Skin refers to an organization of tissues on the
surface of the body and is the largest organ of the human body,
accounting for 16% of the body weight (1). Skin is the physical, chemical and
immunological barrier of organisms. Thus, skin prevents the loss of
useful substances from an organism. Skin also undertakes various
functions, including repairing itself, perspiring, detecting
temperature and pressure, and providing additional structural
support (2–4). The development and function of the skin
are affected by multiple factors, including environmental factors
and hormones (5). Recently, exosome
studies have indicated that some exosomes are involved in the
physiological and pathological processes of the skin (6,7). Such
findings have provided a novel perspective for the understanding of
the molecular mechanisms involved in these processes.
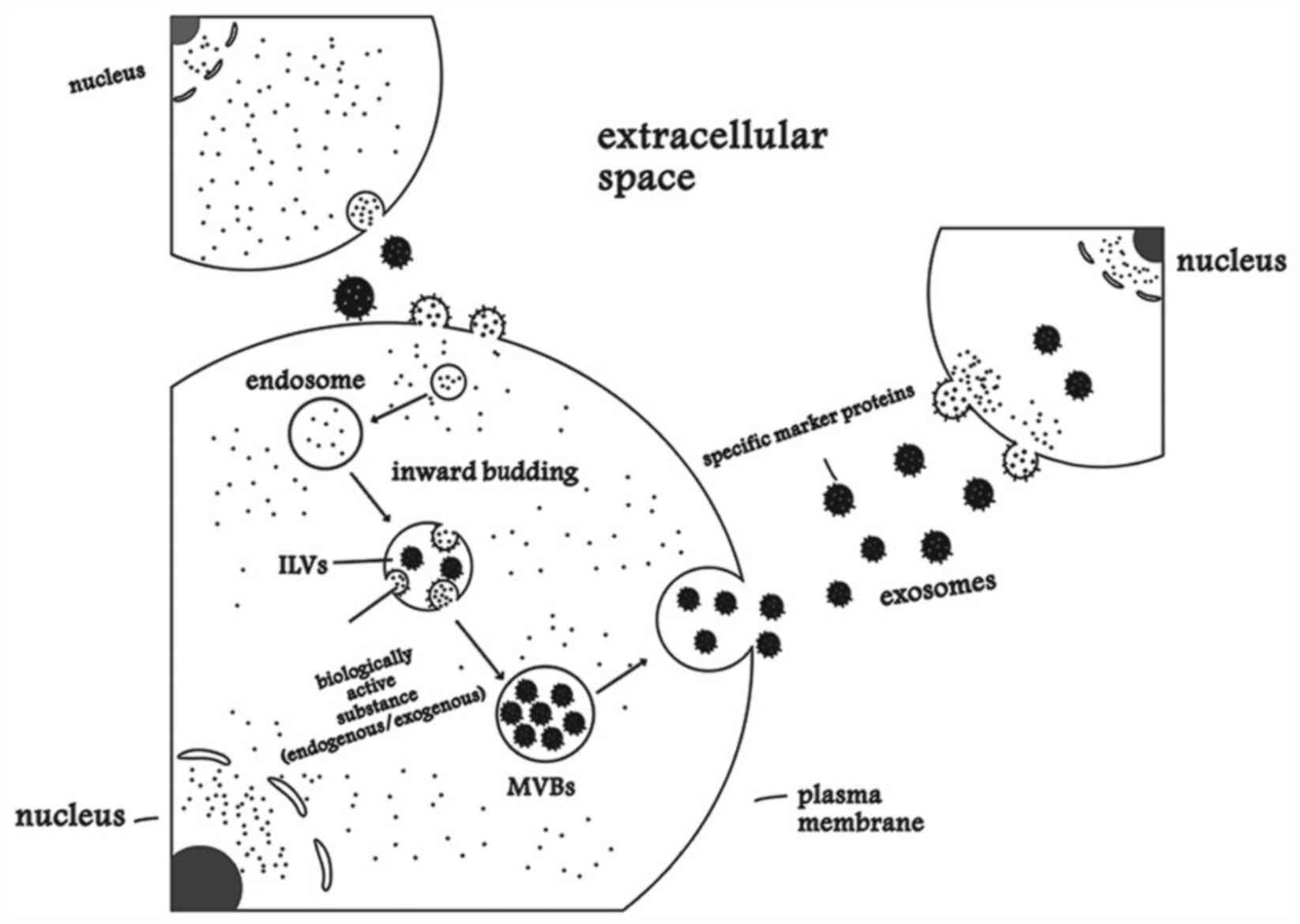
Exosomes are small cell-derived vesicles (~100 nm in
diameter) that are found in the majority of, if not all, biological
fluids (8). Exosomes originate from
endosomes and deliver various bioactive molecules to target cells
(9). The recognition of exosomes by
target cells is a specific, and involves the following events: i)
Recognition between surface receptors; ii) direct fusion of the
exosome with target cell membrane; and iii) ingestion by the target
cell through endocytosis (10). They
are important mediators of cell-to-cell communication, which is
involved in multiple processes, including the immune response,
apoptosis, angiogenesis and inflammation (11). Cells secrete exosomes that present
signal molecules to nearby or distant tissues or cells, serving a
regulatory role (12). To date, the
mechanism of action of exosomes is poorly understood. Therefore,
further studies are required to determine how exosomes regulate
skin diseases and functions.
To date, studies on exosomes have become gradually
applicable in clinical practice. For instance, exosome mRNA may be
used as a diagnostic marker for lung cancer (13), exosome-derived glutathione peroxidase
1 may restore hepatic oxidant injury (14), and exosomal glypican 1 and its
regulatory miRNAs may be specific markers for the detection and
targeted therapy of colorectal cancer (15). With further study, the clinical
applications of exosome markers may be increased.
The skin is a complex structure composed of multiple
cell types and proteins, which comprise the epidermis, dermis and
subcutaneous tissue (3). Skin is the
indispensable barrier between the organism and the environment.
Barrier function is an irreplaceable function of the skin (16), and the skin contains various accessory
organs (17). There are multiple
types of cells that secrete exosomes in the skin, for example
keratinocytes, fibroblasts and adipocytes secrete exosomes into
other cells or body fluids to participate in biological activity
(6,18,19).
Skin diseases, including viral dermatosis, fungal
dermatosis, connective tissue disease, pigmented dermatosis and
skin tumors, are among the most common diseases in humans (20,21).
Damage to the physiological function and normal structure of the
skin may lead to skin diseases (22).
Understanding the pathogenesis of various types of skin diseases
may contribute to the establishment of effective therapeutics.
Therefore, more research should focus on ways to prevent and treat
skin diseases. Various studies have identified an association
between exosomes and skin diseases (23–25). For
instance, melanoma cells secrete exosomes, which are abundant in
the tumor microenvironment, to impact on the pathological process
(26).
Exosomes are a kind of 40–100-nm biological
membrane-bound structure, which are secreted by a variety of cells,
including hematopoietic cells, mesenchymal stem cells (MSCs),
adipocytes, keratinocytes and tumor cell lines, and released into
the extracellular environment (27).
Exosomes are widely distributed in body fluids (blood, urine and
breast milk) (28). In 1983, Pan and
Johnstone (29) first discovered
exosomes in mammalian reticulocytes from mature sheep.
Exosomes derived from different cells or tissues
have varying functions. A previous study on exosomes demonstrated
that they were associated with cellular functions and disease
states (36). Research has
investigated the relationship between exosomes and the immune
response (37). For example, exosomes
released by stem cells have been indicated to contribute to the
treatment of autoimmune diseases (38). Alloreactive T cells may be activated
by the exosomes secreted by donor dendritic cells (39). Furthermore, the roles of exosomes in
cancer have been reported. Cancer-associated fibroblast-derived
exosomes have been demonstrated to alter the metabolic mechanism of
tumor cells (40). High-grade ovarian
cancer cells secrete effective exosomes that serve an influential
role in tumor angiogenesis (41).
Tumor cell-derived exosomes are involved in tumor progression by
multiple mechanisms, including proliferation, invasion,
immunosuppression and formation of the premetastatic niche
(42). For instance, tumor
cell-derived exosomes are able to destroy the host immune system,
and enhance tumor cell survival, growth and migration (43).
In addition, exosomes contain biologically active
substances, including proteins, mRNA, microRNA (miRNA), cytokines
and transcription factors (44).
miRNA, single-chain and non-coding RNA with a length of 18–25 nt,
regulate gene expression at the post-transcriptional level
(45). miRNA interact with mRNA and
have been indicated to degrade target mRNA or inhibit target mRNA
translation (7). A single miRNA may
regulate several target mRNA, and multiple miRNA may also bind to
one target mRNA (46). Increasing
evidence suggests that miRNA serve a critical role in multiple
biological functions (47). Exosomes
derived from MSCs attenuate renal fibrosis via miRNA let7c
(48). Adipose tissue-derived MSC
exosomes enhance hepatocellular carcinoma chemosensitivity by
delivering miRNA-122 to the tumor cells (49). miRNA-9 and miRNA-124 of exosomes
released into the serum may be utilized as biomarkers for acute
ischemic stroke and to evaluate the degree of damage caused by
ischemic injury (50). However, in
the present review, the predominant focus will be on the regulation
of skin functions and diseases by exosomes. These physiological and
pathological processes in which exosomes are involved are outlined
in Table I.
The development of the skin, particularly the
epidermis, is essential for the survival of organisms. If the
epidermis cannot be sufficiently self-renewed and differentiated,
it may lead to a variety of skin diseases (51). Balance between this self-renewal and
differentiation needs to be regulated by progenitor and stem cells
(52). A previous study demonstrated
that subunits of the exosomes (EXOSC7, EXOSC9 and EXOSC10) are
abundant in epidermal progenitor cells, and are essential to avoid
premature differentiation of progenitor cells (53), the mechanism for which may involve
reduced production of grainyhead like transcription factor 3, a
transcription factor essential for epidermal differentiation,
through targeted degradation of its mRNA (53,54).
Wnt signaling pathways serve an important role in
various developmental processes, such as the self-renewal of
mammalian skin or tissue (55). A
previous study identified that Wnt proteins are transported through
the exosomes (56). Wnt induces the
activation of β-catenin in endothelial cells by exosomes, thus
promoting skin repair (57).
Hair of humans is the role of necessity and
metabolism, and its predominant function is for warmth in mammals.
Androgenetic alopecia is a common chronic skin condition that leads
to hair loss in humans (58). A
patented clinical study suggested that a pharmaceutical composition
prepared by using exosomes may promote or enhance hair growth,
making the hair thicker and longer. The exosomes were secreted by
stem cells, such as MSCs (59).
The occurrence and mechanisms of skin and hair color
development in mammals and humans are complicated and affected by
multiple factors. Pigmentation is the core of the process, and is
predominantly regulated by α-melanocyte-stimulating
hormone/melanocortin 1 receptor, mitogen-activated protein kinase
(stem cell factor/c-Kit), eosinophil-derived neurotoxin and the Wnt
signaling pathway (60). Skin
pigmentation requires close communication between cells. In the
skin epidermis, melanocytes and keratinocytes form the epidermal
melanin unit (6). Ultraviolet B is
able to induce melanin production in melanoma cells (6). The difference in skin color is
controlled by the melanin content (61). Abnormal skin pigmentation may lead to
hyper and hypopigmentation disorder; a study by Lo Cicero et
al (6) recently indicated that
human keratinocyte-derived exosomes carry specific miRNA
(miRNA-3196 and miRNA-203) and target melanocytes to regulate
pigmentation, through the alteration of gene expression and the
activation of key enzymes.
Vitiligo is a common pigmentation disorder of the
skin, caused by the loss of functional melanocytes (62). The data presented in a previous study
demonstrated that exosomes isolated from sera of active vitiligo
patients differentially expressed 47 miRNA compared with healthy
controls (23). This indicated a
difference in the miRNAs between the serum exosomes of patients
with vitiligo and those of healthy individuals, and contributed to
understanding the role of melanocytes in the pathogenesis of
vitiligo. Such research may allow for the development of a novel
approach to manipulate pigmentation via exosomes, including
crosstalk between melanocytes and keratinocytes, and the
pathogenesis of abnormal pigment disorders.
Previous studies, listed below, have indicated that
exosomes promote skin wound healing through a paracrine mechanism,
and suggest that the principal mechanism depends on the source of
the exosomes. However, cells derived from different sources,
including adipose mesenchymal cells and human amniotic epithelial
cells, have demonstrated similar characteristics to MSCs (63). Exosomes of endothelial progenitor
cells from human umbilical cord blood may also accelerate skin
wound healing (64). Exosomes
released from human induced pluripotent stem cell-derived MSCs have
the ability of tissue repair by promoting collagen synthesis and
angiogenesis (63). Similarly,
exosomes secreted by cardiosphere-derived cells contribute to scar
reduction (65).
In addition, adipose MSC-derived exosomes
(ASCs-Exos) are internalized by fibroblasts to influence cell
migration, proliferation and collagen synthesis (24). Histological analyses have indicated
that the effect of ASCs-Exos on collagen is different between the
early and later stages of wound healing, with increased collagen I
and III synthesis at early stages and decreased collagen synthesis
at later stages (24). ASCs-Exos may
also promote cutaneous wound healing via optimizing the properties
of fibroblasts (24). Human amniotic
epithelial cell-derived exosomes have been demonstrated to expedite
wound healing and inhibit scar formation by promoting proliferation
and migration of fibroblasts (66).
Human umbilical cord MSC-derived exosomes (hucMSC-Exos) through the
Wnt4 pathway are also involved in skin wound healing. HucMSC-Exos
facilitate the function of endothelial cells; various studies have
demonstrated that the hucMSC-Exo-mediated Wnt4 pathway induced
β-catenin activation in endothelial cells and enhanced vascular
regeneration (57,67). Furthermore, research has suggested
that exosomes from the serum may accelerate wound healing in mice,
predominantly due to the impact on collagen synthesis (68).
At present, the most notably obstacle in the
treatment of skin wounds is how to prolong healing, particularly in
the cases of wound recovery in diabetic patients and patients who
have received surgery. Therefore, exosomes may provide a promising
approach for skin wound repair.
Skin cancer is a common disease and the morbidity is
increasing dramatically, particularly in Caucasians. For example,
the incidence of skin cancer among Caucasians in America reached
165 per 10,000 individuals in 2015 (69,70). Skin
cancers include melanoma, basal cell carcinoma and squamous cell
carcinoma (SCC); the latter two diseases are non-melanoma (69). Tumor cell-derived exosomes are
abundant in the tumor microenvironment; they contribute to the
formation of the premetastatic niche and attack the host's immune
system (26). Exosomes may be used as
an effective tool in cancer diagnosis and prognosis due to their
particular pro-tumorigenic characteristics (25).
Melanoma, also known as malignant melanoma, forms
through malignant transformation of melanocytes and often occurs in
the skin. Melanoma is the most malignant tumor of the skin tumors
(71). Due to the aggressiveness,
ease of distant metastasis and treatment resistance, melanoma has a
high mortality rate (72–74). Early diagnosis and treatment is
therefore particularly important.
Previous studies have indicated that miRNA are also
closely related to melanoma (7,78–80). For instance, melanoma cells secrete
exosomes carrying miR-222 to promote tumorigenesis by activating
the phosphoinositide 3-kinase/AKT pathway (7). The expression level of miRNA-125b was
downregulated in serum exosomes of patients with melanoma compared
with the levels in healthy individuals. miRNA-125b is a
characteristic miRNA from melanoma-derived exosomes, which may
directly inhibit the expression of c-Jun protein and control the
development of melanoma (81).
miRNA-17, −19a, −21, −126 and −149 were highly expressed in the
exosomes of patients with metastatic sporadic melanoma compared
with the levels in patients with familial melanoma and normal
controls. Notably, there was no significant difference in the miRNA
expression level between the healthy controls and the patients with
familial melanoma (82).
Non-melanoma, predominantly SCC, may use exosomes to
increase crosstalk between tumor cells and the tumor
microenvironment (83). Transforming
growth factor-β (TGF-β) signaling is important for cancer
occurrence and development. It has been demonstrated that exosomes
in the tumor microenvironment have the ability to re-activate TGF-β
signaling in SSC (84). Exosomes
secreted from head and neck SCC cells change their protein
composition in response to ionizing radiation, which may induce
changes in translation or transcription (85). Relevant research data has indicated
that the exosomes of head and neck SCC cells also promote the
survival of recipient cells (83).
In summary, exosomes serve an important role in the
occurrence and development of tumors. Full elucidation of the
mechanism of exosome function is conducive to the diagnosis and
treatment of skin cancer.
Scleroderma is a connective tissue disease, and its
main features include endothelial cell dysfunction, immune
activation and excessive skin fibrosis (86). A previous study demonstrated that
patients with scleroderma had a reduced level of exosomes in their
serum, which was predominantly due to vascular abnormalities
disrupting the transportation of exosomes derived from the
fibrocytes of skin tissue (68,87).
However, in dermatomyositis and scleroderma overlap syndrome,
exosomes exist as an autoantigenic complex (88,89).
Another disease associated with skin is obesity.
Obesity is a common metabolic syndrome and a systemic inflammatory
state that affects almost every organ system in the body (90). Adipocytes of people with obesity may
induce inflammation (91).
Fortunately, adipocyte-derived exosomes may carry miRNA to regulate
end-organ TGF-β and Wnt/β-catenin signaling, which are inflammatory
and fibrotic signaling pathways (92). A recent study supports this view; the
exosome-like vesicles secreted by adipose tissues carry ~one third
of miRNA relevant to adipogenesis to induce adipogenesis of adipose
tissue-derived stem cells (93). A
study by Skowron et al (94)
suggested that obesity increased the risk of melanoma malignant
metastasis. Recently, a mechanism by which exosomes are linked to
obesity and melanoma was identified. It was revealed that adipocyte
exosomes carried the majority of proteins involved in lipid
metabolism, primarily for fatty acid oxidation. Crucially, this
feature contributed to the migration and invasion of melanoma cells
(19,95). These results may explain why patients
with obesity and melanoma have a poor prognosis, and may lead to a
novel strategy for the treatment of melanoma by reducing body fat
content or inhibiting fatty acid oxidation.
The discovery and confirmation of the role of
exosomes has had a profound influence on the understanding of
organism physiology and pathological processes. Exosomes not only
carry pathological or physiological marker proteins/RNA of donor
cells, but may also act as a vehicle for the transfer of drugs and
other molecules to lesion sites. Furthermore, exosomes also have
the potential to be modified and processed. In the present review,
exosomes involved in skin development, function and diseases were
discussed. Further in-depth studies of exosomes are required to
determine whether they may be utilized as high-performance tools
and as a novel pathway for regulating skin development and diseases
in the future.
Not applicable.
The present study was supported by grants from the
National Natural Science Foundation of China (grant nos. 81700650
and 31772690).
Not applicable.
YL drafted the article, HW revised the article and
JW provided ideas and helped to improve language accuracy.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Ng KW and Lau WM: Skin Deep: The Basics of
Human Skin Structure and Drug Penetration. Springer-Verlag; Berlin
Heidelberg, New York: 2015
|
|
2
|
Menon GK: Skin basics; structure and
functionLipids and Skin Health. Springer International Publishing;
Switzerland: pp. 9–23. 2015
|
|
3
|
Barbieri JS, Wanat K and Seykora J: Skin:
Basic structure and functionPathobiology of Human Disease. Academic
Press; pp. 1134–1144. 2014, View Article : Google Scholar
|
|
4
|
Mcgrath JA, Eady RAJ and Pope FM: Anatomy
and organization of human skinRook's Textbook of Dermatology. 7th.
Wiley; pp. 45–128. 2008
|
|
5
|
Hay RJ, Johns NE, Williams HC, Bolliger
IW, Dellavalle RP, Margolis DJ, Marks R, Naldi L, Weinstock MA,
Wulf SK, et al: The global burden of skin disease in 2010: An
analysis of the prevalence and impact of skin conditions. J Invest
Dermatol. 134:1527–1534. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lo Cicero A, Delevoye C, Gilles-Marsens F,
Loew D, Dingli F, Guéré C, André N, Vié K, van Niel G and Raposo G:
Exosomes released by keratinocytes modulate melanocyte
pigmentation. Nat Commun. 6:75062015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Felicetti F, De Feo A, Coscia C, Puglisi
R, Pedini F, Pasquini L, Bellenghi M, Errico MC, Pagani E and Carè
A: Exosome-mediated transfer of miR-222 is sufficient to increase
tumor malignancy in melanoma. J Transl Med. 14:562016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lin J, Li J, Huang B, Liu J, Chen X, Chen
XM, Xu YM, Huang LF and Wang XZ: Exosomes: Novel biomarkers for
clinical diagnosis. Sci World J. 2015:6570862015. View Article : Google Scholar
|
|
9
|
Properzi F, Logozzi M and Fais S:
Exosomes: The future of biomarkers in medicine. Biomarkers Med.
7:769–778. 2013. View Article : Google Scholar
|
|
10
|
Cocucci E, Racchetti G and Meldolesi J:
Shedding microvesicles: Artefacts no more. Trends Cell Biol.
19:43–51. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bach DH, Hong JY, Park HJ and Lee SK: The
role of exosomes and miRNAs in drug-resistance of cancer cells. Int
J Cancer. 141:220–230. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lai RC, Chen TS and Lim SK: Mesenchymal
stem cell exosome: A novel stem cell-based therapy for
cardiovascular disease. Regen Med. 6:481–492. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rabinowits G, Gerçel-Taylor C, Day JM,
Taylor DD and Kloecker GH: Exosomal microRNA: A diagnostic marker
for lung cancer. Clin Lung Cancer. 10:42–46. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yan Y, Jiang W, Tan Y, Zou S, Zhang H, Mao
F, Gong A, Qian H and Xu W: hucMSC exosome-derived GPX1 is required
for the recovery of hepatic oxidant injury. Mol Ther. 25:465–479.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li J, Chen Y, Guo X, Zhou L, Jia Z, Peng
Z, Tang Y, Liu W, Zhu B, Wang L and Ren C: GPC1 exosome and its
regulatory miRNAs are specific markers for the detection and target
therapy of colorectal cancer. J Cell Mol Med. 21:838–847. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Proksch E, Brandner JM and Jensen JM: The
skin: An indispensable barrier. Exp Dermatol. 17:1063–1072. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Forslind B and Lindberg M: Skin, Hair, and
Nails: Structure and Function. CRC Press; 2003
|
|
18
|
Bang C, Batkai S, Dangwal S, Gupta SK,
Foinquinos A, Holzmann A, Just A, Remke J, Zimmer K, Zeug A, et al:
Cardiac fibroblast-derived microRNA passenger strand-enriched
exosomes mediate cardiomyocyte hypertrophy. J Clin Invest.
124:2136–2146. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Holmes D: Adipose tissue: Adipocyte
exosomes drive melanoma progression. Nat Rev Endocrinol.
12:4362016. View Article : Google Scholar
|
|
20
|
Bickers DR and Athar M: Oxidative stress
in the pathogenesis of skin disease. J Invest Dermatol.
126:2565–2575. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Akita N, Sawamura D, Matsumura K and
Nomura K: Clinical study of diflorasone diacetate ointment
(Diflal® ointment) in various types of skin diseases.
Skin Res. 29:115–119. 2010.
|
|
22
|
Balato N, Megna M, Ayala F, Balato A,
Napolitano M and Patruno C: Effects of climate changes on skin
diseases. Expert Rev Anti Infect Ther. 12:171–181. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu L, Song P, Yi X, Li C and Gao T: 067
Serum-derived exosomes contribute to abnormal melanocyte function
in patients with active vitiligo. J Invest Dermatol. 136:S12. 2016.
View Article : Google Scholar
|
|
24
|
Hu L, Wang J, Zhou X, Xiong Z, Zhao J, Yu
R, Huang F, Zhang H and Chen L: Exosomes derived from human adipose
mensenchymal stem cells accelerates cutaneous wound healing via
optimizing the characteristics of fibroblasts. Sci Rep.
6:329932016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Goedert L, Koya R, Hu-Lieskovan S and
Ribas A: Exosomes as a predictor tool of acquired resistance to
melanoma treatment. BMC Proc. 8 Suppl 4:pp. P282014; View Article : Google Scholar
|
|
26
|
Xiao D, Barry S, Kmetz D, Egger M, Pan J,
Rai SN, Qu J, McMasters KM and Hao H: Melanoma cell-derived
exosomes promote epithelial-mesenchymal transition in primary
melanocytes through paracrine/autocrine signaling in the tumor
microenvironment. Cancer Lett. 376:318–327. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Simpson RJ, Jensen SS and Lim JW:
Proteomic profiling of exosomes: Current perspectives. Proteomics.
8:4083–4099. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Vlassov AV, Magdaleno S, Setterquist R and
Conrad R: Exosomes: Current knowledge of their composition,
biological functions, and diagnostic and therapeutic potentials.
Biochim Biophys Acta. 1820:940–948. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Pan BT and Johnstone RM: Fate of the
transferrin receptor during maturation of sheep reticulocytes in
vitro: Selective externalization of the receptor. Cell. 33:967–978.
1983. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mathivanan S, Ji H and Simpson RJ:
Exosomes: Extracellular organelles important in intercellular
communication. J Proteomics. 73:1907–1920. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Simpson RJ, Lim JW, Moritz RL and
Mathivanan S: Exosomes: Proteomic insights and diagnostic
potential. Expert Rev Proteomics. 6:267–283. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lakkaraju A and Rodriguez-Boulan E:
Itinerant exosomes: Emerging roles in cell and tissue polarity.
Trends Cell Biol. 18:199–209. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
van Niel G, Porto-Carreiro I, Simoes S and
Raposo G: Exosomes: A common pathway for a specialized function. J
Biochem. 140:13–21. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Segura MF, Hanniford D, Menendez S, Reavie
L, Zou X, Alvarez-Diaz S, Zakrzewski J, Blochin E, Rose A,
Bogunovic D, et al: Aberrant miR-182 expression promotes melanoma
metastasis by repressing FOXO3 and microphthalmia-associated
transcription factor. Proc Natl Acad Sci USA. 106:pp. 1814–1819.
2009; View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Conde-Vancells J, Rodriguez-Suarez E,
Embade N, Gil D, Matthiesen R, Valle M, Elortza F, Lu SC, Mato JM
and Falcon-Perez JM: Characterization and comprehensive proteome
profiling of exosomes secreted by hepatocytes. J Proteome Res.
7:5157–5166. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhou H, Cheruvanky A, Hu X, Matsumoto T,
Hiramatsu N, Cho ME, Berger A, Leelahavanichkul A, Doi K, Chawla
LS, et al: Urinary exosomal transcription factors, a new class of
biomarkers for renal disease. Kidney Int. 74:613–621. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Théry C, Ostrowski M and Segura E:
Membrane vesicles as conveyors of immune responses. Nat Rev
Immunol. 9:581–593. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ichim T and Bogin V: Therapeutic immune
modulation by stem cell secreted exosomes. US Patent 20160361399
A1. August 4–2016.issued December 15, 2016.
|
|
39
|
Liu Q, Rojas-Canales DM, Divito SJ,
Shufesky WJ, Stolz DB, Erdos G, Sullivan ML, Gibson GA, Watkins SC,
Larregina AT, et al: Donor dendritic cell-derived exosomes promote
allograft-targeting immune response. J Clin Invest. 126:2805–2820.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhao H, Yang L, Baddour J, Achreja A,
Bernard V, Moss T, Marini JC, Tudawe T, Seviour EG, San Lucas FA,
et al: Tumor microenvironment derived exosomes pleiotropically
modulate cancer cell metabolism. Elife. 5:e102502016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yi H, Ye J, Yang XM, Zhang LW, Zhang ZG
and Chen YP: High-grade ovarian cancer secreting effective exosomes
in tumor angiogenesis. Int J Clin Exp Pathol. 8:5062–5070.
2015.PubMed/NCBI
|
|
42
|
Tickner JA, Urquhart AJ, Stephenson SA,
Richard DJ and O'Byrne KJ: Functions and therapeutic roles of
exosomes in cancer. Front Oncol. 4:1272014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Saleem SN and Abdel-Mageed AB:
Tumor-derived exosomes in oncogenic reprogramming and cancer
progression. Cell Mol Life Sci. 72:1–10. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Chevillet JR, Kang Q, Ruf IK, Briggs HA,
Vojtech LN, Hughes SM, Cheng HH, Arroyo JD, Meredith EK,
Gallichotte EN, et al: Quantitative and stoichiometric analysis of
the microRNA content of exosomes. Proc Natl Acad Sci USA. 111:pp.
14888–14893. 2014; View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Bi S, Wang C, Jin Y, Lv Z, Xing X and Lu
Q: Correlation between serum exosome derived miR-208a and acute
coronary syndrome. Int J Clin Exp Med. 8:4275–4280. 2015.PubMed/NCBI
|
|
46
|
Wang H and Wang B: Extracellular vesicle
microRNAs mediate skeletal muscle myogenesis and disease. Biomed
Rep. 5:296–300. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Humphries B: Dissecting the mechanism by
which microRNA-200b inhibits breast cancer metastasis. PhD
dissertationMichigan State University 2016
|
|
48
|
Wang B, Yao K, Huuskes BM, Shen HH, Zhuang
J, Godson C, Brennan EP, Wilkinson-Berka JL, Wise AF and Ricardo
SD: Mesenchymal stem cells deliver exogenous microRNA-let7c via
exosomes to attenuate renal fibrosis. Mol Ther. 24:1290–1301. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Lou G, Song X, Yang F, Wu S, Wang J, Chen
Z and Liu Y: Exosomes derived from miR-122-modified adipose
tissue-derived MSCs increase chemosensitivity of hepatocellular
carcinoma. J Hematol Oncol. 8:1222015. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Ji Q, Ji Y, Peng J, Zhou X, Chen X, Zhao
H, Xu T, Chen L and Xu Y: Increased brain-specific miR-9 and
miR-124 in the serum exosomes of acute ischemic stroke patients.
PLoS One. 11:e01636452016. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Jackson SJ, Zhang Z, Feng D, Flagg M,
O'Loughlin E, Wang D, Stokes N, Fuchs E and Yi R: Rapid and
widespread suppression of self-renewal by microRNA-203 during
epidermal differentiation. Development. 140:1882–1891. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Nijhof JG, van Pelt C, Mulder AA, Mitchell
DL, Mullenders LH and de Gruijl FR: Epidermal stem and progenitor
cells in murine epidermis accumulate UV damage despite NER
proficiency. Carcinogenesis. 28:792–800. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Mistry DS, Chen Y and Sen GL: Progenitor
function in self-renewing human epidermis is maintained by the
exosome. Cell Stem Cell. 11:127–135. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Noiret M, Mottier S, Angrand G,
Gautier-Courteille C, Lerivray H, Viet J, Paillard L, Mereau A,
Hardy S and Audic Y: Ptbp1 and Exosc9 knockdowns trigger skin
stability defects through different pathways. Dev Biol.
409:489–501. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Clevers H: Wnt/β-catenin signaling in
development and disease. Cell. 127:469–480. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Gross JC, Chaudhary V, Bartscherer K and
Boutros M: Active Wnt proteins are secreted on exosomes. Nat Cell
Biol. 14:1036–1045. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Zhang B, Wang M, Gong A, Zhang X, Wu X,
Zhu Y, Shi H, Wu L, Zhu W, Qian H and Xu W: HucMSC-exosome
mediated-Wnt4 signaling is required for cutaneous wound healing.
Stem Cells. 33:2158–2168. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Varothai S and Bergfeld WF: Androgenetic
alopecia: An evidence-based treatment update. Am J Clin Dermatol.
15:217–230. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Lim SK, Yeo MSW, Chen TS and Lai RC: Use
of exosomes to promote or enhance hair growth EP Patent 2629782 A1.
October 17–2011, issued August 28, 2013.
|
|
60
|
Lin JY and Fisher DE: Melanocyte biology
and skin pigmentation. Nature. 445:843–850. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Rawlings AV: Ethnic skin types: Are there
differences in skin structure and function? Int J Cosmet Sci.
28:79–93. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Whitton M, Pinart M, Batchelor JM,
Leonardi-Bee J, Gonzalez U, Jiyad Z, Eleftheriadou V and Ezzedine
K: Evidence-based management of vitiligo: Summary of a Cochrane
systematic review. Br J Dermatol. 174:962–969. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Zhang J, Guan J, Niu X, Hu G, Guo S, Li Q,
Xie Z, Zhang C and Wang Y: Exosomes released from human induced
pluripotent stem cells-derived MSCs facilitate cutaneous wound
healing by promoting collagen synthesis and angiogenesis. J Transl
Med. 13:492015. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Li X, Jiang C and Zhao J: Human
endothelial progenitor cells-derived exosomes accelerate cutaneous
wound healing in diabetic rats by promoting endothelial function. J
Diabetes Complications. 30:986–992. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Gallet R, Dawkins J, Valle J, Simsolo E,
de Couto G, Middleton R, Tseliou E, Luthringer D, Kreke M, Smith
RR, et al: Exosomes secreted by cardiosphere-derived cells reduce
scarring, attenuate adverse remodelling, and improve function in
acute and chronic porcine myocardial infarction. Eur Heart J.
38:201–211. 2017.PubMed/NCBI
|
|
66
|
Zhao B, Zhang Y, Han S, Zhang W, Zhou Q,
Guan H, Liu J, Shi J, Su L and Hu D: Exosomes derived from human
amniotic epithelial cells accelerate wound healing and inhibit scar
formation. J Mol Histol. 48:121–132. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Zhang B, Wu X, Zhang X, Sun Y, Yan Y, Shi
H, Zhu Y, Wu L, Pan Z, Zhu W, et al: Human umbilical cord
mesenchymal stem cell exosomes enhance angiogenesis through the
Wnt4/β-catenin pathway. Stem Cells Transl Med. 4:513–522. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Nakamura K, Jinnin M, Fukushima S and Ihn
H: Exosome expression in the skin and sera of systemic sclerosis
patients, and its possible therapeutic application against skin
ulcer. J Dermatol Sci. 84:e97–e98. 2016. View Article : Google Scholar
|
|
69
|
Diepgen TL and Mahler V: The epidemiology
of skin cancer. Br J Dermatol. 146 Suppl 61:1–6. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Wernli KJ, Henrikson NB, Morrison CC,
Nguyen M, Pocobelli G and Blasi PR: Screening for skin cancer in
adults: Updated evidence report and systematic review for the US
Preventive Services Task Force. JAMA. 316:436–447. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Hodi FS, O'Day SJ, McDermott DF, Weber RW,
Sosman JA, Haanen JB, Gonzalez R, Robert C, Schadendorf D, Hassel
JC, et al: Improved survival with ipilimumab in patients with
metastatic melanoma. N Engl J Med. 363:711–723. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Alegre E, Zubiri L, Perez-Gracia JL,
González-Cao M, Soria L, Martín-Algarra S and González A:
Circulating melanoma exosomes as diagnostic and prognosis
biomarkers. Clin Chim Acta. 454:28–32. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Gray-Schopfer V, Wellbrock C and Marais R:
Melanoma biology and new targeted therapy. Nature. 445:851–857.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Gajos-Michniewicz A, Duechler M and Czyz
M: miRNA in melanoma-derived exosomes. Cancer Lett. 347:29–37.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Hood JL, San RS and Wickline SA: Exosomes
released by melanoma cells prepare sentinel lymph nodes for tumor
metastasis. Cancer Res. 71:3792–3801. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Ekström EJ, Bergenfelz C, von Bülow V,
Serifler F, Carlemalm E, Jönsson G, Andersson T and Leandersson K:
WNT5A induces release of exosomes containing pro-angiogenic and
immunosuppressive factors from malignant melanoma cells. Mol
Cancer. 13:882014. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Hood JL: Melanoma exosomes enable tumor
tolerance in lymph nodes. Med Hypotheses. 90:11–13. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Shoshan E, Mobley AK, Braeuer RR, Kamiya
T, Huang L, Vasquez ME, Salameh A, Lee HJ, Kim SJ, Ivan C, et al:
Reduced adenosine-to-inosine miR-455-5p editing promotes melanoma
growth and metastasis. Nat Cell Biol. 17:311–321. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Zhou J, Xu D, Xie H, Tang J, Liu R, Li J,
Wang S, Chen X, Su J, Zhou X, et al: miR-33a functions as a tumor
suppressor in melanoma by targeting HIF-1α. Cancer Biol Ther.
16:846–855. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Bhattacharya A, Schmitz U, Raatz Y,
Schönherr M, Kottek T, Schauer M, Franz S, Saalbach A, Anderegg U,
Wolkenhauer O, et al: miR-638 promotes melanoma metastasis and
protects melanoma cells from apoptosis and autophagy. Oncotarget.
6:2966–2980. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Alegre E, Sanmamed MF, Rodriguez C,
Carranza O, Martín-Algarra S and González A: Study of circulating
microRNA-125b levels in serum exosomes in advanced melanoma. Arch
Pathol Lab Med. 138:828–832. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Pfeffer SR, Grossmann KF, Cassidy PB, Yang
CH, Fan M, Kopelovich L, Leachman SA and Pfeffer LM: Detection of
exosomal miRNAs in the plasma of melanoma patients. J Clin Med.
4:2012–2027. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Mutschelknaus L, Peters C, Winkler K,
Yentrapalli R, Heider T, Atkinson MJ and Moertl S: Exosomes derived
from squamous head and neck cancer promote cell survival after
ionizing radiation. PLoS One. 11:e01522132016. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Languino LR, Singh A, Prisco M, Inman GJ,
Luginbuhl A, Curry JM and South AP: Exosome-mediated transfer from
the tumor microenvironment increases TGFβ signaling in squamous
cell carcinoma. Am J Transl Res. 8:2432–2437. 2016.PubMed/NCBI
|
|
85
|
Jelonek K, Wojakowska A, Marczak L, Muer
A, Tinhofer-Keilholz I, Lysek-Gladysinska M, Widlak P and
Pietrowska M: Ionizing radiation affects protein composition of
exosomes secreted in vitro from head and neck squamous cell
carcinoma. Acta Biochim Pol. 62:265–272. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Toki S, Motegi S, Yamada K, Uchiyama A,
Kanai S, Yamanaka M and Ishikawa O: Clinical and laboratory
features of systemic sclerosis complicated with localized
scleroderma. J Dermatol. 42:283–287. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Nakamura K, Jinnin M, Harada M, Kudo H,
Nakayama W, Inoue K, Ogata A, Kajihara I, Fukushima S and Ihn H:
Altered expression of CD63 and exosomes in scleroderma dermal
fibroblasts. J Dermatol Sci. 84:30–39. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Brouwer R, Pruijn GJ and van Venrooij WJ:
The human exosome: An autoantigenic complex of exoribonucleases in
myositis and scleroderma. Arthritis Res. 3:102–106. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Gutiérrez-Ramos R, Gonz Lez-Díaz V,
Pacheco-Tovar MG, López-Luna A, Avalos-Díaz E and Herrera-Esparza
R: A dermatomyositis and scleroderma overlap syndrome with a
remarkable high titer of anti-exosome antibodies. Reumatismo.
60:296–300. 2008.PubMed/NCBI
|
|
90
|
Barkai L and Paragh G: Metabolic syndrome
in childhood and adolescence. Orv Hetil. 147:243–250. 2006.(In
Hungarian). PubMed/NCBI
|
|
91
|
Rajala MW and Scherer PE: Minireview: The
adipocyte - at the crossroads of energy homeostasis, inflammation,
and atherosclerosis. Endocrinology. 144:3765–3773. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Ferrante SC, Nadler EP, Pillai DK, Hubal
MJ, Wang Z, Wang JM, Gordish-Dressman H, Koeck E, Sevilla S, Wiles
AA and Freishtat RJ: Adipocyte-derived exosomal miRNAs: A novel
mechanism for obesity-related disease. Pediatr Res. 77:447–454.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Zhang Y, Yu M, Dai M, Chen C, Tang Q, Jing
W, Wang H and Tian W: miR-450a-5p within rat adipose tissue
exosome-like vesicles promotes adipogenic differentiation by
targeting WISP2. J Cell Sci. 130:1158–1168. 2017.PubMed/NCBI
|
|
94
|
Skowron F, Bérard F, Balme B and
Maucort-Boulch D: Role of obesity on the thickness of primary
cutaneous melanoma. J Eur Acad Dermatol Venereol. 29:262–269. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Lazar I, Clement E, Dauvillier S, Milhas
D, Ducoux-Petit M, LeGonidec S, Moro C, Soldan V, Dalle S, Balor S,
et al: Adipocyte exosomes promote melanoma aggressiveness through
fatty acid oxidation: a novel mechanism linking obesity and cancer.
Cancer Res. 76:4051–4057. 2016. View Article : Google Scholar : PubMed/NCBI
|