Introduction
To facilitate next generation sequencing techniques,
the discovery of pervasive transcription in the human genome was
one of the major advances of genome research in the last decade
(1,2).
The vast majority (~98%) of the mammalian genome is pervasively
transcribed into countless and multifunctional forms of RNA
molecules termed non-coding RNAs (ncRNAs) (3,4). Based on
the length of the functional transcripts, human ncRNAs have been
largely classified into two classes: Small (20–30 nt.) and long
(>200 nt.) non-coding transcripts. The small ncRNA class
includes housekeeping and regulatory RNAs (tRNAs and rRNAs),
microRNAs (miRNAs), as well as other small RNAs.
The long ncRNA (lncRNA) class includes an ambiguous,
functionally distinct class of RNAs that encompasses thousands of
lncRNA molecules with >15,000 encoded transcripts (5,6). Certain
molecules have been reported to be similar to protein-coding genes
with respect to splicing and polyadenylation. As a result of
relatively high functional diversity, confusing and overlapping
terminology has been used to name and classify lncRNAs (7). Various attributes, including transcript
length, association with annotated protein-coding gene or other DNA
elements/repeats, association with a biochemical pathway or
stability, or sequence and structure conservation have been used
for classification. Accumulating evidence reveals that lncRNAs are
important in various biologic processes, including gene regulation,
chromatin modification, immune surveillance, differentiation, cell
cycle control and apoptosis, or act as nuclear architecture,
subnuclear compartments, or enhancers and promoters in RNA
processing (8–10).
The current study evaluates lncRNA, growth arrest
specific 5 (GAS5). It is a recently identified tumor
suppressor and displays reduced expression levels in various types
of cancer, such as breast, prostate, lung and colorectal cancer. As
GAS5 is involved in the regulation of cell proliferation,
growth arrest and apoptosis (11), its
low-expression pattern confers the elevated capacity of tumor cells
for proliferation (12). GAS5,
among others, was previously shown to be detectable in secreted
exosomes (13). The existence of
lncRNAs in exosomes may be of relevance in cancer, as exosomes are
important mediators of intercellular communication, which regulates
a diverse range of biologic processes (14), and their pathophysiologic roles in
disease, including cancer are increasingly being recognized. As a
promoter of apoptosis, exosomal GAS5 may be involved in the
communication of dying tumor cells in their local microenvironment.
The aim of the current study was to determine whether exosomal
expression levels of GAS5 are adjusted during apoptosis
induction, which is triggered by different mechanisms, in cultured
breast cancer cells.
Materials and methods
Cell lines and cell culture
Two breast cancer cell lines (MCF-7 and MDA-MB-231)
were used in the present study and purchased from the American Type
Culture Collection (ATCC; Manassas, VA, USA]. The MCF-7 cell line
represents hormone-responsive human-invasive breast adenocarcinoma
with receptor positivity resulting in a luminal A subtype. The
MDA-MB-231 cells were of basal subtype, as these cells do not
express hormone receptors and possess the mutant p53 protein
(15). Cells were grown in Dulbecco's
modified Eagle's medium (Biochrom GmbH, Berlin, Germany), including
exosome-free serum, in standard culture conditions. To obtain
exosome-free serum, fetal calf serum (Biochrom GmbH) was
centrifuged at 120,000 × g for 2 h. Taxol and bleomycin (AppliChem
GmbH, Darmstadt, Germany) served as apoptosis-inducer anticancer
compounds. Taxol is an anti-mitotic that interferes with normal
breakdown of microtubules during cell division and induces cell
death. Bleomycin induces apoptotic cell death via DNA strand
breaks.
Evaluation of cytotoxicity
Taxol- and bleomycin-induced cytotoxicity were
evaluated using real-time cellular analysis in the microelectronic
biosensor-based iCELLigence instrument (ACEA Biosciences, San
Diego, CA, USA). In this system, numeric cell changes affect the
local ionic environment at the electrode/solution interface, which
leads to an increase or decrease in electrode impedance. Data are
recorded as impedance change and expressed as the cell index (CI),
as a mean of cell proliferation. For that analysis, 25,000 cells
were plated and Taxol (2 and 10 nm) or bleomycin (2 and 10 nm/ml)
(AppliChem GmbH) was added after 24 h; the cells were monitored for
an additional 24 h. Untreated cells were used as control cells.
Measurement of apoptosis
For a quantitative analysis of apoptosis,
cytoplasmic levels of mono- and oligonucleosomes released into the
cytoplasm during apoptosis prior to membrane breakdown were
measured using the Cell Death Detection ELISA kit (Roche
Diagnostics GmbH, Mannheim, Germany). Detection of nucleosomal
fragments as a result of DNA degradation has widely served as a
hallmark of apoptotic cell death (16). The assay is based on an immunoassay
using specific monoclonal antibodies against histones and DNA,
allowing the determination of nucleosomes in the cytoplasmic
fraction of cell lysates. In the present study, cells in culture
suspension were removed by discarding the medium and only intact
cells were used. For this assay, 150,000 cells were used and
analyzed as previously described (17).
RNA isolation and gene expression
analysis
Total cellular RNA was extracted using the RNA
isolation solution, Tripure (Roche Diagnostics GmbH) according to
the manufacturer's instructions. Exosome extraction from the
culture medium was performed using the Total Exosome Isolation
reagent (Thermo Fisher Scientific, Inc., Waltham, MA, USA) as per
the manufacturer's instructions. RNA was isolated from the
extracted exosomes using the Total Exosome RNA & Protein
Isolation kit (Thermo Fisher Scientific, Inc.). GAS5
expression levels were quantified as previously reported (13), using GAPDH as internal control. The
results of three different experiments were used.
Statistical analysis
The present study evaluates whether GAS5 in
exosomes is a marker of apoptosis. Results of three independent
cell culture experiments were used. Cell viability in the
drug-treated cells is expressed as the percentage of control cells
while oligonucleosome-associated apoptosis and GAS5
expression levels are presented as fold-change relative to the
control cells. Mean changes relative to basal values were
statistically analyzed using the independent t-test where P<0.05
was considered to indicate a statistically significant difference.
Statistical analyses were performed using SPSS version 15 (SPSS
Inc., Chicago, IL, USA).
Results
Drug-induced cytotoxicity
Taxol- and bleomycin-associated cytotoxicity in
MCF-7 and MDA-MB-231 cells was determined. Cells were treated with
each of the drugs for 24 h, and the anti-proliferative effect was
measured using real-time cellular analysis. The CI, as a mean of
cell proliferation, served as the measure of cytotoxicity.
Different rates of cell viability were obtained, which indicated
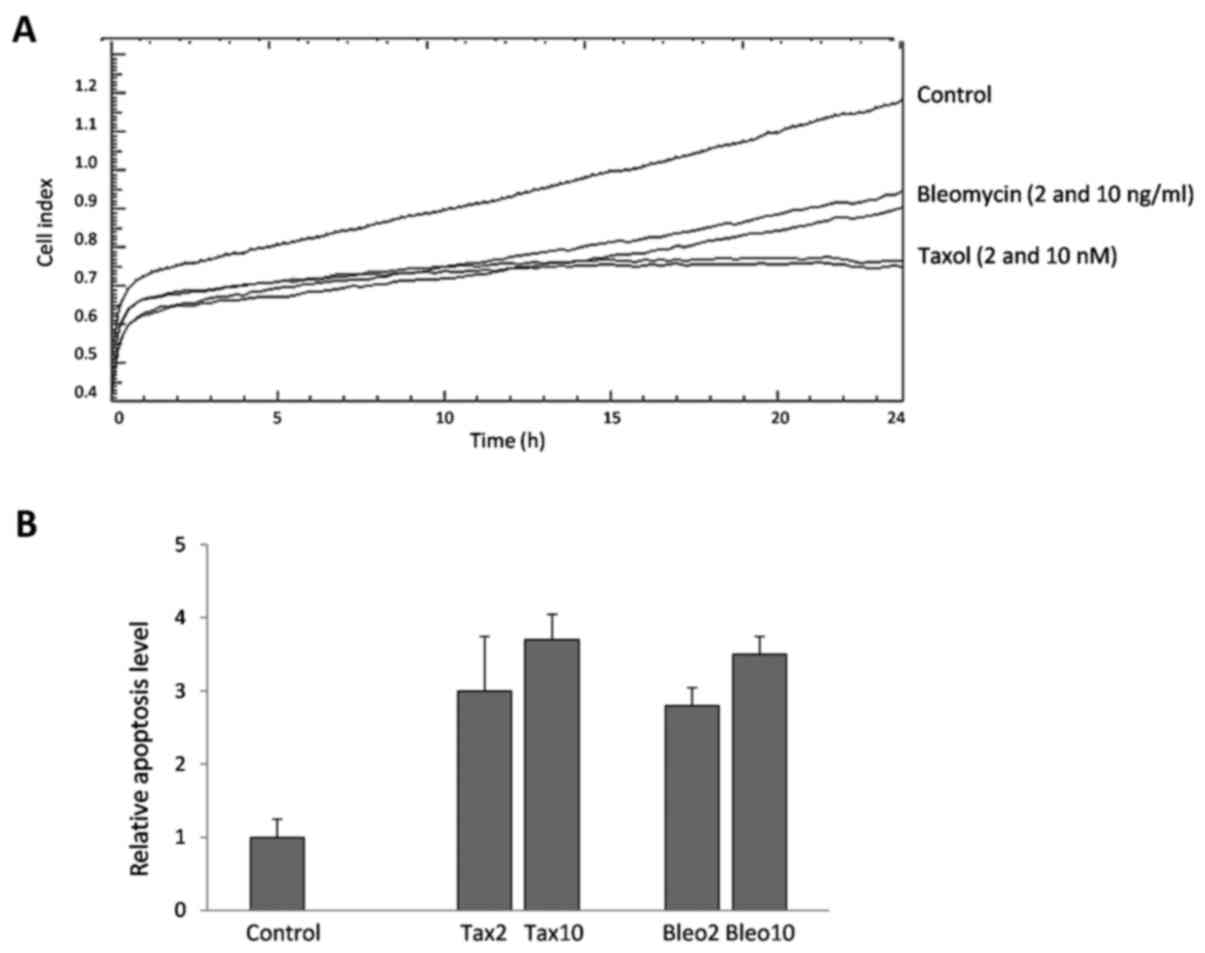
the variable sensitivity of the cell lines to those drugs. Fig. 1A representatively depicts the CI of the
MCF-7 cells, and Table I shows the
percentages of viable cells (% of control) in the MCF-7 and
MDA-MB-231 cells following treatment. MCF-7 cells were more
sensitive to the drugs than the MDA-MB-231 cells, and Taxol (2 and
10 nM) was identified to be more potent than bleomycin (2 and 10
ng/ml) at the applied doses.
 | Table I.Cell index and apoptosis levels in
drug-treated breast cancer cells. |
Table I.
Cell index and apoptosis levels in
drug-treated breast cancer cells.
|
| Mean cell index (% of
control) | Oligonucleosomes
(−fold of control) |
|---|
|
|
|
|
|---|
| Cell line | Taxol 2 | Taxol 10 | Bleo 2 | Bleo 10 | Taxol 2 | Taxol 10 | Bleo 2 | Bleo 10 |
|---|
| MCF-7 | 69 | 67 | 91 | 87 | 3.0 | 3.7 | 2.8 | 3.5 |
| MDA-MB-231 | 80 | 75 | 98 | 97 | 1.8 | 2.1 | 1.5 | 1.9 |
Apoptosis induction
Taxol- and bleomycin-induced apoptosis was measured
in breast cancer cells following 24 h of treatment. Despite
differences in cell viability, considerable levels of cytoplasmic
nucleosomes were detectable in the two cell lines. For example,
similar quantities of nucleosomes were measured in the Taxol- and
bleomycin-treated MCF-7 cells (Fig.
1B), although a difference of ~20% was noted in the mean CI
between the two drugs. Similarly, although the toxicity of the two
drugs was lower in the MDA-MB-231 cells, a considerable level of
apoptosis was also detectable in these cells (Table I).
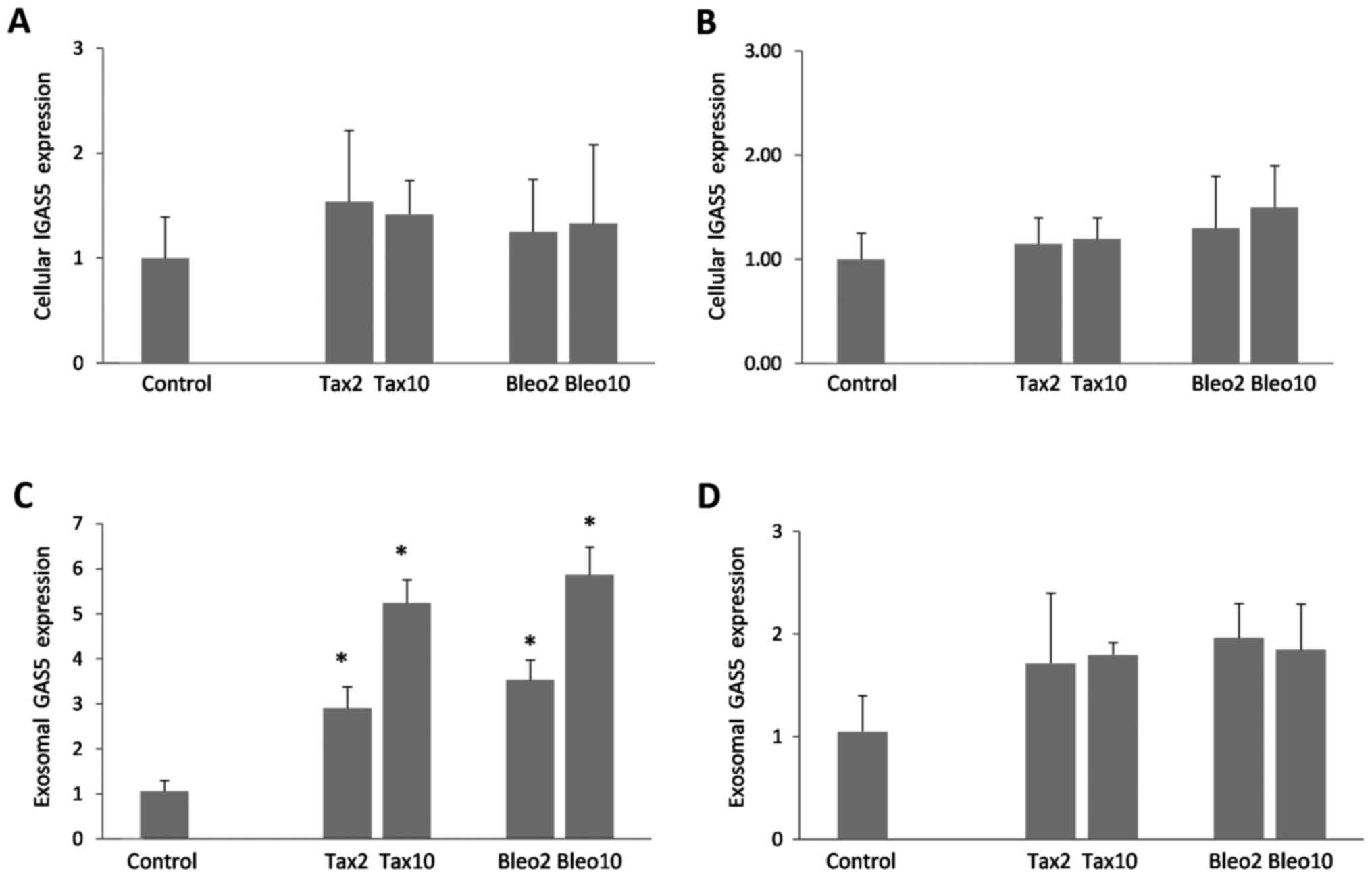
Cellular and exosomal GAS5 expression
levels
Subsequently, GAS5 expression levels in
Taxol- and bleomycin-treated cells, and exosomes secreted from the
drug-treated cells were investigated. In the two cell lines, an
increase of up to 1.5-fold was detected in the cellular expression
levels of GAS5 (Fig. 2A and B,
P>0.05). Compared with cellular GAS5, exosomal expression
levels of GAS5 appeared to be more responsive to apoptosis
induction, as the extent of accumulation in secreted exosomes was
marked, particularly in the MCF-7 cells (Fig. 2C and D). As seen in Fig. 2C and D, dose-dependent increases of
≤5.8-fold (P<0.001) and ≤1.9-fold (P>0.05) were detected in
the MCF-7 and MDA-MB-231 cells, respectively.
Discussion
As important mediators of intercellular
communication, exosomes have been implicated in cancer development
and progression (18). Their
involvement in malignant development is associated with their cargo
in the form of proteins and nucleic acids of different types,
including lncRNAs. In this respect, lncRNAs and exosomes may
function together to disseminate cell signals that alter and/or
control local cellular microenvironments (19). Such an intercellular communication via
RNA molecules may also occur within tumors with high cell turnover.
The present study focused on clarifying whether GAS5 is
enriched in exosomes during the apoptotic processes. To accomplish
this goal, cellular and exosomal accumulation of GAS5 was
investigated in relation to Taxol- and bleomycin-induced apoptosis
in two breast cancer cell lines that differ in hormone receptor
expression and p53 mutational status.
At the administered doses, Taxol was identified to
be more potent than bleomycin in the MCF-7 and MDA-MB-231 cells.
This is consistent with expectations, as Taxol directly interferes
with cell division, whereas bleomycin-associated toxicity occurs
via DNA damage. Similarly, Taxol-associated toxicity was stronger
in MCF-7 cells when compared with MDA-MB-231 cells, which indicates
the involvement of the hormone receptor and p53 status in MD-MB-231
cells. However, a previous study reported that Taxol-induced
apoptosis in MCF-7 and MDA-MB-231 cells is estrogen receptor- and
p53-independent (20). Despite
variances in toxicity rates, similar levels of cell
death-associated nucleosomes were measured following treatment with
the two drugs, thereby indicating the varying efficacy in blocking
cell proliferation and apoptosis induction. In addition, the
current findings confirm that cytoplasmic nucleosomal fragments are
useful tools for evaluating apoptosis in intact cells, as DNA
degradation occurs several hours before plasma membrane breakdown
during the apoptotic process (21).
The primary goal of the present study was to
investigate GAS5 accumulation in exosomes during apoptotic
induction triggered by two different mechanisms, e.g., microtubule
stabilization and DNA strand breaks. Upon apoptosis induction,
cellular expression of GAS5 was induced; however, to a
limited extent (≤1.5-fold) whereas exosomal enrichment was more
pronounced, particularly in MCF-7 cells. These findings indicate
that even a small increase of cellular GAS5 expression leads
to exosomal accumulation. Comparable levels of GAS5
accumulation were found in exosomes that were released from Taxol-
and bleomycin-treated MCF-7 cells, suggesting that GAS5
enrichment in exosomes is associated with apoptosis initiation
rather than the antiproliferative effect of anticancer agents
(Table I). Accordingly, the extent of
exosomal GAS5 enrichment was lower in MDA-MB-231 cells, possibly as
a result of lower cell death rates in these cells.
In conclusion, the current study provides evidence
that GAS5 accumulation in exosomes is a marker of apoptotic
induction. Therefore, it is plausible to hypothesize that
GAS5 is involved in communication of tumor cells upon cell
death-promoting signals. Tracking GAS5 in secreted exosomes
may be useful for evaluating apoptosis and assessing the efficacy
of therapeutic interventions in cancer, as radiotherapy and many
chemotherapeutic agents usually depend upon the efficient
engagement of the apoptotic machinery for their action (22). However, any possible involvement of
GAS5 in exosomal communication of dying tumor cells requires
further research.
Acknowledgements
The present study is part of MSc Thesis of Oguz
Koldemir and was supported by the Istanbul University Scientific
Projects Coordination Unit (grant no. 56669). The authors would
like to thank Mr. David F. Chapman for language editing of the
manuscript.
Glossary
Abbreviations
Abbreviations:
|
lncRNAs
|
long non-coding RNAs
|
|
ncRNAs
|
non-coding RNAs
|
References
|
1
|
Jacquier A: The complex eukaryotic
transcriptome: Unexpected pervasive transcription and novel small
RNAs. Nat Rev Genet. 10:833–844. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jensen TH, Jacquier A and Libri D: Dealing
with pervasive transcription. Mol Cell. 52:473–484. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Berretta J and Morillon A: Pervasive
transcription constitutes a new level of eukaryotic genome
regulation. EMBO Rep. 10:973–982. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mercer TR, Gerhardt DJ, Dinger ME,
Crawford J, Trapnell C, Jeddeloh JA, Mattick JS and Rinn JL:
Targeted RNA sequencing reveals the deep complexity of the human
transcriptome. Nat Biotechnol. 30:99–104. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Derrien T, Johnson R, Bussotti G, Tanzer
A, Djebali S, Tilgner H, Guernec G, Martin D, Merkel A, Knowles DG,
et al: The GENCODE v7 catalog of human long noncoding RNAs:
Analysis of their gene structure, evolution, and expression. Genome
Res. 22:1775–1789. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Harrow J, Frankish A, Gonzalez JM,
Tapanari E, Diekhans M, Kokocinski F, Aken BL, Barrell D, Zadissa
A, Searle S, et al: GENCODE: The reference human genome annotation
for The ENCODE Project. Genome Res. 22:1760–1774. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
St Laurent G, Wahlestedt C and Kapranov P:
The Landscape of long noncoding RNA classification. Trends Genet.
31:239–251. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wilusz JE, Sunwoo H and Spector DL: Long
noncoding RNAs: Functional surprises from the RNA world. Genes Dev.
23:1494–1504. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mercer TR, Dinger ME and Mattick JS: Long
non-coding RNAs: Insights into functions. Nat Rev Genet.
10:155–159. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang KC and Chang HY: Molecular mechanisms
of long noncoding RNAs. Mol Cell. 43:904–914. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Pickard MR and Williams GT: Molecular and
cellular mechanisms of action of tumour suppressor GAS5 lncRNA.
Genes (Basel). 6:484–499. 2015.PubMed/NCBI
|
|
12
|
Ma C, Shi X, Zhu Q, Li Q, Liu Y, Yao Y and
Song Y: The growth arrest-specific transcript 5 (GAS5): A pivotal
tumor suppressor long noncoding RNA in human cancers. Tumour Biol.
37:1437–1444. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gezer U, Özgür E, Cetinkaya M, Isin M and
Dalay N: Long non-coding RNAs with low expression levels in cells
are enriched in secreted exosomes. Cell Biol Int. 38:1076–1079.
2014.PubMed/NCBI
|
|
14
|
EL Andaloussi S, Mäger I, Breakefield XO
and Wood MJ: Extracellular vesicles: Biology and emerging
therapeutic opportunities. Nat Rev Drug Discov. 12:347–357. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Subik K, Lee JF, Baxter L, Strzepek T,
Costello D, Crowley P, Xing L, Hung MC, Bonfiglio T, Hicks DG, et
al: The expression patterns of ER, PR, HER2, CK5/6, EGFR, Ki-67 and
AR by immunohistochemical analysis in breast cancer cell lines.
Breast Cancer (Auckl). 4:35–41. 2010.PubMed/NCBI
|
|
16
|
Zhang J and Xu M: Apoptotic DNA
fragmentation and tissue homeostasis. Trends Cell Biol. 12:84–89.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Özgür E, Mert U, Isin M, Okutan M, Dalay N
and Gezer U: Differential expression of long non-coding RNAs during
genotoxic stress-induced apoptosis in HeLa and MCF-7 cells. Clin
Exp Med. 13:119–126. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Whiteside TL: Tumor-derived exosomes and
their role in cancer progression. Adv Clin Chem. 74:103–141. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hewson C and Morris KV: Form and function
of exosome-associated long non-coding RNAs in cancer. Curr Top
Microbiol Immunol. 394:41–56. 2016.PubMed/NCBI
|
|
20
|
Choi YH and Yoo YH: Taxol-induced growth
arrest and apoptosis is associated with the upregulation of the Cdk
inhibitor, p21WAF1/CIP1, in human breast
cancer cells. Oncol Rep. 28:2163–2169. 2012.PubMed/NCBI
|
|
21
|
Neukirchen J, Meier A, Rohrbeck A,
Garcia-Pardillos G, Steidl U, Fenk R, Haas R, Kronenwett R and Rohr
UP: The proteasome inhibitor bortezomib acts differently in
combination with p53 gene transfer or cytotoxic chemotherapy on
NSCLC cells. Cancer Gene Ther. 14:431–439. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Indran IR, Tufo G, Pervaiz S and Brenner
C: Recent advances in apoptosis, mitochondria and drug resistance
in cancer cells. Biochim Biophys Acta. 1807:735–745. 2011.
View Article : Google Scholar : PubMed/NCBI
|