Introduction
Renal cell carcinoma (RCC) is the most prevalent
type of cancer of the urinary system, and is characterized by being
a highly malignant tumor (1).
Based on its morphological classification, RCC can be mainly
divided into three subtypes: i) Kidney renal clear cell carcinoma
(KIRC), ii) kidney renal papillary cell carcinoma and iii)
suspicious cell malignant tumors. KIRC accounts for >70% of RCC
cases (2). RCC is one of the 10
most common cancer types worldwide, causing nearly 140,000
mortalities annually (3). At
present, the main treatment for localized RCC is surgical
treatment, while immunotherapy, targeted drugs and chemotherapy are
the treatment of choice for advanced and metastatic cases (4). Despite advancements in treatment
strategies, the 5-year overall survival (OS) rate is just 12% for
patients with metastatic KIRC. However, ~16% of patients already
have distant metastases upon their initial diagnosis of RCC
(5). Therefore, although
significant advances in diagnostic techniques and targeted
therapies have been made, the prognosis remains poor for the
majority of patients (2,6). The high recurrence and incidence
rates of KIRC emphasize the urgency to find novel molecular targets
for disease treatment.
Ribosomal proteins (RPs) are one of the main
components of ribosomes, which play key roles in the regulation of
intracellular protein biosynthesis. Specifically, RPs can
coordinate interactions between ribosomes, genes, elongation
factors and initiators (7). The
unique extra-ribosomal effects and functions of distinct RPs have
been previously reported (8).
These effects are involved in regulating cell proliferation,
differentiation and apoptosis, which are crucial functions for cell
proliferation and development. Although several studies have found
abnormal expression patterns of different RPs in a variety of
diseases, the specific roles of these proteins and their
participation in the underlying molecular mechanisms in the
development of human cancer remain unclear.
Ribosomal protein S20 (RPS20) belongs to the S10P
family of RPs (9). RPS20 is mainly
involved in the regulation of ribosomal RNA processing (10). It is primarily localized in the
cytoplasm (11), unlike other RPs,
which are usually found in the nucleoli (12–17).
Recent evidence suggested that RPS20 is involved in non-ribosomal
regulation. RPS20 was shown to be involved in regulating the
p53-mouse double minute 2 homolog signaling pathway (18–29).
Furthermore, the interaction between GNL1 and RPS20 was found to be
capable of regulating cell proliferation (30). RPS20 has also been identified to
act as an oncogene in a variety of tumors. However, the specific
role of RPS20 in the onset and advancement of RCC remains to be
elucidated.
The aim of the present study was to validate the
high expression levels of RPS20 in RCC [as observed in The Cancer
Genome Atlas (TCGA) database] and to explore its regulatory
mechanism. In line with our in silico analyses, RPS20 was
observed to be highly expressed in RCC tissues. The lentiviral
transduction knockdown results further indicated that the
inhibition of RPS20 expression could significantly reduce the
proliferation, migration, and invasion of RCC cells in
vitro. Similarly, RPS20 knockdown considerably inhibited the
growth of subcutaneous tumors in nude mice. Furthermore, inhibiting
RPS20 expression in an RCC cell line reduced the expression of
CDK4, cyclin D1 and N-cadherin, and increased the expression of
E-cadherin. Lastly, RPS20 was revealed to positively regulate
several downstream signaling pathways, including the mTOR and ERK
pathways.
Materials and methods
In silico analyses using the Oncomine
and TCGA databases
The mRNA expression levels of RPS20 in different
tumors were explored using the Oncomine database (31). The threshold and query details were
set as follows: Fold-change=2, P=0.05, checked (all) gene ranking
and mRNA data. The gene expression profile and clinical records
from patients with KIRC were obtained from TCGA website (https://portal.gdc.cancer.gov/), which contained
539 tumor samples and 72 normal samples (32). R software version 3.6.3 with
Strawberry Perl was used for data processing. Samples with
incomplete information were removed from the datasets before
performing statistical analyses.
In silico analysis based on the Gene
Expression Profiling Interactive Analysis (GEPIA) database
GEPIA is an open-source database from which
expression data of RNA sequencing can be obtained from 10,000 tumor
and normal samples (33). Using
this tool, the association of high RPS20 expression with the OS and
progression-free survival (PFS) of patients with KIRC were
analyzed. Moreover, differential gene expression of RPS20 among
different tumor types was investigated using the Tumor IMmune
Estimation Resource (TIMER) database (https://cistrome.shinyapps.io/timer/).
Clinical patients and tissue
samples
In total, 43 RCC tissue samples were collected from
patients with renal cancer undergoing partial or radical
nephrectomy at The Second Affiliated Hospital of Nantong University
(Nantong, China) between January 2017.01 and December 2019, and
were compared with their normal tissue counterparts. The tissue
samples were immediately stored at −80°C upon collection. Human
studies were approved (approval no. 2021YL012) by the Ethics
Committee of The Second Affiliated Hospital of Nantong University
(Nantong, China) according to the Declaration of Helsinki of 1964.
Written informed consent was provided by all participating
patients.
Cell culture
Human RCC cell lines (786-O, ACHN and OS-RC-2) were
obtained from the Shanghai Institute of Biological Sciences. ACHN
cells were cultured in MEM (Gibco; Thermo Fisher Scientific, Inc.)
containing 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.), whereas 786-O and OS-RC-2 cells were cultured in
RPMI-1640 medium with 10% FBS (Gibco; Thermo Fisher Scientific,
Inc.). All cell lines were maintained in a sterile incubator
(Thermo Fisher Scientific, Inc.) at 37°C with 5%
CO2.
RPS20 knockdown using lentiviral
transduction
Cells were infected using lentiviral particles
containing short hairpin (sh)RPS20 constructs to establish stable
RPS20 knockdown cell lines. The shRPS20 sequence used for knockdown
was 5′-GATCGTTTCCAGATGAGAATT-3′, while the shRNA control sequence
used was 5′-TTCTCCCGAACGTGTCACG-3′. RPS20 knockdown lentiviral
particles (designated as LV-shRPS20) and negative control (NC)
GV248 vector (designated as LV-shNC) were used to infect 786-O and
OS-RC-2 cells (MOI=5) following the manufacturer's instructions
(Shanghai GeneChem Co., Ltd.). After 48 h of transfection, cells
were collected for additional studies. The success of gene
knockdown was evaluated by observing the generation of green
fluorescent cells under a fluorescence microscope and by using
puromycin selection (3 µg/ml). The knockdown efficiency of RPS20
was then confirmed via reverse transcription-quantitative PCR
(RT-qPCR) and western blotting.
Western blotting
Protease inhibitors and ice-cold RIPA buffer were
premixed with fresh RCC patient tissues or cells to extract
proteins. The supernatant was collected, and the concentration of
total protein was quantified by utilizing a BCA protein assay kit
(Thermo Fisher Scientific, Inc.). Western blotting was performed as
previously described (34). The
following primary antibodies were used: Anti-RPS20 (cat. no.
15692-1-AP; 1:2,000), anti-β-actin (cat. no. 81115-1-RR; 1:5,000),
anti-AKT (cat. no. 60203-2-lg; 1:1,000), anti-phosphorylated
(p)-AKT (cat. no. 28731-1-AP; 1:1,000), anti-ERK (cat. no.
51068-1-S-AP; 1:1,000), anti-p-ERK (cat. no. 28733-1-AP; 1:1,000),
anti-cyclin D1 (cat. no. 26939-1-AP; 1:2,000), anti-CDK4 (cat. no.
11026-1-AP; 1:1,000), anti-E-cadherin (cat. no. 20874-1-AP;
1:5,000) and anti-N-cadherin (cat. no. 22018-1-AP; 1:1,000; all
purchased from ProteinTech Group, Inc.). HRP-conjugated goat
anti-rabbit IgG (cat. no. PR3001; 1:5,000) and HRP-conjugated goat
anti-mouse IgG (cat. no. PR3002; 1:5,000; both purchased from
ProteinTech Group, Inc.) were the secondary antibodies used in the
present study.
Extraction of RNA and RT-qPCR
TRIzol® (Qiagen, Inc.) was used to
extract total RNA from RCC cells and tissues. Synthesis of cDNA was
performed using the Thermo-Script Reverse Transcription kit (Thermo
Fisher Scientific, Inc.), following the reagent manufacturer's
instructions. qPCR was performed in 10-µl reactions loaded into
96-well plates with SYBR Green reagent (Takara Bio, Inc.) using the
CFX96™ Real-Time PCR system (Bio-Rad Laboratories, Inc.). The
thermocycling conditions were as follows: 95°C for 5 min, followed
by 40 cycles of 95°C for 15 sec, 60°C for 25 sec and 72°C for 30
sec. Relative gene expression was calculated using the
2−ΔΔCq method (35).
Gene expression was normalized using β-actin as a
housekeeping gene. The following primer sequences were utilized:
RPS20 forward, 5′-ATCACCCTAACAAGCCGCAA-3′ and reverse,
5′-AGGCATTCGAACTGGTCCTT-3′; and actin forward,
5′-GGGCATGGGTCAGAAGGATT-3′ and reverse,
5′-CATGTCGTCCCAGTTGGTGA-3′.
Immunohistochemistry (IHC)
Collected RCC and adjacent normal tissues were
subjected to IHC analysis. After incubation at 60°C for 60 min, the
paraffin-embedded tissue sections were deparaffinized in xylene and
then immersed in graded ethanol solutions for hydration. The slides
were blocked for 5 min using Ultra V Block (Shilian Boyan
Technology Co., Ltd), and the tissue sections were subsequently
subjected to overnight incubation at 4°C with an anti-RPS20 primary
antibody (1:100; ProteinTech Group, Inc.). Next, the slides were
washed with PBS and incubated with a anti-rabbit IgG (cat. no.
15692-1-AP; 1:200; ProteinTech Group, Inc.) secondary antibody at
37°C for 10–30 min. Finally, the tissue samples were stained with
diaminobenzidine before evaluation by light microscopy. Specimens
were categorized as negative, positive, ++ positive, or +++
positive based on the total of the staining intensity and staining
extent scores, which ranged from weak to strong.
Cell Counting Kit (CCK)-8 assay
CCK-8 assay (Dojindo Laboratories, Inc.) was
utilized to evaluate the proliferation rate of RCC cells. Cells
transduced with either shRPS20 or shNC were seeded in five 96-well
plates with a cell density of 3×103 cells/well in
triplicates. The proliferation of cells was observed daily for 3
days by incubating the cells with CCK-8 solution (10 µl) and
serum-free medium (190 µl) for at 37°C for 2 h. The colorimetric
absorbance at 450 nm was estimated using a microplate reader.
Colony formation assay
Transduced OS-RC-2 and 786-O cells, as well as
control cells were seeded into six-well plates at a cell density of
800 cells/well. RPMI-1640 with 20% FBS was used for culturing the
cells for 7 days at 37°C with 5% CO2. Colony fixation
was performed using 4% paraformaldehyde solution at room
temperature for 30 min, followed by washes with PBS and stained
using a 0.1% crystal violet solution at room temperature for 5 min,
then visualized and counted manually. Colonies consisted of >50
cells.
Wound healing test
Transduced OS-RC-2 and 786-O cells, along with their
corresponding control cells, were seeded into six-well plates. When
the cell layer reached 100% cell confluency, a scratch wound was
made on it with a pipette tip. Cells were cultured in serum-free
medium. The migration rates of the two cell groups were evaluated
through light microscopy. Images of the cell layers were captured
every 6 h.
Invasion assay
Matrigel diluted with serum-free medium (1:6
dilution) was added to the cell culture chamber, and after Matrigel
solidification, the cells were cultured. The upper chambers of a
Transwell plate (8.0-µm pore size polycarbonate filter) were filled
with 200 µl basal serum-free medium, while in the lower chamber,
600 µl complete medium containing 20% FBS was added. Cells were
cultured for 1 day at 37°C with 5% CO2. A cotton swab
was used for removing the residual cells from the upper chamber
surface. Cells that had migrated to the lower chamber were fixed in
paraformaldehyde solution (4%) at room temperature for 30 min,
stained using crystal violet (0.1%) at room temperature for 5 min.
Then, the mean number of cells was computed by randomly picking
five fields under a light microscope and calculating the number of
cells in each.
Flow cytometry
Transduced OS-RC-2 and 786-O cells, and their
corresponding control cells were seeded into separate dishes for 24
h. Following two washes with ice-cold PBS, the cells were fixed in
ethanol solution (70%) overnight at 4°C, trypsinized in PBS (100
µl) and stained at room temperature with propidium iodide (20
µg/ml) for 30 min. Analysis of the cell cycle was performed (ModFit
LT 4.1 software) on the different groups of cells using the Beckman
MoFlo XDP instrument (A00-1-1102; Beckman Coulter, Inc.).
In vivo xenograft experiments
Female four-week-old BALB/c nude mice (n=10; weight,
16–18 g) were bred at the Animal Research Center of Nantong
University. Transduced 786-O-shNC and 786-O-shRPS20 cell
resuspension of 100 µl (~5.0×106) was collected and
implanted in the right armpits of each nude mouse. The animals were
housed in microisolator cages with autoclaved bedding with food and
water provided ad libitum. The mice were maintained on a
daily 12/12-h light/dark cycle. The tumor volume and total body
weight of the mice were recorded every other week. The tumor volume
was calculated based on the following formula: Volume
(mm3)=length × width2 × 0.52. After the
tumors reached a volume of ~1,000 mm3, the mice were
euthanized using cervical dislocation, and the tumors were removed,
measured and weighed in preparation for further experiments. The
animal experiments were approved (approval no. S20210227-041) by
the Animal Ethics Committee of Nantong University (Nantong, China)
and the experiments were conducted according to the National
Institutes of Health Guide for the Care and Use of Laboratory
Animals.
Statistical analysis
GraphPad software (version 5.02; GraphPad Software,
Inc.) was applied for statistical analyses. The experiments were
performed three times for validating their reproducibility. The
relationship between RPS20 expression and stage and tumor size was
examined using the Kruskal test. The correlation between RPS20
expression and lymph node status and distant metastasis was
evaluated by the Wilcox test. Moreover, logistic regression
analysis was also applied to estimate the correlation between gene
expression and clinicopathological parameters. Univariate and
multivariate Cox analyses were adopted to evaluate the value of the
RPS20 gene as a prognostic indicator. Kaplan-Meier analysis with
the log-rank test was used to conduct the survival study.
Statistical comparisons across two groups were performed with a
two-tailed unpaired Student's t-test, while one-way ANOVA with
Tukey's post hoc test was used for the comparison of multiple
groups. *P<0.05 and **P<0.01 were considered to indicate a
statistically significant difference.
Results
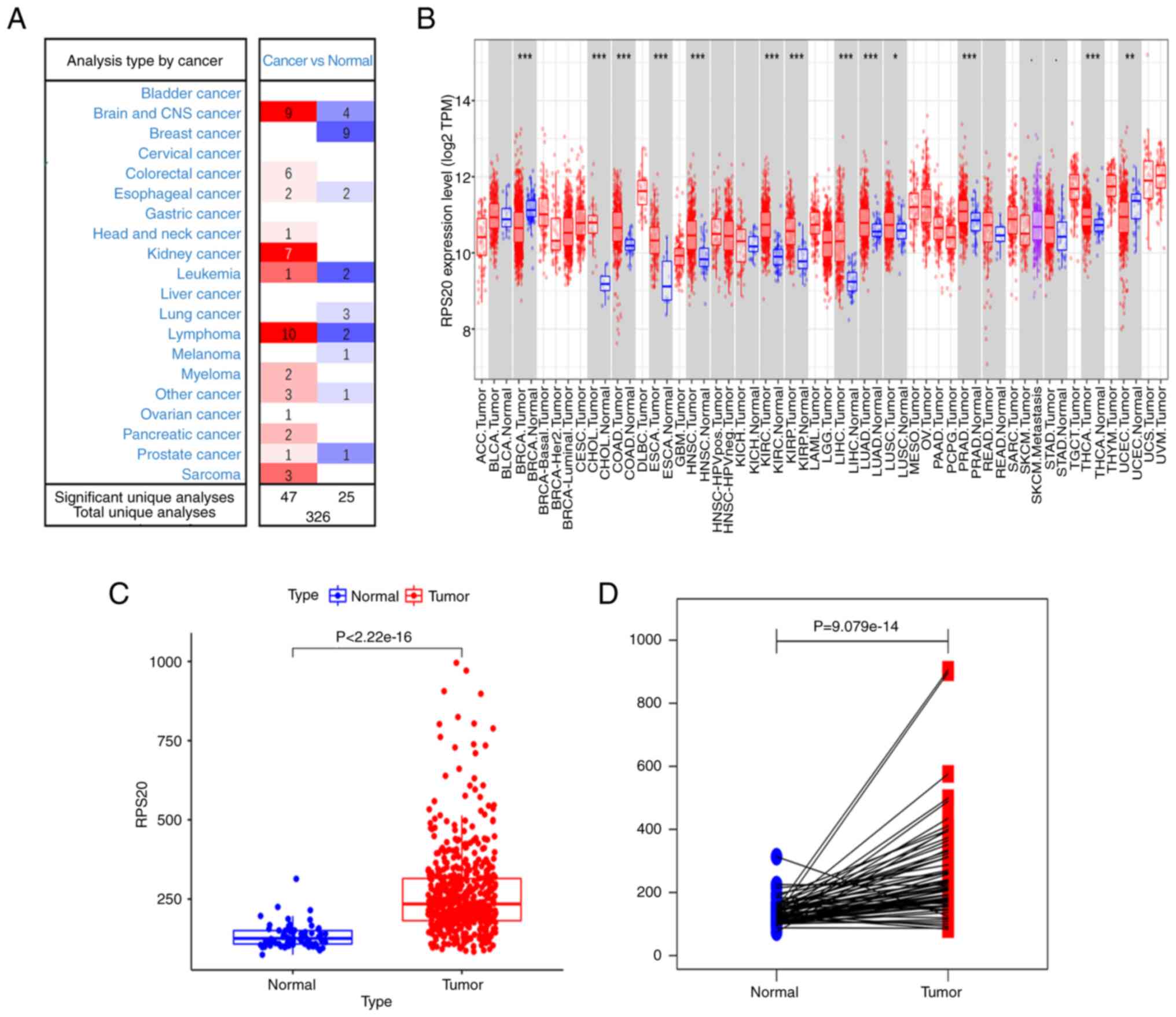
RPS20 expression in human tumors
Using the Oncomine database to evaluate seven
different datasets, higher expression of RPS20 mRNA was found in
RCC tissues compared with that in normal samples based on the set
threshold. Furthermore, RPS20 was found to be highly expressed in
the majority of brain tumors, lymphomas and sarcomas, while its
expression was lower in breast tumors and leukemias (Fig. 1A). Similarly, using TIMER, the
RPS20 gene was found to be differentially expressed in tumor
tissues in comparison with its expression in adjacent normal
tissues in patients with RCC (P<0.001; Fig. 1B). Additional analyses using the
data retrieved from TCGA database showed that KIRC tissues (n=539)
had higher RPS20 mRNA expression levels than adjacent normal
tissues (n=72; Fig. 1C and D).
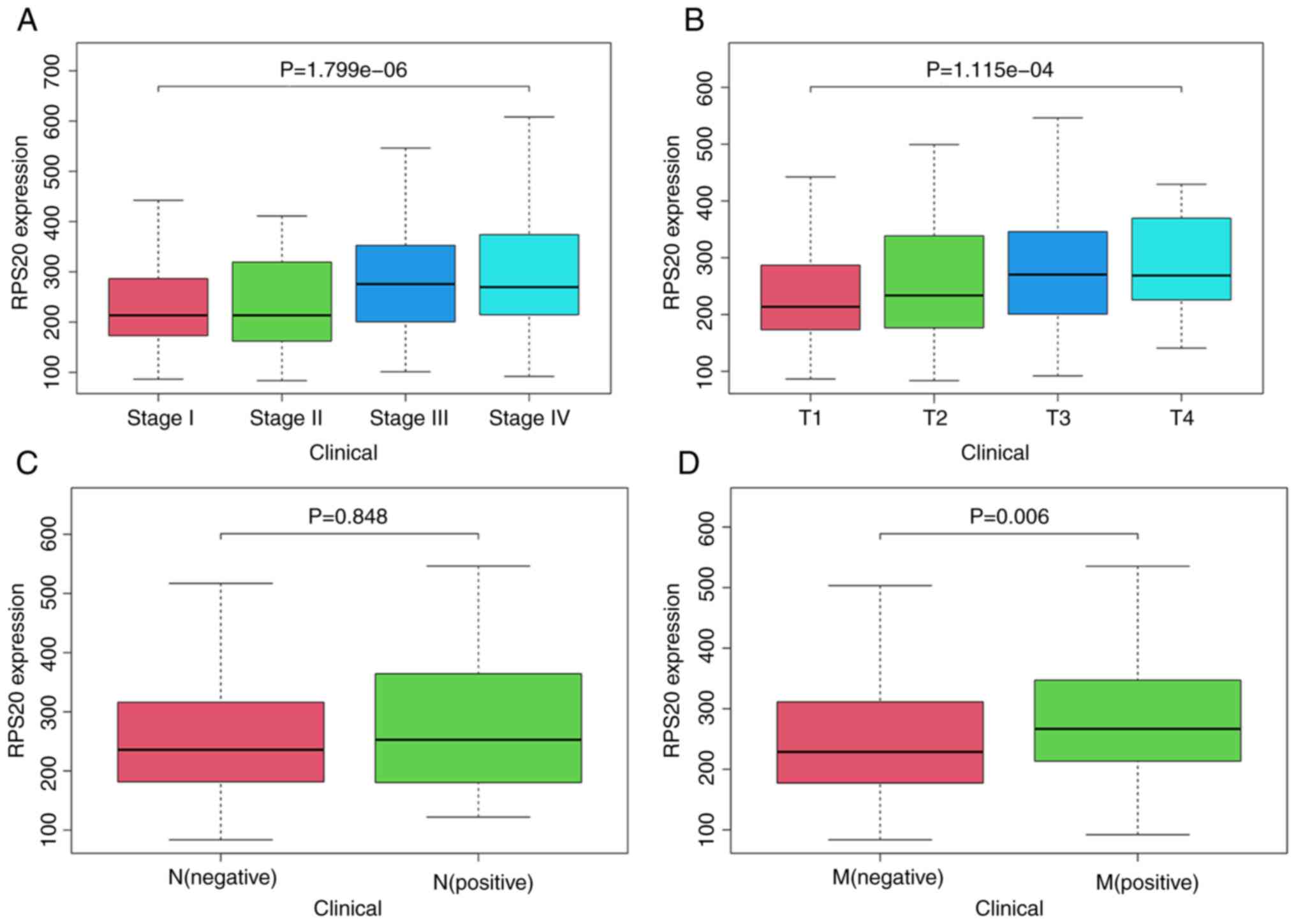
Association of RPS20 expression and
clinicopathological variables
The association between RPS20 expression and various
clinicopathological characteristics of patients with RCC was
investigated, since RPS20 is highly expressed in RCC.
Kruskal-Wallis test showed that RPS20 mRNA levels were correlated
with disease stage (P<0.001) and tumor size (P<0.001;
Fig. 2A and B). Wilcoxon test
showed that high expression of RPS20 was associated with distant
metastasis (P=0.006; Fig. 2D).
Notably, RPS20 expression had no association with lymph node
metastasis (P=0.848). It was hypothesized that this was due to the
removal of 289 samples with unknown transfer details (Fig. 2C). Furthermore, logistic regression
revealed associations of RPS20 expression with sex (P<0.001),
disease stage (P=0.010) and T stage (P=0.001), but not with age,
distant metastasis or lymph node metastasis (Table I). Kaplan-Meier survival analysis
revealed that RPS20 expression was associated with the OS of
patients with RCC (P=0.006; Fig.
4B).
 | Table I.Association between expression of
RPS20 and various clinicopathological parameters. |
Table I.
Association between expression of
RPS20 and various clinicopathological parameters.
|
|
| Expression level of
RPS20 |
|
|
|---|
| Clinicopathological
variables | Number of
cases |
|
|
|
|---|
| Low | High | χ2 | P-value |
|---|
| All cases | 250 | 125 | 125 |
|
|
| Sex |
|
|
| 33.865 | 0.000 |
|
Male | 99 | 72 | 27 |
|
|
|
Female | 151 | 53 | 98 |
|
|
| Age |
|
|
| 0.148 | 0.701 |
|
<60 | 105 | 51 | 54 |
|
|
|
≥60 | 145 | 74 | 71 |
|
|
| Tumor stage |
|
|
| 6.624 | 0.01 |
| I or
II | 136 | 81 | 55 |
|
|
| III or
IV | 114 | 44 | 71 |
|
|
| T grade |
|
|
| 11.306 | 0.001 |
| T1 or
T2 | 148 | 84 | 64 |
|
|
| T3 or
T4 | 102 | 41 | 61 |
|
|
| Distant
metastasis |
|
|
| 0.139 | 0.709 |
|
Negative | 209 | 106 | 103 |
|
|
|
Positive | 41 | 19 | 21 |
|
|
| Lymph nodes |
|
|
| 0.638 | 0.424 |
|
Negative | 235 | 119 | 116 |
|
|
|
Positive | 15 | 6 | 9 |
|
|
| Years of
survival |
|
|
| 1.939 | 0.164 |
|
<5 | 197 | 94 | 103 |
|
|
| ≥5 | 53 | 31 | 22 |
|
|
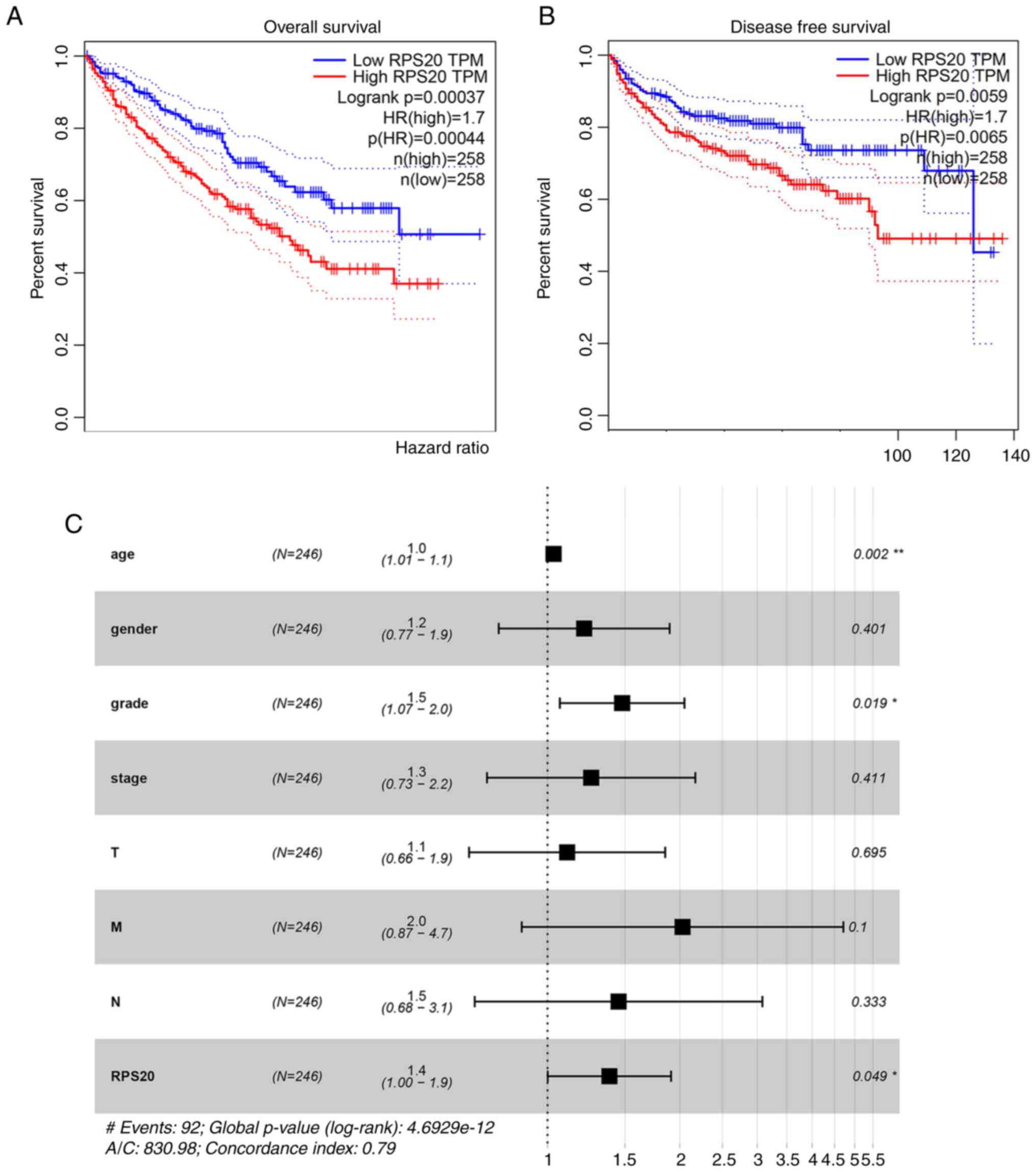
Prognostic potential of RPS20
expression in RCC
Using GEPIA, a negative correlation between RPS20
expression and OS was found [hazard ratio (HR)=1.7; P=0.00044;
Fig. 3A] and DFS (HR=1.7;
P=0.0065; Fig. 3B). To validate
these findings, data from TCGA was further analyzed. Univariate Cox
analysis revealed that OS was associated with RPS20 expression
(HR=1.03; P<0.05), age (HR=1.02; P<0.05), disease stage
(HR=1.86; P<0.001), tumor size (HR=1.94; P<0.001), lymph node
status (HR=4.07; P<0.001) and distant metastasis (HR=2.93;
P<0.01) (Table II).
Multivariate analysis revealed that age (HR=1.03; P<0.05) and
RPS20 expression (HR=1.38; P<0.05) were independent prognostic
values (Table II and Fig. 3C).
| Table II.Univariate and multivariate Cox
analyses of clinicopathological parameters and overall survival in
patients with clear cell renal cell carcinoma. |
Table II.
Univariate and multivariate Cox
analyses of clinicopathological parameters and overall survival in
patients with clear cell renal cell carcinoma.
|
| Univariate
analysis | Multivariate
analysis |
|---|
| Clinicopathological
parameters |
|
|
|---|
| HR (95% CI) | P-value | HR (95% CI) | P-value |
|---|
| Age | 1.02
(1.00-1.04) | 0.012 | 1.03
(1.01-1.05) | 0.001 |
| Sex | 1.01
(0.67-1.54) | 0.951 | 1.21
(0.77-1.90) | 0.400 |
| Grade | 2.24
(1.68-2.99) | 0.000 | 1.48
(1.07-2.05) | 0.018 |
| Stage | 1.86
(1.54-2.25) | 0.000 | 1.26
(0.73-2.17) | 0.411 |
| T | 1.94
(1.54-2.46) | 0.000 | 1.11
(0.66-1.85) | 0.695 |
| N | 4.07
(2.63-6.30) | 0.000 | 2.03
(0.87-4.71) | 0.099 |
| M | 2.93
(1.52-5.67) | 0.001 | 1.45
(0.68-3.08) | 0.333 |
| Ribosomal protein
S20 | 1.00
(1.00-1.00) | 0.025 | 1.38
(1.00-1.90) | 0.049 |
Expression of RPS20 in KIRC
tissues
The differential expression of RPS20 was validated
via western blotting and IHC in various RCC cell lines as well as
43 paired RCC and normal tissue samples. High expression of RPS20
in the three analyzed RCC cell lines and renal cell tissues were
revealed by western blotting (Fig. 4A
and B). Similarly, the IHC results showed a significant
upregulation of RPS20 in RCC samples (Fig. 4C).
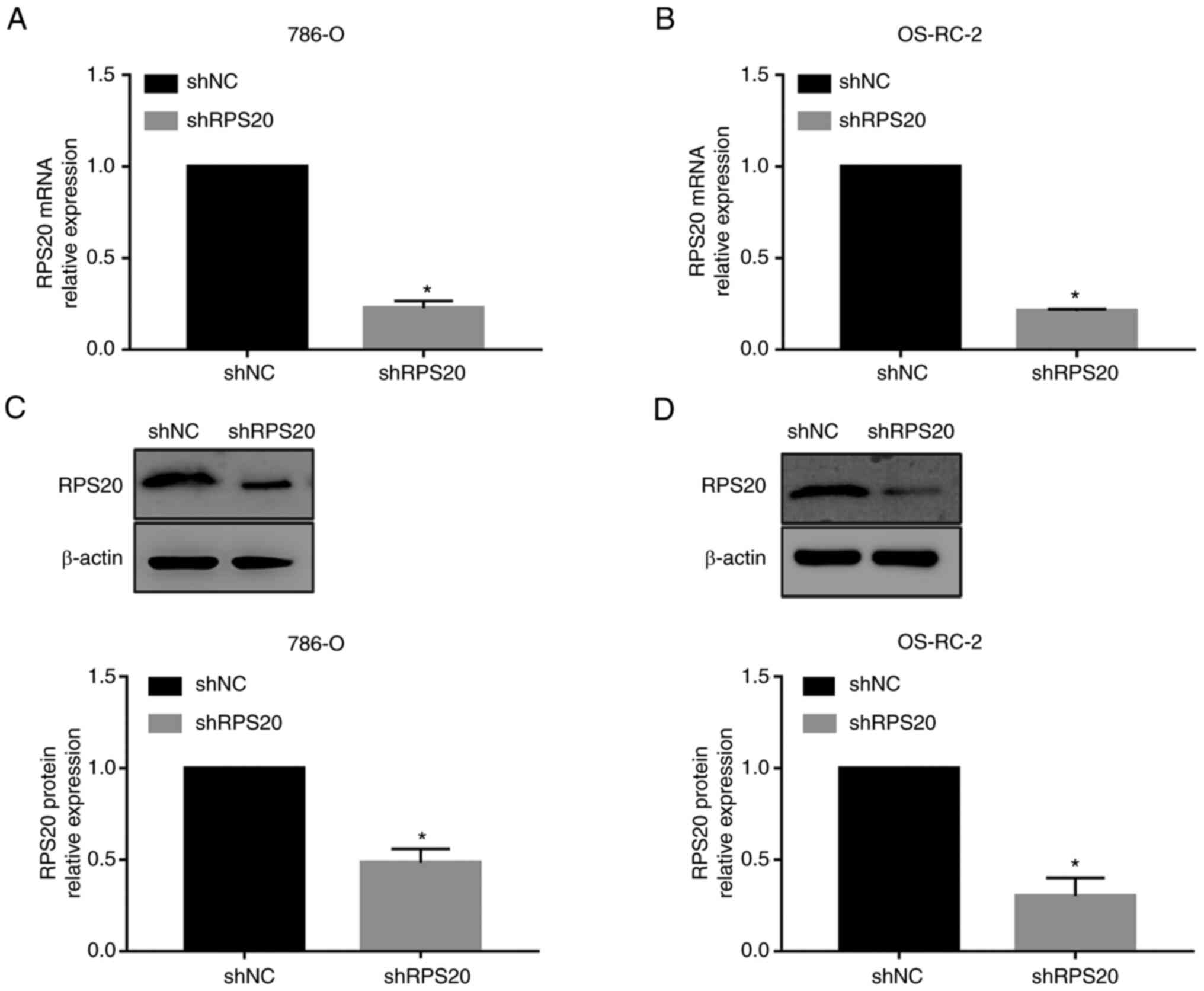
RPS20 is downregulated in 786-O and
OS-RC-2 RCC cell lines
To investigate the role of RPS20 in RCC, shRNA
constructs for RPS20 were generated. The 786-O and OS-RC-2 KIRC
cell lines were transduced with RPS20 shRNA and control lentiviral
particles to construct stable RPS20 knockdown cell lines.
Successful knockdown of RPS20 was confirmed by RT-qPCR which showed
a 90% decrease in the expression levels of RPS20 mRNA in the
aforementioned cell lines compared with those in the control vector
group (P<0.01; Fig. 5A and B).
Similarly, RPS20 protein levels were measured by western blotting,
and the knockdown efficiency was found to exceed 50% (Fig. 5C and D).
Knockdown of RPS20 suppresses the
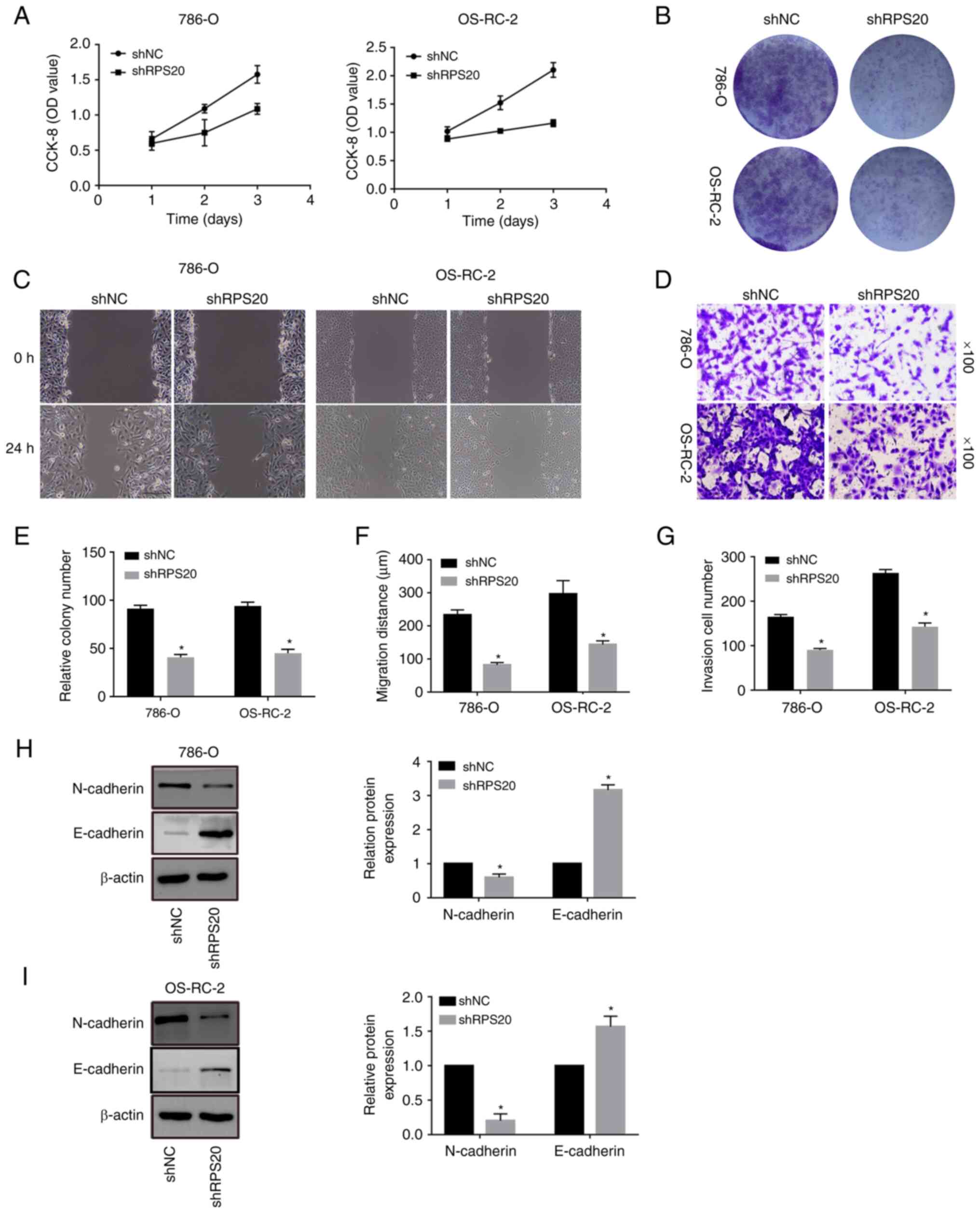
proliferation and mobility of RCC cells in vitro
To further elucidate the effect of RPS20 on the
proliferation of RCC cells, CCK-8 assays were performed on the
knockdown cell lines generated in the present study. The results
showed that the proliferation of RPS20 knockdown cell lines was
significantly inhibited compared with that of control cells
(Fig. 6A). Colony formation assay
was used to assess the possible function of RPS20 in cell
proliferation. Similarly, RPS20 knockdown cells formed fewer
colonies than control cells (Fig. 6B
and E). Furthermore, wound-healing assays were performed on the
RPS20 knockdown cell lines (Fig. 6C
and F). Knocking down RPS20 considerably led to inhibition of
RCC cell invasion and migration, according to the results of
Transwell assays (Fig. 6D and G).
Consistent with these findings, RPS20 knockdown cells displayed a
significantly increased E-cadherin level and a decreased N-cadherin
level (Fig. 6H and I).
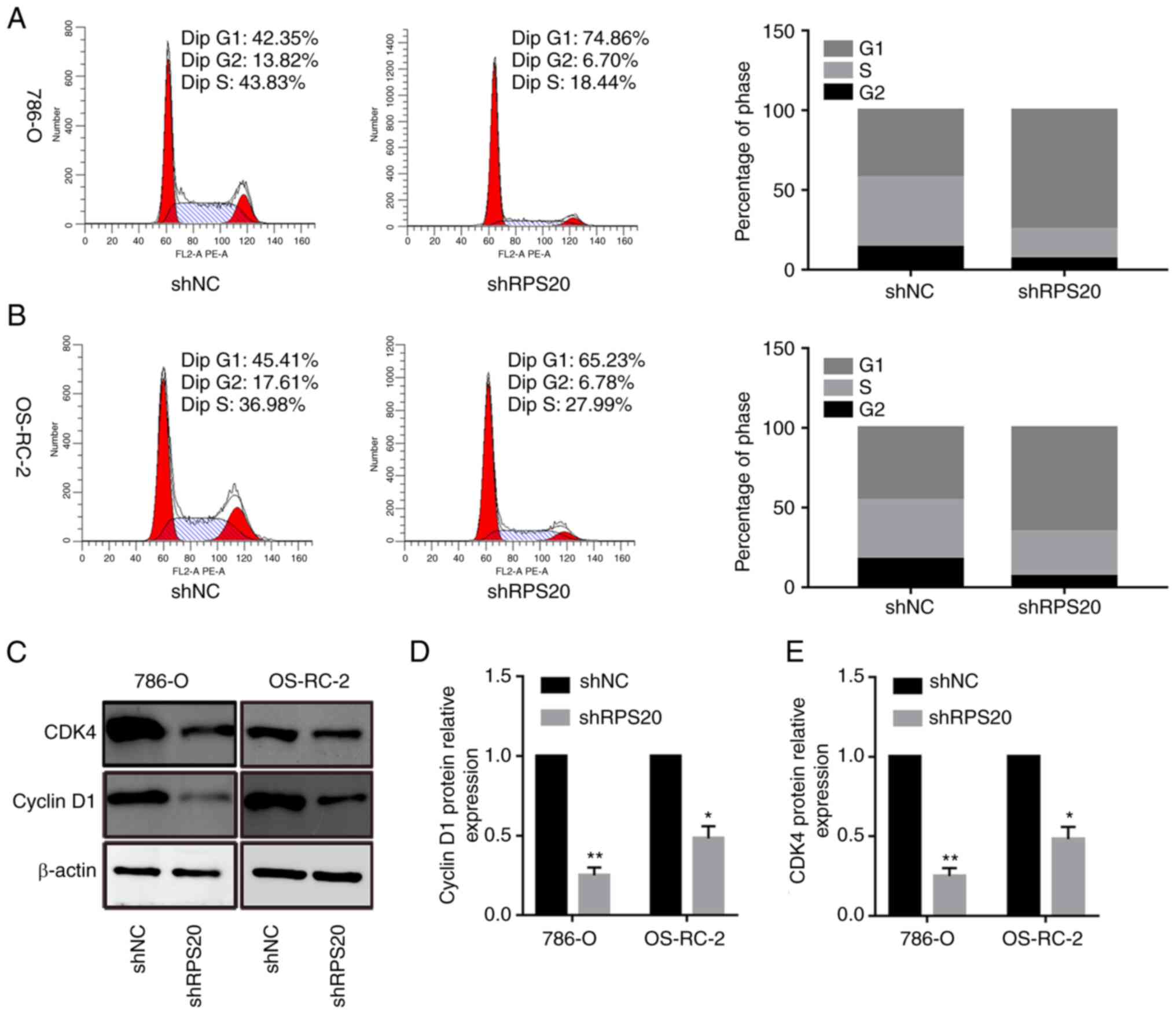
RPS20 knockdown induces cell cycle
arrest
To investigate the mechanism by which RPS20
contributed to the proliferation of 786-O and OS-RC-2 cells, cell
cycle analysis was performed. The results showed that the
proportion of cells in the G0/G1 phase was
74.86 and 65.23% in LV-shRPS20 786-O and OS-RC-2 cells,
respectively, while it was 42.35 and 45.41% in LV-shNC 786-O and
OS-RC-2 cells, respectively (P<0.05; Fig. 7A and B). These data suggested that
the cell cycle was arrested in G0/G1 phase
after RPS20 was knocked down. Consistent with these results, the
expression levels of the cell cycle-associated proteins CDK4 and
cyclin D1 were found to be lower in shRPS20 786-O and OS-RC-2 cells
(Fig. 7C-E).
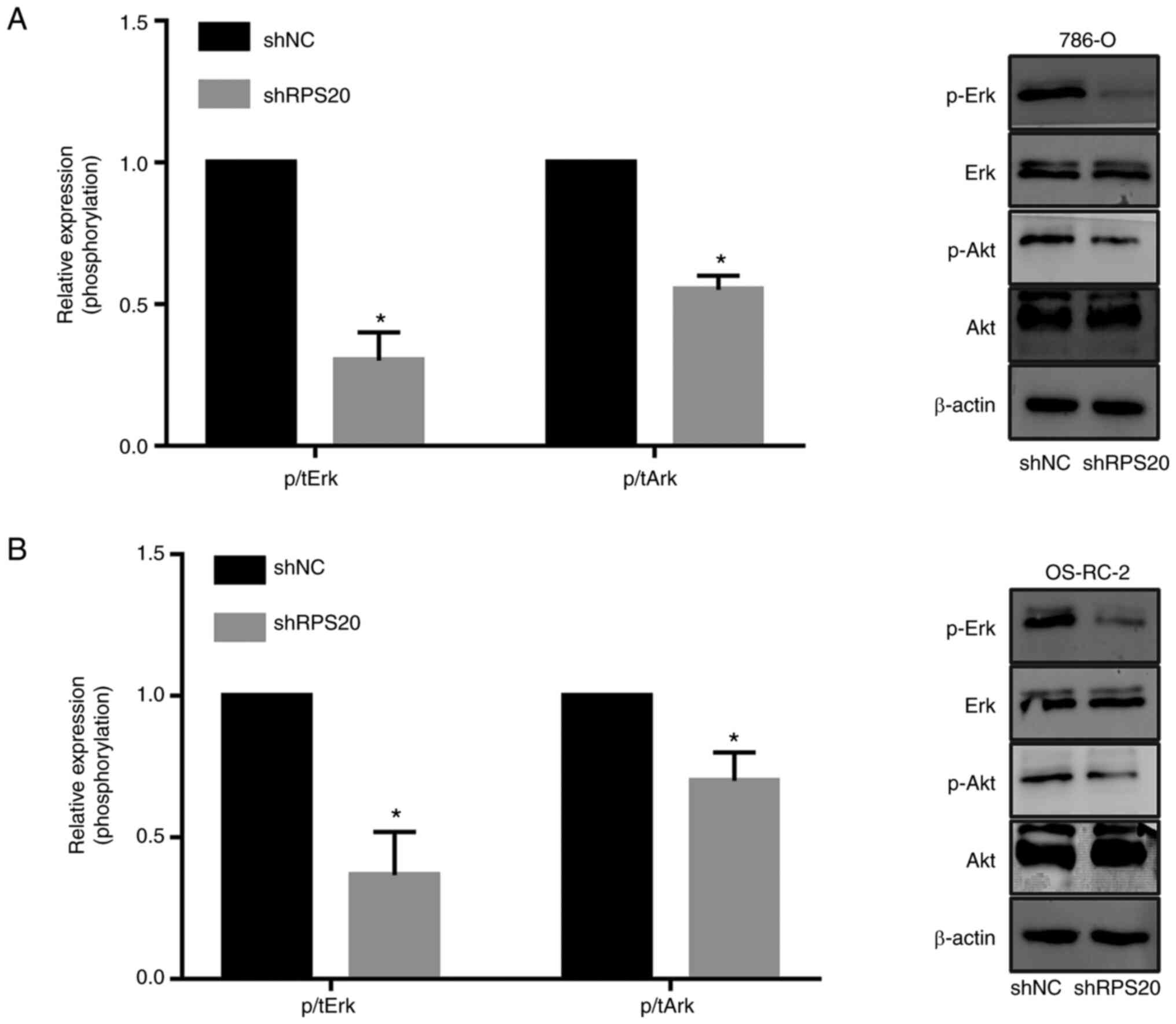
RPS20 knockdown inhibits the mTOR and
ERK signaling pathways
To explore the signaling pathways that may be
regulated by RPS20, the effects of knocking down RPS20 in the mTOR
and ERK signaling pathways were evaluated using western blotting.
The results revealed that knocking down RPS20 considerably
inhibited the phosphorylation of AKT and ERK, while total AKT and
ERK protein levels were not affected (Fig. 8A and B). These results suggested a
key role of RPS20 in the ERK-MAPK and AKT-mTOR signaling
pathways.
RPS20 promotes tumor development in
vivo
To further support the present in vitro
experiments, the current study further analyzed in vivo
whether RPS20 shRNA may affect tumorigenesis. For establishing
xenograft mice models, cell suspensions of the aforementioned RPS20
knockdown cell lines were implanted into both armpits of each nude
mouse. The results revealed that tumor growth was considerably
inhibited in the RPS20 knockdown group compared with that of the
control group (Fig. 9A and B).
Consistently, tumor weight in the RPS20 knockdown group was
significantly lower compared with that of the control group
(Fig. 9C and D). Overall, these
results indicated that RPS20 knockdown may lead to the inhibition
of RCC tumor cell proliferation in vivo.
Discussion
Kidney cancer is a heterogeneous disease. Current
evidence suggests that the majority of RCCs develop due to several
factors, including dysregulation of hypoxia-inducible factor
signaling, mutations in key histone and chromatin-modifying
enzymes, and metabolic reprogramming of cellular metabolism
(36,37). While advancements in diagnostic
techniques and targeted therapies have been achieved in recent
decades, disease prognosis remains unsatisfactory. Thus, screening
for new biomarkers may offer novel strategies for diagnosing and
treating KIRC. The rapid advancement of high-throughput sequencing
technologies and bioinformatics techniques may provide novel
methods for diagnosing and treating KIRC.
Various RPs have critical importance in the onset
and progression of different human tumors. For example, RPL11
interacts with Myc and suppresses its transcriptional activity
(38), while RP S29 induces cell
apoptosis by regulating p53 and Bcl-2 (39). Moreover, abnormal RP expression can
lead to non-neoplastic diseases such as hemochromatosis and anemia
(40). Previous research has
slowly unveiled the role of RPS20 outside of the ribosome. Cell
proliferation can be regulated by human nucleolar GNL1 and RPS20
interaction (30). The
Mdm2-p53-MdmX network may be controlled by the ribosomal proteins
RPL37, RPS15 and RPS20 (41).
RPS20 mutations enhance the risk of developing hereditary
non-polyposis colorectal cancer (42). According to the findings of a
recent bioinformatics study, the member of the ribosomal family
known as RPS20 may be helpful as a prognostic predictor in patients
diagnosed with ccRCC (43). The
function of RPS20 in ccRCC still needs to be fully understood. The
findings of the present study suggested that RPS20 is upregulated
in ccRCC tissues and cell lines, associated with
clinicopathological characteristics (grade, stage) in ccRCC
patients. Furthermore, it was found that the knockdown of RPS20 may
limit cell proliferation, migration, and invasion via inhibition of
MAPK and AKT signaling pathways.
The multistep and intricate process of tumor
invasion and metastasis has become a significant barrier to the
clinical treatment of several malignancies (44,45).
Furthermore, metastasis can result in ccRCC treatment failure,
which lowers 5-year survival rates (5). As a result, limiting cell migration
and invasion can effectively control cancer cell spread. According
to the results of the current investigation, migration and invasion
assays revealed that RPS20 knockdown significantly reduced the
invasion and metastasis of ccRCC cells.
Additionally, patients with ccRCC have a poor
prognosis when MAPK and AKT signaling pathways are activated
(46). The MAPK pathway comprises
three proteins: ERK, JNK and p38. ERK is a crucial signal
transducer for cell survival, and JNK and p38 help cells learn to
invade and migrate (47). There is
growing evidence that MAPKs may stop tumor invasion and metastasis
in various tumor types (48,49).
AKT signaling is additionally engaged in numerous biological
processes, including glucose metabolism and cell cycle,
particularly in cancer cells (50). AKT signaling is suppressed in ccRCC
cells, which reduces invasion and metastasis (51). In the present study, it was
demonstrated that RPS20 knockdown suppresses MAPK and AKT signaling
in ccRCC.
To conclude, the current results clearly
demonstrated that RPS20 was upregulated in human RCC tissues.
Mechanistically, it was found that RPS20 protein could promote not
only the proliferation and migration but also the invasion of RCC
cells via regulating the AKT-mTOR and ERK-MAPK signaling pathways.
However, the present study was based on the hypothesis that RPS20
has dual functions as a ribosomal component and signal transducer.
Consequently, no experiments were conducted to investigate whether
RPS20 mediated direct activation of the ERK-MAPK and AKT-mTOR
signaling pathways. In future studies, the specific mechanisms by
which RPS20 affects the aforementioned pathways will be
investigated.
Acknowledgements
The authors are grateful to the Second Affiliated
Hospital of Nantong University for their support of the present
study.
Funding
The present study was supported by the Natural Science
Foundation of Jiangsu (grant no. BE2017682), Nantong Science and
Technology Bureau (grant no. MS22019009), Youth Project of Health
Commission of Nantong City (grant no. QN2022017), and Basic
Research and Social Minsheng Plan Project (grant no.
JC12022008).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
CS, ZC, YZ, WX and RP performed the experiments. CS
and BZ confirm the authenticity of all the raw data. JJ, YF and WZ
analyzed the data. CS and ZC wrote the manuscript. BZ designed this
study and polished the manuscript. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
Human studies were approved (approval no. 2021YL012)
by the Ethics Committee of The Second Affiliated Hospital of
Nantong University (Nantong, China) according to the Declaration of
Helsinki of 1964. Written informed consent was provided by all
participating patients. The animal experiments were approved
(approval no. S20210227-041) by the Animal Ethics Committee of
Nantong University (Nantong, China) and the experiments were
conducted according to the National Institutes of Health Guide for
the Care and Use of Laboratory Animals.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Miller KD, Nogueira L, Mariotto AB,
Rowland JH, Yabroff KR, Alfano CM, Jemal A, Kramer JL and Siegel
RL: Cancer treatment and survivorship statistics, 2019. CA Cancer J
Clin. 69:363–385. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fernández-Pello S, Hofmann F, Tahbaz R,
Marconi L, Lam TB, Albiges L, Bensalah K, Canfield SE, Dabestani S,
Giles RH, et al: A systematic review and meta-analysis comparing
the effectiveness and adverse effects of different systemic
treatments for non-clear cell renal cell carcinoma. Eur Urol.
71:426–436. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rhoades Smith KE and Bilen MA: A review of
papillary renal cell carcinoma and MET inhibitors. Kidney Cancer.
3:151–161. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen VJ, Hernandez-Meza G, Agrawal P,
Zhang CA, Xie L, Gong CL, Hoerner CR, Srinivas S, Oermann EK and
Fan AC: Time on therapy for at least three months correlates with
overall survival in metastatic renal cell carcinoma. Cancers
(Basel). 11:10002019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2018. CA Cancer J Clin. 68:7–30. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gao C, Guo X, Xue A, Ruan Y, Wang H and
Gao X: High intratumoral expression of eIF4A1 promotes
epithelial-to-mesenchymal transition and predicts unfavorable
prognosis in gastric cancer. Acta Biochim Biophys Sin (Shanghai).
52:310–319. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ruggero D and Pandolfi PP: Does the
ribosome translate cancer? Nat Rev Cancer. 3:179–192. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lindström MS: Emerging functions of
ribosomal proteins in gene-specific transcription and translation.
Biochem Biophys Res Commun. 379:167–170. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chu W, Presky DH, Swerlick RA and Burns
DK: Human ribosomal protein S20 cDNA sequence. Nucleic Acids Res.
21:16721993. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
O'Donohue MF, Choesmel V, Faubladier M,
Fichant G and Gleizes PE: Functional dichotomy of ribosomal
proteins during the synthesis of mammalian 40S ribosomal subunits.
J Cell Biol. 190:853–866. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tai LR, Chou CW, Wu JY, Kirby R and Lin A:
Late-assembly of human ribosomal protein S20 in the cytoplasm is
essential for the functioning of the small subunit ribosome. Exp
Cell Res. 319:2947–2953. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ko JR, Wu JY, Kirby R, Li IF and Lin A:
Mapping the essential structures of human ribosomal protein L7 for
nuclear entry, ribosome assembly and function. FEBS Lett.
580:3804–3810. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen IJ, Wang IA, Tai LR and Lin A: The
role of expansion segment of human ribosomal protein L35 in nuclear
entry, translation activity, and endoplasmic reticulum docking.
Biochem Cell Biol. 86:271–277. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Schmidt C, Lipsius E and Kruppa J: Nuclear
and nucleolar targeting of human ribosomal protein S6. Mol Biol
Cell. 6:1875–1885. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shu-Nu C, Lin CH and Lin A: An acidic
amino acid cluster regulates the nucleolar localization and
ribosome assembly of human ribosomal protein L22. FEBS Lett.
484:22–28. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Da Costa L, Tchernia G, Gascard P, Lo A,
Meerpohl J, Niemeyer C, Chasis JA, Fixler J and Mohandas N:
Nucleolar localization of RPS19 protein in normal cells and
mislocalization due to mutations in the nucleolar localization
signals in 2 Diamond-Blackfan anemia patients: Potential insights
into pathophysiology. Blood. 101:5039–5045. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Antoine M, Reimers K, Wirz W, Gressner AM,
Müller R and Kiefer P: Identification of an unconventional nuclear
localization signal in human ribosomal protein S2. Biochem Biophys
Res Commun. 335:146–153. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ferreira-Cerca S, Pöll G, Gleizes PE,
Tschochner H and Milkereit P: Roles of eukaryotic ribosomal
proteins in maturation and transport of pre-18S rRNA and ribosome
function. Mol Cell. 20:263–275. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rouquette J, Choesmel V and Gleizes PE:
Nuclear export and cytoplasmic processing of precursors to the 40S
ribosomal subunits in mammalian cells. EMBO J. 24:2862–2872. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang Y and Lu H: Signaling to p53:
Ribosomal proteins find their way. Cancer Cell. 16:369–377. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Boulon S, Westman BJ, Hutten S, Boisvert
FM and Lamond AI: The nucleolus under stress. Mol Cell. 40:216–227.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang Y, Wolf GW, Bhat K, Jin A, Allio T,
Burkhart WA and Xiong Y: Ribosomal protein L11 negatively regulates
oncoprotein MDM2 and mediates a p53-dependent ribosomal-stress
checkpoint pathway. Mol Cell Biol. 23:8902–8912. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dai MS and Lu H: Inhibition of
MDM2-mediated p53 ubiquitination and degradation by ribosomal
protein L5. J Biol Chem. 279:44475–44482. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jin A, Itahana K, O'Keefe K and Zhang Y:
Inhibition of HDM2 and activation of p53 by ribosomal protein L23.
Mol Cell Biol. 24:7669–7680. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang Y, Wang J, Yuan Y, Zhang W, Guan W,
Wu Z, Jin C, Chen H, Zhang L, Yang X and He F: Negative regulation
of HDM2 to attenuate p53 degradation by ribosomal protein L26.
Nucleic Acids Res. 38:6544–6554. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen D, Zhang Z, Li M, Wang W, Li Y,
Rayburn ER, Hill DL, Wang H and Zhang R: Ribosomal protein S7 as a
novel modulator of p53-MDM2 interaction: Binding to MDM2,
stabilization of p53 protein, and activation of p53 function.
Oncogene. 26:5029–5037. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xiong X, Zhao Y, He H and Sun Y: Ribosomal
protein S27-like and S27 interplay with p53-MDM2 axis as a target,
a substrate and a regulator. Oncogene. 30:1798–1811. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sun XX, DeVine T, Challagundla KB and Dai
MS: Interplay between ribosomal protein S27a and MDM2 protein in
p53 activation in response to ribosomal stress. J Biol Chem.
286:22730–22741. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Frum R, Busby SA, Ramamoorthy M, Deb S,
Shabanowitz J, Hunt DF and Deb SP: HDM2-binding partners:
Interaction with translation elongation factor EF1alpha. J Proteome
Res. 6:1410–1417. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Krishnan R, Boddapati N and Mahalingam S:
Interplay between human nucleolar GNL1 and RPS20 is critical to
modulate cell proliferation. Sci Rep. 8:114212018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Rhodes DR, Kalyana-Sundaram S, Mahavisno
V, Varambally R, Yu J, Briggs BB, Barrette TR, Anstet MJ,
Kincead-Beal C, Kulkarni P, et al: Oncomine 3.0: Genes, pathways,
and networks in a collection of 18,000 cancer gene expression
profiles. Neoplasia. 9:166–180. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lánczky A, Nagy Á, Bottai G, Munkácsy G,
Szabó A, Santarpia L and Győrffy B: miRpower: A web-tool to
validate survival-associated miRNAs utilizing expression data from
2178 breast cancer patients. Breast Cancer Res Treat. 160:439–446.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tang Z, Li C, Kang B, Gao G, Li C and
Zhang Z: GEPIA: A web server for cancer and normal gene expression
profiling and interactive analyses. Nucleic Acids Res. 45:W98–W102.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zheng B, Mao JH, Qian L, Zhu H, Gu DH, Pan
XD, Yi F and Ji DM: Pre-clinical evaluation of AZD-2014, a novel
mTORC1/2 dual inhibitor, against renal cell carcinoma. Cancer Lett.
357:468–475. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wettersten HI, Aboud OA, Lara PN and Weiss
RH: Metabolic reprogramming in clear cell renal cell carcinoma. Nat
Rev Nephrol. 13:410–419. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Xie M, Ma T, Xue J, Ma H, Sun M, Zhang Z,
Liu M, Liu Y, Ju S, Wang Z and De W: The long intergenic
non-protein coding RNA 707 promotes proliferation and metastasis of
gastric cancer by interacting with mRNA stabilizing protein HuR.
Cancer Lett. 443:67–79. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Dai MS, Sears R and Lu H: Feedback
regulation of c-Myc by ribosomal protein L11. Cell Cycle.
6:2735–2741. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Khanna N, Sen S, Sharma H and Singh N: S29
ribosomal protein induces apoptosis in H520 cells and sensitizes
them to chemotherapy. Biochem Biophys Res Commun. 304:26–35. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Warner JR and McIntosh KB: How common are
extraribosomal functions of ribosomal proteins? Mol Cell. 34:3–11.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Daftuar L, Zhu Y, Jacq X and Prives C:
Ribosomal proteins RPL37, RPS15 and RPS20 regulate the
Mdm2-p53-MdmX network. PLoS One. 8:e686672013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Nieminen TT, O'Donohue MF, Wu Y, Lohi H,
Scherer SW, Paterson AD, Ellonen P, Abdel-Rahman WM, Valo S,
Mecklin JP, et al: Germline mutation of RPS20, encoding a ribosomal
protein, causes predisposition to hereditary nonpolyposis
colorectal carcinoma without DNA mismatch repair deficiency.
Gastroenterology. 147:595–598.e5. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Li CX, Chen J, Xu ZG, Yiu WK and Lin YT:
The expression and prognostic value of RNA binding proteins in
clear cell renal cell carcinoma. Transl Cancer Res Dec.
9:7415–7431. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Uekita T and Sakai R: Roles of CUB
domain-containing protein 1 signaling in cancer invasion and
metastasis. Cancer Sci. 102:1943–1948. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Friedl P, Locker J, Sahai E and Segall JE:
Classifying collective cancer cell invasion. Nat Cell Biol.
14:777–783. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Fan D, Liu Q, Wu F, Liu N, Qu H, Yuan Y,
Li Y, Gao H, Ge J, Xu Y, et al: Prognostic significance of
PI3K/AKT/mTOR signaling pathway members in clear cell renal cell
carcinoma. PeerJ. 8:e92612020. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Igaki T, Pagliarini RA and Xu T: Loss of
cell polarity drives tumor growth and invasion through JNK
activation in Drosophila. Curr Biol. 16:1139–1146. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Chen PN, Hsieh YS, Chiang CL, Chiou HL,
Yang SF and Chu SC: Silibinin inhibits invasion of oral cancer
cells by suppressing the MAPK pathway. J Dent Res. 85:220–225.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Reddy KB, Nabha SM and Atanaskova N: Role
of MAP kinase in tumor progression and invasion. Cancer Metastasis
Rev. 22:395–403. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Matsushima-Nishiwaki R, Toyoda H,
Takamatsu R, Yasuda E, Okuda S, Maeda A, Kaneoka Y, Yoshimi N,
Kumada T and Kozawa O: Heat shock protein 22 (HSPB8) reduces the
migra-tion of hepatocellular carcinoma cells through the
suppression of the phosphoinositide 3-kinase (PI3K)/AKT pathway.
Biochim Biophys Acta Mol Basis Dis. 1863:1629–1639. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Wang L, Fang Z, Gao P and Zheng J: GLUD1
suppresses renal tumorigenesis and development via inhibiting
PI3K/Akt/mTOR pathway. Front Oncol. 12:9755172022. View Article : Google Scholar : PubMed/NCBI
|