Introduction
Pancreatic cancer (PC) is the fourth leading cause
of cancer-associated mortality worldwide (1), and the majority of patients succumb
within 6 months of the initial diagnosis (2). Poor prognosis is closely associated with
the unsatisfactory results of currently available treatments, which
can be largely ascribed to the unique pancreatic tumor
microenvironment (TME) (3). Various
stromal components act in concert with tumor cells, allowing a
dynamic response to changing environments during tumor development
and following exposure to chemotherapeutic drugs (4). PC cells usually promote a supportive TME
by activating a fibro-inflammatory reaction (3). Conversely, normal host cells can be
educated and activated by PC cells to promote tumor progression
through the secretion of growth factors and chemokines. The
majority of studies have highlighted the involvement of fibroblasts
and inflammatory cells (5–8). However, to the best of our knowledge,
little attention has been paid to the function of mature adipocytes
in the pancreatic TME. Normal adipocytes have primarily been
recognized as an energy reservoir to store surplus fuel, but now,
there is solid evidence to suggest that they are vital endocrine
cells with the ability to produce multiple hormones and chemokines
involved in inflammatory and immune responses (9), indicating that stromal adipocytes may be
excellent candidates to impact pancreatic tumor behavior and the
TME.
Tumor-surrounding host cells can be educated to
undergo morphological, functional and epigenetic changes and form
unique tumor-associated cell types (10,11). Both
in vitro and in vivo studies in breast cancer have
confirmed the identity of distinct cancer-associated adipocytes,
which are characterized by decreased lipid droplets and a
fibroblast-like shape (12–14). In metastatic ovarian cancer, omental
adipocytes at the invasive front can also undergo a lipolytic
process (15). Furthermore, it has
been demonstrated that cancer-associated adipocytes acquire the
ability to produce numerous proinflammatory factors (12,15,16).
However, little is known about the potentially altered phenotypes
of PC-surrounding adipocytes. It is noteworthy that intrapancreatic
fatty infiltration (pancreatic steatosis) has been demonstrated to
be independently associated with the increased risk of PC precursor
lesions and pancreatic carcinoma from clinical investigations
(17,18). In addition, various studies have
confirmed that cancer-associated adipocytes are positively involved
in PC progression (19–22). Therefore, in order to further reveal
the mechanisms of adipocytes in PC progression, research focusing
on pancreatic cancer-associated adipocytes is required. The present
study demonstrated that pancreatic cancer-associated adipocytes
exhibit profound phenotypic alterations in human cancer and in
vitro. The transcriptome profiling of these modified adipocytes
is also outlined, which provides the basic information required for
further studies.
Materials and methods
Pathological examination of adipocytes
in PC specimens
The human tissue samples used in the present study
were obtained from consecutive archival paraffin blocks of patients
who were diagnosed with pancreatic ductal adenocarcinoma and
underwent surgery in Huadong Hospital (Shanghai, China) between
November 2018 and January 2019. The demographic and pathological
features of the patients are listed in Table I. Written informed consent for the use
of tissues in scientific research was obtained from all patients.
The collection of the pathological data from patients with PC was
approved by the Institutional Research Ethics Committee of Huadong
Hospital, Fudan University (approval no. 2018K098).
Immunohistochemical staining of paraffin-embedded tissues with
rabbit anti-FABP4 polyclonal antibody (pAb; cat. no. 12802-1-AP;
ProteinTech Group, Inc.) was performed in order to observe the
characteristics of the adipocytes. For each slide, representative
images of adipocytes surrounding the normal tissue and adipocytes
in the vicinity of the PC cells were obtained. Adipocyte cell sizes
were assessed using ImageJ software (National Institutes of Health)
(23).
 | Table I.The demographic and pathological
features of the patients. |
Table I.
The demographic and pathological
features of the patients.
| No. | Sex | Age | Location | Tumor
sizea | Tumor
differentiation | TNM
stageb |
|---|
| 1 | Male | 48 | Head | 4.6 cm | Moderate | IIB |
| 2 | Male | 66 | Head | 6.8 cm | Poor | III |
| 3 | Female | 76 | Head | 4.9 cm | Poor | III |
| 4 | Male | 67 | Head | 4.1 cm | Well | IIA |
| 5 | Male | 62 | Tail | 4.3 cm | Moderate | IIB |
| 6 | Female | 61 | Body | 5.7 cm | Poor | IIA |
| 7 | Male | 69 | Tail | 3.3 cm | Moderate | IB |
| 8 | Female | 79 | Head | 2.4 cm | Well | IB |
| 9 | Female | 64 | Head | 3.6 cm | Poor | IIA |
| 10 | Female | 66 | Body | 5.1 cm | Poor | IIB |
Cell culture and coculture
The human PC cell lines Panc-1 and Mia PaCa2 were
purchased from the Cell Bank of the Type Culture Collection of
Chinese Academy of Sciences and cultured according to standard
protocols. In brief, Panc-1 cells were maintained in Dulbecco's
modified Eagle's medium (DMEM; Gibco; Thermo Fisher Scientific,
Inc.), containing 10% fetal bovine serum (FBS) and 1%
penicillin/streptomycin. Mia PaCa2 cells were cultured in DMEM,
supplemented with 10% FBS and 2.5% horse serum. Murine 3T3-L1
preadipocytes were purchased from the American Type Culture
Collection and cultured in DMEM supplemented with 10% newborn calf
serum. All cells were cultured at 37°C (5% CO2) in a
humidified incubator. Two days after reaching 100% confluence
(designated as Day 0), the 3T3-L1 preadipocytes were then induced
to differentiate in DMEM containing 10% FBS, 0.5 mM
3-isobutyl-1-methylxanthine (M), 1 µM dexamethasone (D) and 1 µg/ml
insulin (I). After 48 h (Day 2), these cells were sustained in DMEM
supplemented with 10% FBS and 1 µg/ml insulin for another 2 days.
Next, 3T3-L1 mature adipocytes were cultured in DMEM supplemented
with 10% FBS. After induction (at Day 8), mature adipocytes were
cocultured with PC cells using a Transwell indirect coculture
system (0.4 µm pore size; Corning), and mature adipocytes cultured
alone at the same time-points were used as controls.
RNA sequencing (RNA-seq) of 3T3-L1
cells
Total RNA was extracted from 3T3-L1 mature
adipocytes using TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocol.
RNA-seq libraries were prepared according to the TruSeq™ RNA Sample
Preparation kit from Illumina. Briefly, 5 µg total RNA was isolated
according to the polyA mRNA selection method by oligo(dT) beads and
then fragmented. Double-stranded cDNA was synthesized using a
SuperScript Double-stranded cDNA Synthesis kit (Invitrogen; Thermo
Fisher Scientific, Inc.) with random primers (Illumina). Libraries
were size selected for cDNA target fragments of 200–300 bp on 2%
Low Range Ultra Agarose, and the cDNA library was amplified using
Phusion DNA polymerase (NEB) for 15 PCR cycles. Quantified by
TBS380, amplified cDNA fragments were sequenced with the Illumina
HiSeq X10 platform (2×150 bp read length). The expression level of
each gene was calculated using the fragments per kilobase of
transcript per million (FPKM) base pairs sequenced method (24). In order to identify the differentially
expressed genes (DEGs) between 2 groups (3 biological replicates
per group), Cuffdiff was used (25).
Genes with a fold change ≥2 and a false discovery rate (FDR)
<0.05 were identified as significant DEGs. In order to
comprehend the functions of these DEGs, Gene Ontology (GO)
functional enrichment and the Kyoto Encyclopedia of Genes and
Genomes (KEGG) pathway analysis were performed using Goatools and
KOBAS, respectively (26). Heat-map
representations of the expression of selected genes expression were
generated using the ‘pheatmap’ R package.
Oil Red O staining
Following three washes with PBS, 3T3-L1 adipocytes
were fixed with 3.7% formaldehyde for 20 min. Stock Oil Red O
solution (0.5% in isopropanol) was diluted with distilled water
(3:2). Fixed cells were subsequently stained with Oil Red O
solution at room temperature for 1 h (27). The cells were then washed with water
and visualized by light microscopy.
Reverse transcriptase-quantitative PCR
(RT-qPCR)
Following total RNA extraction, PrimeScript RT
Master Mix (Takara Bio) was used for first-strand cDNA synthesis
with random primers. The expression of the considered genes was
determined via RT-qPCR, which was carried out with SYBR-Green PCR
Master Mixture reagent (Applied Biosystems; Thermo Fisher
Scientific, Inc.) using an ABI 7500 Real-Time PCR system (Applied
Biosystems; Thermo Fisher Scientific, Inc.). Thermocycling
conditions were as follows: 10 min denaturation at 95°C, followed
by 40 cycles of 95°C for 15 sec and 60°C for 1 min. Relative RNA
quantity was calculated using the 2−ΔΔCq method with 18S
rRNA as an endogenous control (28).
All reactions were performed in triplicate. The primer sequences
used in the present study are listed in Table II.
| Table II.Primers used in the present
study. |
Table II.
Primers used in the present
study.
| Gene name | Forward primer
sequence (5′ to 3′) | Reverse primer
sequence (5′ to 3′) |
|---|
| 18S |
CGCCGCTAGAGGTGAAATTCT |
CATTCTTGGCAAATGCTTTCG |
| C/EBPα |
CAAGAACAGCAACGAGTACCG |
GTCACTGGTCAACTCCAGCAC |
| PPAR-γ |
CCGAAGAACCATCCGATTGA |
TTTGTGGATCCGGCAGTTAAG |
| FABP4 |
TTCGATGAAATCACCGCAGA |
GGTCGACTTTCCATCCCACTT |
| HSL |
GGCTTACTGGGCACAGATACCT |
CTGAAGGCTCTGAGTTGCTCAA |
| ATGL |
CTGAGAATCACCATTCCCACATC |
CACAGCATGTAAGGGGGAGA |
| FASN |
AGAGATCCCGAGACGCTTCT |
GCTTGGTCCTTTGAAGTCGAAGA |
| CIDEA |
GCCTGCAGGAACTTATCAGC |
AGAACTCCTCTGTGTCCACCA |
| Glut4 |
ACACTGGTCCTAGCTGTATTC |
CCAGCCACGTTGCATTGTA |
| IRS1 |
CCAGCCTGGCTATTTAGCTG |
CCCAACTCAACTCCACCACT |
| APN |
TGGAATGACAGGAGCTGAAGG |
TATAAGCGGCTTCTCCAGGCT |
| RESISTIN |
TCGTGGGACATTCGTGAAGA |
GGGCTGCTGTCCAGTCTATCC |
| LEPTIN |
GGGCTTCACCCCATTCTGA |
TGGCTATCTGCAGCACATTTTG |
| α-SMA |
GGAGAAGCCCAGCCAGTCGC |
AGCCGGCCTTACAGAGCCCA |
| FSP-1 |
GCTGCCCAGATAAGGAACCC |
TGCGAAGAAGCCAGAGTAAGG |
| MMP-9 |
TGAGTCCGGCAGACAATCCT |
CGCCCTGGATCTCAGCAATA |
| PAI-1 |
TCTCCAATTACTGGGTGAGTCAGA |
GCAGCCGGAAATGACACAT |
| MMP11 |
GCCCTCATGTCCCCTTTCTAC |
CCTTCGGTCATCTGGGCTAA |
Western blot analysis
Total lysates were extracted using lysis buffer
containing 50 mM Tris-HCl (pH 6.8), 2% SDS, protease inhibitor
mixture (all from Roche Diagnostics), 100 mmol/l NaF, and 1 mmol/l
PMSF. The protein concentration was determined by the bicinchoninic
acid assay. Equal amounts (20 µg) of protein were separated via 12%
SDS-PAGE, transferred to polyvinylidene fluoride membranes (EMD
Millipore), blocked with 5% skim milk at room temperature for 1 h
and immunoblotted with the indicated primary antibodies (1:1,000)
at 4°C overnight. Rabbit anti-PPAR-γ pAb (cat. no. 16643-1-AP),
rabbit anti-FABP4 pAb (cat. no. 12802-1-AP), rabbit anti-HSL pAb
(LIPE; cat. no. 17333-1-AP), rabbit anti-FASN pAb (cat. no.
10624-2-AP) and rabbit anti-α-tubulin pAb (cat. no. 11224-1-AP)
were purchased from ProteinTech Group, Inc. Rabbit anti-phospho-Akt
monoclonal antibody (mAb; cat. no. 4060) and rabbit anti-Akt mAb
(cat. no. 4691) were purchased from Cell Signaling Technology, Inc.
The following day, membranes were incubated with horseradish
peroxidase-conjugated anti-rabbit IgG antibodies (1:5,000; cat. no.
SA00001-2; ProteinTech Group, Inc.) at 37°C for 1 h. Blots were
then visualized with enhanced chemiluminescence detection kit
(Thermo Fisher Scientific, Inc.) using the ImageQuant LAS 4000
System (GE Healthcare). Semi-quantification analysis was conducted
by Quantity One software, version 4.6 (Bio-Rad Laboratories).
Transwell cell migration assay
PC cells (Panc-1 and Mia PaCa2) alone and cocultured
with mature adipocytes were assessed for their ability to migrate
through an 8 µm pore membrane (24-well insert; Corning). After 5
days of coculture with adipocytes, tumor cells were trypsinized and
seeded (5×104) into the upper chamber in serum-free
DMEM. The lower chamber was filled with medium containing 10% FBS.
After incubation for 36 h, cells on the bottom of the insert were
fixed with 100% methanol and stained in 0.1% crystal violet at room
temperature for 30 min. The number of migrating cells was counted
separately in three randomly selected fields using a light
microscope (IX71; Olympus).
Statistical analysis
Statistical analysis of all data was completed using
GraphPad Prism software (version 5.0; GraphPad Software, Inc.). All
experiments were performed in triplicate. The results are expressed
as the mean ± standard deviation. Unpaired two-tailed Student's
t-test was performed to determine significance of differences
between two groups. P<0.05 was considered to indicate a
statistically significant difference. P-values are presented in the
figures (*P<0.05; **P<0.01; ***P<0.001).
Results
Pancreatic cancer cells stimulate
delipidation in mature adipocytes
A series of studies have demonstrated crosstalk
between cancer cells and adipocytes, but these have largely focused
on the postive effect of adiopocytes on tumor progression (29). In order to determine the direct impact
of PC cells on mature adipocytes, the presence of adipocytes in
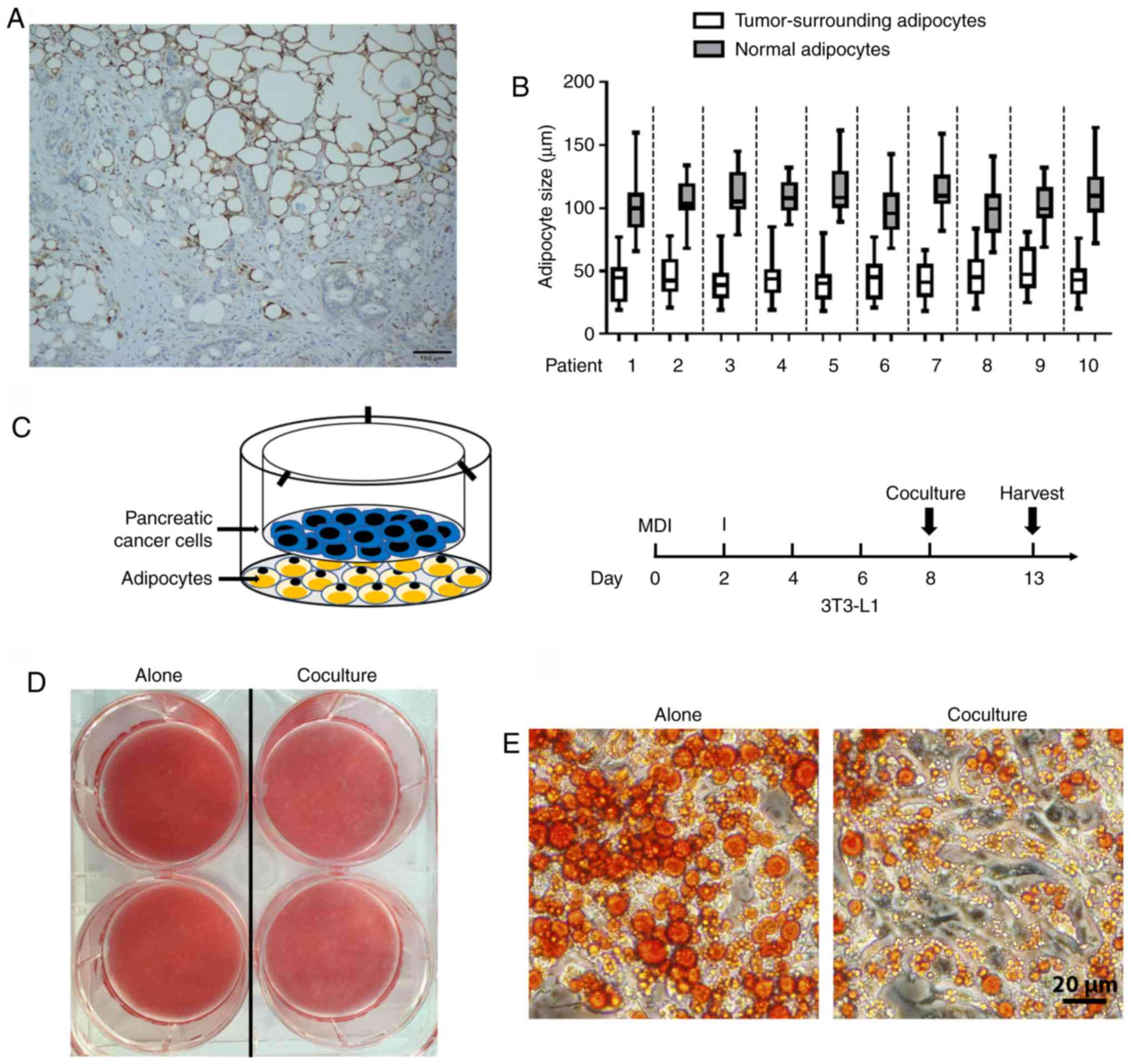
human pancreatic tumors was first investigated. To address this
issue, immunohistochemical staining of FABP4 (highly expressed in
mature adipocytes) was performed in tumor samples. The presence of
FABP4-stained adipocytes at the invasive front of pancreatic tumors
was observed (Fig. 1A). Furthermore,
tumor-neighboring adipocytes were smaller in size compared with
those normal adipocytes further away (Fig. 1B). In order to assess the
morphological changes of PC-related adipocytes, an adipocyte-PC
cell indirect coculture system was established (Fig. 1C, left panel). First, 3T3-L1 cells
were induced to differentiate into mature adipocytes, and human PC
cells were then cultured on top (Fig.
1C, right panel). As indicated by Oil Red O staining, following
coculture with Panc-1 for 5 days, mature adipocytes exhibited a
decrease in lipid content (Fig. 1D)
with a marked decrease in the size of lipid droplets (Fig. 1E) compared with those cultured alone,
which is consistent with the clinical features of small adipocytes
adjacent to the tumor tissues.
Altered gene expression profiling is
identified in pancreatic cancer-associated adipocytes
Given the great phenotypic changes of
cancer-associated adipocytes in other types of tumor, such as
breast and prostate cancer (13,16), we
reasoned that PC-associated adipocytes may change their gene
expression patterns. RNA-sequencing was performed on 3T3-L1 cells
cultured with or without PC cells, and identified 1,081 genes that
were differentially expressed in cancer-associated adipocytes,
including 541 upregulated and 540 downregulated genes (Fig. 2A). Next, all DEGs were characterized
through GO term mapping according to their major GO categories
[i.e., the GO terms ‘biological processes’ (BP), ‘cellular
components’ (CC), and ‘molecular functions’ (MF)] (Fig. 2B). The top BP GO annotation was
‘positive regulation of cell migration/motility/cellular component
movement’. Within the CC of ontology, the most abundant term was
‘proteinaceous extracellular matrix’. Furthermore, ‘receptor
regulator activity’ was primarily enriched in the MF group. DEGs
were also mapped to the KEGG pathway database (Fig. 2C). Numerous metabolic pathways, such
as ‘insulin resistance’, ‘glucagon signaling pathway’ and
‘cholesterol metabolism’, were enriched, indicating that diverse
metabolic changes were occurring in the cancer-associated
adipocytes. It was also revealed that ‘cytokine-cytokine receptor
interaction’ was the most significantly enriched KEGG pathway.
 | Figure 2.Altered gene expression profiling is
identified in mature adipocytes following coculture with Panc-1
cancer cells. (A) Left, volcano plot of DEGs. Upregulated genes in
coculture adipocytes are colored in red, and downregulated genes in
blue. Right, schematic representation of the DEGs. Of 1,081 DEGs,
541 were upregulated, while 540 were downregulated in the coculture
group compared with the control group. (B) Histogram of GO
annotations. The three major categories, BP, CC and MF, were
analyzed. (C) A scatter diagram of enriched KEGG pathways. The
level of KEGG enrichment was measured using GeneRatio. The color
and size of the dots represent the range of P-adjusted values and
gene counts, respectively. DEGs, differentially expressed genes;
GO, Gene Ontology; BP, biological process; CC, cellular component;
MF, molecular function; KEGG, Kyoto Encyclopedia of Genes and
Genomes. |
Pancreatic cancer induces dysregulated
metabolism in adipocytes
The aforementioned data revealed that upon coculture
with PC cells, a marked decrease in lipid accumulation and a
profound impact on gene expression networks associated with
metabolism were exhibited in adipocytes. Furthermore, a broad
spectrum of genes involved in fatty acid and triglyceride synthesis
for lipogenesis, and their upstream regulators, as well as
triglyceride storage and degradation, were significantly decreased
(Fig. 3A), revealing the dysregulated
lipid homeostasis in cocultured adipocytes. RT-qPCR analysis of
adipocytes cocultured with Panc-1 (Fig.
3B) or with another PC cell line Mia PaCa2 (Fig. 3C) was performed, which confirmed the
substantial inhibition of lipid metabolism-associated genes,
including FABP4, HSL, ATGL, CIDEA and FASN, and of two key
adipogenic upstream factors, PPAR-γ and C/EBPα. In line with the
RT-qPCR data, immunoblotting analysis also confirmed decreased
levels of FABP4, HSL, FASN and PPAR-γ in cocultured adipocytes
(Fig. 4A and B). The present study
also investigated the effect of coculture on adipocyte glucose
metabolism and revealed that there was a marked decrease in the
insulin-induced phosphorylation of Akt (Fig. 4C), as well as the transcriptional
expression of Glut4 and IRS1 (Fig.
4D).
Pancreatic cancer-associated
adipocytes undergo dedifferentiation
Adipocytes are commonly regarded as terminally
differentiated cells, but increasing evidence suggests they may
have greater plasticity under certain circumstances (12,30). In
the present study, the ‘fat cell differentiation’ pathway was
highly enriched in cocultured adipocytes. A total of 25 DEGs were
involved in this process (Table
III) among which several mature adipocyte markers, such as
leptin and resistin, were significantly downregulated. It was also
observed that, following coculture with pancreatic cancer cells for
5 days, numerous adipocytes exhibited a stretched, fibroblastic
shape (Fig. 5A). Further analysis
using the GO BP demonstrated that the mesenchymal cell
differentiation process was activated in cocultured adipocytes
(Fig. 5B). There was also a marked
increase in the expression of mesenchymal state-associated genes
(31,32), indicating a shift towards more
mesenchymal phenotypes in these cocultured adipocytes (Fig. 5C). The RT-qPCR results demonstrated
that adipocytes cocultured with Panc-1 exhibited a loss in
expression of mature adipocyte markers, including adiponectin
(APN), leptin and resistin, and a gain in expression of fibroblast
markers, including α-SMA, FSP-1, MMP-9, PAI-1 and MMP-11 (Fig. 5D). Similar results were obtained by
coculturing adipocytes with Mia PaCa2 (Fig. 5E).
| Table III.DEGs involved in the fat
differentiation pathway in coculture adipocytes. |
Table III.
DEGs involved in the fat
differentiation pathway in coculture adipocytes.
| Gene ID | Description | log2
FC | Regulation |
|---|
| RNASEL | Ribonuclease L | 1.034174 | Up |
| ENPP1 | Ectonucleotide
pyrophosphatase/phosphodiesterase 1 | 1.108244 | Up |
| MMP11 | Matrix
metallopeptidase 11 | 2.89554 | Up |
| VSTM2A | V-set and
transmembrane domain containing 2A | 1.242257 | Up |
| ARL4A | ADP-ribosylation
factor-like 4A | −1.43843 | Down |
| ERO1L | ERO1-like | 1.709092 | Up |
| KLF5 | Kruppel-like factor
5 | 1.345372 | Up |
| CEBPD | CCAAT/enhancer
binding protein (C/EBP), delta | 1.382325 | Up |
| ADGRF5 | Adhesion G
protein-coupled receptor F5 | 1.84409 | Up |
| ZBTB7C | Zinc finger and BTB
domain containing 7C | 1.041932 | Up |
| CCDC85B | Coiled-coil domain
containing 85B | 1.669037 | Up |
| SCD1 | Stearoyl-Coenzyme A
desaturase-1 | −1.01976 | Down |
| ADIG | Adipogenin | −1.21847 | Down |
| NOCT | Nocturnin | −1.14398 | Down |
| KLF4 | Kruppel-like factor
4 | 1.189328 | Up |
| TRPV4 | Transient receptor
potential cation channel subfamily V, member 4 | 1.235702 | Up |
| TMEM120A | Transmembrane
protein 120A | −1.1772 | Down |
| MEDAG | Mesenteric estrogen
dependent adipogenesis | 1.111338 | Up |
| LEP | Leptin | −1.84249 | Down |
| CREB5 | cAMP responsive
element binding protein 5 | 1.663459 | Up |
| CLIP3 | CAP-GLY domain
containing linker protein 3 | 1.930421 | Up |
| AAMDC | Adipogenesis
associated Mth938 domain containing | −1.15184 | Down |
| RETN | Resistin | −1.56133 | Down |
| ADRB3 | Adrenergic
receptor, β3 | −1.69788 | Down |
| ANGPTL8 | Angiopoietin-like
8 | −1.03671 | Down |
Cancer-associated adipocytes increase
PC cell aggressiveness
Previous research has demonstrated that adipocytes
activated by tumor cells exerted an active role in tumor
progression and TME remodeling (33).
In the present study, bioinformatics analysis of the whole
transcriptome of adipocytes revealed several enriched pathways
associated with the remodeling of the pancreatic cancer stroma
(Table IV), including the
cytokine-mediated signaling pathway, angiogenesis and extracellular
matrix organization. In order to directly assess the role of
cancer-associated adipocytes in PC progression, the present study
performed Transwell assays to assess the ability of the PC cells to
traverse the membrane. PC cells were cocultured with or without
adipocytes for 5 days, and it was revealed that both
adipocyte-exposed Panc-1 and Mia PaCa2 tumor cells had a
significantly increased ability to fully migrate through a
Transwell membrane (Fig. 6). Overall,
these results indicated that adipocytes can increase PC cell
aggressiveness.
| Table IV.Summary of enriched pathways
potentially related to the remodeling of the pancreatic tumor
microenvironment in cocultured adipocytes. |
Table IV.
Summary of enriched pathways
potentially related to the remodeling of the pancreatic tumor
microenvironment in cocultured adipocytes.
| Category | Gene count | Genes | P-value |
|---|
| Cytokine-mediated
signaling pathway | 28 |
ACKR3/GPR35/GREM2/IRAK3/CCL11/CCL8/CCL9/CCL6/SPHK1SOCS3/LRTM1/OSMR/PRLR/IL2RB/PDGFB/LRRC15/TREM2/RTN4RL2/ECM1/CXCL9/CXCL10/CXCL13/EPO/LEP/IRF7/ACSL1/CX3CL1/CASP4 | 2.65 e-6 |
| Angiogenesis | 37 |
NRP2/FN1/IHH/ACKR3/CHIL1/ITGB2/ADORA2B/CCL11/SPHK1/AHR/PRL2C2/PLAU/KLF5/RUNX1/DLL1/FGF1/ENG/GREM1/ANGPT4/ECM1/CYR61/KLF4/NOS3/CXCL10/ANXA3/HSPB1/SERPINE1/EPO/LEP/AQP1/ADM/HMOX1/CX3CL1/FOXC2/THY1/CAMP/TGFBR2 | 7.28 e-8 |
| Extracellular
matrix organization | 27 |
FN1/IHH/LAMC2/MMP11/ECM2/ERO1L/EGFLAM/FBLN1/MYH11
PHLDB2/HAS1/LOX/ADAMTSL2/ENG/OLFML2A/WT1/GREM1/FERMT1/NPNT/CYR61/EMILIN1/ELN/FSCN1/BCL3/APLP1/FOXC2/MMP13 | 4.73 e-10 |
Discussion
Research focusing on the occult PC stroma has
resulted in an increased level of knowledge and understanding;
however, the current understanding of cancer-associated adipocytes
remains unclear compared with that of other cell types, forming the
impetus for the research presented. It was consistently observed
that adipocytes adjacent to human pancreatic tumors had smaller
lipid droplet sizes. In the in vitro coculture system of the
present study, adipocytes demonstrated a status of delipidation and
impaired lipid homeostasis, indicating that PC cells are able to
blunt energy-consuming activities, including adipogenesis in
cocultured adipocytes. The impact of the tumor shutting down
adipocyte metabolism should serve to save nutrients (such as
glucose) that would be accessible for the tumor. Regarding the
mechanisms whereby PC cells decreased energy utilization in
adipocytes, it was observed that dysregulated lipid metabolism was
accompanied by insulin resistance, demonstrated by a decrease in
insulin-stimulated phosphorylation of Akt, as well as decreased
Glut4 and IRS1 mRNA expression in cocultured adipocytes. Previous
studies have revealed that Glut4-mediated adipose tissue glucose
influx could activate de novo lipogenesis, and the latter
favored enhanced systemic insulin sensitivity (34,35). This
is of clinical relevance since it has been recognized that PC is a
powerful diabetogenic state and new-onset diabetes in patients with
PC is likely to be induced by the secreted products of cancer cells
(36). The metabolic reprogramming of
adipocytes orchestrated by cancer cells would help PC deal with the
high energy demand during tumor progression through the control of
non-cancer cell insulin sensitivity. Furthermore, it is well
recognized that adipocyte-derived lipids are potent energy sources
that fulfill the energetic needs of tumor cells (15,37,38). In
this regard, the phenomenon of delipidation could also result from
the abundant release of free fatty acids in cocultured adipocytes.
In fact, the increased accumulation of lipid in PC cells during
coculturing with adipocytes was observed in the present study (data
not shown). Collectively, these results indicated that multiple
modes of metabolic crosstalk occur between stromal adipocytes and
PC cells.
Another key concern arising from the present study
is the process of dedifferentiation in adipocytes induced by PC
cells. A number of studies have suggested that adipocytes retain
their capacity to return to a precursor state. Cell lineage tracing
studies in mouse models have revealed that adipocytes could undergo
reprogramming into fibroblasts within fibrotic dermis (39), or into preadipocytes during lactation
(40). In agreement with previously
reported results demonstrating such adipocyte plasticity during
adipocyte-cancer crosstalk (13,41–43), the
present study revealed that cocultured adipocytes underwent
dedifferentiation, forming the mesenchymal cell type and giving
rise to fibroblast-like cells. It will be of great interest to
determine the underlying mechanisms of this process. In this
regard, the factors secreted by tumor cells are likely to be the
key regulators. Both Wnt3a released by breast cancer cells
(13) and Wnt5a secreted from
melanoma cells (42) have been
reported as triggering factors of adipocyte dedifferentiation in
the tumor microenvironment. In addition, the aforementioned role of
adipocyte metabolic alteration on the dedifferentiation process
cannot be excluded. It is not currently known whether metabolic
changes have a critical impact on the adipocyte differentiation and
vice versa, therefore further research is required.
Notably, for the adipocytes exposed to PC cells, in
addition to the impairment of metabolism and concomitant
dedifferentiation process, it is of great interest to determine
whether they have acquired pluripotency, and can reprogram their
genomes to form new subsets. The data from the present study
demonstrated that adipocytes cocultured with cancer cells maintain
a mesenchymal state, which may facilitate the entrance into a
stem-cell state (44). In addition,
bioinformatics analysis of the transcriptome of adipocytes
revealed, following coculture, a marked change in gene expression
networks associated with the modulation of matrix remodeling. These
adipocytes could obtain new competencies that contribute to the
remodeling of the tumor stroma. In breast cancer, cancer-associated
adipocytes revert to a fibroblastic cell population and take part
in the desmoplastic reaction (13).
PC is a highly desmoplastic malignancy characterized by excessive
deposition of the extracellular matrix (ECM) (45). Obesity is positively correlated with
increased ECM deposition in pancreatic TME, and adipocytes in the
fatty PC stroma can activate pancreatic stellate cells, indirectly
leading to excessive desmoplasia (19). Collectively, these results highlight
the role of cancer-associated adipocytes in the PC desmoplastic
stroma. The present study also characterized the cytokine-mediated
signaling and the positive regulation of angiogenesis in PC-induced
adipocytes. The inappropriate release of proinflammatory factors by
adipose tissue has been extensively recognized as the primary cause
of obesity-associated disorders, including cancer (46,47). In
adipose tissue, the presence of pancreatic tumor cells may result
in a marked increased inflammatory response. In addition, adipose
tissue is one of the most vascularized tissues in the body with the
highest capacity for angiogenesis (48), and highly vascularized adipose tissue
may provide a predetermined environment to foster tumors. Thus, it
is reasonable to speculate that upon interplay with adipose tissue,
tumors may mobilize the angiogenetic process of adipocytes for
their own sake.
In summary, the present study revealed the distinct
phenotypes of pancreatic cancer-associated adipocytes. The model
utilized, as presented in Fig. 7,
proposes systemic crosstalk between PC and adipocytes, providing
critical insights towards the understanding of the contribution of
adipocytes to the formation of a unique pancreatic tumor stroma,
which ultimately leads to increased tumor malignancy.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The sequencing data have been deposited in the Gene
Expression Omnibus with the assigned accession number, GSE123939.
The datasets used and/or analyzed during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
ZC, YL, WW and CJ participated in the conception and
design of the present study. ZC, YL, CX performed the statistical
analysis and were involved in the preparation of the figures. HW,
JL and PH participated in the analysis of the figures and data. ZC,
YL, WW and CJ prepared and revised the manuscript. HH reviewed the
results and revised the manuscript. All authors have read and
approved the final manuscript and agree to be accountable for all
aspects of the research in ensuring that the accuracy or integrity
of any part of the work are appropriately investigated and
resolved.
Ethics approval and consent to
participate
The present study was approved by the Institutional
Research Ethics Committee of Huadong Hospital, Fudan University
(approval no. 2018K098). Written informed consent for the use of
tissues in scientific research was obtained from all patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2018. CA Cancer J Clin. 68:7–30. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Michl P and Gress TM: Current concepts and
novel targets in advanced pancreatic cancer. Gut. 62:317–326. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Feig C, Gopinathan A, Neesse A, Chan DS,
Cook N and Tuveson DA: The pancreas cancer microenvironment. Clin
Cancer Res. 18:4266–4276. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Quail DF, Bowman RL, Akkari L, Quick ML,
Schuhmacher AJ, Huse JT, Holland EC, Sutton JC and Joyce JA: The
tumor microenvironment underlies acquired resistance to CSF-1R
inhibition in gliomas. Science. 352:62882016. View Article : Google Scholar
|
|
5
|
von Ahrens D, Bhagat TD, Nagrath D, Maitra
A and Verma A: The role of stromal cancer-associated fibroblasts in
pancreatic cancer. J Hematol Oncol. 10:762017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Helm O, Held-Feindt J, Grage-Griebenow E,
Reiling N, Ungefroren H, Vogel I, Kruger U, Becker T, Ebsen M,
Rocken C, et al: Tumor-associated macrophages exhibit pro- and
anti-inflammatory properties by which they impact on pancreatic
tumorigenesis. Int J Cancer. 135:843–861. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hwang RF, Moore T, Arumugam T,
Ramachandran V, Amos KD, Rivera A, Ji B, Evans DB and Logsdon CD:
Cancer-associated stromal fibroblasts promote pancreatic tumor
progression. Cancer Re. 68:918–926. 2008. View Article : Google Scholar
|
|
8
|
Bailey JM, Swanson BJ, Hamada T, Eggers
JP, Singh PK, Caffery T, Ouellette MM and Hollingsworth MA: Sonic
hedgehog promotes desmoplasia in pancreatic cancer. Clin Cancer
Res. 14:5995–6004. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Deng Y and Scherer PE: Adipokines as novel
biomarkers and regulators of the metabolic syndrome. Ann N Y Acad
Sci. 1212:E1–E19. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xiao Q, Zhou D, Rucki AA, Williams J, Zhou
J, Mo G, Murphy A, Fujiwara K, Kleponis J, Salman B, et al:
Cancer-associated fibroblasts in pancreatic cancer are reprogrammed
by tumor-induced alterations in genomic DNA methylation. Cancer
Res. 76:5395–5404. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Apte MV, Wilson JS, Lugea A and Pandol SJ:
A starring role for stellate cells in the pancreatic cancer
microenvironment. Gastroenterology. 144:1210–1219. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dirat B, Bochet L, Dabek M, Daviaud D,
Dauvillier S, Majed B, Wang YY, Meulle A, Salles B, Le Gonidec S,
et al: Cancer-associated adipocytes exhibit an activated phenotype
and contribute to breast cancer invasion. Cancer Res. 71:2455–2465.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bochet L, Lehuede C, Dauvillier S, Wang
YY, Dirat B, Laurent V, Dray C, Guiet R, Maridonneau-Parini I, Le
Gonidec S, et al: Adipocyte-derived fibroblasts promote tumor
progression and contribute to the desmoplastic reaction in breast
cancer. Cancer Res. 73:5657–5668. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang YY, Attane C, Milhas D, Dirat B,
Dauvillier S, Guerard A, Gilhodes J, Lazar I, Alet N, Laurent V, et
al: Mammary adipocytes stimulate breast cancer invasion through
metabolic remodeling of tumor cells. JCI Insight. 2:e874892017.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nieman KM, Kenny HA, Penicka CV, Ladanyi
A, Buell-Gutbrod R, Zillhardt MR, Romero IL, Carey MS, Mills GB,
Hotamisligil GS, et al: Adipocytes promote ovarian cancer
metastasis and provide energy for rapid tumor growth. Nat Med.
17:1498–1503. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Laurent V, Guerard A, Mazerolles C, Le
Gonidec S, Toulet A, Nieto L, Zaidi F, Majed B, Garandeau D,
Socrier Y, et al: Periprostatic adipocytes act as a driving force
for prostate cancer progression in obesity. Nat Commun.
7:102302016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hori M, Takahashi M, Hiraoka N, Yamaji T,
Mutoh M, Ishigamori R, Furuta K, Okusaka T, Shimada K, Kosuge T, et
al: Association of pancreatic fatty infiltration with pancreatic
ductal adenocarcinoma. Clin Transl Gastroenterol. 5:e532014.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rebours V, Gaujoux S, d'Assignies G,
Sauvanet A, Ruszniewski P, Levy P, Paradis V, Bedossa P and
Couvelard A: Obesity and fatty pancreatic infiltration are risk
factors for pancreatic precancerous lesions (PanIN). Clin Cancer
Res. 21:3522–3528. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Incio J, Liu H, Suboj P, Chin SM, Chen IX,
Pinter M, Ng MR, Nia HT, Grahovac J, Kao S, et al: Obesity-Induced
inflammation and desmoplasia promote pancreatic cancer progression
and resistance to chemotherapy. Cancer Discov. 6:852–869. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Okumura T, Ohuchida K, Sada M, Abe T, Endo
S, Koikawa K, Iwamoto C, Miura D, Mizuuchi Y, Moriyama T, et al:
Extra-pancreatic invasion induces lipolytic and fibrotic changes in
the adipose microenvironment, with released fatty acids enhancing
the invasiveness of pancreatic cancer cells. Oncotarget.
8:18280–18295. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ziegler KM, Considine RV, True E,
Swartz-Basile DA, Pitt HA and Zyromski NJ: Adipocytes enhance
murine pancreatic cancer growth via a hepatocyte growth factor
(HGF)-mediated mechanism. Int J Surg. 28:179–184. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Carbone C, Piro G, Gaianigo N, Ligorio F,
Santoro R, Merz V, Simionato F, Zecchetto C, Falco G, Conti G, et
al: Adipocytes sustain pancreatic cancer progression through a
non-canonical WNT paracrine network inducing ROR2 nuclear
shuttling. Int J Obes (Lond). 42:334–343. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Schneider CA, Rasband WS and Eliceiri KW:
NIH image to imageJ: 25 years of image analysis. Nat Methods.
9:671–675. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Trapnell C, Williams BA, Pertea G,
Mortazavi A, Kwan G, van Baren MJ, Salzberg SL, Wold BJ and Pachter
L: Transcript assembly and quantification by RNA-Seq reveals
unannotated transcripts and isoform switching during cell
differentiation. Nat Biotechnol. 28:511–515. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Trapnell C, Hendrickson DG, Sauvageau M,
Goff L, Rinn JL and Pachter L: Differential analysis of gene
regulation at transcript resolution with RNA-seq. Nat Biotechnol.
31:46–53. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mao X, Cai T, Olyarchuk JG and Wei L:
Automated genome annotation and pathway identification using the
KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics.
21:3787–3793. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Huang H, Song TJ, Li X, Hu L, He Q, Liu M,
Lane MD and Tang QQ: BMP signaling pathway is required for
commitment of C3H10T1/2 pluripotent stem cells to the adipocyte
lineage. Proce Natl Acad Sci USA. 106:12670–12675. 2009. View Article : Google Scholar
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Park J, Morley TS, Kim M, Clegg DJ and
Scherer PE: Obesity and cancer-mechanisms underlying tumour
progression and recurrence. Nat Rev Endocrinol. 10:455–465. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Corsa CAS and MacDougald OA: Cyclical
dedifferentiation and redifferentiation of mammary adipocytes. Cell
Metab. 28:187–189. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kalluri R and Weinberg RA: The basics of
epithelial-mesenchymal transition. J Clin Invest. 119:1420–1428.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Nieto MA, Huang RY, Jackson RA and Thiery
JP: EMT: 2016. Cell. 166:21–45. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lengyel E, Makowski L, DiGiovanni J and
Kolonin MG: Cancer as a matter of fat: The crosstalk between
adipose tissue and tumors. Trends Cancer. 4:374–384. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Herman MA, Peroni OD, Villoria J, Schon
MR, Abumrad NA, Bluher M, Klein S and Kahn BB: A novel ChREBP
isoform in adipose tissue regulates systemic glucose metabolism.
Nature. 484:333–338. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Iizuka K, Bruick RK, Liang G, Horton JD
and Uyeda K: Deficiency of carbohydrate response element-binding
protein (ChREBP) reduces lipogenesis as well as glycolysis. Proc
Natl Acad Sci USA. 101:7281–7286. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sah RP, Nagpal SJ, Mukhopadhyay D and
Chari ST: New insights into pancreatic cancer-induced
paraneoplastic diabetes. Nat Rev Gastroenterol Hepatol. 10:423–433.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hoy AJ, Balaban S and Saunders DN:
Adipocyte-tumor cell metabolic crosstalk in breast cancer. Trends
Mol Med. 23:381–392. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Miranda F, Mannion D, Liu S, Zheng Y,
Mangala LS, Redondo C, Herrero-Gonzalez S, Xu R, Taylor C, Chedom
DF, et al: Salt-inducible kinase 2 couples ovarian cancer cell
metabolism with survival at the adipocyte-rich metastatic niche.
Cancer Cell. 30:273–289. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Marangoni RG, Korman BD, Wei J, Wood TA,
Graham LV, Whitfield ML, Scherer PE, Tourtellotte WG and Varga J:
Myofibroblasts in murine cutaneous fibrosis originate from
adiponectin-positive intradermal progenitors. Arthritis Rheumatol.
67:1062–1073. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wang QA, Song A, Chen W, Schwalie PC,
Zhang F, Vishvanath L, Jiang L, Ye R, Shao M, Tao C, et al:
Reversible de-differentiation of mature white adipocytes into
preadipocyte-like precursors during lactation. Cell Metab.
28:282–288. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zoico E, Darra E, Rizzatti V, Budui S,
Franceschetti G, Mazzali G, Rossi AP, Fantin F, Menegazzi M, Cinti
S and Zamboni M: Adipocytes WNT5a mediated dedifferentiation: A
possible target in pancreatic cancer microenvironment. Oncotarget.
7:20223–20235. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zoico E, Darra E, Rizzatti V, Tebon M,
Franceschetti G, Mazzali G, Rossi AP, Fantin F and Zamboni M: Role
of adipose tissue in melanoma cancer microenvironment and
progression. Int J Obes (Lond). 42:344–352. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Chirumbolo S and Bjorklund G: Can wnt5a
and wnt non-canonical pathways really mediate adipocyte
de-differentiation in a tumour microenvironment? Eur J Cancer.
64:96–100. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ye X and Weinberg RA:
Epithelial-mesenchymal plasticity: A central regulator of cancer
progression. Trends Cell Biol. 25:675–686. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Whatcott CJ, Diep CH, Jiang P, Watanabe A,
LoBello J, Sima C, Hostetter G, Shepard HM, Von Hoff DD and Han H:
Desmoplasia in primary tumors and metastatic lesions of pancreatic
cancer. Clin Cancer Res. 21:3561–3568. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Bluher M: Adipose tissue dysfunction in
obesity. Exp Clin Endocrinol Diabetes. 117:241–250. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Khandekar MJ, Cohen P and Spiegelman BM:
Molecular mechanisms of cancer development in obesity. Nat Rev
Cancer. 11:886–895. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Lemoine AY, Ledoux S and Larger E: Adipose
tissue angiogenesis in obesity. Thromb Haemost. 110:661–668. 2013.
View Article : Google Scholar : PubMed/NCBI
|