Introduction
Hepatocellular carcinoma (HCC) is one of the most
common malignant tumors worldwide and the third most common cause
of cancer-related mortality (1).
The prognosis of patients with HCC remains unsatisfactory, despite
the fact that various treatment strategies have been developed. HCC
is a highly aggressive tumor. Frequent intrahepatic and distant
metastases are the main reasons for the high recurrence rate and
the low survival rate of patients with liver cancer following
surgery (2). Therefore, in-depth
studies on the developmental mechanisms of HCC are urgently
required to identify safer and more effective therapeutic
strategies for patients with HCC in order to prevent tumor
recurrence and improve the survival rate of patients.
Normal cells rely on mitochondrial oxidative
phosphorylation to provide energy for growth and differentiation
under well-oxygenated conditions. In tumors and other proliferating
or developing cells, even in the presence of oxygen and fully
functioning mitochondria, the cells opt to undergo cellular
metabolism and acquire an energy supply in the form of glycolysis,
which leads to a marked increase in the rate of glucose uptake into
the cells and promotes the production of large amounts of lactic
acid. This phenomenon is known as the ‘Warburg’ effect or ‘aerobic
glycolysis’ (3).
In glycolysis, when glucose is metabolized to
lactate, only two ATPs are generated per glucose molecule, whereas
the oxidative phosphorylation of a glucose molecule following
complete oxidation can generate up to 36 ATPs for cellular energy
supply, and cancer cells tend to favor glycolysis. This appears to
be a ‘disadvantageous’ metabolic pattern, with the inefficient
production of ATP. Indeed, the preference of tumors for glycolysis
as their primary metabolic energy source represents an adaptive
response to the environmental growth constraints commonly
encountered in cancer development. Firstly, aerobic glycolysis
provides the free energy or co-factors (for example ATP, NADPH and
NADH) required for sustained cancer cell proliferation (4). Secondly, glycolysis metabolizes one
molecule of glucose 10–100-fold more rapidly than the complete
oxidation of one molecule of glucose in the mitochondria. These two
different strategies of metabolizing glucose produce comparable
amounts of ATP in any given time period (5). Therefore, a plausible explanation is
that glycolytic metabolism occurs more rapidly than oxidative
phosphorylation (OXPHOS) and ATP production is rapid enough to
compensate for the insufficient amount of ATP produced (6,7).
Thirdly, using glycolytic metabolism, tumor cells escape oxidative
stress and are protected from reactive oxygen species-induced
damage (8). In fact, the liver is
one of the largest metabolic organs and a center for
gluconeogenesis. Aerobic glycolysis affects the progression of HCC,
including the maintenance of cell proliferation, the induction of
immune escape, invasion and metastasis, and the promotion of
angiogenesis and tumor resistance (9,10).
RNA sequencing techniques and transcriptional
profiling have demonstrated that although the human genome is
universally transcribed, only ~2% of RNAs code for proteins
(11,12). Even if they do not code for
proteins, long non-coding RNAs (lncRNAs) are not functionless
‘noise sequences’. lncRNAs can be involved in biological
activities, including tumor growth, metastasis and metabolism
(13–15).
In recent years, lncRNAs have been found to play a
key role in the pathogenesis of cancers, including HCC.
Understanding the novel regulatory roles of lncRNAs in glucose
metabolism, particularly in HCC, could provide new insight into the
underlying mechanisms of cancer onset and progression, which could
ultimately lead to the identification of novel therapeutic targets.
Therefore, in the present review, the major factors affecting the
function of lncRNAs in HCC were briefly discussed and the
mechanisms through which lncRNAs regulate HCC progression through
glycolysis were summarized.
Overview of lncRNAs
High-throughput sequencing technologies and
computing platforms have demonstrated that ~75% of the human genome
can be transcribed into RNAs, of which 74% are encoded as
non-protein coding RNAs (ncRNAs). Although not translated, ncRNAs
play a regulatory role in the developmental and pathophysiological
stages of human cells. According to the length of RNAs, ncRNAs can
be mainly divided into small ncRNAs and lncRNAs. Among ncRNAs,
those with >200 nucleotides are classified as lncRNAs. They are
mostly transcribed by RNA polymerase II (pol II) and include
various types of intergenic transcripts, enhancer RNAs and positive
or antisense transcripts that overlap with other genes. lncRNAs can
be classified according to four factors as follows: Genomic
localization and influence on DNA sequence, functional mechanism
and targeting mechanisms (16).
lncRNAs have different subcellular localizations in cells, and the
unique subcellular localization is closely related to the function
of interacting molecules, post-transcriptional or
co-transcriptional regulatory modifications and external
stimuli.
Major factors affecting the function of
lncRNAs in HCC
In HCC, lncRNAs can play a key role as
oncogenes/tumor suppressor gene, and the main factors affecting the
function of lncRNAs are the following: Epigenetic modification,
selective splicing, transcriptional regulation, binding to RNAs and
the encoding of small peptides (Fig.
1).
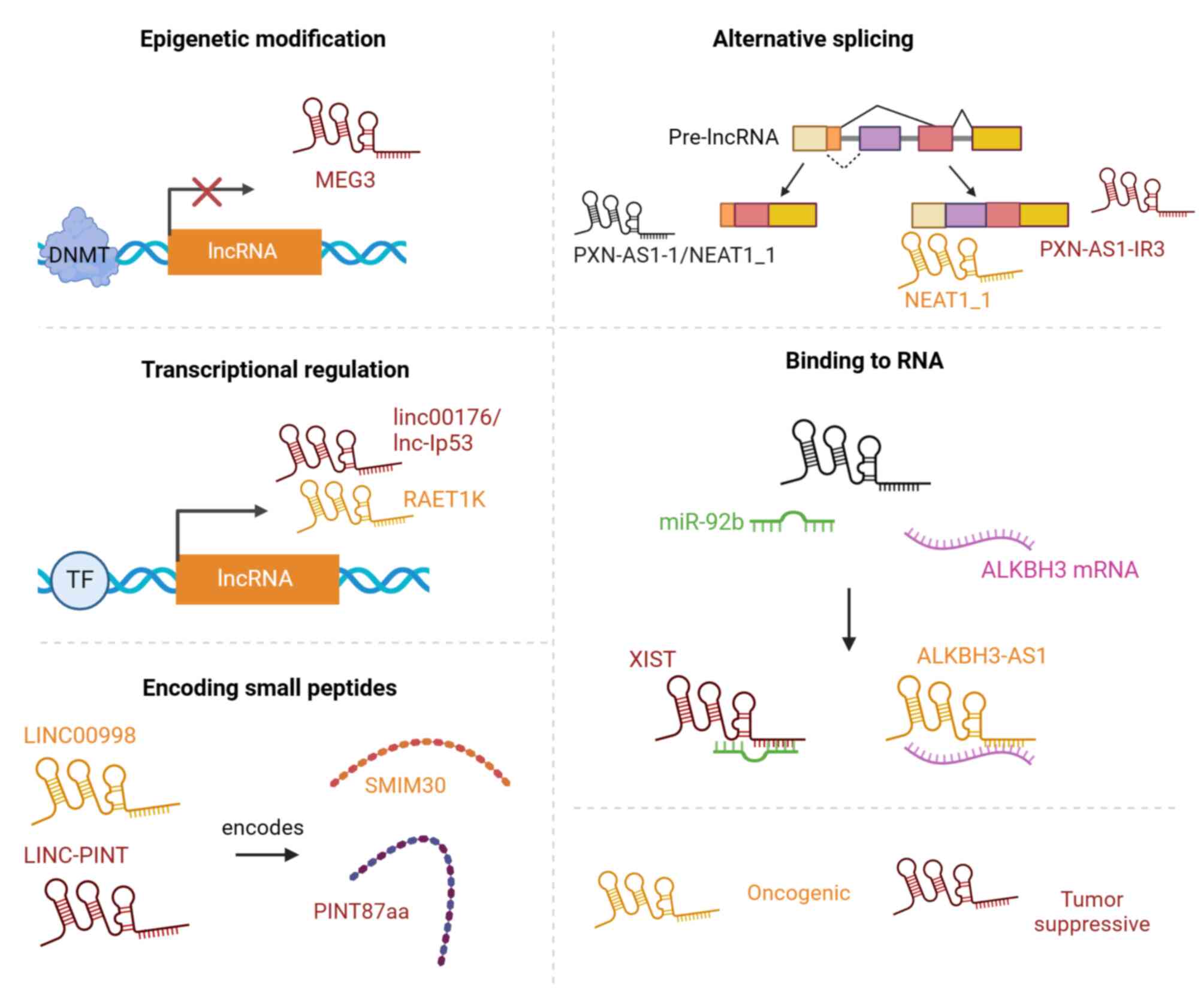 | Figure 1.Functional mechanisms of lncRNA in
HCC. DNMT, DNA Methyltransferase; MEG3, maternally expressed 3;
RAET1K, retinoic acid early transcript 1K; miR-92b, microRNA 92b;
ALKBH3: AlkB Homolog 3, alpha-ketoglutarate dependent dioxygenase;
ALKBH3-AS1, ALKBH3 antisense RNA 1; XIST, X inactive specific
transcript; LINC-PINT, long intergenic non-protein coding RNA,
P53-induced transcript; SMIM30, small integral membrane protein
30. |
Epigenetic modification
Common epigenetic modifications in HCC progression
include DNA methylation and histone modification (17). The regulation of epigenetic
modification determines the function of lncRNAs. lncRNAs can
recruit some chromatin remodeling complexes to mediate gene
silencing and thus play a role in promoting or suppressing HCC. For
example, linc00441 has been shown to recruit DNA methyltransferase
(DNMT)3A for methylation, inducing the silencing of its neighboring
gene, RB1, and thereby promoting the proliferation of HCC cells
(18). In addition, DNMT1 and DNMT3
induce the hypermethylation of the MEG3 promoter of the tumor
suppressor lncRNA, reducing MEG3 expression and leading to
apoptotic resistance and tumor growth in HCC cells (19). Of note, Zhou et al (20) reported that lncRNA ID2-AS1 inhibited
HCC tumor metastasis by blocking histone deacetylase 8.
Alternative splicing
Different forms of selective splicing of lncRNA
precursors (pre-lncRNAs) generate different isoforms of lncRNAs,
which may enable them to perform different functions in HCC. For
example, lncRNA PXN-AS1 can be spliced into several different
isoforms, among which the PXN-AS1-L isoform inhibits apoptosis in
myeloid leukemia in a PXN-dependent manner (21). However, another isoform
(PXN-AS1-IR3) promotes HCC metastasis by inducing transcriptional
activation of MYC (22). In
addition, the lncRNA NEAT1 can be alternatively spliced to produce
two isoforms, NEAT1_1 and NEAT1_2. It has been found that the low
expression of NEAT1_2 is significantly associated with the overall
survival of patients with HCC. Follow-up data suggest that NEAT1_2,
but not NEAT1_1, mediates mTORC1 signaling to control aerobic
glycolysis in HCC cells, contributing to the Warburg effect and HCC
development (23).
Transcriptional regulation
lncRNAs are subject to different transcriptional
regulations, which markedly affect the cancer- or
cancer-suppressive functions of lncRNAs. A number of tumor
suppressor genes or oncogene transcription factors, including p53,
hypoxia inducible factor-1α (HIF-1α) and myelocytomatosis (myc)
have been shown to induce the transcription of lncRNA genes. For
example, myc is a key proto-oncogene, and in HCC, lncRNA linc00176,
which is transcribed by myc, has been found to play a role in
promoting cell proliferation and survival by inhibiting cancer cell
cycle arrest and cell necrosis (24). Another study demonstrated that
HIF-1α binds to the promoter region of lncRNA RAET1K to activate
its transcription, thereby enhancing its cancer-promoting effect in
HCC (25). However, MEG3 has been
identified to function as a tumor suppressor gene in hepatoma cells
by interacting with tumor suppressor p53 protein to activate
p53-mediated transcriptional activity and affect the expression of
certain p53 target genes (26).
Binding to RNAs
The binding of lncRNAs to RNAs is another key factor
in determining their role in cancer. ALKBH3-AS1 is a carcinogenic
lncRNA, which directly binds ALKBH3 mRNA in HCC cells, upregulates
the expression level of ALKBH3, and promotes the proliferation of
cancer cells (27). In addition to
directly regulating mRNAs, lncRNAs also affect the expression of
their target genes by controlling microRNA (miRNA or miR)
expression. For example, it has been demonstrated that lncRNA XIST
binds miR-92b as a sponge in liver cancer tissues, preventing
miR-92b from binding to its downstream target Smad7 mRNA, thereby
inhibiting the proliferation and metastasis of HCC (28).
Encoding small peptides
Encoded small peptides play a crucial role in cancer
development and determine the functions of lncRNAs in cancer.
SMIM30, encoded by LINC00998 in HCC, has been found to be
associated with a low survival rate of patients with HCC and is
able to promote the development of HCC by inducing SRC/YES1
membrane anchoring and MAPK pathway activation (29). By contrast, LINC-PINT encodes a
PINT87aa micropeptide that binds to the transcription factor,
FOXM1, as a potential anticancer micropeptide, while promoting
cancer cell senescence (30). A
recent correlative study using ribosome analysis methods found that
a 99-aa peptide termed KRASIM encoded by lncRNA NCBP2-AS2 bound to
KRAS proteins to inhibit ERK signaling, thereby inhibiting the
growth of HCC cells (31).
Interaction between lncRNAs, glycolysis and
HCC progression
Aerobic glycolysis is strongly
associated with the progression of HCC
Aerobic glycolysis in HCC cells has been reported to
be closely related to their sustained proliferation, invasive
metastasis, the induction of stem cell activity and the generation
of therapeutic resistance. In the aerobic glycolytic metabolism of
HCC cells, an increase in the glycolytic flux of the cancer cells,
accompanied by an increase in intermediate metabolites, provides
sufficient energy and metabolites for the biosynthetic molecules
required for the sustained proliferation of the cancer cells. As
previously demonstrated, in xenograft tumors, the growth rate of
HCC tumors was reduced by ~50% by decreasing the aerobic glycolytic
enzyme, hexokinase 2 (HK2) (32).
The production of large amounts of lactate and H+ in
aerobic glycolysis leads to the acidification of the extracellular
environment, induces the transformation of cancer cell epithelial
cells into mesenchymal cells, and promotes the invasion and
metastasis of tumor cells. It has been found that the highly
metastatic HCC cell lines, MHCC97H and LM3, exhibit higher levels
of aerobic glycolysis compared with those with a lesser invasive
ability (33). Increased lactate
levels further enhance the stemness of tumor stem cells, and the
elevated lactylation of H3 histone effectively promotes the
oncogenicity of HCC stem cells (34). Furthermore, the
substantial production of lactic acid and hydrogen ions
(H+) during aerobic glycolysis leads to acidification of
the tumor microenvironment, promoting immunosuppression and
facilitating tumor immune evasion, as well as conferring resistance
to immunotherapy (35). Therefore,
targeting glycolytic metabolic pathways and controlling lactate
levels may be an effective strategy with which to prevent or
attenuate the progression of HCC.
lncRNAs involved in the regulation of
glycolysis in HCC
In the tumor microenvironment, some lncRNAs can
affect proliferation, metastasis and drug resistance in HCC by
promoting or inhibiting aerobic glycolysis in tumor cells. For
example, the knockdown of NPSR1-AS1 in HCC cells has been shown to
reduce their glycolytic metabolism and abolish their tumorigenic
potential. This suggests that NPSR1-AS1 has glycolysis-dependent
tumorigenic activity in HCC (36).
It has been demonstrated that overexpression of lncRNA
RP11-620J15.3 in HCC suggests that it functions as a competitive
endogenous RNA to upregulate glucose-6-phosphate isomerase via
sponge-binding to miR-326 and promoting aerobic glycolysis, which
in turn promotes HCC cell proliferation and metastasis (37). In addition to promoting tumor
progression, targeting changes in cancer cell metabolism is
currently providing new insight into tumor drug therapy. Tretinoin
is a widely known compound capable of inhibiting the development of
HCC. Zhang et al (38)
demonstrated that lncRNA MBNL1-AS1 reduced the sensitivity of HCC
cells to tretinoin by inhibiting miR-708-5p-mediated glycolysis.
This finding revealed an effective therapeutic target for the
treatment of HCC. The majority of the lncRNAs involved in
glycolysis in HCC are summarized in Table I and are discussed below.
 | Table I.Role of lncRNAs in the regulation of
glycolysis in HCC. |
Table I.
Role of lncRNAs in the regulation of
glycolysis in HCC.
| Regulation of
glycolysis | lncRNAs | Target | Function of
HCC | (Refs.) |
|---|
| Promote | NPSR1-AS1 | MAPK/ERK | Promoting
proliferation | (36) |
| Promote | RP11-620J15.3 | miR-326/GPI | Promoting
proliferation and metastasis, invasion | (37) |
| Promote | MBNL1-AS1 | miR-708-5p/HK2 | Promoting
resistance to tripterine and metastasis, invasion | (38) |
| Promote | TUG1 | miR-524-5p/SIX1;
miR-455-3p/AMPKβ2/HK2 | Promoting
proliferation and invasion | (39,40) |
| Promote | HOTAIR |
miR-130a-3p/HIF1α/mTOR/GLUT1 | Promoting
proliferation | (41,42) |
| Promote | MALAT1 | TCF7L2 | Promoting
proliferation | (43) |
| Promote | lncMMPA | miR-548
s/ALDH1A3 | Promoting
proliferation | (44) |
| Promote | Ftx | PPARγ | Promoting
proliferation and metastasis, invasion | (45) |
| Promote | SNHG6 | BOP1 | Promoting
proliferation | (46) |
| Promote | FTO-IT1 |
GLUT1/PKM2/c-myc | Promoting
proliferation | (47) |
| Promote | SOX2OT | PKM2 | Promoting
metastasis and invasion | (48) |
| Promote | miR4458HG | IGF2BP2 | Promoting
proliferation | (49) |
| Promote | NR2F1-AS1 | miR-140/HK2 | Promoting
invasion | (50) |
| Promote | PANTR1 | miR-587/BCL2A1 | Promoting
proliferation and invasion | (51) |
| Promote | SLC2A1-AS1 |
STAT3/FOXM1/GLUT1 | Promoting
proliferation and metasis | (52) |
| Promote | FIRRE | CREB/PFKFB4 | Promoting
proliferation | (53) |
| Promote | LINC01572 |
miR-195-5p/PFKFB4/PI3K-AKT | Promoting
proliferation and metastasis, invasion | (54) |
| Promote | ZFPM2-AS1 | miR-18b-5p/PKM | Promoting
proliferation, Metastasis, cancer stem macrophage polarization and
infiltration | (55) |
| Promote | SNHG1 | miR-326/PKM2 | Promoting
proliferation | (56) |
| Promote | CERS6-AS1 |
miR-30b-3p/MDM2/p53 | Promoting
proliferation and invasion | (57) |
| Promote | MNX1-AS1 | PKM2 | Promoting
proliferation | (58) |
| Promote | UPK1A-AS1 | HIF-1α | Promoting
proliferation | (59) |
| Inhibit | NEAT1 | mTORC1 | Inhibiting
proliferation | (23) |
| Inhibit | LINC00659 | FUS/SLC10A1 | Inhibiting
proliferation and metastasis | (60) |
| Inhibit | LINC01554 | PKM2, Akt/mTOR | Inhibiting
proliferation | (61) |
| Inhibit | WFDC21P | Nur77 | Inhibiting
proliferation and metastasis | (62) |
| Inhibit | NONHSAT024276 |
PTBP1/(PKM1/PKM2) | Inhibiting
proliferation and metastasis | (63) |
Glycolysis-promoting lncRNAs
Taurine upregulated gene 1 (TUG1)
TUG1 has been found to function as an oncogenic
lncRNA that is abnormally upregulated in the majority of cancers,
including HCC tissues (64). Its
overexpression has been found to be associated with the promotion
of glycolysis and has been studied in HCC. Partially, TUG1 mediates
its biological function by segregating with miRNAs. The
overexpression of TUG1 has been shown to be significantly
associated with HK2, and luciferase reporter gene-based assays have
revealed that the TUG1/miR-455-3p/AMPKβ2 axis affects HCC cell
growth, metastasis and glycolysis through the regulation of HK2
(40). SIX1 is a therapeutic target
in HCC and directly regulates lactate levels in cancer cells, which
in turn affects tumor cell proliferation, apoptosis and metastasis
(65). Lu et al (39) found that the overexpression of TUG1
impaired the miR-524-5p mimic-mediated inhibition of SIX1
expression, and suppressed glucose uptake, LDHA activity, lactate
levels and ATP levels. This suggests that the TCG1/miR-524-5p/SIX1
axis plays a key role in glycolysis, invasion and metastasis in
HCC.
HOX transcriptional antisense
intergenic RNA (HOTAIR)
HOTAIR is a new class of oncogenic lncRNA often
involved in regulating chromatin remodeling and epigenetic changes
(66). Previous research has
indicated that HOTAIR is highly expressed in a variety of malignant
tumors and is involved in cell proliferation, metastasis, DNA
repair and metabolism (67). As
previously demonstrated in HCC, HOTAIR promotes glycolysis through
the upregulation of glucose transporter (GLUT)1 and the activation
of the mTOR signaling pathway (42). In addition, has been revealed to
promote glycolysis in HCC under hypoxic conditions by targeting and
inhibiting miR-130a-3p (41).
Urothelial carcinoma associated 1
(UCA1)
The lncRNA UCA1 was originally identified in human
bladder cancer. It has an aberrant expression in embryogenesis and
in a wide range of cancerous tissues and cells, and plays a role in
tumor growth and glycolysis (68).
The expression of UCA1 has been found to be higher in HCC tissues
than in paracancerous tissues. Upwardly mobile coding protein 1
(UPF1) is an evolutionarily conserved and ubiquitously expressed
phosphoprotein that promotes cellular processes through G1/S
(69). RIP experiments have
revealed that UPF1 specifically binds to UCA1, and the
downregulation of UPF1 significantly upregulates the expression
level of UCA1 and effectively increases the rate of lactate
production in HCC (70).
Metastasis-associated lung
adenocarcinoma transcript 1 (MALAT1)
TCF7L2 has been identified to be an effector of the
Wnt signaling pathway and directly binds to several genes which
play a key role in the regulation of glucose metabolism. In
addition, genome-wide association studies have identified single
nucleotide polymorphisms in the TCF7L2 gene associated with
diabetes mellitus (71). It has
been previously indicated that MALAT1 plays a pivotal role in the
regulation of HCC cell proliferation, migration, and metastasis
(72). Furthermore, MALAT1 has been
found to regulate the expression of glycolytic genes in HCC by
increasing the translation of the transcription factor, TCF7L2,
which increases lactate and glucose fluxes (43). In addition, gluconeogenesis, a major
component of normal hepatocyte glucose metabolism, is negatively
regulated by MALAT1.
Glycolysis-inhibiting lncRNAs
LINC00659
Member 1 of 10 of the solute carrier family
(SLC10A1) encodes Na+-taurine bile acid cotransporter
peptide (73). The expression of
SLC10A1 in tumor tissues has been shown to be lower than that in
normal tissues, and it has been found to be associated with the
poor survival of patients with HCC (74). SLC10A1 has also been demonstrated to
inhibit glycolysis in HCC cells through rates of glucose
utilization, lactic acid production and extracellular acidification
(75). The transfection of
pcDNA3.1LINC00659 can upregulate the expression of SLC10A1 in HCC
cells. Similarly, seahorse experiments have revealed that an
increase in the oxygen consumption rate and a decrease in the
extracellular acidification rate of HepG2 and Huh7 cells induced by
the overexpression of LINC00659 can be partially reversed by
co-transfection with sh-SLC10A1. These results suggested that
LINC00659 inhibits the aerobic glycolysis of HCC cells by
modulating SLC10A1. Thus, LINC00659 is a potential therapeutic
target in HCC (60).
LINC01554
LINC01554 is a tumor suppressor lncRNA and inhibits
aerobic glycolysis in HCC (61).
The primary target of LINC01554 is pyruvate kinase M2 (PKM2), which
is the late rate-limiting enzyme of aerobic glycolysis. In
vitro ubiquitination experiments demonstrated enhanced PKM2
degradation mediated by ubiquitination in LINC01554 cells compared
with controls. In LINC01554-KO cells, the ubiquitination-mediated
degradation of PKM2 by LINC01554 was significantly attenuated. This
resulted in a reduced glucose consumption, lactate production and
pyruvate production, and in increased ATP levels. LINC01554 has the
characteristic of weakening the advantage of cancer cells in
acquiring high glycolysis; therefore, LINC01554 functions as a
tumor suppressor in HCC and may be used as a potential therapeutic
target in patients with HCC.
lncRNAs affect glycolysis by regulating
transporter proteins, enzymes and signaling pathways in HCC
Dysregulated lncRNAs in HCC are involved in an
altered cancer metabolism and are considered to play a key role in
regulating the dysregulation of transporter proteins, metabolic
enzymes and related signaling pathways in tumors that are dependent
on high rates of glycolysis (Fig.
2).
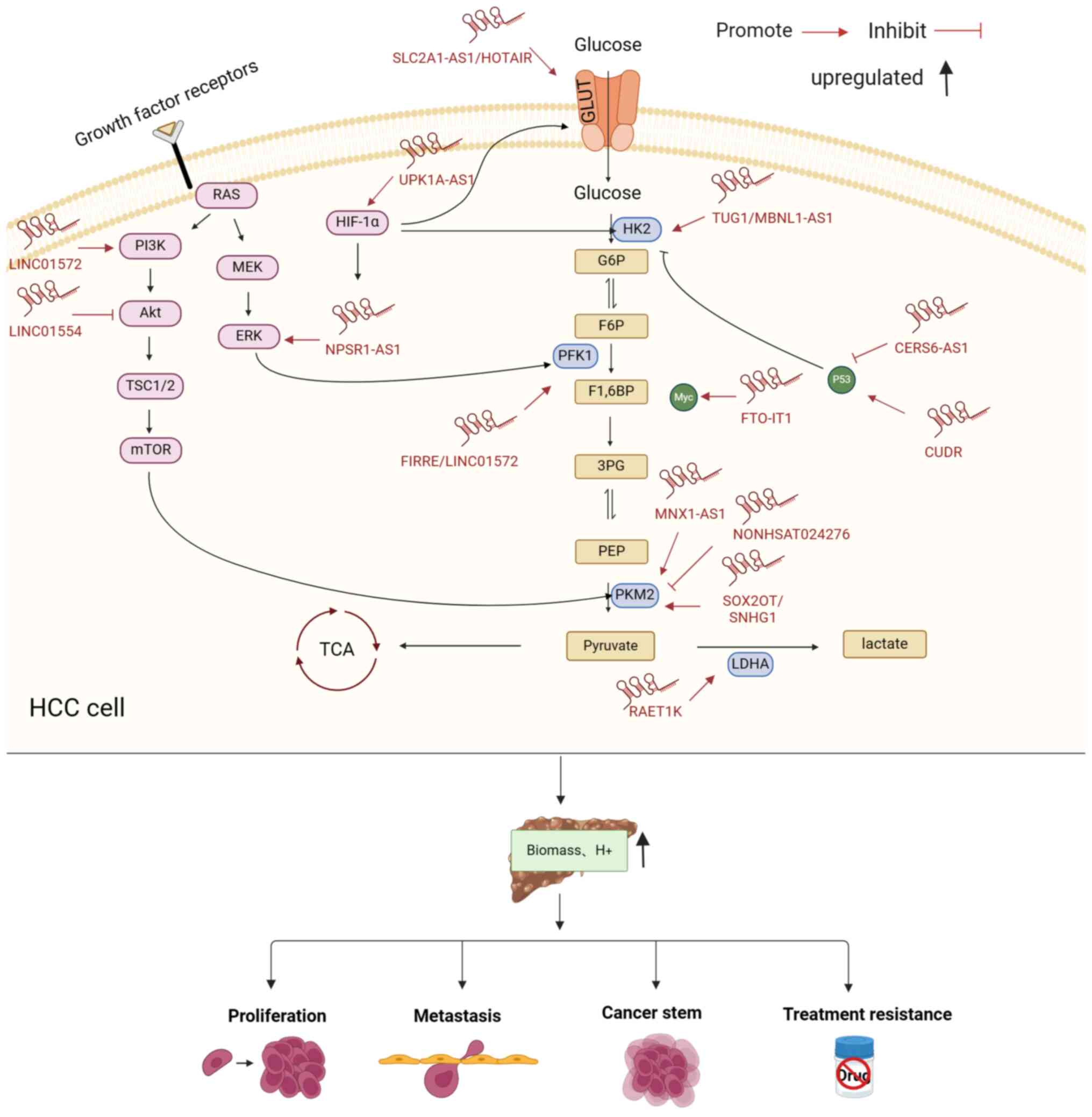 | Figure 2.lncRNAs affect glycolysis in HCC by
regulating glycolytic enzymes, transporter proteins and related
pathways. RAS, reliability availability and serviceability; PI3K,
phosphoinositide 3-kinase; Akt, protein kinase B; TSC1/2, tuberous
sclerosis complex 1/2; mTOR, mammalian target of rapamycin; MEK,
mitogen-activated extracellular signal-regulated kinase; ERK,
extracellular regulated protein kinases; HIF-1α, hypoxia inducible
factor-1α; GLUT1, glucose transporter type 1; HK2, hexokinase 2;
G6P, glucose-6-phosphatase G-6-pase; F6P, fructose 6-phosphate;
PFK1, phosphofructokinase-1; F1,6-BP, fructose 1,6-bisphosphate;
3PG, 3-phosphoglycerate; PEP, phosphoenolpyruvate; PKM2, pyruvate
kinase isozyme typeM2; LDHA, lactate dehydrogenase A; Myc, v-myc
avian myelocytomatosis viral oncogene homolog; ATP, adenosine
5′-triphosphate; H+, hydrogen; HOTAIR, HOX transcript
antisense RNA; UPK1A-AS1, UPK1A antisense RNA 1; NPSR1-AS1, NPSR1
antisense RNA 1; FIRRE, firre intergenic repeating RNA element;
TUG1, long non-coding RNA taurine-upregulated gene 1; FTO-IT1, FTO
intronic transcript 1; SNHG1, small nucleolar RNA host gene 1. |
lncRNAs regulate glycosylation-related
transporters and enzymes
GLUT
Often cancer cells exhibit a higher glucose uptake,
and GLUT proteins play a crucial role in the transmembrane
transport of glucose. GLUT proteins can be classified into three
isoforms. GLUT1, a facilitator of glucose transporter proteins, is
highly expressed in a variety of human cancers, including lung
cancer (76), colorectal cancer
(77), prostate cancer (78) and HCC (89). GLUT1 has also been shown to be
associated with a poor patient prognosis and plays a role in
glycolysis in cancer. It has been demonstrated that lncRNAs can
regulate glycolysis by affecting GLUT1 expression, thereby
influencing the development of HCC. For example, a previous study
found that SLC2A1-AS1-mediated the downregulation of GLUT1 and
significantly inhibited glycolysis in HCC (52). Another study demonstrated that
lncRNA FTO-IT1 promoted glycolysis and progression in HCC by
regulating FTO-mediated N6-methyladenosine modification
on GLUT1 (47). Furthermore, it has
been revealed that lncRNA HOTAIR promotes glycolysis by
upregulating GLUT1 and activating the mTOR signaling pathway
(42). GLUT has emerged as a target
for cancer therapy in recent years due to the dependence of tumor
cell growth on extracellular glucose (80).
HK2
HK catalyzes the first step in glycolytic
metabolism, catalyzing the formation of glucose-6-phosphate. It is
considered to be a key regulator of cellular energy metabolism and
contributes to the Warburg effect by promoting intracellular
glucose uptake (81–83). HK2 has often been reported to be
highly expressed in HCC cells and induces tumor development by
promoting glycolysis. The lncRNA TUG1 has been shown to be
significantly associated with HK2 overexpression and the poor
prognosis of patients with HCC. It has also been demonstrated that
TUG1 positively regulates HK2 expression by binding to miR-455-3p,
which promotes glycolysis in tumor cells, and accelerates tumor
growth and metastasis (40). In
addition, MBNL1-AS1 and NR2F1-AS1 have been reported to positively
regulate the expression level of HK2, which promotes aerobic
glycolysis in HCC cells, and contributes to cancer cell
proliferation, migration and resistance to therapeutic drugs
(38,50). Therefore, an in-depth study of HK2
may provide insight into the tumorigenesis and progression of HCC,
and may lead to the development of novel therapeutics.
Lactate dehydrogenase (LDH) A
LDHA is a member of the LDH family, which is the
rate-limiting enzyme for the interconversion of pyruvate and
lactate in the glycolytic pathway. LDHA is upregulated in a number
of types of cancer and is associated with the clinicopathological
features and prognosis of patients (84,85).
The lncRNA RAET1K has been found to be positively associated with
the expression of LDHA in cells and regulate its activity,
contributing to increased levels of glycolysis, thereby promoting
cell proliferation and invasion (25).
PKM2
PK functions as the final rate-limiting enzyme of
glycolysis, converting phosphoenolpyruvate to pyruvate. PKM2 is one
of the four isozymes of PK and plays a crucial role in cancer
development (86). lncRNAs can
participate in the regulation of aerobic glycolysis by affecting
the expression of PKM2, thereby influencing HCC progression. For
example, the lncRNA SOX2OT has been shown to promote PKM2-mediated
glycolytic activation by targeting the binding to miR-122-5p,
increasing its expression level and promoting the metastasis of HCC
cells (48). In addition, the
lncRNA SNHG1 has been revealed to function as a molecular sponge
for miR-326, isolating the interaction of miR-326 with PKM2, and
promoting PKM2 expression. The activation of PKM2 expression is one
of the mechanisms through which SNHG1 promotes glycolysis and HCC
cell proliferation (56).
NONHSAT024276, a potential oncogene for HCC, directly binds to
polypyrimidine bundle-binding protein 1 (PTBP1), increases the
ratio of M1 to M2 isoforms of PKM1/PKM2 and blocks
PTBP1/PKM-mediated glycolysis to inhibit cancer cell proliferation
and migration (63).
6-Phosphofructo-2-kinase/fructose-2,6-biphosphatase4 (PFKFB4)
PFKFB4, a member of the PFKFB family, which has been
identified to be a key regulator of glycolysis, controls the
synthesis and degradation of fructose-2,6-bisphosphate (F-2,6-BP)
(87). It has been recently
revealed that lncRNAs are involved in the regulation of PFKFB4
expression in tumor tissues and that they play a key role in tumor
glycolysis. The high expression of the lncRNA FIRRE has been shown
to be associated with malignant clinical features and with the poor
survival of patients with HCC. Mechanistically, FIRRE promotes HCC
cell proliferation and glycolysis by facilitating PFKFB4
transcription and expression, mainly through cAMP-responsive
element-binding protein (53).
Another study confirmed that LINC01572 is aberrantly upregulated in
HCC tissues, particularly in patients with type 2 diabetes.
Mechanistically, LINC01572 increases the level of PFKFB4 by
competitively inhibiting miR-195-5p, thereby enhancing glycolysis
and triggering HCC development (54).
lncRNAs regulate glycolysis-related
signaling pathways p53 signaling pathway
p53 (also known as TP53) is a well-known tumor
suppressor gene. It has been demonstrated that lncRNAs, as
functional components of the p53 pathway, may play a regulatory
role in this pathway. lncRNA CERS6-AS1 can sponge miR-30b-3p to
elevate MDM2, which promotes MDM2-mediated ubiquitin-dependent
degradation of the p53 oncogene, and facilitates glycolysis in HCC
cells (57). On the contrary, the
interaction between p53 and lncRNA can promote cancer by
influencing glycolytic enzymes. p53 forms a complex with the lncRNA
CUDR, which binds to the promoter region of PKM2 to enhance PKM2
expression, phosphorylation and polymer formation, and, ultimately,
p53 accelerates the growth of the HCC cell line, Hep3B, by
lengthening telomeres through a cascade of reactions that promotes
HCC development (88).
c-Myc signaling pathway
c-Myc is a transcription factor, mainly found in the
nucleus, that has been implicated in the development and
progression of cancer. c-Myc, as a major regulator of aerobic
glycolysis, can regulate aerobic glycolysis by directly activating
glycolytic enzymes (89). In
addition to a large number of coding genes, lncRNAs function as
downstream targets of c-Myc and participate in glycolysis in cancer
cells. lncRNA FTO-IT1 has been shown to enhance glycolysis in HCC
by inducing the stabilization of FTO mRNA, leading to the
overexpression of c-Myc. Moreover, c-Myc has been found to regulate
the expression of FTO-IT1 by binding to its promoter region under
low glucose conditions, forming a positive feedback loop between
c-Myc and FTO-IT1 (47). Wu et
al (58) also demonstrated that
MNX1-AS1, a c-Myc target gene, was upregulated in HCC, promoting
aerobic glycolysis and tumorigenesis.
HIF-1α signaling pathway
When tumor cells continue to proliferate and expand
to a certain limit, vascular hypoxia leads to the appearance of
HIF, and HIF-1α plays a key role as a member of the aerobic
glycolysis and lactate pathways (90). In hypoxic environments, NPSR1-AS1
induces the activation of the MAPK/ERK pathway in HCC cells, which
promotes the proliferation and glycolysis of HCC cells (36). The overexpression of lncRNA
UPK1A-AS1 has been reported to significantly increase the stability
of the HIF-1α ubiquitin-modified expression of upregulated
glycolysis-related genes, thereby promoting glycolysis levels in
HCC cells (59).
Phosphatidylinositol 3-kinase
(PI3K)/AKT/mTOR signaling pathway
The dysregulation of the PI3K/AKT/mTOR pathway is a
prevalent occurrence in the majority of human cancers. In HCC, this
signaling cascade plays a crucial role in facilitating glucose
metabolism, tumor metastasis, and resistance to drugs (91–93).
It has been demonstrated that lncRNA is a regulator of
PI3K/AKT/mTOR signaling in a variety of cancer types, and can
indirectly affect the expression of enzymes by regulating this
pathway. In HCC tissues and cell lines, the upregulation of
LINC01572 increases the expression of the glycolytic enzyme,
PFKFB4, by activating PI3K/AKT signaling, thereby enhancing
glycolysis and triggering HCC malignancy (54). mTOR is a serine/threonine kinase. It
has been previously shown that the upregulation of lncRNA HOTAIR
can induce the glycolysis of HCC cells by activating the mTOR
signaling pathway (42). By
contrast, LINC01554, a novel oncogene in HCC, has been identified
to inhibit the AKT/mTOR signaling pathway and eliminate aerobic
glycolysis in HCC cells, thereby suppressing tumor growth (61).
In summary, lncRNAs mediate changes in the
expression of glycolysis-associated transporter proteins, enzymes
and signaling pathways in HCCs, affecting the level of tumor
aerobic glycolysis, and thus cancer formation and progression.
Therefore, these glycolysis-associated lncRNAs have gradually
become key targets for cancer research, and the inhibition of these
lncRNAs is critical for controlling tumor development. Follow-up
studies should continue to explore the mechanisms of the lncRNA
regulation of glycolysis to provide new directions for cancer
treatment.
Conclusions and future perspectives
Although there have been major breakthroughs in the
study of malignant tumors, HCC remains a lethal disease. Due to the
high aggressiveness of HCC, this type of cancer is very likely to
metastasize, and the majority of cases are diagnosed in the middle
and late stages; thus, the option of surgical treatment is only
<40%, which is reflected in the low long-term survival rate of
patients with HCC (94). Therefore,
the influence of other molecular therapeutic targets on the
progression and mechanism of HCC may have a key impact on the
prevention and control of HCC and the long-term survival rate of
patients.
Cancer cells are metabolically active, and they can
alternate between glycolysis and mitochondrial OXPHOS in response
to nutritional stress caused by environmental changes (95). Therefore, blocking the glycolytic
pathway in tumor cells or inducing the transformation of cancer
cells from aerobic glycolytic to mitochondrial OXPHOS may lead to
novel approaches for the treatment of HCC. The present review
focused on the effects of lncRNAs on HCC progression, and their
specific functions and mechanisms in cancer metabolic pathways. In
terms of the mechanisms, lncRNAs significantly affect the process
of glucose metabolism mainly through glycolytic-related
transporters, metabolic enzymes or related signaling pathways, and
thus participate in the progression of HCC. lncRNAs are involved in
the regulation of glucose metabolism in tumor cells, which suggests
that lncRNAs, related glycolytic regulatory factors, may become
novel targets for cancer therapy.
In the current stage of HCC research, clinical
trials involving lncRNAs primarily focus on exosomal lncRNAs that
can serve as detectable biomarkers (96). lncRNAs can participate in tumor cell
metabolism through different mechanisms, which may have extensive
therapeutic significance and may provide new insight into the
treatment of HCC. Therefore, the role of lncRNAs in regulating
aerobic glycolysis should be carefully considered when considering
the development of future therapeutic drugs and methods. Further
studies on lncRNAs inhibitors may provide strategies with which to
block the progression of HCC. These may include: i) Targeting
lncRNAs capable of regulating glycolytic enzymes; ii) targeting
lncRNAs that regulate the glucose transporter GLUT to inhibit
glucose uptake; and iii) lncRNAs that target glycolysis-related
regulatory factors or signaling pathways. Therefore, the further
in-depth exploration of the mechanisms of lncRNAs in HCC glycolysis
may aid in the development of more effective therapeutic strategies
with which to inhibit tumor progression.
Acknowledgements
Not applicable.
Funding
The present study was supported in part by the Key R & D
program Natural Science Foundation of Guangxi (grant no.
AB19110007). The present study was also supported in part by the
Natural Science Foundation of Guangxi (grant no. 2017GXNSFAA198015)
and the Innovation Project of Guangxi Graduate Education (grant no.
YCSW2023215).
Availability of data and materials
Not applicable.
Authors' contributions
CO designed the present review. QioH wrote the
manuscript. ZL and QiqH was involved in article revision. XL, JX,
LH and LBH surveyed the literature and contributed to the
revisions. All authors have read and approved the final manuscript.
Data authentication is not applicable.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bertuccio P, Turati F, Carioli G,
Rodriguez T, La Vecchia C, Malvezzi M and Negri E: Global trends
and predictions in hepatocellular carcinoma mortality. J Hepatol.
67:302–309. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yang JD, Hainaut P, Gores GJ, Amadou A,
Plymoth A and Roberts LR: A global view of hepatocellular
carcinoma: Trends, risk, prevention and management. Nat Rev
Gastroenterol Hepatol. 16:589–604. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Vander Heiden MG, Cantley LC and Thompson
CB: Understanding the Warburg effect: The metabolic requirements of
cell proliferation. Science. 324:1029–1033. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lunt SY and Vander Heiden MG: Aerobic
glycolysis: meeting the metabolic requirements of cell
proliferation. Annu Rev Cell Dev Biol. 27:441–464. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Shestov AA, Liu X, Ser Z, Cluntun AA, Hung
YP, Huang L, Kim D, Le A, Yellen G, Albeck JG, et al: Quantitative
determinants of aerobic glycolysis identify flux through the enzyme
GAPDH as a limiting step. Elife. 3:e033422014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Guido C, Whitaker-Menezes D, Capparelli C,
Balliet R, Lin Z, Pestell RG, Howell A, Aquila S, Andò S,
Martinez-Outschoorn U, et al: Metabolic reprogramming of
cancer-associated fibroblasts by TGF-β drives tumor growth:
Connecting TGF-β signaling with ‘Warburg-like’ cancer metabolism
and L-lactate production. Cell Cycle. 11:3019–3035. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liberti MV and Locasale JW: The Warburg
effect: How does it benefit cancer cells? Trends Biochem Sci.
41:211–218. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sattler UGA and Mueller-Klieser W: The
anti-oxidant capacity of tumour glycolysis. Int J Radiat Biol.
85:963–971. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Beyoğlu D, Imbeaud S, Maurhofer O,
Bioulac-Sage P, Zucman-Rossi J, Dufour JF and Idle JR: Tissue
metabolomics of hepatocellular carcinoma: Tumor energy metabolism
and the role of transcriptomic classification. Hepatology.
58:229–238. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bustamante E and Pedersen PL: High aerobic
glycolysis of rat hepatoma cells in culture: Role of mitochondrial
hexokinase. Proc Natl Acad Sci USA. 74:3735–3739. 1977. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Djebali S, Davis CA, Merkel A, Dobin A,
Lassmann T, Mortazavi A, Tanzer A, Lagarde J, Lin W, Schlesinger F,
et al: Landscape of transcription in human cells. Nature.
489:101–108. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
ENCODE Project Consortium, Birney E,
Stamatoyannopoulos JA, Dutta A, Guigó R, Gingeras TR, Margulies EH,
Weng Z, Snyder M, Dermitzakis ET, et al: Identification and
analysis of functional elements in 1% of the human genome by the
ENCODE pilot project. Nature. 447:799–816. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Deng H, Zhang J, Shi J, Guo Z, He C, Ding
L, Tang JH and Hou Y: Role of long non-coding RNA in tumor drug
resistance. Tumour Biol. 37:11623–11631. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li J, Meng H, Bai Y and Wang K: Regulation
of lncRNA and its role in cancer metastasis. Oncol Res. 23:205–217.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shen XH, Qi P and Du X: Long non-coding
RNAs in cancer invasion and metastasis. Mod Pathol. 28:4–13. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Statello L, Guo CJ, Chen LL and Huarte M:
Gene regulation by long non-coding RNAs and its biological
functions. Nat Rev Mol Cell Biol. 22:96–118. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yuan SX, Zhang J, Xu QG, Yang Y and Zhou
WP: Long noncoding RNA, the methylation of genomic elements and
their emerging crosstalk in hepatocellular carcinoma. Cancer Lett.
379:239–244. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tang J, Xie Y, Xu X, Yin Y, Jiang R, Deng
L, Tan Z, Gangarapu V, Tang J and Sun B: Bidirectional
transcription of Linc00441 and RB1 via H3K27 modification-dependent
way promotes hepatocellular carcinoma. Cell Death Dis. 8:e26752017.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Braconi C, Kogure T, Valeri N, Huang N,
Nuovo G, Costinean S, Negrini M, Miotto E, Croce CM and Patel T:
microRNA-29 can regulate expression of the long non-coding RNA gene
MEG3 in hepatocellular cancer. Oncogene. 30:4750–4756. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhou Y, Huan L, Wu Y, Bao C, Chen B, Wang
L, Huang S, Liang L and He X: LncRNA ID2-AS1 suppresses tumor
metastasis by activating the HDAC8/ID2 pathway in hepatocellular
carcinoma. Cancer Lett. 469:399–409. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yuan JH, Liu XN, Wang TT, Pan W, Tao QF,
Zhou WP, Wang F and Sun SH: The MBNL3 splicing factor promotes
hepatocellular carcinoma by increasing PXN expression through the
alternative splicing of lncRNA-PXN-AS1. Nat Cell Biol. 19:820–832.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhou HZ, Li F, Cheng ST, Xu Y, Deng HJ, Gu
DY, Wang J, Chen WX, Zhou YJ, Yang ML, et al: DDX17-regulated
alternative splicing that produced an oncogenic isoform of PXN-AS1
to promote HCC metastasis. Hepatology. 75:847–865. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang H, Su X, Burley SK and Zheng XFS:
mTOR regulates aerobic glycolysis through NEAT1 and nuclear
paraspeckle-mediated mechanism in hepatocellular carcinoma.
Theranostics. 12:3518–3533. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tran DDH, Kessler C, Niehus SE, Mahnkopf
M, Koch A and Tamura T: Myc target gene, long intergenic noncoding
RNA, Linc00176 in hepatocellular carcinoma regulates cell cycle and
cell survival by titrating tumor suppressor microRNAs. Oncogene.
37:75–85. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhou Y, Huang Y, Hu K, Zhang Z, Yang J and
Wang Z: HIF1A activates the transcription of lncRNA RAET1K to
modulate hypoxia-induced glycolysis in hepatocellular carcinoma
cells via miR-100-5p. Cell Death Dis. 11:1762020. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhu J, Liu S, Ye F, Shen Y, Tie Y, Zhu J,
Wei L, Jin Y, Fu H, Wu Y and Zheng X: Long noncoding RNA MEG3
interacts with p53 protein and regulates partial p53 target genes
in hepatoma cells. PLoS One. 10:e1397902015.
|
|
27
|
Lu Q, Wang H, Lei X, Ma Q, Zhao J, Sun W,
Guo C, Huang D and Xu Q: LncRNA ALKBH3-AS1 enhances ALKBH3 mRNA
stability to promote hepatocellular carcinoma cell proliferation
and invasion. J Cell Mol Med. 26:5292–5302. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhuang LK, Yang YT, Ma X, Han B, Wang ZS,
Zhao QY, Wu LQ and Qu ZQ: MicroRNA-92b promotes hepatocellular
carcinoma progression by targeting Smad7 and is mediated by long
non-coding RNA XIST. Cell Death Dis. 7:e22032016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Pang Y, Liu Z, Han H, Wang B, Li W, Mao C
and Liu S: Peptide SMIM30 promotes HCC development by inducing
SRC/YES1 membrane anchoring and MAPK pathway activation. J Hepatol.
73:1155–1169. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Xiang X, Fu Y, Zhao K, Miao R, Zhang X, Ma
X, Liu C, Zhang N and Qu K: Cellular senescence in hepatocellular
carcinoma induced by a long non-coding RNA-encoded peptide PINT87aa
by blocking FOXM1-mediated PHB2. Theranostics. 11:4929–4944. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Xu W, Deng B, Lin P, Liu C, Li B, Huang Q,
Zhou H, Yang J and Qu L: Ribosome profiling analysis identified a
KRAS-interacting microprotein that represses oncogenic signaling in
hepatocellular carcinoma cells. Sci China Life Sci. 63:529–542.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li S, Li J, Dai W, Zhang Q, Feng J, Wu L,
Liu T, Yu Q, Xu S, Wang W, et al: Genistein suppresses aerobic
glycolysis and induces hepatocellular carcinoma cell death. Br J
Cancer. 117:1518–1528. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Li Y, Lu Z, Liang Z, Ji D, Zhang P, Liu Q,
Zheng X and Yao Y: Metastasis-associated in colon cancer-1 is
associated with poor prognosis in hepatocellular carcinoma, partly
by promoting proliferation through enhanced glucose metabolism. Mol
Med Rep. 12:426–434. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Pan L, Feng F, Wu J, Fan S, Han J, Wang S,
Yang L, Liu W, Wang C and Xu K: Demethylzeylasteral targets lactate
by inhibiting histone lactylation to suppress the tumorigenicity of
liver cancer stem cells. Pharmacol Res. 181:1062702022. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ganapathy-Kanniappan S: Linking tumor
glycolysis and immune evasion in cancer: Emerging concepts and
therapeutic opportunities. Biochim Biophys Acta Rev Cancer.
1868:212–220. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
He H, Chen T, Mo H, Chen S, Liu Q and Guo
C: Hypoxia-inducible long noncoding RNA NPSR1-AS1 promotes the
proliferation and glycolysis of hepatocellular carcinoma cells by
regulating the MAPK/ERK pathway. Biochem Biophys Res Commun.
533:886–892. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Liu C, Xu K, Liu J, He C, Liu P, Fu Q,
Zhang H and Qin T: LncRNA RP11-620J15.3 promotes HCC cell
proliferation and metastasis by targeting miR-326/GPI to enhance
glycolysis. Biol Direct. 18:152023. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhang H, Zhao L, Ren P and Sun X: LncRNA
MBNL1-AS1 knockdown increases the sensitivity of hepatocellular
carcinoma to tripterine by regulating miR-708-5p-mediated
glycolysis. Biotechnol Genet Eng Rev. 1–18. 2023.(Epub ahead of
print). View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lu L, Huang J, Mo J, Da X, Li Q, Fan M and
Lu H: Exosomal lncRNA TUG1 from cancer-associated fibroblasts
promotes liver cancer cell migration, invasion, and glycolysis by
regulating the miR-524-5p/SIX1 axis. Cell Mol Biol Lett. 27:172022.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Lin YH, Wu MH, Huang YH, Yeh CT, Cheng ML,
Chi HC, Tsai CY, Chung IH, Chen CY and Lin KH: Taurine up-regulated
gene 1 functions as a master regulator to coordinate glycolysis and
metastasis in hepatocellular carcinoma. Hepatology. 67:188–203.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hu M, Fu Q, Jing C, Zhang X, Qin T and Pan
Y: LncRNA HOTAIR knockdown inhibits glycolysis by regulating
miR-130a-3p/HIF1A in hepatocellular carcinoma under hypoxia. Biomed
Pharmacother. 125:1097032020. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wei S, Fan Q, Yang L, Zhang X, Ma Y, Zong
Z, Hua X, Su D, Sun H, Li H and Liu Z: Promotion of glycolysis by
HOTAIR through GLUT1 upregulation via mTOR signaling. Oncol Rep.
38:1902–1908. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Malakar P, Stein I, Saragovi A, Winkler R,
Stern-Ginossar N, Berger M, Pikarsky E and Karni R: Long Noncoding
RNA MALAT1 regulates cancer glucose metabolism by enhancing
mTOR-mediated translation of TCF7L2. Cancer Res. 79:2480–2493.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Xu M, Zhou C, Weng J, Chen Z, Zhou Q, Gao
J, Shi G, Ke A, Ren N, Sun H and Shen Y: Tumor associated
macrophages-derived exosomes facilitate hepatocellular carcinoma
malignance by transferring lncMMPA to tumor cells and activating
glycolysis pathway. J Exp Clin Cancer Res. 41:2532022. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Li X, Zhao Q, Qi J, Wang W, Zhang D, Li Z
and Qin C: lncRNA Ftx promotes aerobic glycolysis and tumor
progression through the PPARγ pathway in hepatocellular carcinoma.
Int J Oncol. 53:551–566. 2018.PubMed/NCBI
|
|
46
|
Chen K, Wang X, Wei B, Sun R, Wu C and
Yang HJ: LncRNA SNHG6 promotes glycolysis reprogramming in
hepatocellular carcinoma by stabilizing the BOP1 protein. Anim
Cells Syst (Seoul). 26:369–379. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wang F, Hu Y, Wang H, Hu P, Xiong H, Zeng
Z, Han S, Wang D, Wang J, Zhao Y, et al: LncRNA FTO-IT1 promotes
glycolysis and progression of hepatocellular carcinoma through
modulating FTO-mediated N6-methyladenosine modification on GLUT1
and PKM2. J Exp Clin Cancer Res. 42:2672023. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Liang Y, Zhang D, Zheng T, Yang G, Wang J,
Meng F, Liu Y, Zhang G, Zhang L, Han J, et al: lncRNA-SOX2OT
promotes hepatocellular carcinoma invasion and metastasis through
miR-122-5p-mediated activation of PKM2. Oncogenesis. 9:542020.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Ye Y, Wang M, Wang G, Mai Z, Zhou B, Han
Y, Zhuang J and Xia W: lncRNA miR4458HG modulates hepatocellular
carcinoma progression by activating m6A-dependent glycolysis and
promoting the polarization of tumor-associated macrophages. Cell
Mol Life Sci. 80:992023. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Li X, Li Y, Bai S, Zhang J, Liu Z and Yang
J: NR2F1-AS1/miR-140/HK2 axis regulates hypoxia-induced glycolysis
and migration in hepatocellular carcinoma. Cancer Manag Res.
13:427–437. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Ma X, Mao Z, Zhu J, Liu H and Chen F:
lncRNA PANTR1 upregulates BCL2A1 expression to promote
tumorigenesis and warburg effect of hepatocellular carcinoma
through restraining miR-587. J Immunol Res. 2021:17368192021.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Shang R, Wang M, Dai B, Du J, Wang J, Liu
Z, Qu S, Yang X, Liu J, Xia C, et al: Long noncoding RNA SLC2A1-AS1
regulates aerobic glycolysis and progression in hepatocellular
carcinoma via inhibiting the STAT3/FOXM1/GLUT1 pathway. Mol Oncol.
14:1381–1396. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Shen C, Ding L, Mo H, Liu R, Xu Q and Tu
K: Long noncoding RNA FIRRE contributes to the proliferation and
glycolysis of hepatocellular carcinoma cells by enhancing PFKFB4
expression. J Cancer. 12:4099–4108. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Lai S, Quan Z, Hao Y, Liu J, Wang Z, Dai
L, Dai H, He S and Tang B: Long non-coding RNA LINC01572 promotes
hepatocellular carcinoma progression via sponging miR-195-5p to
enhance PFKFB4-mediated glycolysis and PI3K/AKT activation. Front
Cell Dev Biol. 9:7830882021. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Ji W, Bai J and Ke Y: Exosomal ZFPM2-AS1
contributes to tumorigenesis, metastasis, stemness, macrophage
polarization, and infiltration in hepatocellular carcinoma through
PKM mediated glycolysis. Environ Toxicol. 38:1332–1346. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Wang Y, Yang F, Peng Q, Mei K, He H and
Yang Q: Long non-coding RNA SNHG1 activates glycolysis to promote
hepatocellular cancer progression through the miR-326/PKM2 axis. J
Gene Med. 24:e34402022. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Xu B, Wei Y, Liu F, Li L, Zhou S, Peng Y
and Li B: Long noncoding RNA CERS6-AS1 modulates glucose metabolism
and tumor progression in hepatocellular carcinoma by promoting the
MDM2/p53 signaling pathway. Cell Death Discov. 8:3482022.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Wu Y, Wang Y, Yao H, Li H, Meng F, Li Q,
Lin X and Liu L: MNX1-AS1, a c-Myc induced lncRNA, promotes the
Warburg effect by regulating PKM2 nuclear translocation. J Exp Clin
Cancer Res. 41:3372022. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Zhang D, Zou X, Song Y and Wu D: Long
non-coding RNA UPK1A-AS1 promotes glycolysis in hepatocellular
carcinoma cells via stabilization of HIF-1α. Nan Fang Yi Ke Da Xue
Xue Bao. 41:193–199. 2021.(In Chinese). PubMed/NCBI
|
|
60
|
Chen B, Xu X, Wu W, Zheng K and Yu Y:
LINC00659 inhibits hepatocellular carcinoma malignant progression
by blocking aerobic glycolysis through FUS recruitment and SLC10A1
modulation. Anal Cell Pathol (Amst). 2023:58529632023.PubMed/NCBI
|
|
61
|
Zheng YL, Li L, Jia YX, Zhang BZ, Li JC,
Zhu YH, Li MQ, He JZ, Zeng TT, Ban XJ, et al: LINC01554-mediated
glucose metabolism reprogramming suppresses tumorigenicity in
hepatocellular carcinoma via downregulating PKM2 expression and
inhibiting Akt/mTOR signaling pathway. Theranostics. 9:796–810.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Guan YF, Huang QL, Ai YL, Chen QT, Zhao
WX, Wang XM, Wu Q and Chen HZ: Nur77-activated lncRNA WFDC21P
attenuates hepatocarcinogenesis via modulating glycolysis.
Oncogene. 39:2408–2423. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Li Y, Chen X, Huang H, Liao L, Chong H, Li
G, Yuan T, Lu W, Deng S and Huang Q: A feedback loop between
NONHSAT024276 and PTBP1 inhibits tumor progression and glycolysis
in HCC by increasing the PKM1/PKM2 ratio. Cancer Sci.
114:1519–1540. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Huang MD, Chen WM, Qi FZ, Sun M, Xu TP, Ma
P and Shu YQ: Long non-coding RNA TUG1 is up-regulated in
hepatocellular carcinoma and promotes cell growth and apoptosis by
epigenetically silencing of KLF2. Mol Cancer. 14:1652015.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Chen K, Wei H, Pan J, Chen Z, Pan D, Gao
T, Huang J, Huang M, Ou M and Zhong W: Six1 is negatively
correlated with poor prognosis and reduces 5-fluorouracil
sensitivity via attenuating the stemness of hepatocellular
carcinoma cells. Eur J Pharmacol. 861:1725992019. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Raju G, Pavitra E, Bandaru SS, Varaprasad
GL, Nagaraju GP, Malla RR, Huh YS and Han YK: HOTAIR: A potential
metastatic, drug-resistant and prognostic regulator of breast
cancer. Mol Cancer. 22:652023. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Zhang J, Zhang P, Wang L, Piao HL and Ma
L: Long non-coding RNA HOTAIR in carcinogenesis and metastasis.
Acta Biochim Biophys Sin (Shanghai). 46:1–5. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Li T, Sun X and Jiang X: UCA1 involved in
the metformin-regulated bladder cancer cell proliferation and
glycolysis. Tumour Biol. 39:10104283177108232017. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Lou CH, Shao A, Shum EY, Espinoza JL,
Huang L, Karam R and Wilkinson MF: Posttranscriptional control of
the stem cell and neurogenic programs by the nonsense-mediated RNA
decay pathway. Cell Rep. 6:748–764. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Zhou Y, Li Y, Wang N, Li X, Zheng J and Ge
L: UPF1 inhibits the hepatocellular carcinoma progression by
targeting long non-coding RNA UCA1. Sci Rep. 9:66522019. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Grant SFA, Thorleifsson G, Reynisdottir I,
Benediktsson R, Manolescu A, Sainz J, Helgason A, Stefansson H,
Emilsson V, Helgadottir A, et al: Variant of transcription factor
7-like 2 (TCF7L2) gene confers risk of type 2 diabetes. Nat Genet.
38:320–323. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
72
|
Li GZ, Meng GX, Pan GQ, Zhang X, Yan LJ,
Li RZ, Ding ZN, Tan SY, Wang DX, Tian BW, et al: MALAT1/mir-1-3p
mediated BRF2 expression promotes HCC progression via inhibiting
the LKB1/AMPK signaling pathway. Cancer Cell Int. 23:1882023.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Döring B, Lütteke T, Geyer J and Petzinger
E: The SLC10 carrier family: Transport functions and molecular
structure. Curr Top Membr. 70:105–168. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Tran QH, Nguyen VG, Tran CM and Nguyen MN:
Down-regulation of solute carrier family 10 member 1 is associated
with early recurrence and poorer prognosis of hepatocellular
carcinoma. Heliyon. 7:e064632021. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Lu C, Fang S, Weng Q, Lv X, Meng M, Zhu J,
Zheng L, Hu Y, Gao Y, Wu X, et al: Integrated analysis reveals
critical glycolytic regulators in hepatocellular carcinoma. Cell
Commun Signal. 18:972020. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Zhang B, Xie Z and Li B: The
clinicopathologic impacts and prognostic significance of GLUT1
expression in patients with lung cancer: A meta-analysis. Gene.
689:76–83. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Kuai XY, Lei ZY, Liu XS and Shao XY: The
interaction of GLUT1 and FOXM1 leads to a poor prognosis in
colorectal cancer. Anticancer Agents Med Chem. 20:941–950. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Xiao H, Wang J, Yan W, Cui Y, Chen Z, Gao
X, Wen X and Chen J: GLUT1 regulates cell glycolysis and
proliferation in prostate cancer. Prostate. 78:86–94. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Sun HW, Yu XJ, Wu WC, Chen J, Shi M, Zheng
L and Xu J: GLUT1 and ASCT2 as predictors for prognosis of
hepatocellular carcinoma. PLoS One. 11:e01689072016. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Barbosa AM and Martel F: Targeting glucose
transporters for breast cancer therapy: The effect of natural and
synthetic compounds. Cancers (Basel). 12:1542020. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
DeWaal D, Nogueira V, Terry AR, Patra KC,
Jeon SM, Guzman G, Au J, Long CP, Antoniewicz MR and Hay N:
Hexokinase-2 depletion inhibits glycolysis and induces oxidative
phosphorylation in hepatocellular carcinoma and sensitizes to
metformin. Nat Commun. 9:4462018. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Wu Q, Wang SP, Sun XX, Tao YF, Yuan XQ,
Chen QM, Dai L, Li CL, Zhang JY and Yang AL: HuaChanSu suppresses
tumor growth and interferes with glucose metabolism in
hepatocellular carcinoma cells by restraining Hexokinase-2. Int J
Biochem Cell Biol. 142:1061232022. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Ros S and Schulze A: Glycolysis back in
the limelight: Systemic targeting of HK2 blocks tumor growth.
Cancer Discov. 3:1105–1107. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Zhang R, Su J, Xue SL, Yang H, Ju LL, Ji
Y, Wu KH, Zhang YW, Zhang YX, Hu JF and Yu M: HPV E6/p53 mediated
down-regulation of miR-34a inhibits Warburg effect through
targeting LDHA in cervical cancer. Am J Cancer Res. 6:312–320.
2016.PubMed/NCBI
|
|
85
|
Zhang K, Zhang T, Yang Y, Tu W, Huang H,
Wang Y, Chen Y, Pan K and Chen Z:
N6-methyladenosine-mediated LDHA induction potentiates
chemoresistance of colorectal cancer cells through metabolic
reprogramming. Theranostics. 12:4802–4817. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Christofk HR, Vander Heiden MG, Harris MH,
Ramanathan A, Gerszten RE, Wei R, Fleming MD, Schreiber SL and
Cantley LC: The M2 splice isoform of pyruvate kinase is important
for cancer metabolism and tumour growth. Nature. 452:230–233. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Kotowski K, Rosik J, Machaj F, Supplitt S,
Wiczew D, Jabłońska K, Wiechec E, Ghavami S and Dzięgiel P: Role of
PFKFB3 and PFKFB4 in cancer: Genetic basis, impact on disease
development/progression, and potential as therapeutic targets.
Cancers (Basel). 13:9092021. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Wu M, An J, Zheng Q, Xin X, Lin Z, Li X,
Li H and Lu D: Double mutant P53 (N340Q/L344R) promotes
hepatocarcinogenesis through upregulation of Pim1 mediated by PKM2
and LncRNA CUDR. Oncotarget. 7:66525–66539. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Tang J, Yan T, Bao Y, Shen C, Yu C, Zhu X,
Tian X, Guo F, Liang Q, Liu Q, et al: LncRNA GLCC1 promotes
colorectal carcinogenesis and glucose metabolism by stabilizing
c-Myc. Nat Commun. 10:34992019. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Yu Z, Wang Y, Deng J, Liu D, Zhang L, Shao
H, Wang Z, Zhu W, Zhao C and Ke Q: Long non-coding RNA COL4A2-AS1
facilitates cell proliferation and glycolysis of colorectal cancer
cells via miR-20b-5p/hypoxia inducible factor 1 alpha subunit axis.
Bioengineered. 12:6251–6263. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Badoiu SC, Greabu M, Miricescu D,
Stanescu-Spinu II, Ilinca R, Balan DG, Balcangiu-Stroescu AE, Mihai
DA, Vacaroiu IA, Stefani C and Jinga V: PI3K/AKT/mTOR dysregulation
and reprogramming metabolic pathways in renal cancer: Crosstalk
with the VHL/HIF axis. Int J Mol Sci. 24:83912023. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Zhu YJ, Zheng B, Wang HY and Chen L: New
knowledge of the mechanisms of sorafenib resistance in liver
cancer. Acta Pharmacol Sin. 38:614–622. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Zhang H, Wang Q, Liu J and Cao H:
Inhibition of the PI3K/Akt signaling pathway reverses
sorafenib-derived chemo-resistance in hepatocellular carcinoma.
Oncol Lett. 15:9377–9384. 2018.PubMed/NCBI
|
|
94
|
Li J, Xing J, Yang Y, Liu J, Wang W, Xia
Y, Yan Z, Wang K, Wu D, Wu L, et al: Adjuvant
131I-metuximab for hepatocellular carcinoma after liver
resection: A randomised, controlled, multicentre, open-label, phase
2 trial. Lancet Gastroenterol Hepatol. 5:548–560. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Elgendy M, Cirò M, Hosseini A, Weiszmann
J, Mazzarella L, Ferrari E, Cazzoli R, Curigliano G, DeCensi A,
Bonanni B, et al: Combination of hypoglycemia and metformin impairs
tumor metabolic plasticity and growth by modulating the
PP2A-GSK3β-MCL-1 axis. Cancer Cell. 35:798–815. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Sasaki R, Kanda T, Yokosuka O, Kato N,
Matsuoka S and Moriyama M: Exosomes and hepatocellular carcinoma:
From bench to bedside. Int J Mol Sci. 20:14062019. View Article : Google Scholar : PubMed/NCBI
|














