Introduction
Adriamycin (ADR; doxorubicin hydrochloride), is an
anthracycline antibiotic that is clinically used as an
antineoplastic agent. The clinical efficacy of ADR, particularly
for long-term treatment, is limited by the induction of hepatic and
cardiac toxicities that are frequently lethal (1). Specific studies have proposed that
ADR-induced toxicity is possibly mediated by oxidative damage to
cellular components, including membrane lipids in the plasma
membranes and mitochondria (2).
When the concentration of generated reactive oxygen species exceeds
the antioxidant capability of the cell, cellular oxidative damage
occurs. Oxygen-derived free radicals and lipid peroxidation play a
critical role in the pathogenesis of various liver diseases,
including hepatic fibrosis (3,4).
Doxorubicin has been shown to cause an imbalance between free
oxygen radicals and antioxidant enzymes, resulting in tissue injury
(5,6). Doxorubicin induces toxic effects on
the liver by increasing the levels of superoxide dismutase,
catalase and glutathione peroxidase enzymes in liver tissue
(7,8). The modulation of these mediators has
been indicated to prevent doxorubicin-induced toxicities in various
organs (9).
Diverse antioxidants have been shown to prevent
ADR-induced hepatotoxicity in rats (10,11).
A unique nutrient formulation (NM), containing primarily ascorbic
acid, lysine, proline, N-acetyl cysteine and green tea
extract, has previously been shown to exhibit a broad spectrum of
pharmacological, therapeutic, cardiovascular and chemoprotective
properties (12). In previous
studies, it was found that NM significantly inhibited
acetaminophen- and carbon tetrachloride-induced hepatic and renal
damage (13,14).
In the present study, the in vivo effects of
the NM diet were examined in mice treated with ADR, focusing on
renal and hepatic enzyme levels.
Materials and methods
Materials
ADR powder, obtained from Sigma-Aldrich (St. Louis,
MO, USA), was diluted in warm saline (pH 7.4) to 20 mg/ml. The
stock solution of the NM was composed of the following in the
quantities indicated: 700 mg vitamin C (as ascorbic acid and as Mg,
Ca and palmitate ascorbate); 1,000 mg L-lysine; 750 mg L-proline;
500 mg L-arginine; 200 mg N-acetyl cysteine; 1,000 mg
standardized green tea extract (80% polyphenol); 30 μg selenium; 2
mg copper; 1 mg manganese; and 50 mg quercetin.
Animals
Male BALB/c mice free of murine viruses, bacteria
and parasites and ~6 weeks of age on arrival, were purchased from
Simonsen Laboratories (Gilroy, CA, USA). The mice were maintained
in microisolator cages under pathogen-free conditions on a 12-h
light/dark schedule for one week. Animals were cared for in
accordance with the institutional guidelines for the care and use
of experimental animals.
Experimental design
Following one week of isolation, mice were divided
into four groups (A–D) with six animals per group. Mice in groups A
and C were fed a regular Purina mouse chow diet (Laboratory Rodent
Diet 5001 from Purina Mills, LLC, purchased from Newco Distributing
Inc., Rancho Cucamonga, CA, USA) for three weeks, while those in
groups B and D were fed a regular mouse chow diet supplemented with
1% (w/w) NM during that period. During the study, the mice
consumed, on the average, 4 g of their respective diets per day.
Thus, the supplemented mice received ~20 mg NM per day. After three
weeks, the mice in groups C and D received a single injection of 20
mg ADR/kg intraperitoneally, while those in groups A and B received
saline alone. After 24 h, mice were sacrificed, blood samples were
obtained by cardiac puncture, serum was collected for clinical
chemistry and livers, kidneys, hearts and lungs were excised and
weighed.
Statistical analysis
Results are expressed as mean + SD for each group.
Data was analyzed by independent sample t tests using MedCalc
Software (Ostend, Belgium). P<0.05 was considered to indicate a
statistically significant difference.
Results
Renal toxicity
ADR-induced renal toxicity in BALB/c male mice was
measured by renal serum enzyme levels. These changes were protected
by the NM 1% diet.
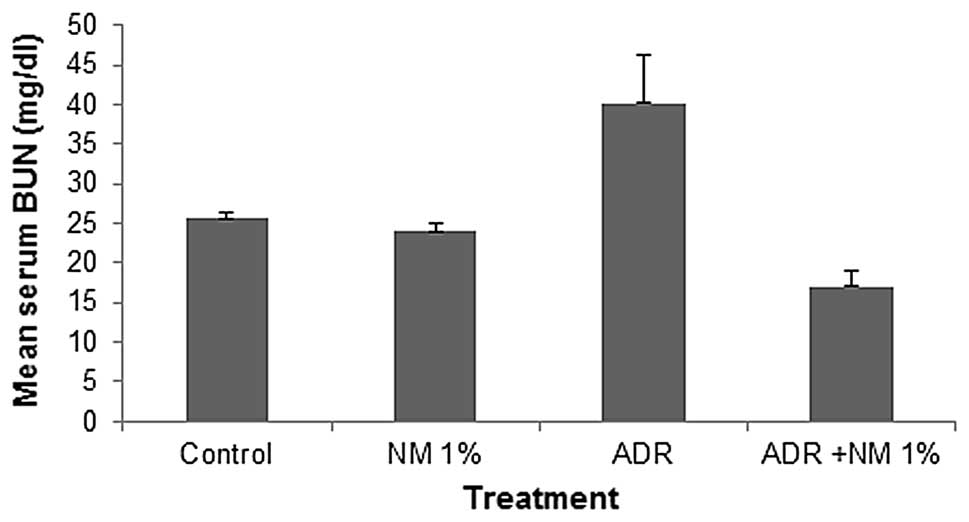
Mean serum blood urea nitrogen (BUN)
In the untreated BALB/c mice that were fed the
supplemented NM 1% diet, the mean serum BUN level was 93% of that
in the control diet mice. In the mice fed the control diet, the
administration of ADR increased the serum BUN level by 157% of the
values in the saline-treated controls (P=0.0001). Of the BALB/c
mice injected with ADR, the mice fed the NM 1% diet showed a mean
serum BUN level that was 42% (P<0.0001) of that in mice fed the
control diet. Furthermore, mice injected with ADR and fed NM 1%
showed a 43.6% (P<0.001) reduction in mean serum BUN level
compared with that in the control mice not injected with ADR
(Fig. 1).
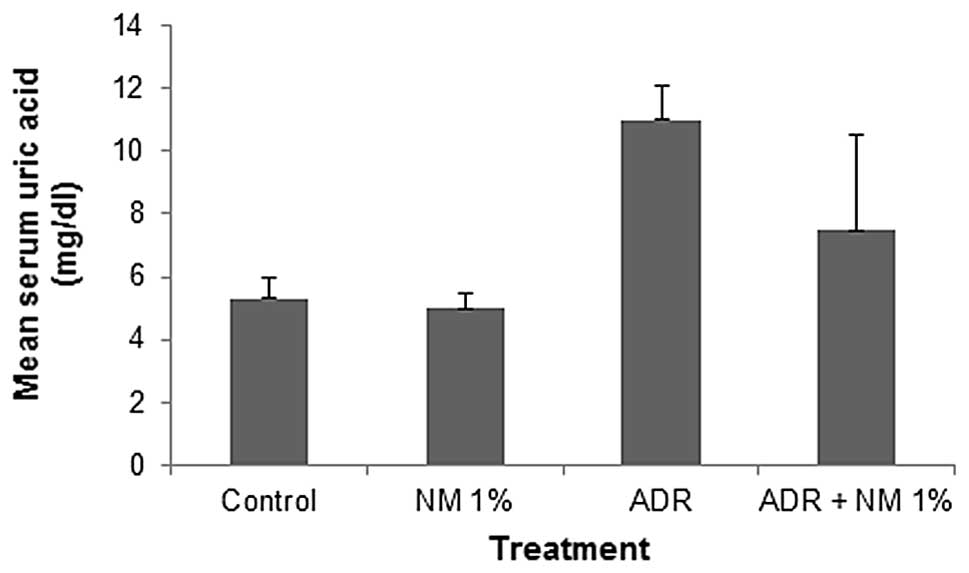
Mean serum uric acid
In the mice fed the control diet, the administration
of ADR increased the mean serum uric acid level by 207%
(P<0.0001) of that in the saline-treated mice. Of the BALB/c
mice injected with ADR, the mice fed the NM 1% diet showed a mean
serum uric acid level that was 68.2% (P=0.02) of that in mice fed
the control diet. Mice injected with ADR and fed NM 1% showed no
significant difference in mean serum uric acid level compared with
those in the mice not injected with ADR on the control or NM 1%
diets (Fig. 2).
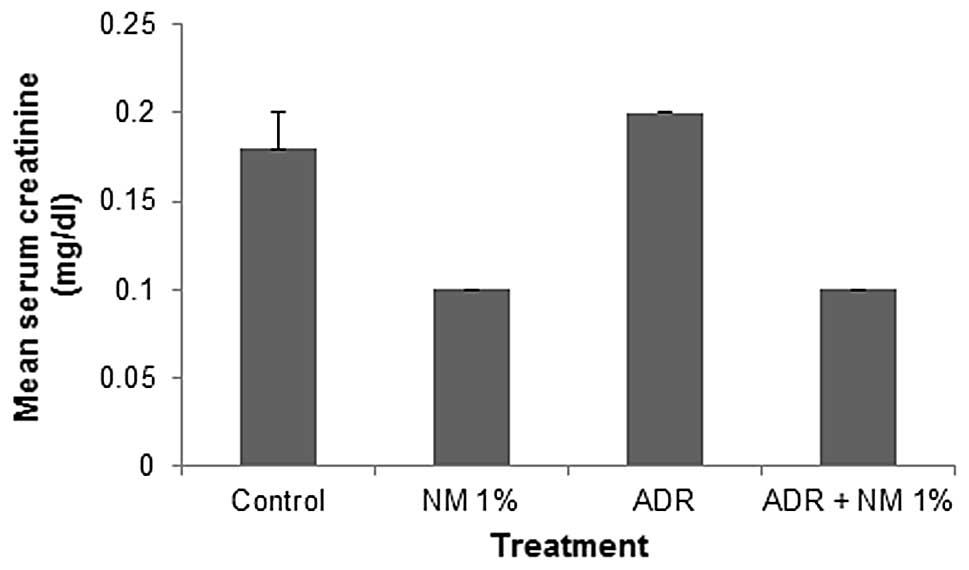
Mean serum creatinine
The mean serum creatinine level in untreated BALB/c
mice fed the supplemented NM 1% diet was 56% (P<0.001) of that
in control diet mice. In the mice fed the control diet, the
administration of ADR increased the mean level of creatinine by
111% (P=0.03) of that in the saline-treated mice. Of the BALB/c
mice injected with ADR, the mice fed the NM 1% diet showed a mean
serum creatinine level that was 50% (P<0.0001) of that in mice
fed the control diet. Furthermore, mice injected with ADR and fed
NM 1% showed a 44.4% (P<0.0001) reduction in mean serum
creatinine level compared with that in control diet mice not
injected with ADR (Fig. 3).
Hepatic toxicity
ADR-induced hepatic toxicity in BALB/c male mice was
measured by hepatic serum enzyme levels. These changes were
protected against by the NM 1% diet.
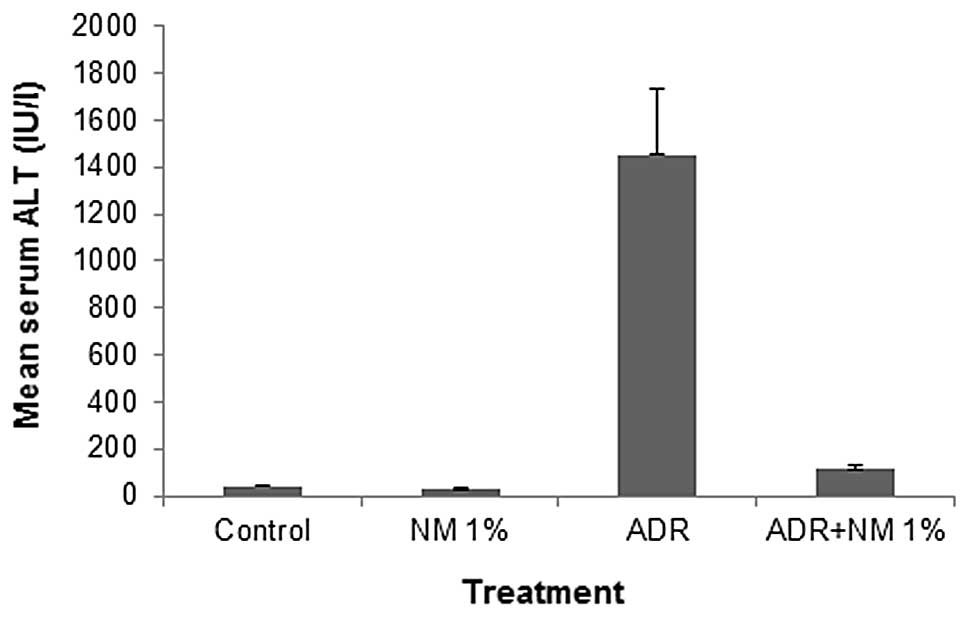
Mean serum alanine aminotransferase
(ALT)
Among the untreated BALB/c mice, mean serum ALT
level decreased by 31% (P=0.004) following NM supplementation. In
the mice fed the control diet, ADR treatment increased the ALT
level by 3,718% (P<0.0001) of the level in the saline-treated
control. Of the BALB/c mice injected with ADR, the mice fed the NM
1% diet showed a mean serum ALT level that was 7.9% (P<0.0001)
of that in mice fed the control diet. Mice injected with ADR and
fed NM 1% showed a mean serum ALT level that was 292% (P<0.001)
of that shown by control mice not injected with ADR (Fig. 4).
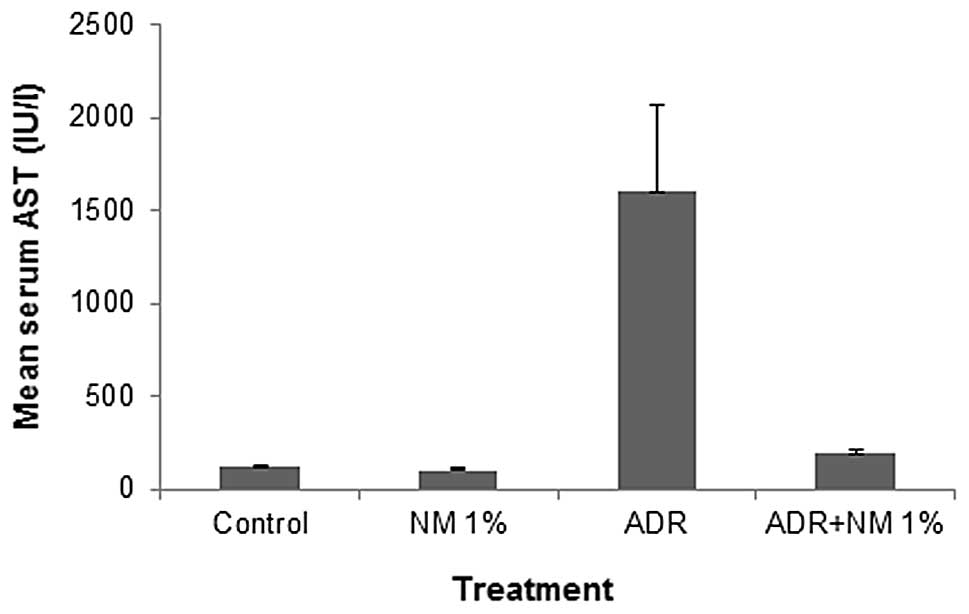
Mean serum aspartate aminotransferase
(AST)
Among the untreated BALB/c mice, the mean serum AST
level decreased by 12% (P=0.03) with NM supplementation. In the
mice fed the control diet, the administration of ADR increased the
mean serum AST level by 1,334% (P<0.0001) of the value in the
saline-treated controls. Of the BALB/c mice injected with ADR, the
mice fed the NM 1% diet showed a mean serum ALT level that was 12%
(P<0.0001) of that in mice fed the control diet. Mice injected
with ADR and fed NM 1% showed a mean serum AST level that was 163%
(P<0.0001) of that shown by control mice not injected with ADR
(Fig. 5).
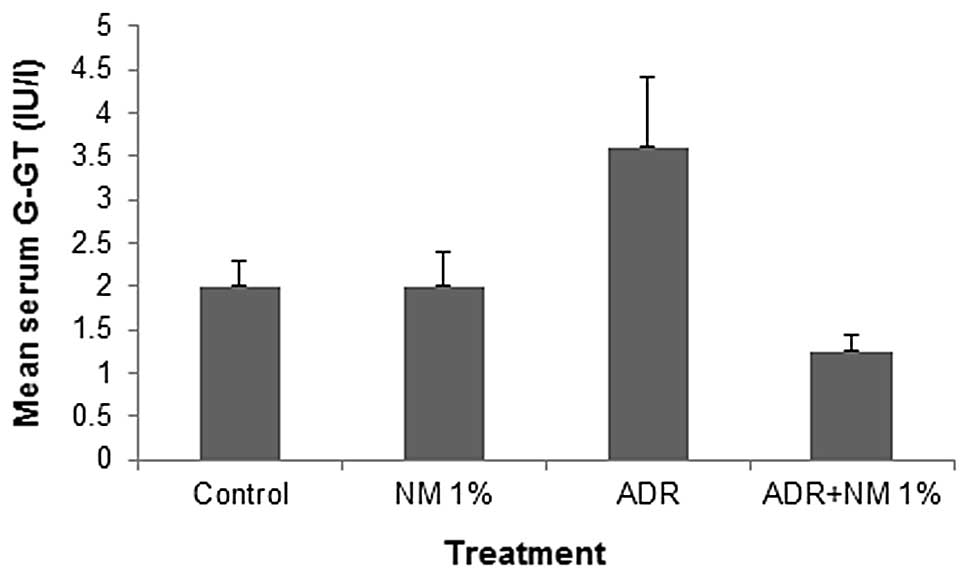
Mean serum γ-glutamyl transferase
(γ-GT)
In the mice fed the control diet, the mean serum
γ-GT level increased by 180% (P=0.001) of the value in the
saline-treated controls. Of the BALB/c mice injected with ADR, the
mice fed the NM 1% diet showed a mean serum γ-GT level that was
34.7% (P<0.0001) of that in mice fed the control diet. Mice
injected with ADR and fed NM 1% showed a mean serum γ-GT level that
was 62.5% (P=0.0005) of that shown by control mice not injected
with ADR. No significant difference was identified between the γ-GT
levels of untreated mice in the two diet groups (Fig. 6).
Vital organ weights
ADR injection into BALB/c mice did not have a
significant effect on the weights of vital organs, as shown in
Table I.
 | Table IEffect of ADR and the NM 1% diet on
the mean weight of vital organs in BALB/c mice. |
Table I
Effect of ADR and the NM 1% diet on
the mean weight of vital organs in BALB/c mice.
| Organ weight (g) |
|---|
|
|
|---|
| Organ | Control diet | NM 1% diet | ADR + control
diet | ADR + NM 1% diet |
|---|
| Liver | 1.4±0.10 | 1.43±0.11 | 1.21±0.14 | 1.18±0.05 |
| Kidney | 0.41±0.04 | 0.47±0.05 | 0.37±0.07 | 0.41±0.03 |
| Lung | 0.14±0.02 | 0.15±0.22 | 0.14±0.01 | 0.14±0.02 |
| Heart | 0.12±0.15 | 0.13±0.01 | 0.10±0.01 | 0.12±0.01 |
| Spleen | 0.11±0.04 | 0.09±0.004 | 0.06±0.04 | 0.06±0.01 |
Final body weights of mice
ADR injection into BALB/c mice reduced the mean
final weight of the mice fed the control diet by 14.5% (P=0.0001)
and the mice fed the NM 1% diet by 5% (P=0.04). In the mice not
treated with ADR, the mean final weights of the control diet and NM
1% diet mice did not significantly differ. The mean initial weight
of all the mice was 24.1±1.4 g, whereas the mean final weights for
the untreated groups were 26.2±0.7 g for control diet mice and
26.4±1.4 g for the NM 1% diet mice. For the ADR-treated groups, the
mean final weights were 22.4±1.2 g for control diet mice and
24.9±1.2 g for NM 1% diet mice (data not shown).
Dietary intake
The mean dietary consumption by the mice in the two
groups of mice fed the NM 1% diet (3.5±0.5 g) was 83.3% (P=0.036)
of that consumed by the mice in the control diet groups (4.2±0.5 g)
(data not shown).
Discussion
The results of the present study demonstrate that
pretreatment for three weeks with a diet supplemented with NM 1%
reduced hepatic and renal damage in male BALB/c mice injected with
a toxic dose (20 mg/kg body weight) of ADR. ADR treatment caused
marked increases in the levels of hepatic serum markers, AST, ALT
and γ-GT, in non-supplemented mice. Supplementation with NM
retained the AST, ALT and γ-GT levels at normal levels. Elevated
ALT and AST levels reflect hepatocellular inflammation, damage and
necrosis, as additional AST and ALT are released into the
bloodstream when a body tissue or organ, including the heart or
liver, is diseased or damaged. The amount of AST in the blood is
directly associated with the extent of tissue damage. Increased
levels of γ-GT are associated with early liver cell damage or
cholestatic disease. These elevated levels of serum indices for
hepatocellular damage have been previously reported in a
doxycycline-induced hepatoxicity model (15).
The ADR-treated mice also showed significantly
increased levels of renal markers, including creatinine, uric acid
and BUN. NM 1% dietary supplementation attenuated the increases in
renal serum marker levels, to provide almost normal levels. The
BUN/creatinine ratio is useful for the differential diagnosis of
acute or chronic renal disease. Reduced renal perfusion, congestive
heart failure or recent onset of urinary tract obstruction is
likely to result in an increase in the BUN/creatinine ratio. The
BUN/creatinine ratio for the untreated control BALB/c mice was 142,
whereas when the mice were treated with ADR, the ratio increased to
200. NM 1% dietary pretreatment in BALB/c mice injected with ADR
reduced the mean BUN/creatinine ratio to 170, a reduction of 15%
compared with that observed in mice receiving the control diet
prior to ADR administration.
Various antioxidants have been shown to prevent
ADR-induced toxicity in vivo. A previous review by
Grandados-Principal et al provided new evidence for the
chemoprevention of doxorubicin toxicity using natural antioxidants,
including vitamin E, vitamin C, coenzyme Q, carotenoids, vitamin A,
flavonoids, polyphenol, resveratrol, antioxidants from virgin olive
oil and selenium. The study offered new insights into the molecular
mechanisms of doxorubicin toxicity with respect to DNA damage, free
radicals and other parameters (16). The NM tested was formulated based
on targeting various physiological processes involved in a wide
spectrum of pathological conditions at the cellular level.
The antioxidant, vitamin C, was shown to protect
against doxorubicin-induced cardiotoxicity and prolong the lives of
mice and guinea pigs without interfering with the anticancer
function of the drug (17). An
additional component of the NM that is important for protecting the
liver against toxicity is N-acetyl cysteine. This is used as
an antidote for acetaminophen toxicity, as it increases glutathione
stores, providing a glutathione substitute and directly conjugates
with N-acetyl-p-benzoquinoneimine (NAPQI), a toxic metabolic
by-product. N-Acetyl cysteine has been shown to protect
animals from the cardiotoxicity of doxorubicin (18).
Green tea polyphenols have also shown protective
effects against the administration of toxic chemicals. Pretreatment
with epigallocatechin gallate (EGCG) led to a dose-dependent
reduction of all the histological and biochemical variables of
liver injury observed in carbon tetrachloride-treated mice
(19). Green tea polyphenols
reduced the severity of liver injury with lower concentrations of
lipid peroxidation and proinflammatory nitric oxide generated
mediators. Hasegawa et al reported that pretreatment of male
rats with green tea as drinking water provided effective protection
against the induction of hepatic degenerative changes by the
carcinogen 2-nitropropane (20).
Patil et al observed that green tea extract protected rats
from doxorubicin-induced electrocardiographic changes and changes
in biochemical markers, including lactate dehydrogenase, creatine
kinase and glutamic oxaloacetate transaminase in serum, as well as
superoxide dismutase, catalase and reduced glutathione, membrane
bound enzymes and decreased lipid peroxidation in heart tissue
(21).
Based on previously published studies, we
hypothesize that metabolic effects are likely to result from the
synergy of ascorbic acid, lysine, proline, green tea extract,
arginine, N-acetyl cysteine, quercetin, selenium, copper and
manganese. Combining these micronutrients expands metabolic
targets, maximizing biological impact with lower doses of
components. A previous study of the comparative effects of NM,
green tea extract and EGCG on the inhibition of MMP-2 and MMP-9
secretion in various cancer cell lines with varying MMP secretion
patterns, revealed the superior potency of NM over green tea
extract and EGCG at equivalent doses (22).
In conclusion, the present study demonstrated that
pretreatment with a diet supplemented with 1% NM for three weeks
reduced hepatic and renal damage in BALB/c mice following the
administration of a toxic dose of ADR. Supplementation with dietary
NM reduced the ADR-induced elevated hepatic and renal serum markers
in mice. Although clinical studies are required, the results
obtained indicate the therapeutic potential of using NM
adjunctively with ADR to protect against ADR-induced liver and
kidney damage.
Acknowledgements
Liver and kidney enzyme analyses were provided by
IDEXX Laboratories, Inc. (Westbrook, ME, USA). The study was funded
by the Dr. Rath Health Foundation (Santa Clara, CA, USA), a
non-profit organization.
References
|
1
|
Saad SY, Najjar TA and Al-Rikabi AC: The
preventive role of deferoxamine against acute doxorubicin-induced
cardiac, renal and hepatic toxicity in rats. Pharmacol Res.
43:211–218. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
DeGraff W, Hahn SM, Mitchell JB and
Krishna MC: Free radical modes of cytotoxicity of adriamycin and
streptonigrin. Biochem Pharmacol. 48:1427–1435. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Poli G: Pathogenesis of liver fibrosis:
role of oxidative stress. Mol Aspects Med. 21:49–98. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Loguercio C and Federico A: Oxidative
stress in viral and alcoholic hepatitis. Free Radic Biol Med.
34:1–10. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen Y, Jungsuwadee P, Vore M, Butterfield
DA and St Clair DK: Collateral damage in cancer chemotherapy:
oxidative stress in nontargeted tissues. Mol Interv. 7:147–156.
2007. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Essick EE and Sam F: Oxidative stress and
autophagy in cardiac disease, neurological disorders, aging and
cancer. Oxid Med Cell Longev. 3:168–177. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Feng YQ, Zuo XL, Li RF, Zhang KJ, Chen F
and Xiao H: Protection against doxorubicin-induced oxidative damage
in normal blood cells by naringenin. Zhongguo Shi Yan Xue Ye Xue Za
Zhi. 16:790–793. 2008.(In Chinese).
|
|
8
|
Kasapović J, Pejić S, Stojiljković V,
Todorović A, Radošević-Jelić L, Saičić ZS and Pajović SB:
Antioxidant status and lipid peroxidation in the blood of breast
cancer patients of different ages after chemotherapy with
5-fluorouracil, doxorubicin and cyclophosphamide. Clin Biochem.
43:1287–1293. 2010.PubMed/NCBI
|
|
9
|
Bouayed J and Bohn T: Exogenous
antioxidants - double-edged swords in cellular redox state: Health
beneficial effects at physiologic doses versus deleterious effects
at high doses. Oxid Med Cell Longev. 3:228–237. 2010. View Article : Google Scholar
|
|
10
|
Alshabanah OA, Hafez MM, Al-Harbi MM,
Hassan ZK, Al Rejaie SS, Asiri YA and Sayed-Ahmed MM: Doxorubicin
toxicity can be ameliorated during antioxidant L-carnitine
supplementation. Oxid Med Cell Longev. 3:428–433. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Venkatesan N, Punithavathi D and Arumugam
V: Curcumin prevents adriamycin nephrotoxicity in rats. Br J
Pharmacol. 129:231–234. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Niedzwiecki A, Roomi MW, Kalinovsky T and
Rath M: Micronutrient synergy – a new tool in effective control of
metastasis and other key mechanisms of cancer. Cancer Metastasis
Rev. 29:529–542. 2010.
|
|
13
|
Roomi MW, Kalinovsky T, Ivanov V, Rath M
and Niedzwiecki A: A nutrient mixture prevents acetaminophen
hepatic and renal toxicity in ICR mice. Hum Exp Toxicol.
27:223–230. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Roomi M, Kalinovsky T, Roomi NW, Ivanov V,
Rath M and Niedzwiecki A: A nutrient mixture suppresses carbon
tetrachloride-induced acute hepatic toxicity in ICR mice. Hum Exp
Toxicol. 27:559–566. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Andreadou I, Sigala F, Iliodromitis EK,
Papaefthimiou M, Sigalas C, Aligiannis N, Savvari P, Gorgoulis V,
Papalabros E and Kremastinos DT: Acute doxorubicin cardiotoxicity
is successfully treated with the phytochemical oleuropein through
suppression of oxidative and nitrosative stress. J Mol Cell
Cardiol. 42:549–558. 2007. View Article : Google Scholar
|
|
16
|
Granados-Principal S, Quiles JL,
Ramirez-Tortosa CL, Sanchez-Rovira P and Ramirez-Tortosa MC: New
advances in molecular mechanisms and the prevention of adriamycin
toxicity by antioxidant nutrients. Food Chem Toxicol. 48:1425–1438.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shimpo K, Nagatsu T, Yamada K, Sato T,
Niimi H, Shamoto M, Takeuchi T, Umezawa H and Fujita K: Ascorbic
acid and adriamycin toxicity. Am J Clin Nutr. 54(6 Suppl):
1298S–1301S. 1991.PubMed/NCBI
|
|
18
|
Doroshow JH, Locker GY, Ifrim I and Myers
CE: Prevention of doxorubicin cardiac toxicity in the mouse by
N-acetylcysteine. J Clin Invest. 68:1053–1064. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen JH, Tipoe GL, Liong EC, So HS, Leung
KM, Tom WM, Fung PC and Nanji AA: Green tea polyphenols prevent
toxin-induced hepatotoxicity in mice by down-regulating inducible
nitric oxide-derived prooxidants. Am J Clin Nutr. 80:742–751.
2004.PubMed/NCBI
|
|
20
|
Hasegawa R, Chujo T, Sai-Kato K, Umemura
T, Tanimura A and Kurokawa Y: Preventive effects of green tea
against liver oxidative DNA damage and hepatotoxicity in rats
treated with 2-nitropropane. Food Chem Toxicol. 33:961–970. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Patil LJ, Bothara SB and Balaraman R:
Effect of chronic administration of green tea extract on chemically
induced electrocardiographic and biochemical changes in rat heart.
Int J Green Pharm. 4:170–173. 2010. View Article : Google Scholar
|
|
22
|
Roomi MW, Monterrey JC, Kalinovsky T, Rath
M and Niedzwiecki A: Comparative effects of EGCG, green tea and a
nutrient mixture on the patterns of MMP-2 and MMP-9 expression in
cancer cell lines. Oncol Rep. 24:747–757. 2010.PubMed/NCBI
|