Introduction
Infertility is defined as the inability to conceive
following >1 year of regular unprotected sexual intercourse and
now affects 10–15% of couples of reproductive age (1,2).
Defective sperm function is a common contributing factor,
accounting for 30–40% of couples attending infertility clinics
(3). Obesity, which is the sixth
most important risk factor contributing to the overall burden of
infertility worldwide (4), has been
reported to impair male infertility (5,6).
Accumulating evidence suggests that male infertility is regulated
by an orchestrated network comprising numerous pathological changes
(7,8). The mechanisms underlying
obesity-associated infertility are complex and remain unclear;
however, previous studies have indicated that excessive cell
apoptosis serves an important role (7,9–11).
Apoptosis is essential for cellular homeostasis and
male germ cell development (12,13).
Increased apoptosis has been observed in the spermatozoa of male
patients with infertility, as well as in the sperm of infertile
mice (14,15). High-fat diet (HFD) has been
demonstrated to induce apoptosis in rodents, which in turn promotes
the progression of infertility (16). Conversely, inhibiting excessive
apoptosis attenuates HFD-induced impairment of spermatogenesis
(17). Therefore, pharmacological
agents that are able to inhibit testicular cell apoptosis are of
great therapeutic interest.
Asiatic acid (AA), which is a pentacyclic triterpene
isolated from Centella asiatica, has been demonstrated to
possess a number of pharmacological activities (18). AA is able to protect against cardiac
hypertrophy (19), reduce islet
fibrosis in animal models of diabetes (20) and ameliorate hepatic lipid
accumulation (21). Furthermore,
previous studies have indicated that AA attenuates
glutamate-induced apoptosis in SH-SY5Y cells (22) and inhibits apoptosis in the striatum
of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-treated mice
(23). However, the effect of AA on
obesity-induced impaired spermatogenesis has not yet been reported.
The aim of the present study was to investigate whether AA is able
to protect against HFD-induced defective spermatogenesis
function.
Materials and methods
Reagents
AA (purity, 97%; cat. no. 546712) was purchased from
Sigma-Aldrich (Merck KGaA, Darmstadt, Germany). Rabbit anti-B-cell
lymphoma (Bcl)-xl antibody (cat. no. ab32370), rabbit anti-Fas
antibody (cat. no. ab82419, and anti-GAPDH antibody (cat. no.
ab8245) were obtained from Abcam (Cambridge, UK). Anti-Bcl-2
antibody (cat. no. 2870) and anti-Bcl-2-associated X protein (Bax)
antibody (cat. no. 2722) were purchased from Cell Signaling
Technology, Inc. (Danvers, MA, USA). TUNEL kits (cat. no.
11684817910) were purchased from Roche Applied Science (Penzberg,
Germany).
Animal treatment
All experiments in the present study were performed
in compliance with National Institutes of Health Guide for the Care
and Use of Laboratory Animals and were approved by the Animal Care
and Use Committees of Puai Hospital of Huazhong University of
Science and Technology (Wuhan, China). A total of 24 adult male
Sprague-Dawley rats (180–200 g, 8–9 weeks) were obtained from the
Institute of Laboratory Animal Science, Chinese Academy of Medical
Sciences (Beijing, China). All rats were housed with a 12-h
light/dark cycle at 20–25°C and 50±5% humidity, with ad
libitum access to food and water. Rats were randomly divided
into three groups: Control group (n=8), HFD group (n=8) and the AA
+ HFD (n=8) group. Rats in the control group were fed with a normal
diet, whereas the other rats were fed with an HFD (protein, 18.1%;
fat, 61.6%; carbohydrates, 20.3%) for 12 weeks to induce obesity.
AA was dissolved in 1% CMC-Na as a vehicle for in vivo
experiments. Rats in the AA + HFD group were orally administered
with 50 mg/kg AA once per day for 12 weeks, and the other two
groups received the same volume of vehicle as control. At the end
of the study period, rats were euthanized with an overdose of
sodium pentobarbital (200 mg/kg, Sigma-Aldrich; Merck KGaA) and
blood samples were harvested from the abdominal aorta for further
analysis. Finally, rats were sacrificed via cervical dislocation
and testes and adipose tissues were harvested.
Histological analysis
Testis samples were fixed in 4% paraformaldehyde at
room temperature for 24 h, dehydrated and embedded in paraffin.
Tissues were cut into 5-µm sections for further analysis. The
sections were stained with hematoxylin and eosin at room
temperature and observed under a light microscope (magnification,
×400; E100; Nikon Corporation, Tokyo, Japan). In each group, 30
fields in 6 rats were randomly selected and the number of
spermatogonia, Leydig cells and Sertoli cells were calculated using
Image-Pro Plus 6.0 (Media Cybernetics, Inc., Rockville, MD,
USA).
Blood pressure
Prior to sacrifice, rats were anesthetized using
0.5% isoflurane (Sigma-Aldrich; Merck KGaA), and a microtip
catheter transducer (SPR-839; Millar, Inc., Houston, TX) was
inserted into the right carotid artery and left ventricle to detect
the systolic blood pressure according to the manufacturers
protocol.
Hormone detection
Serum was collected from the tail vein of animals
and fasting insulin was determined using a rat insulin ELISA Kit
(cat. no. EZRMI-13K, EMD Millipore, Billerica, MA, USA) 3 days
prior to sacrifice. Sex hormones were detected using kits for
estradiol (E2; cat. no. E-EL-R0065c), testosterone (T; cat. no.
E-EL-R0072c), follicle stimulating hormone (FSH; cat. no.
E-EL-R0391c) and luteinizing hormone (LH; cat. no. E-EL-R0026c)
purchased from Elabscience Biotechnology Co., Ltd. (Wuhan, China)
according to the manufacturers protocol. The homeostasis model
assessment of insulin resistance (HOMA-IR) was calculated as
previously described (24).
Semen analysis
Isolated epididymides were immediately placed in
Ringers solution (Wuhan Servicebio Technology Co., Ltd., Wuhan,
China) and cut into pieces. The concentration, viability and
motility of sperm were determined as previously described (25). The sperm gradually left the
epididymis and semen samples were carefully collected. The number
of sperm was counted using a hemocytometer (AMQAX1000, Thermo
Fisher Scientific, Inc., Waltham, MA, USA) and the concentrations
were calculated according to the manufacturers protocol.
Eosin-nigrosin staining solution was used to determine sperm
viability at room temperature for 5 min, and light microscopy
(magnification, ×400) was used to observe the spermatozoa. Using
this staining live spermatozoa are white in color, whereas dead
spermatozoa are pink or red (26).
Sperm motility was detected by computer-assisted sperm analysis
(CASA). Sperm was incubated in Ringers solution at room temperature
for 30 min and subsequently placed in CASA assay chambers (Hamilton
Thorne Research, Beverly, MA, USA). Sperm tracks (1.5 sec, 30
frames) were captured (frequency, 60 Hz) and further analyzed by
HTM-IVOS Sperm Analyzer software (version 12.2L; Hamilton Thorne
Research) (27).
Immunohistochemistry and TUNEL
staining
Immunohistochemistry was performed to detect the
expression of Fas and Bcl-xl. Testis tissue sections were
deparaffinized and boiled in sodium citrate buffer (pH=7.0, 5 min,
MXB Biotechnologies, Fuzhou, China) for antigen retrieval after
rehydration in a descending alcohol series. Sections were
subsequently incubated with primary antibodies (anti-Fas, 1:1,000;
anti-Bcl-xl, 1:500) at 4°C overnight after eliminating the internal
peroxidase activity using 3% hydrogen peroxide incubation at room
temperature for 20 min. Sections were subsequently incubated with
the secondary antibody (EnVision™+/HRP reagent; 1:100; cat. no.
GK500610A, Gene Technology Co., Ltd., Shanghai, China) at 37°C for
30 min. Sections were incubated with diaminobenzidine at room
temperature for 2 min and observed under a light microscope
(magnification, ×100 and ×400; E100; Nikon Corporation). A total of
30 fields were randomly selected in 6 rats from each group and the
expression of Fas and Bcl-xl were identified using integrated
optical density. Apoptosis was detected using the TUNEL kit
according to the manufacturers protocol. A total of 100 cells were
randomly selected in each group and the number of positive cells
was calculated manually.
Western blot analysis and reverse
transcription-quantitative polymerase chain analysis (RT-qPCR)
Total proteins from fresh testis tissues were
isolated using radioimmunoprecipitation assay lysis buffer (Wuhan
Servicebio Technology Co., Ltd.). Protein concentrations were
determined using a bicinchoninic acid assay kit (cat. no. 23225;
Thermo Fisher Scientific, Inc.). Proteins (50 µg) were separated by
10% SDS-PAGE and transferred to a polyvinylidene fluoride membrane.
The membrane was blocked with 5% non-fat milk at room temperature
for 1 h, and subsequently incubated overnight at 4°C with the
following primary antibodies: Anti-GAPDH antibody, anti-Bax
antibody and anti-Bcl-2 antibody (all 1:1,000). The membrane was
subsequently incubated with IRDye 800CW-conjugated secondary
antibody (1;10,000, cat. no. LI 926-32211; LI-COR Biosciences,
Lincoln, NE, USA) at room temperature for 1 h. Finally, the
membrane was scanned using a two-color infrared imaging system
(Odyssey; LI-COR Biosciences) and protein expression levels were
normalized to GAPDH.
RNA was isolated from tissues using an RNeasy mini
kit (Qiagen AB, Sollentuna, Sweden) and RT-qPCR was performed with
a Bio-Rad iCycler (Bio-Rad Laboratories, Inc., Hercules, CA, USA)
using PrimeScriptTM RT reagent kit with gDNA Eraser (RR047A, Takara
Bio, Inc., Otsu, Japan). SYBR Premix Ex TaqTM II was obtained from
Takara Bio, Inc. (DRR820A). The temperature protocol for reverse
transcription was: 37°C for 15 min, 85°C for 5 sec. The
thermocycling conditions for PCR were: Initial denaturation at 95°C
for 30 sec; 40 cycles of 95°C for 5 sec and 60°C for 45 sec;
dissociation at 95°C for 15 sec and 60°C for 30 sec. The primers
used were as follows: Bax, forward 5′-ATC CAG GAT CGA GCA GGG AGG
ATG G-3 and reverse, 5′-TGC CCG CCT ACT TCA ACG A-3; Bcl-2, forward
5′-CTT CCA GCC TGA GAG CAA CC-3 and reverse 5′-CAT CCC AGC CTC CGT
TAT CC-3; GAP DH, forward 5′GAC ATG CCG CCT GGA GAA AC-3 and
reverse 5′AGCC CAG GAT GCC CTT TAG T-3. Relative mRNA expression
levels were analyzed using the 2−∆∆Cq method (28). The mRNA levels were normalized to
GAPDH.
Data analysis
Results in each group are expressed as the mean +
standard deviation. All statistical tests were conducted using SPSS
19.0 (IBM Corp., Armonk, NY, USA). Multiple group comparisons were
made using one-way ANOVA followed by a post hoc Tukeys test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
AA improves cardiometabolic profile in
rats subjected to HFD
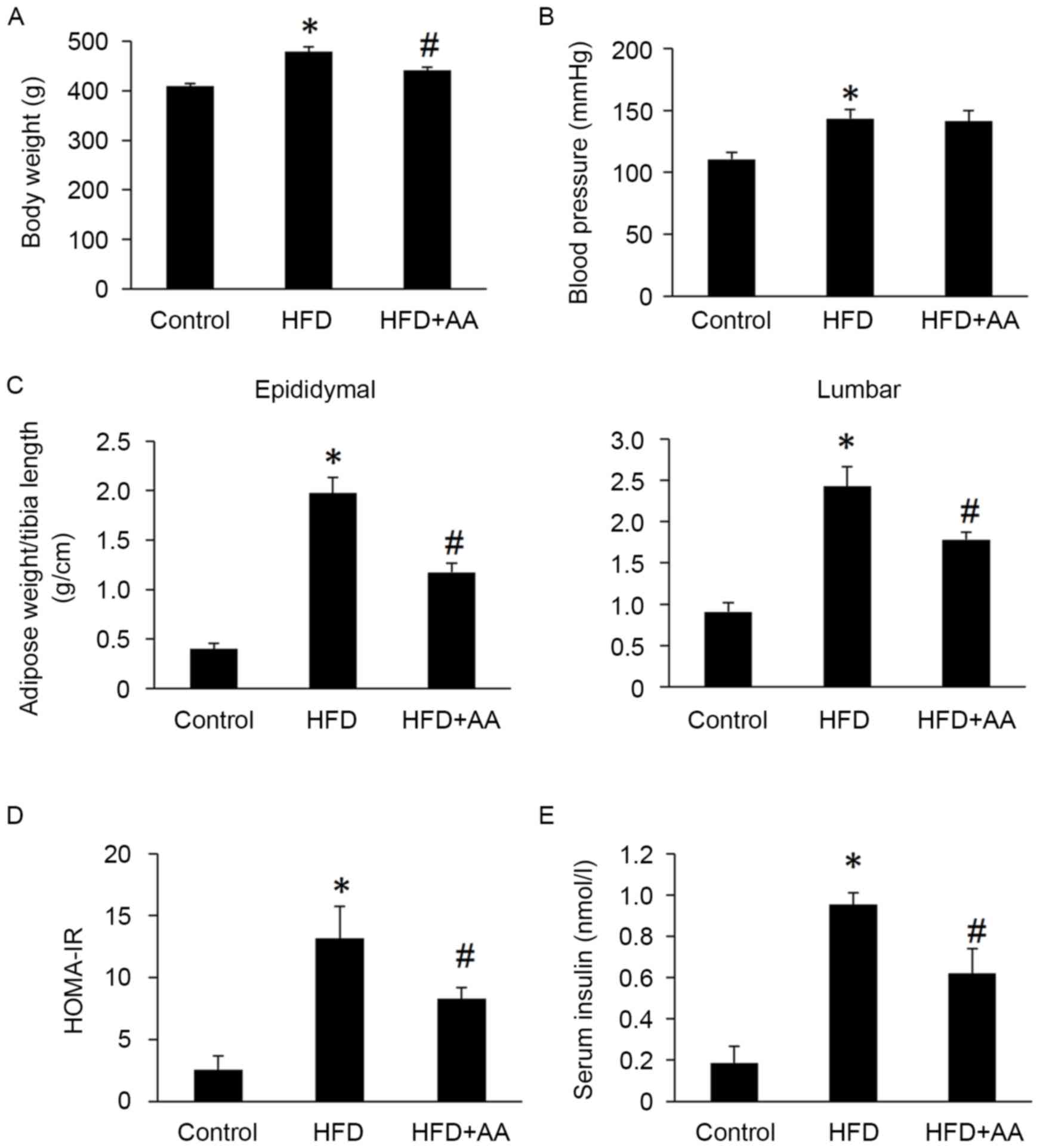
Body weight, HOMA-IR, serum insulin, epididymal and
lumbar adipose tissues weights were all significantly increased in
rats with HFD compared with control rats (P<0.05; Fig. 1). However, AA treatment significantly
attenuated these HFD-induced increases (P<0.05; Fig. 1). Rats in the HFD also had
significantly increased systolic blood pressure compared with the
control group (P<0.05); however, no significant difference was
observed between the HFD and AA + HFD group (Fig. 1).
AA treatment improves pathological
changes of testes induced by HFD
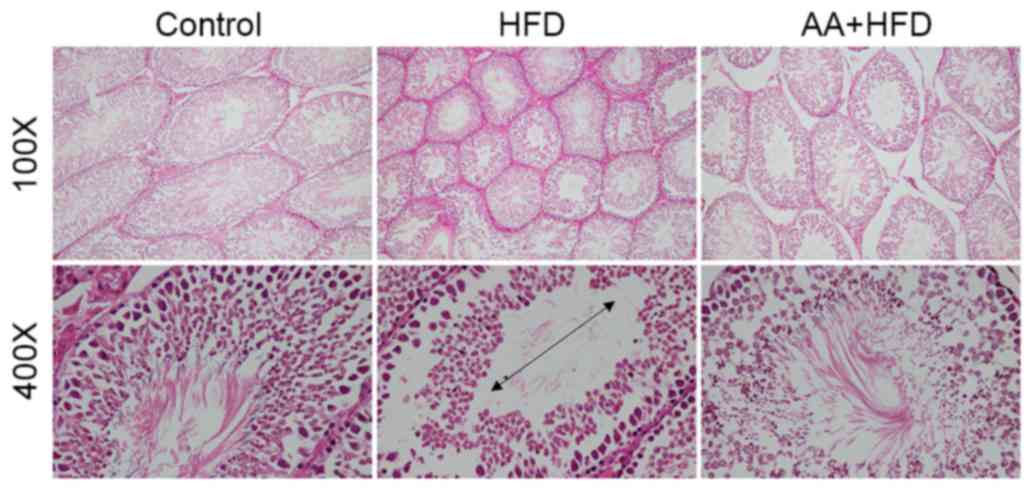
Following 12 weeks of HFD, rats in the HFD group had
significantly decreased testis weight and testis weight/body weight
compared with the control group (P<0.05; Table I), which was also confirmed by
histological analysis (Fig. 2).
Atrophic seminiferous tubules with smaller diameters were also
observed in the testis of rats with HFD (Fig. 2). HFD also resulted in a significant
reduction in spermatogonia, Leydig cells and Sertoli cells compared
with the control group (P<0.05; Table
I). Sperm concentration, sperm viability and motility were
significantly decreased in rats subjected to an HFD diet compared
with control rats (P<0.05; Table
I). AA treatment significantly attenuated the decreased testis
weight, testis/body weight, spermatogonia, Leydig cells and Sertoli
cells (P<0.05; Table I), and
markedly improved the HFD-induced atrophy of seminiferous tubules
(Fig. 2).
 | Table I.Effects of AA on testis weight, germ
cell count and sperm quality. |
Table I.
Effects of AA on testis weight, germ
cell count and sperm quality.
| Parameter | Control | HFD | AA + HFD |
|---|
| Testis weight
(g) |
3.32±0.13 |
2.64±0.09a |
2.86±0.09b |
| Testis weight/body
weight (g/kg) |
8.11±0.39 |
5.52±0.14a |
6.54±0.21b |
| Spermatogonia
(number/field) |
25.26±2.18 |
15.52±1.79a |
23.10±1.39b |
| Leydig cells
(number/field) |
8.21±0.15 |
4.53±0.33a |
5.21±0.08b |
| Sertoli cells
(number/field) |
9.21±0.14 |
5.34±0.24a |
7.16±0.13b |
| Sperm concentration
(×106/ml) |
57.61±4.21 |
45.36±3.17a |
53.11±2.54b |
| Sperm viability
(%) |
93.39±2.65 |
85.31±1.70a |
92.02±1.33b |
| Sperm motility
(%) |
75.07±1.60 |
66.28±1.88a |
71.10±1.48b |
AA treatment attenuates the
HFD-induced abnormal serum sexual hormone levels
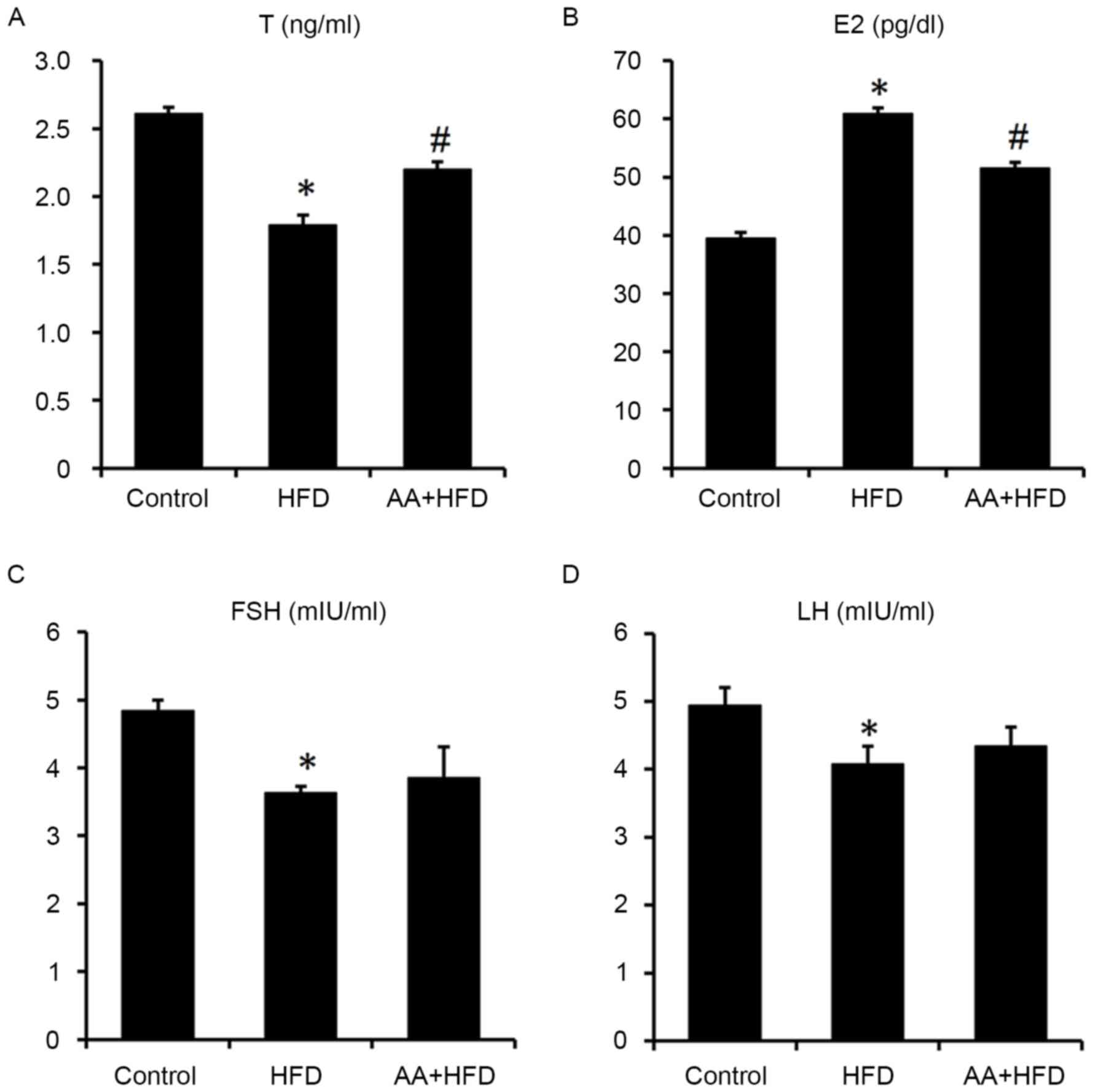
Serum E2 levels were significantly increased and T
levels were significantly decreased in the HFD compared with the
control group (P<0.05; Fig. 3A and
B). However, AA treatment for 12 weeks significantly
ameliorated the HFD-induced abnormal serum sexual hormone levels
(Fig. 3A and B). Serum FSH and LH
were both significantly reduced in HFD rats compared with the
control group (P<0.05), whereas no significant difference was
observed between the HFD and AA + HFD groups (Fig. 3C and D).
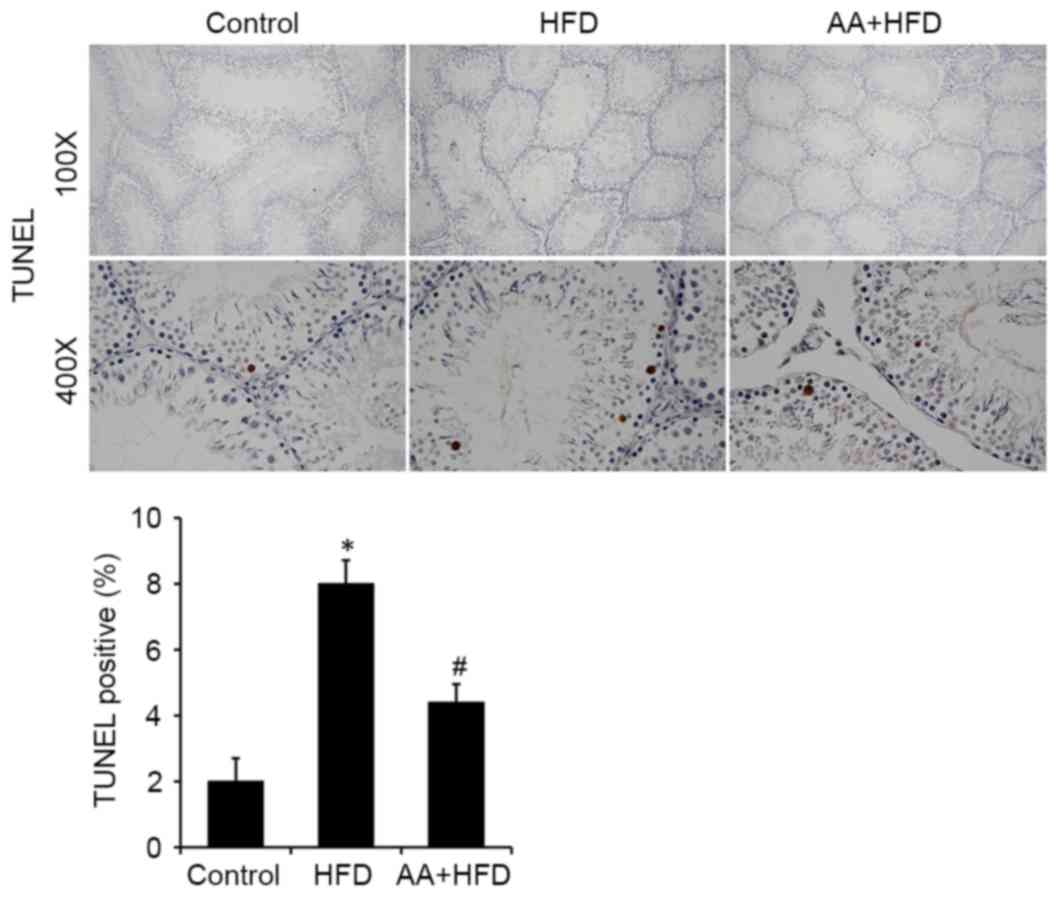
AA suppresses apoptosis in the testis
of rats fed with HFD
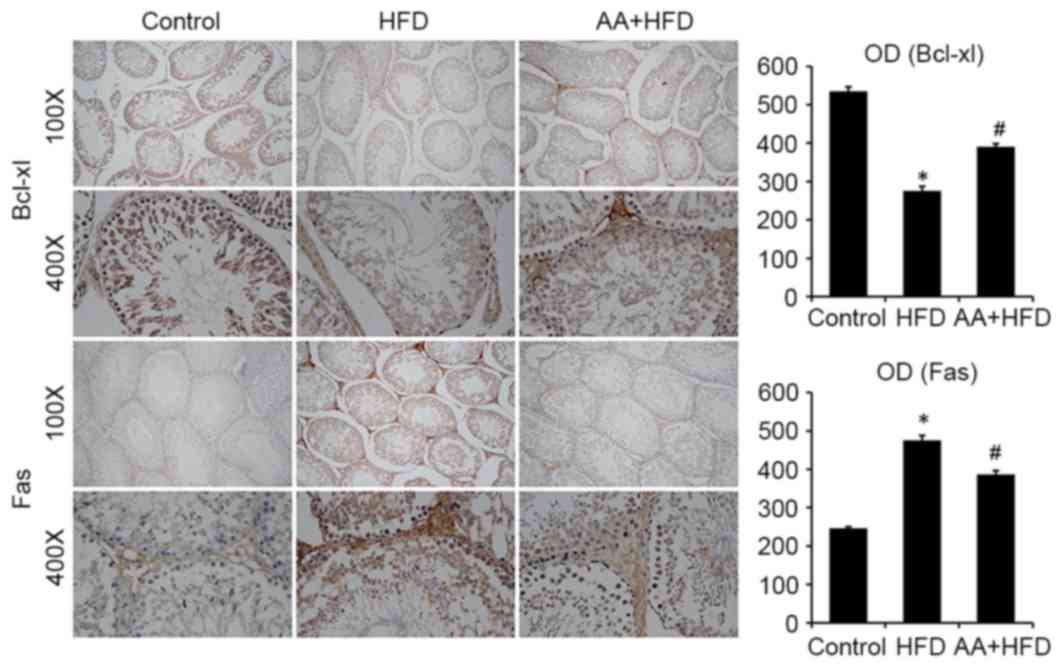
Immunohistological analysis revealed that the
protein level of Bcl-xl was significantly decreased in the HFD rats
compared with the control group (P<0.05) and that AA treatment
significantly ameliorated this effect (P<0.05; Fig. 4). Fas was also significantly
upregulated in the HFD group compared with the control rats
(P<0.05) and this effect was significantly ameliorated with AA
treatment (P<0.05; Fig. 4). TUNEL
analysis was used to investigate the apoptotic rate of germ cells
in the testis. The results revealed that rats in the HFD group had
a significantly higher rate of apoptosis compared with the control
rats (P<0.05), and that AA significantly inhibited HFD-induced
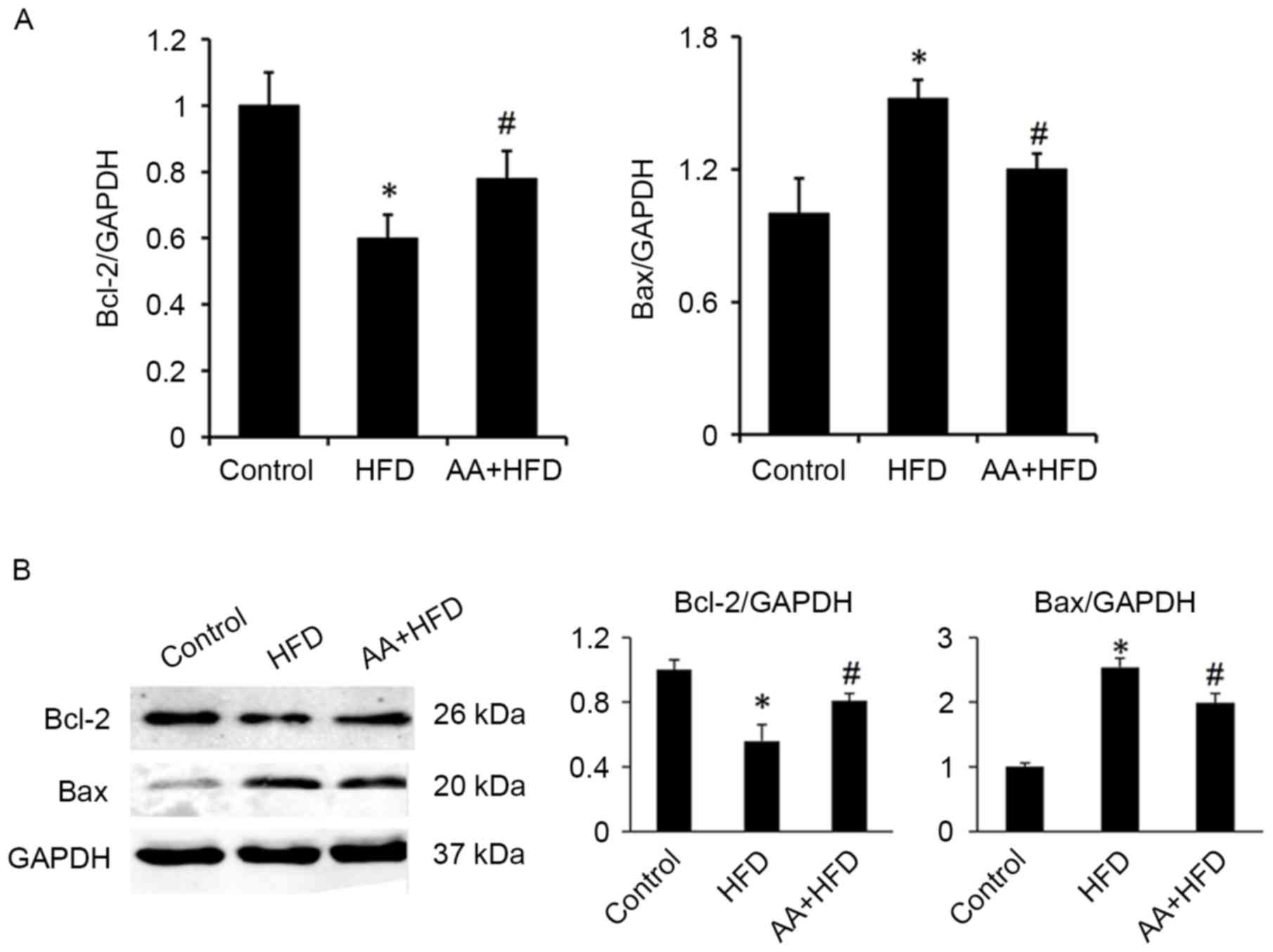
testicular cell apoptosis (P<0.05; Fig. 5). These results were corroborated by
subsequent analysis of mRNA and protein levels, which indicated
that HFD induced a significant downregulation in Bcl-2 and a
significant increase in Bax expression compared with the control
group (P<0.05; Fig. 6). Treatment
with AA, however, significantly ameliorated these effects, inducing
a significant increase in Bcl-2 and decrease in Bax compared with
the HFD group (P<0.05; Fig.
6).
Discussion
The number of worldwide overweight individuals has
grown rapidly, resulting in an escalation of obesity-associated
health problems including infertility (29). As a result, there is a greater need
to develop pharmacological agents for and to explore the novel and
specific regulators of obesity-associated infertility. The results
of the present study indicate that AA may attenuate HFD-induced
impaired spermatogenesis. AA was also demonstrated to ameliorate
endocrine disorders and suppress HFD-induced testicular cell
apoptosis.
Endocrine disorders are key features of
spermatogenesis dysfunction (30). T
is able to promote spermatogenesis via intracellular signaling
pathways (31). A recent study
indicated that metformin-induced T level increases were able to
improve reproductive function in obese male rats (17). Consistent with this, the present
study demonstrated that AA upregulates the level of T and reduces
the level of E2, which suggests that improved sex hormone levels
may contribute to the protective effects of AA. AA had no
significant effect on FSH and LH levels, which indicates that it
does not affect pituitary hormones.
It is known that spermatogenesis is a complex
process that relies on coordinated cell proliferation and apoptosis
(32). Excessive cell apoptosis is
reported to be a prevalent phenomenon in defective spermatogenesis
(33); therefore, inhibiting
excessive cell apoptosis and reconstructing the balance between
cell proliferation and apoptosis may be an effective treatment for
defective spermatogenesis. In view of the antiapoptotic properties
of AA (22,23), it was hypothesized that AA may
suppress HFD-induced apoptosis in the testes. The results of the
present study revealed that AA significantly inhibits testicular
cell apoptosis, which suggests that apoptosis may be one of the
underlying mechanisms by which AA protects against HFD-induced
defective spermatogenesis. Conversely, it has previously been
reported that AA induces tumor cell apoptosis (34,35). The
reason for these incompatible results may be that apoptosis serves
different roles in different pathological processes.
The precise mechanisms that mediate the
antiapoptotic effects of AA remain to be elucidated. A recent study
indicated that AA is able to activate AMP-activated protein kinase
α (19), which has been demonstrated
to be a negative regulator of apoptosis (36,37). AA
has also been reported to suppress inflammation and oxidative
injury in human bronchial epithelial cells (38), which is associated with apoptosis.
Further study is required to determine the precise mechanisms
underlying the protective effects of AA.
In conclusion, the results of the present study
demonstrated that AA is able to attenuate HFD-induced
spermatogenesis dysfunction via inhibiting excessive apoptosis.
These findings provide theoretical evidence for the use of AA as a
treatment for obesity-associated infertility.
References
|
1
|
Botelho F, Figueiredo L, Leite R, Carvalho
A, Tomada N and Vendeira P: Predictive factors of a successful
testicular biopsy and subsequent clinical pregnancy. Andrologia.
44:237–242. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ferlin A, Arredi B and Foresta C: Genetic
causes of male infertility. Reprod Toxicol. 22:133–141. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Adamson GD and Baker VL: Subfertility:
Causes, treatment and outcome. Best Pract Res Clin Obstet Gynaecol.
17:169–185. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Barnett R: Obesity. Lancet. 366:9842005.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Katib A: Mechanisms linking obesity to
male infertility. Cent European J Urol. 68:79–85. 2015.PubMed/NCBI
|
|
6
|
Hammoud AO, Meikle AW, Reis LO, Gibson M,
Peterson CM and Carrell DT: Obesity and male infertility: A
practical approach. Semin Reprod Med. 30:486–495. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bellver J, Melo MA, Bosch E, Serra V,
Remohi J and Pellicer A: Obesity and poor reproductive outcome: The
potential role of the endometrium. Fertil Steril. 88:446–451. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hammoud AO, Wilde N, Gibson M, Parks A,
Carrell DT and Meikle AW: Male obesity and alteration in sperm
parameters. Fertil Steril. 90:2222–2225. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lee J, Richburg JH, Younkin SC and
Boekelheide K: The Fas system is a key regulator of germ cell
apoptosis in the testis. Endocrinology. 138:2081–2088. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sinha HA and Swerdloff RS: Hormonal and
genetic control of germ cell apoptosis in the testis. Rev Reprod.
4:38–47. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tapanainen JS, Tilly JL, Vihko KK and
Hsueh AJ: Hormonal control of apoptotic cell death in the testis:
Gonadotropins and androgens as testicular cell survival factors.
Mol Endocrinol. 7:643–650. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Aitken RJ and Baker MA: Causes and
consequences of apoptosis in spermatozoa; contributions to
infertility and impacts on development. Int J Dev Biol. 57:265–272.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
MacFarlane M and Williams AC: Apoptosis
and disease: A life or death decision. EMBO Rep. 5:674–678. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Barroso G, Morshedi M and Oehninger S:
Analysis of DNA fragmentation, plasma membrane translocation of
phosphatidylserine and oxidative stress in human spermatozoa. Hum
Reprod. 15:1338–1344. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang H, Zhao R, Guo C, Jiang S, Yang J, Xu
Y, Liu Y, Fan L, Xiong W, Ma J, et al: Knockout of BRD7 results in
impaired spermatogenesis and male infertility. Sci Rep.
6:217762016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bhat GK, Sea TL, Olatinwo MO, Simorangkir
D, Ford GD, Ford BD and Mann DR: Influence of a leptin deficiency
on testicular morphology, germ cell apoptosis and expression levels
of apoptosis-related genes in the mouse. J Androl. 27:302–310.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yan WJ, Mu Y, Yu N, Yi TL, Zhang Y, Pang
XL, Cheng D and Yang J: Protective effects of metformin on
reproductive function in obese male rats induced by high-fat diet.
J Assist Reprod Genet. 32:1097–1104. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bonte F, Dumas M, Chaudagne C and Meybeck
A: Influence of asiatic acid, madecassic acid and asiaticoside on
human collagen I synthesis. Planta Med. 60:133–135. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ma ZG, Dai J, Wei WY, Zhang WB, Xu SC,
Liao HH, Yang Z and Tang QZ: Asiatic acid protects against cardiac
hypertrophy through activating AMPKα signalling pathway. Int J Biol
Sci. 12:861–871. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang X, Lu Q, Yu DS, Chen YP, Shang J,
Zhang LY, Sun HB and Liu J: Asiatic acid mitigates hyperglycemia
and reduces islet fibrosis in Goto-Kakizaki rat, a spontaneous type
2 diabetic animal model. Chin J Nat Med. 13:529–534.
2015.PubMed/NCBI
|
|
21
|
Yan SL, Yang HT, Lee YJ, Lin CC, Chang MH
and Yin MC: Asiatic acid ameliorates hepatic lipid accumulation and
insulin resistance in mice consuming a high-fat diet. J Agric Food
Chem. 62:4625–4631. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xiong Y, Ding H, Xu M and Gao J:
Protective effects of asiatic acid on rotenone- or
H2O2-induced injury in SH-SY5Y cells.
Neurochem Res. 34:746–754. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chao PC, Lee HL and Yin MC: Asiatic acid
attenuated apoptotic and inflammatory stress in the striatum of
MPTP-treated mice. Food Funct. 7:1999–2005. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Matthews DR, Hosker JP, Rudenski AS,
Naylor BA, Treacher DF and Turner RC: Homeostasis model assessment:
Insulin resistance and beta-cell function from fasting plasma
glucose and insulin concentrations in man. Diabetologia.
28:412–419. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ghanayem BI, Bai R, Kissling GE, Travlos G
and Hoffler U: Diet-induced obesity in male mice is associated with
reduced fertility and potentiation of acrylamide-induced
reproductive toxicity. Biol Reprod. 82:96–104. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kokilavani P, Suriyakalaa U, Elumalai P,
Abirami B, Ramachandran R, Sankarganesh A and Achiraman S:
Antioxidant mediated ameliorative steroidogenesis by Commelina
benghalensis L. and Cissus quadrangularis L. against quinalphos
induced male reproductive toxicity. Pestic Biochem Physiol.
109:18–33. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wei ZT, Lu XL, Zhang G, Yu J, Li H, Jia
GH, Li JT and Zhang JM: The long-term effects of superovulation on
fertility and sexual behavior of male offspring in mice. J Assist
Reprod Genet. 31:555–560. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-tie quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Pasquali R, Patton L and Gambineri A:
Obesity and infertility. Curr Opin Endocrinol Diabetes Obes.
14:482–487. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Plant TM and Marshall GR: The functional
significance of FSH in spermatogenesis and the control of its
secretion in male primates. Endocr Rev. 22:764–786. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chang C, Chen YT, Yeh SD, Xu Q, Wang RS,
Guillou F, Lardy H and Yeh S: Infertility with defective
spermatogenesis and hypotestosteronemia in male mice lacking the
androgen receptor in sertoli cells. Proc Natl Acad Sci USA. 101:pp.
6876–6881. 2004; View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sakkas D, Seli E, Bizzaro D, Tarozzi N and
Manicardi GC: Abnormal spermatozoa in the ejaculate: Abortive
apoptosis and faulty nuclear remodelling during spermatogenesis.
Reprod Biomed Online. 7:428–432. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sukhotnik I and Nativ O, Roitburt A, Bejar
D, Coran AG, Mogilner JG and Nativ O: Methotrexate induces germ
cell apoptosis and impairs spermatogenesis in a rat. Pediatr Surg
Int. 29:179–184. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gurfinkel DM, Chow S, Hurren R, Gronda M,
Henderson C, Berube C, Hedley DW and Schimmer AD: Disruption of the
endoplasmic reticulum and increases in cytoplasmic calcium are
early events in cell death induced by the natural triterpenoid
Asiatic acid. Apoptosis. 11:1463–1471. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kavitha CV, Jain AK, Agarwal C, Pierce A,
Keating A, Huber KM, Serkova NJ, Wempe MF, Agarwal R and Deep G:
Asiatic acid induces endoplasmic reticulum stress and apoptotic
death in glioblastoma multiforme cells both in vitro and in vivo.
Mol Carcinog. 54:1417–1429. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Qi D and Young LH: AMPK: Energy sensor and
survival mechanism in the ischemic heart. Trends Endocrinol Metab.
26:422–429. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ma ZG, Yuan YP, Xu SC, Wei WY, Xu CR,
Zhang X, Wu QQ, Liao HH, Ni J and Tang QZ: CTRP3 attenuates cardiac
dysfunction, inflammation, oxidative stress and cell death in
diabetic cardiomyopathy in rats. Diabetologia. 60:1126–1137. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Tsao SM and Yin MC: Antioxidative and
antiinflammatory activities of asiatic acid, glycyrrhizic acid and
oleanolic acid in human bronchial epithelial cells. J Agric Food
Chem. 63:3196–3204. 2015. View Article : Google Scholar : PubMed/NCBI
|