10 sec and 72°C for 10 sec for 45 cycles;
melting at 95°C for 10 sec, 65°C for 60 sec and 97°C for 1 sec. The
fold amplification for gene expression was determined using the
2−∆∆Cq method (15).
Western blotting
SW579/B-CPAP cells were lysed (30 min at 4°C) with
RIPA buffer (Beijing Solarbio Science & Technology Co., Ltd.,
Beijing, China) in the presence of a protease inhibitor cocktail
(Sigma Aldrich; Merck KGaA, Darmstadt, Germany) and phosphatase
inhibitor Cocktail III (Cell Signaling Technology, Inc., Danvers,
MA, USA). Protein concentrations were determined using a BCA
Protein Assay kit (Pierce; Thermo Fisher Scientific, Inc.). The
proteins (20 μg per lane) were resolved using 10% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and then
transferred to polyvinylidene fluoride membranes (EMD Millipore,
Billerica, MA, USA). The membranes were blocked with 5% bovine
serum albumin (1 h at room temperature) and incubated with the
corresponding primary antibodies including KLF5 rabbit polyclonal
antibody (dilution 1:1,000; cat. no. 21017-1-AP; ProteinTech
Group., Inc.), FBW7 mouse monoclonal antibody (dilution 1:1,000;
cat. no. ab74054; Abcam), nuclear matrix protein p84 mouse
monoclonal antibody (dilution 1:1,000; cat. no. ab487; Abcam),
fibronectin rabbit polyclonal antibody (dilution 1:1,000; cat. no.
ab2413; Abcam), E-Cadherin rabbit mAb (dilution 1:1,000; cat. no.
3195; Cell Signaling Technology, Inc.), Vimentin rabbit mAb
(dilution 1:1,000; cat. no. 5741; Cell Signaling Technology, Inc.),
Twist family BHLH transcription factor 1 (Twist1) rabbit mAb
(dilution 1:1,000; cat. no. 46702; Cell Signaling Technology,
Inc.), IκBα Mouse mAb (dilution 1:1,000; cat. no. 4814; Cell
Signaling Technology, Inc.), phosphorylated (p)-IκBα rabbit mAb
(dilution 1:1,000; cat. no. 2859; Cell Signaling Technology, Inc.),
inhibitor of nuclear factor NF-κB subunit β (IKKβ) rabbit mAb
(dilution 1:1,000; cat. no. 8943; Cell Signaling Technology, Inc.),
p-IKKβ rabbit mAb (dilution 1:1,000; cat. no. 2697; Cell Signaling
Technology, Inc.), NF-κB p65 rabbit mAb (dilution 1:1,000; cat. no.
8242; Cell Signaling Technology, Inc.), p44/42 extracellular
signal-regulated kinase 1 and 2 (Erk1/2) rabbit mAb (dilution
1:1,000; cat. no. 4695; Cell Signaling Technology, Inc.), p-p44/42
Erk1/2 rabbit mAb (dilution 1:1,000; cat. no. 4370; Cell Signaling
Technology, Inc.), AKT rabbit mAb (dilution 1:1,000; cat. no. 4685;
Cell Signaling Technology, Inc.), p-AKT rabbit mAb (dilution
1:1,000; cat. no. 4060; Cell Signaling Technology, Inc.) and GAPDH
mouse monoclonal antibody (dilution 1:5,000; cat. no. 60004-1-Ig;
ProteinTech Group., Inc.) at 4°C overnight. The membranes were
washed with tris-buffered saline with Tween-20 (0.1%) at room
temperature (three times for 10 min), and then incubated with
horseradish peroxidase (HRP)-conjugated goat anti-rabbit IgG
(dilution 1:5,000; cat. no. SA00001-2; ProteinTech Group., Inc.) or
HRP-conjugated goat anti-mouse IgG secondary antibodies (dilution
1:5,000; cat. no. SA00001-1; ProteinTech Group., Inc.). Finally,
the protein bands were visualized and quantified by using the
electrochemiluminesence reagent (Thermo Fisher Scientific, Inc.)
and Tanon 5200 Multi Fully Automatic Chemiluminescence/Fluorescence
Image Analysis System which includes a Gel Image system software
(version 4.2.5; Tanon Science and Technology Co., Ltd., Shanghai,
China).
Co-immunoprecipitation (Co-IP)
For the Co-IP assay, lysates of SW579 cells were
incubated with a protein A/G agarose (Beyotime Institute of
Biotechnology) at 4°C for 1 h to remove non-specific hybrid
proteins. Whole cell lysates obtained by centrifugation (12,000 × g
at 4°C for 5 min) were incubated with 2 µg KLF5 rabbit polyclonal
antibody (dilution 1:200; cat. no. 21017-1-AP; ProteinTech Group.,
Inc.) or the control rabbit IgG (dilution 1:200; cat. no. A7016;
Beyotime Institute of Biotechnology), FBW7 mouse monoclonal
antibody (dilution 1:200; cat. no. ab74054; Abcam) or the control
mouse IgG (dilution 1:200; cat. no. A7028; Beyotime Institute of
Biotechnology) at 4°C overnight. Then the samples were incubated
with protein A/G agarose (Beyotime Institute of Biotechnology) for
4 h at 4°C to capture the antigen-antibody complex. Following
centrifugation at 12,000 × g at 4°C for 5 sec, the supernatants
were removed. Then, the agarose bead-antigen-antibody complex was
washed 3 times with pre-cooled PBS. The bound proteins were boiled
in SDS sample buffer and resolved by 10% SDS-PAGE for a western
blotting assay.
Cell proliferation assay
A Cell Counting Kit-8 (CCK-8; Dojindo Molecular
Technologies, Inc., Kumamoto, Japan) assay was used to assess the
cell proliferation ability. A total of 1,000 cells were plated and
adhered onto 96 well plates and optical density values at 450 nm
were measured subsequent to adding CCK-8 (incubation for 2 h at
37°C) on day 1, 2, 3, 4, 5 and 6. B-CPAP/KLF5 cells were cultured
with medium containing NF-κB inhibitor SC75741 (5 μM; Selleck
Chemicals, Houston, TX, USA) or vehicle [dimethyl sulfoxide (DMSO;
Sangon Biotech Co., Ltd., Shanghai, China)] for 1, 2, 3, 4, 5 and 6
days (at 37°C) to evaluate the influence of NF-κB on cell
proliferation.
Anchorage-independent growth assay in
soft agar
For the anchorage-independent growth assay,
low-melting point agarose was soluted in diluted water at
concentrations of 1.2 and 0.6%. The agarose solutions were then
incubated in a thermostat water bath at 40°C for 12 h following
autoclaving. Agarose (1.2%) and 2X DMEM medium (with 20% FBS) were
mixed at a ratio of 1:1, and 3 ml of the mixture was injected into
a 6-well plate. Then 0.6% agarose (1.5 ml) and 2X DMEM medium
containing 2×104 cells (1.5 ml) were mixed at a ratio of
1:1, and the mixture was injected into the upper layer once the
basal layer dried. Colonies were photographed and counted 2–3 weeks
later. Upper agar containing NF-κB inhibitor SC75741 (5 µM; Selleck
Chemicals) or vehicle (DMSO) was used to evaluate the influence of
NF-κB on anchorage-independent growth.
Transwell assay
Cell migration and Matrigel invasion assays were
performed using Transwell inserts (Corning Life Science).
SW579/B-CPAP cells (1×105) in 200 µl serum-free DMEM
were placed in the upper chamber and the lower compartments were
filled with 600 µl DMEM containing 10% FBS. After 48 h culturing at
37°C, tumor cells in the upper side of chamber were removed with
cotton swabs and tumor cells in the lower chamber were stained with
0.5% crystal violet for 30 min at room temperature. Migrated cells
were photographed under an Olympus inverted microscope (Olympus
Corporation, Tokyo, Japan) for counting. To evaluate the influence
of NF-κB on cell migration/invasion, B-CPAP/KLF5 cells were plated
into the upper chamber of the Transwell and complete medium
containing NF-κB inhibitor (SC75741, 5 µM) or vehicle (DMSO) were
added into the lower compartments.
Immunofluorescence (IF)
A total of 1×104 SW579/B-CPAP cells were seeded onto
slides, fixed with 4% paraformaldehyde at room temperature for 10
min, permeabilized with 0.5% NP-40 (Sigma-Aldrich; Merck KGaA) and
blocked with 10% normal goat serum (Beijing Solarbio Science &
Technology Co., Ltd.) for 20 min at room temperature. Then the
slides were incubated with NF-κB p65 Rabbit monoclonal antibody
(mAb; dilution 1:400; cat. no. 8242; Cell Signaling Technology,
Inc.) at 4°C overnight. Subsequent to three washes using PBS for 5
min, the slides were incubated with Alexa Fluor® 555
Conjugate secondary antibody (dilution 1:200; cat. no. 4413; Cell
Signaling Technology, Inc.) for 1 h at 37°C. Finally, the slides
were incubated with DAPI (dilution 1:1,000; cat. no. C1002;
Beyotime Institute of Biotechnology) for 10 min at 37°C. For the
cytoskeleton assay, the slides were incubated with Alexa Fluor 568
dye-conjugated phalloidin (dilution, 1:1,000; cat. no. A12380;
Invitrogen; Thermo Fisher Scientific, Inc.) for 2 h at 37°C and
DAPI (dilution 1:1,000; cat. no. C1002; Beyotime Institute of
Biotechnology) for 10 min at 37°C. Images were obtained using a
fluorescent microscopy (Olympus Corporation).
Protein stability assay
A cycloheximide (CHX) assay was used to determine
the half-life of KLF5. SW579 cells were transfected with an empty
vector or FBW7 expressing plasmid. CHX was added to the cell
culture (at 37°C) at a concentration of 0.1 mg/ml and cells were
harvested at 0.0, 0.5, 1.0 and 2.0 h. Then, cell lysates were
analyzed using western blotting.
In vivo experiments
All experiments were performed strictly in
accordance with a protocol ethically approved by the Institutional
Animal Care and Use Committee of Zhengzhou University. A number of
15 male BALB/c nude mice (4-weeks-old) were obtained from Beijing
Vital River Laboratory Animal Technology Co., Ltd. (Beijing,
China). The mice were maintained under a 12 h dark/light cycle with
ad libitum access to food in specific pathogen-free conditions (55%
humidity and 22°C). SW579/shNC, SW579/shKLF5#1 and SW579/shKLF5#2
cells (1×106 in 100 µl PBS) were subcutaneously injected
into the right flank of nude mice (5 mice per group). Tumor sizes
were measured every 10 days using vernier calipers and the final
volume was calculated using the following formula: V = length ×
width2/2. Mice were sacrificed at the 40th day using
cervical dislocation euthanasia. The subcutaneous xenografts were
weighed, fixed with 4% buffered formaldehyde for 48 h at room
temperature and embedded in paraffin for IHC assays. To evaluate
the effects of KLF5 on metastatic potential, B-CPAP/NC or
B-CPAP/KLF5 cells were intravenously injected (3×106 in
100 µl PBS) into the tail vein of nude mice (5 mice per group). On
day 40, the mice were sacrificed using cervical dislocation. Lungs
were carefully dissected out of the chest and washed with PBS. Then
the metastatic nodules on the lung surface were counted. Then the
lungs of each group were stained with hematoxylin (0.5%) and eosin
(0.05%) for pathological confirmation. The sections (4 µm) were
deparaffinized, rehydrated, stained with hematoxylin for 5 min at
room temperature and then stained with eosin staining solution for
3 min at room temperature. Pictures were taken using an Olympus
inverted microscope (Olympus Corporation).
Statistical analysis
Statistical analysis was performed with SPSS 18.0
(SPSS Inc., Chicago, IL, USA). All data were presented as the mean
± standard deviation. Differences were analyzed with unpaired
Student's t-test between two groups or with one-way analysis of
variance (followed by a Bonferroni post hoc test) among three
groups. The association between KLF5 expression and the
clinicopathological parameters of patients with thyroid cancer were
analyzed using χ2 or Fisher's exact tests. A Spearman's
rank correlation test was used to evaluate the correlation between
KLF5 expression and FBW7 expression. P<0.05 was considered to
indicate a statistically significant difference.
Results
Expression of KLF5 in benign and
malignant thyroid lesions
KLF5 expression in thyroid cancer was determined
using an IHC assay. It was revealed that the protein levels of KLF5
were significantly upregulated in thyroid cancer tissues,
particularly in lymph node metastatic cases, compared with
non-tumor tissues [non-tumor vs. thyroid cancer (LN-), P=0.039;
non-tumor vs. thyroid cancer (LN+), P<0.001; Fig. 1A and B]. The detailed association
between KLF5 expression and the clinicopathological parameters of
patients with thyroid cancer is presented in Table I. KLF5 expression was revealed to be
associated with lymph node metastasis using a χ2 test
(P=0.027). However, no significant differences were observed
between KLF5 expression and other variables, including age, sex,
tumor size and tumor type.
 | Table I.Association of KLF5 expression with
clinicopathological parameters in patients with thyroid cancer. |
Table I.
Association of KLF5 expression with
clinicopathological parameters in patients with thyroid cancer.
|
|
| KLF5
expression |
|
|---|
|
|
|
|
|
|---|
| Variable | Cases (n) | Low | High | P-value |
|---|
| Age (years) |
|
|
| 0.645 |
|
<45 | 63 | 24 | 39 |
|
|
≥45 | 35 | 15 | 20 |
|
| Sex |
|
|
| 0.240 |
|
Male | 24 | 12 | 12 |
|
|
Female | 74 | 27 | 47 |
|
| Tumor size
(cm) |
|
|
| 0.078 |
|
<2 | 70 | 24 | 46 |
|
| ≥2 | 28 | 15 | 13 |
|
| Lymph node
metastasis |
|
|
| 0.027 |
|
Yes | 30 | 7 | 23 |
|
| No | 68 | 32 | 36 |
|
| Tumor types |
|
|
| 0.931 |
|
Papillary | 66 | 26 | 40 |
|
|
Follicular | 27 | 11 | 16 |
|
|
Anaplastic | 5 | 2 | 3 |
|
KLF5 promotes the proliferation and
anchorage-independent growth of thyroid cancer cells
The thyroid cancer cell line SW579 demonstrated
higher levels of KLF5 compared with B-CPAP cells (Fig. 2A). The knockdown efficacy of KLF5 in
SW579 cells and overexpression efficacy of KLF5 in B-CPAP cells
were verified using a western blot assay (Fig. 2A). The result of a CCK-8 assay
revealed that the siRNAs significantly inhibited the proliferation
ability of SW579 cells compared with the scrambled control
(siRNA#1, P=0.002; siRNA#2, P<0.001; Fig. 2B, upper panel), while the
overexpression of KLF5 significantly promoted the proliferation
ability of B-CPAP cells compared with the scrambled control
(P=0.006; Fig. 2B, lower panel). In
the anchorage-independent growth assay, SW579/scrRNA cells induced
a significantly greater number of single-cell-derived colonies
compared with the SW579/siRNA#1 cells or SW579/siRNA#2 cells
(siRNA#1, P<0.001; siRNA#2, P<0.001; Fig. 2C, left panel), while the
overexpression of KLF5 significantly promoted the
anchorage-independent growth of B-CPAP cells compared with the
negative control (NC; P=0.007; Fig.
2C, right panel).
KLF5 promotes the metastatic potential
of thyroid cancer cells
The present study further assessed cell migration
and invasion ability using a Transwell assay. As presented in
Fig. 3A, the knockdown of KLF5 by
the two siRNAs significantly inhibited the migration and invasion
ability of SW579 cells compared with the scrambled control
(migration: SW579/scrRNA vs. SW579/siRNA#1, P<0.001;
SW579/scrRNA vs. SW579/siRNA#2, P<0.001; invasion: SW579/scrRNA
vs. SW579/siRNA#1, P=0.001; SW579/scrRNA vs. SW579/siRNA#2,
P=0.001). In accordance with these results, the knockdown of KLF5
significantly decreased the levels of a number of
metastasis-associated markers, including fibronectin (P<0.01),
Twist1 (P<0.001) and vimentin (P<0.01), while it
significantly promoted the level of metastasis suppressor molecule
E-cadherin (P<0.05; Fig. 3C and
D). Similar results were obtained from IF staining.
SW579/siRNA#1 or SW579/siRNA#2 cells revealed an increased number
of actin stress fibers compared with SW579/scrRNA cells, which
indicated decreased motility (Fig.
3E). The opposite result was observed in B-CPAP/KLF5 cells,
which demonstrated an increased migration and invasion ability
compared with B-CPAP/NC cells (migration, P=0.008; invasion,
P=0.039; Fig. 3B). In addition,
B-CPAP/KLF5 cells demonstrated significantly higher levels of
fibronectin (P<0.01), Twist1 (P<0.01) and vimentin
(P<0.01), significantly lower levels of E-cadherin (P<0.01;
Fig. 3F and G) and fewer actin
stress fibers (Fig. 3H) compared
with B-CPAP/NC cells. Collectively, these data indicated that KLF5
promoted the metastatic potential of thyroid cancer cells.
 | Figure 3.KLF5 promoted the invasive and
metastatic potential of thyroid cancer cells in vitro. (A)
Cell migration/invasion of SW579/scrRNA, SW579/siKLF5#1 and
SW579/siKLF5#2 was analyzed using a Transwell assay. (B) Cell
migration/invasion of B-CPAP/NC and B-CPAP/KLF5 cells was assessed
using a Transwell assay. (C) Western blot analysis of lysates from
SW579/scrRNA, SW579/siKLF5#1 and SW579/siKLF5#2 cells using
anti-E-cadherin, anti-vimentin, anti-Twist1 and anti-fibronectin
and (D) the western blot analysis results quantified. (E)
Immunostaining of phalloidin (F-actin) in SW579/scrRNA,
SW579/siKLF5#1 and SW579/siKLF5#2 cells. (F) Western blot analysis
of lysates from B-CPAP/NC and B-CPAP/KLF5 cells using the indicated
antibodies and (G) the western blot analysis results quantified.
(H) Immunostaining of phalloidin (F-actin) in B-CPAP/NC and
B-CPAP/KLF5 cells. KLF5, Kruppel-like factor 5; scrRNA, scrambled
RNA; si-, small interfering RNA; NC, negative control; Twist1,
Twist family BHLH transcription factor 1. |
KLF5 activates the NF-κB signalling
pathway in thyroid cancer cells
The downstream mechanisms of KLF5 are complicated
and the reported signaling pathways involved include the
extracellular-regulated kinase (ERK) pathway, AKT pathway and the
NF-κB pathway in addition to their crosstalk (13,16–18).
In thyroid cancer cells, KLF5 demonstrated no significant influence
on the activation of EKR or AKT signaling pathways (Fig. 4). However, the NF-κB pathway may be
activated by KLF5 in the two cell lines used in the present study.
As presented in Fig. 5A and B, the
knockdown of KLF5 in SW579 cells significantly decreased the
protein level of p-IκB-α (P<0.001), p-IKK-β (P=0.006) and
nuclear NF-κB p65 (P=0.001). It was additionally observed that the
overexpression of KLF5 in B-CPAP cells significantly promoted the
levels of p-IκB-α (P=0.011), p-IKK-β (P=0.005) and nuclear NF-κB
p65 (P<0.001). Notably, the IF assay for NF-κB p65 in SW579
cells revealed that the knockdown of KLF5 substantially decreased
nuclear NF-κB p65, while the overexpression of KLF5 in B-CPAP cells
promoted nuclear NF-κB p65 (Fig.
5C). In summary, KLF5 promoted NF-κB cytoplasmic-nuclear
translocation in thyroid cancer cells.
 | Figure 4.Effect of KLF5 on the ERK pathway and
the AKT pathway. Western blot analysis of p-ERK, ERK, p-AKT and AKT
in SW579 cells (SW579/scrRNA, SW579/siKLF5#1 and SW579/siKLF5#2)
and B-CPAP cells (B-CPAP/NC and B-CPAP/KLF5). KLF5, Kruppel-like
factor 5; scrRNA, scrambled RNA; si-, small interfering RNA; NC,
negative control; AKT, protein kinase B; p-, phosphorylated; ERK,
extracellular-regulated kinase; NS, not significant. |
 | Figure 5.(A) Western blotting of p-IκB-α,
p-IKK-β and nuclear NF-κB p65 in SW579 cells (SW579/scrRNA,
SW579/siKLF5#1 and SW579/siKLF5#2, left panel) and B-CPAP cells
(B-CPAP/NC and B-CPAP/KLF5, right panel); and p84 was used as a
nuclear marker. (B) Quantified western blotting results. (C)
Immunofluorescent staining of NF-κB p65 in SW579/scrRNA,
SW579/siKLF5#1, SW579/siKLF5#2, B-CPAP/NC and B-CPAP/KLF5 cells.
NF-κB, nuclear factor-κB; p-, phosphorylated-; IKK-β, inhibitor of
nuclear factor κB kinase subunit β; IκB-α, nuclear factor of κ
light polypeptide gene enhancer in B-cells inhibitor, α; scrRNA,
scrambled RNA; si, small interfering RNA; NC, negative control;
KLF5, Kruppel-like factor 5. |
Blocking the NF-κB signalling pathway
abolishes KLF5-induced phenotypes
To investigate the contribution of NF-κB activation
to KLF5-induced phenotypes, B-CPAP/KLF5 cells were treated with the
specific NF-κB inhibitor SC75741. Subsequent to incubation with
SC75741, B-CPAP/KLF5 cells exhibited significantly decreased
nuclear NF-κB p65 (P<0.001; Fig. 6A
and B). Furthermore, the proliferation (P<0.001; Fig. 6C), colony formation (P<0.001;
Fig. 6D), migration (P<0.001)
and invasion (P=0.001; Fig. 6E)
ability of B-CPAP/KLF5+SC75741 cells was significantly reduced
compared with the B-CPAP/KLF5+DMSO group. These data suggest that
KLF5 regulates the aggressive phenotypes of thyroid cancer cells
through activating the NF-κB signaling pathway.
FBW7 is responsible for the
ubiquitination and degradation of KLF5
Although the IHC assay revealed significantly higher
protein levels of KLF5 in thyroid cancer tissues compared with
normal tissues (P<0.001), the results of RT-qPCR revealed no
significant differences in KLF5 mRNA levels between thyroid cancer
tissues and non-tumor tissues (n=98, P>0.05; data not shown).
This result suggested that KLF5 may be regulated by
post-translational mechanisms. A previous study revealed that KLF5
is an unstable protein, which may be regulated by the E3 ubiquitin
ligase FBW7 (19). Thus, the
present study examined the interaction between FBW7 and KLF5 by a
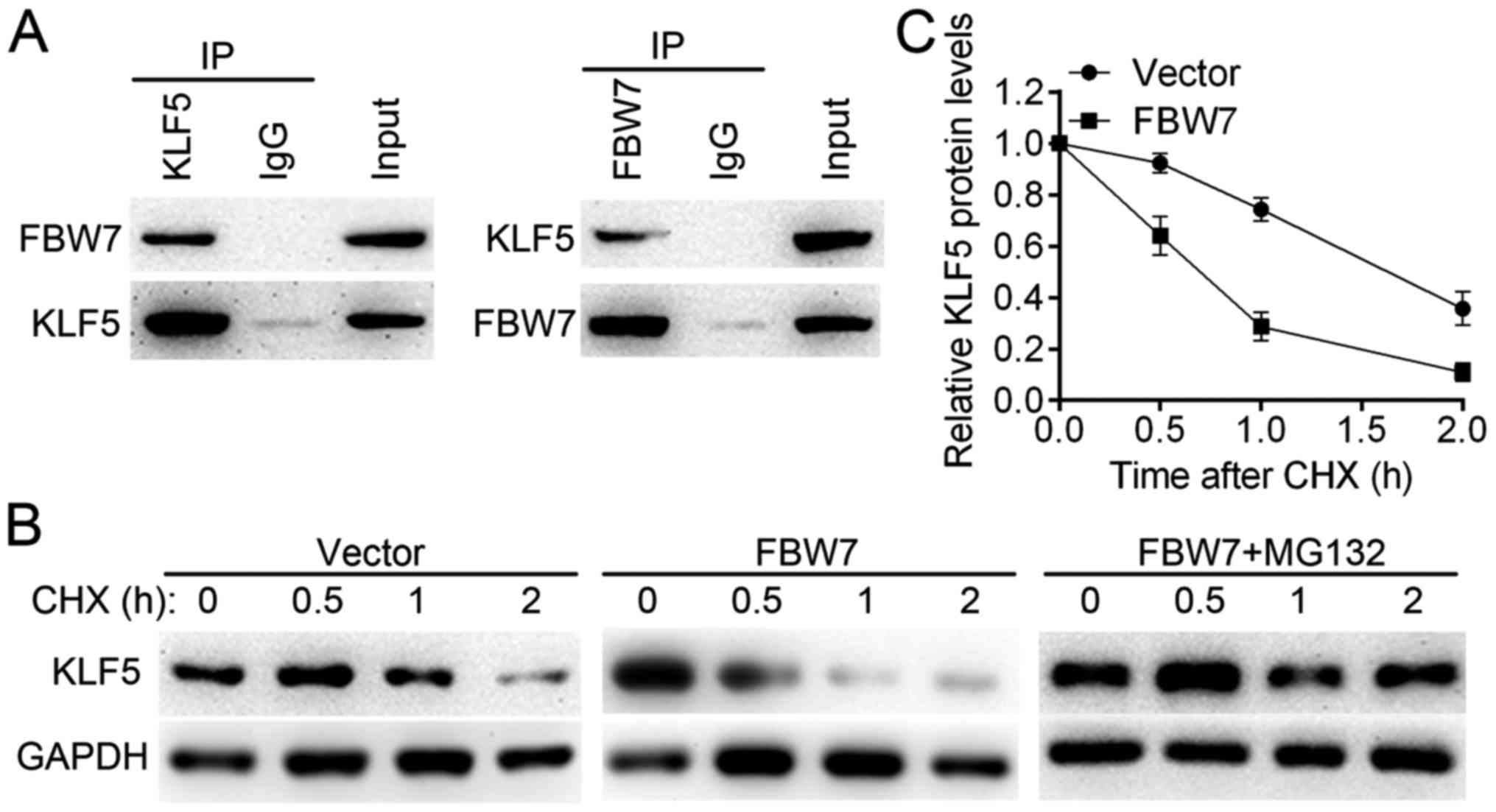
Co-IP assay in SW579 cells (Fig.
7A). It was observed that FBW7 reduced the KLF5 half-life
compared with the control group and that the effect may be blocked
by the proteasome inhibitor MG132 (Fig.
7B and C). These results suggest that FBW7 is an upstream
regulator for KLF5 and may be responsible for the dysregulation of
KLF5 in thyroid cancer cells.
KLF5 expression levels negatively
correlate with FBW7 in tumor tissues
Given that KLF5 was a direct target of FBW7 for
degradation in thyroid cancer cells, the correlation between KLF5
and FBW7 in tumor tissues was further evaluated. As presented in
Fig. 8A, representative tissue
sections revealed that the protein levels of KLF5 and FBW7 were
negatively correlated in thyroid cancer tissues and the results
were summarized in Fig. 8B (r=
−0.435, P<0.001). Collectively, these results revealed that the
FBW7-KLF5-NF-κB p65 axis is critical for the malignant phenotypes
of thyroid cancer (Fig. 8C).
KLF5 promotes in vivo tumor growth and
the metastasis of thyroid cancer cells
To investigate whether KLF5 was able to induce
similar phenotypes in vivo, stable KLF5-silenced cells
(SW579/shNC, SW579/shKLF5#1 and SW579/shKLF5#2) were established
and the knockdown efficacy of KLF5 was verified by western blotting
(Fig. 9A). A subcutaneous xenograft
experiment in nude mice was then performed (Fig. 9B). The growth curves of the tumors
revealed that the knockdown of KLF5 significantly inhibited SW579
cell growth in nude mice (shKLF5#1, P<0.001; shKLF5#2,
P<0.001; Fig. 9C), and the tumor
weight was significantly decreased in the SW579/shKLF5#1 or SW579/
shKLF5#2 group compared with the SW579/shNC group (shKLF5#1,
P<0.001; shKLF5#2, P<0.001; Fig.
9D). Subcutaneous tumor types derived from SW579/shKLF5#1 or
SW579/shKLF5#2 cells demonstrated increased levels of E-cadherin
and decreased levels of KLF5 and ki67 compared with the SW579/shNC
group (Fig. 9E). The results of the
in vivo lung metastasis assay revealed that B-CPAP/KLF5
cells induced a significantly greater number of lung metastatic
nodules in nude mice compared with B-CPAP/NC cells (P=0.002;
Fig. 9F and G), which suggested
KLF5 is capable of manipulating the metastatic ability of B-CPAP
cells in vivo.
Discussion
Extensive local invasion and distant metastasis are
two major causes of recurrence and the mortality of patients with
thyroid cancer (20), but the
mechanisms of these processes remain elusive. In the present study,
it was demonstrated that the protein expression of KLF5 was
upregulated in thyroid cancer tissues compared with non-tumor
tissues. Additionally, the overexpression of KLF5 was correlated
with a higher tumor stage and lymph node metastasis. Thus, it is
suspected that KLF5 may function as an oncogene in thyroid
cancer.
The biological functions of KLF5 were evaluated in
SW579 and B-CPAP cells. Previous studies have indicated that KLF5
has the potential to regulate cell proliferation (21), differentiation (22), metastasis (23) and stem cell characteristics
(24). In the present study, KLF5
loss-of-function and gain-of-function assays were conducted to
determine whether KLF5 was essential for the tumorigenesis of
thyroid cancer cells. It was observed that the knockdown of KLF5 in
SW579 cells significantly inhibited tumorigenesis in vitro
and in vivo, while the overexpression of KLF5 in B-CPAP
cells promoted tumorigenesis in vitro and metastasis in
vivo. These results suggested that KLF5 may be critical for the
unrestricted growth of thyroid cancer cells.
Another major characteristics of tumor cells is
metastatic potential. In the present study, it was revealed that
KLF5 promoted the migration/invasion ability of thyroid cancer
cells. It was also observed that the knockdown of KLF5 decreased
the protein levels of a number of pro-metastatic molecules,
including fibronectin, vimentin and Twist1. However, the level of
E-cadherin, which is generally considered as a metastatic
suppressor accounting for cell adhesion, was upregulated following
KLF5 knockdown. Furthermore, phalloidin staining (red) in SW579
cells expressing siKLF5#1 and siKLF5#2 revealed a significant
reduction in actin stress-fiber filaments, indicating weakened
migratory and invasive capacity in these cells.
The aforementioned in vitro and in
vivo phenotypes prompted further investigation into the
mechanisms of KLF5 in thyroid cancer cells. The molecular
pathogenesis of thyroid cancer involves multiple signalling
pathways, including the PI3K/AKT (25), mitogen activated protein
kinase/extracellular regulated kinase (26), Wnt/β-catenin (27) and NF-κB pathways (28). A previous study indicated that
lipopolysaccharide-induced oxidative stress upregulated KLF5
expression, and thus promoted the phosphorylation and nuclear
translocation of NF-κB p65 (16).
However, it remains unknown whether KLF5 is able to activate the
NF-κB signalling pathway in thyroid cancer cells. Regardless of the
contribution from other mechanisms, the results of the present
study provide strong evidence that the knockdown of KLF5 inhibited,
while the overexpression of KLF5 promoted, the nuclear expression
of NF-κB p65 in thyroid cancer cells. Inhibition of the NF-κB
pathway with a small molecule inhibitor blocked KLF5-mediated
phenotypes including cell growth and metastasis. These results
associated KLF5 with an acknowledged signalling pathway, making
KLF5 an attractive therapeutic target.
KLF5 expression may be regulated by different
mechanisms at the mRNA level and protein level. Fu et al
(21) revealed that DNA
(cytosine-5)-methyltransferase 1-mediated promotor hypermethylation
was responsible for the epigenetic silencing of KLF5 in clear cell
renal cell carcinoma, which induced the frequent downregulation of
KLF5 expression at the mRNA level. Thus, the present study
evaluated the mRNA levels of KLF5 in 36 paired clinical samples.
Unexpectedly, no significant difference in KLF5 mRNA expression was
observed between thyroid cancer tissues and non-tumor tissues. By
screening previously published literature, it was observed that the
level of KLF5 may also be regulated through the
ubiquitin-proteasome system as KLF5 is a short-lived protein
(29,30). The most highly reported protein
responsible for KLF5 degradation was E3 ubiquitin ligase FBW7. Zhao
et al (19) reported that
glycogen synthase kinase 3β inducing the S303 phosphorylation of
KLF5 is essential for FBW7-mediated KLF5 degradation. In colon
cancer cells, FBW7 interacted with KLF5 in a CDC4
phosphodegron-dependent manner and was responsible for
KLF5-mediated cell proliferation (31). Of note, the IHC assay in the present
study revealed that the protein levels of KLF5 in thyroid cancer
tissues were significantly higher compared with non-tumor tissues,
and were negatively correlated with FBW7 levels. Additionally, the
exogenous expression of FBW7 in SW579 cells decreased the KLF5
protein half-life, which may be restored by the proteasome
inhibitor MG132. These results revealed that the incontrollable
expression of KLF5 was induced by the dysregulation of
FBW7-mediated proteasomal degradation. This suggests a potential
regulatory mechanism of KLF5 and requires further study.
In summary, the present study revealed that elevated
expression of KLF5 in thyroid cancer tissues was correlated with
lymph node metastasis. KLF5 promoted cell growth and metastasis of
thyroid cancer cells through the NF-κB signaling pathway. The
present functional and mechanistic investigations suggest that
targeting KLF5 may provide a potential therapeutic strategy for
thyroid cancer.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81570746 and
81770812 to GQ), the Innovation Scientists and Technicians Troop
Construction Projects of Henan Province (grant no. 134200510021 to
GQ) and the Young Foundation of the First Affiliated Hospital of
Zhengzhou University (to LW).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YM and GQ planned the study. YM, QW and FL performed
the experiments. YM, XM and LW performed the data analysis. FG, FH
and SZ participated in the clinical sample collection. All authors
read and approved the manuscript and agreed to be accountable for
all aspects of the research in ensuring that the accuracy or
integrity of any part of the work are appropriately investigated
and resolved.
Ethics approval and consent to
participate
The use of human tissue samples and experimental
protocols were approved by the Medical Ethics Review Committee of
Zhengzhou University (Zhengzhou, China), and written informed
consent was obtained from each participant prior to enrolment in
the study. All experiments were performed strictly in accordance
with a protocol ethically approved by the Institutional Animal Care
and Use Committee of Zhengzhou University.
Patient consent for publication
No identifying patient data were included in the
present study.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
La Vecchia C, Malvezzi M, Bosetti C,
Garavello W, Bertuccio P, Levi F and Negri E: Thyroid cancer
mortality and incidence: A global overview. Int J Cancer.
136:2187–2195. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Xing M: Molecular pathogenesis and
mechanisms of thyroid cancer. Nat Rev Cancer. 13:184–199. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hofstra RM, Landsvater RM, Ceccherini I,
Stulp RP, Stelwagen T, Luo Y, Pasini B, Höppener JW, van Amstel HK,
Romeo G, et al: A mutation in the RET proto-oncogene associated
with multiple endocrine neoplasia type 2B and sporadic medullary
thyroid carcinoma. Nature. 367:375–376. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Russo D, Damante G, Puxeddu E, Durante C
and Filetti S: Epigenetics of thyroid cancer and novel therapeutic
targets. J Mol Endocrinol. 46:R73–R81. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sogawa K, Imataka H, Yamasaki Y, Kusume H,
Abe H and Fujii-Kuriyama Y: cDNA cloning and transcriptional
properties of a novel GC box-binding protein, BTEB2. Nucleic Acids
Res. 21:1527–1532. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dong JT, Boyd JC and Frierson HF Jr: Loss
of heterozygosity at 13q14 and 13q21 in high grade, high stage
prostate cancer. Prostate. 49:166–171. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen C, Brabham WW, Stultz BG, Frierson HF
Jr, Barrett JC, Sawyers CL, Isaacs JT and Dong JT: Defining a
common region of deletion at 13q21 in human cancers. Genes
Chromosomes Cancer. 31:333–344. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kojima S, Kobayashi A, Gotoh O, Ohkuma Y,
Fujii-Kuriyama Y and Sogawa K: Transcriptional activation domain of
human BTEB2, a GC box-binding factor. J Biochem. 121:389–396. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dong JT and Chen C: Essential role of KLF5
transcription factor in cell proliferation and differentiation and
its implications for human diseases. Cell Mol Life Sci.
66:2691–2706. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Du C, Gao Y, Xu S, Jia J, Huang Z, Fan J,
Wang X, He D and Guo P: KLF5 promotes cell migration by
up-regulating FYN in bladder cancer cells. FEBS Lett. 590:408–418.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Maehara O, Sato F, Natsuizaka M, Asano A,
Kubota Y, Itoh J, Tsunematsu S, Terashita K, Tsukuda Y, Nakai M, et
al: A pivotal role of Krüppel-like factor 5 in regulation of cancer
stem-like cells in hepatocellular carcinoma. Cancer Biol Ther.
16:1453–1461. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ci X, Xing C, Zhang B, Zhang Z, Ni JJ,
Zhou W and Dong JT: KLF5 inhibits angiogenesis in PTEN-deficient
prostate cancer by attenuating AKT activation and subsequent HIF1α
accumulation. Mol Cancer. 14:912015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Saiselet M, Floor S, Tarabichi M, Dom G,
Hébrant A, van Staveren WC and Maenhaut C: Thyroid cancer cell
lines: An overview. Front Endocrinol (Lausanne).
3:1332012.PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen HL, Chong IW, Lee YC, Tsai JR, Yuan
SS, Wang HM, Liu WL and Liu PL: Krüppel-like factor 5 mediates
proinflammatory cytokine expression in lipopolysaccharide-induced
acute lung injury through upregulation of nuclear factor-κB
phosphorylation in vitro and in vivo. Mediators Inflamm.
2014:2819842014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jing H and Lee S: NF-κB in cellular
senescence and cancer treatment. Mol Cells. 37:189–195. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yang Y, Goldstein BG, Nakagawa H and Katz
JP: Krüppel-like factor 5 activates MEK/ERK signaling via EGFR in
primary squamous epithelial cells. FASEB J. 21:543–550. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhao D, Zheng HQ, Zhou Z and Chen C: The
Fbw7 tumor suppressor targets KLF5 for ubiquitin-mediated
degradation and suppresses breast cell proliferation. Cancer Res.
70:4728–4738. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Grebe SK and Hay ID: Thyroid cancer nodal
metastases: Biologic significance and therapeutic considerations.
Surg Oncol Clin N Am. 5:43–63. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fu RJ, He W, Wang XB, Li L, Zhao HB, Liu
XY, Pang Z, Chen GQ, Huang L and Zhao KW: DNMT1-maintained
hypermethylation of Krüppel-like factor 5 involves in the
progression of clear cell renal cell carcinoma. Cell Death Dis.
8:e29522017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Oishi Y, Manabe I, Tobe K, Tsushima K,
Shindo T, Fujiu K, Nishimura G, Maemura K, Yamauchi T, Kubota N, et
al: Krüppel-like transcription factor KLF5 is a key regulator of
adipocyte differentiation. Cell Metab. 1:27–39. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jia L, Zhou Z, Liang H, Wu J, Shi P, Li F,
Wang Z, Wang C, Chen W, Zhang H, et al: KLF5 promotes breast cancer
proliferation, migration and invasion in part by upregulating the
transcription of TNFAIP2. Oncogene. 35:2040–2051. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Parisi S, Passaro F, Aloia L, Manabe I,
Nagai R, Pastore L and Russo T: Klf5 is involved in self-renewal of
mouse embryonic stem cells. J Cell Sci. 121:2629–2634. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Qiu W, Xia X, Qiu Z, Guo M and Yang Z:
RasGRP3 controls cell proliferation and migration in papillary
thyroid cancer by regulating the Akt-MDM2 pathway. Gene. 633:35–41.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Guan H, Guo Z, Liang W, Li H, Wei G, Xu L,
Xiao H and Li Y: Trop2 enhances invasion of thyroid cancer by
inducing MMP2 through ERK and JNK pathways. BMC Cancer. 17:4862017.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xiang S, Xiang T, Xiao Q, Li Y, Shao B and
Luo T: Zinc-finger protein 545 is inactivated due to promoter
methylation and functions as a tumor suppressor through the
Wnt/β-catenin, PI3K/AKT and MAPK/ERK signaling pathways in
colorectal cancer. Int J Oncol. 51:801–811. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lv N, Shan Z, Gao Y, Guan H, Fan C, Wang H
and Teng W: Twist1 regulates the epithelial-mesenchymal transition
via the NF-κB pathway in papillary thyroid carcinoma. Endocrine.
51:469–477. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chen C, Sun X, Ran Q, Wilkinson KD, Murphy
TJ, Simons JW and Dong JT: Ubiquitin-proteasome degradation of KLF5
transcription factor in cancer and untransformed epithelial cells.
Oncogene. 24:3319–3327. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhi X, Zhao D, Zhou Z, Liu R and Chen C:
YAP promotes breast cell proliferation and survival partially
through stabilizing the KLF5 transcription factor. Am J Pathol.
180:2452–2461. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liu N, Li H, Li S, Shen M, Xiao N, Chen Y,
Wang Y, Wang W, Wang R, Wang Q, et al: The Fbw7/human CDC4 tumor
suppressor targets proproliferative factor KLF5 for ubiquitination
and degradation through multiple phosphodegron motifs. J Biol Chem.
285:18858–18867. 2010. View Article : Google Scholar : PubMed/NCBI
|