Introduction
Engineered skin substitutes play a potentially
important role in the treatment of extensive burns and chronic
wounds (1,2). However, existing skin substitutes
have not completely replicated native human skin, since they lack
skin appendages (1). Sweat
glands, one of the skin appendages, serve a number of vital
functions, including the maintenance of homeostasis and thermal
regulation (3). Therefore, the
regeneration of sweat glands has become a recent research goal in
skin tissue engineering.
Approaches to sweat gland regeneration include: i)
studies on the tubular morphogenesis in vitro of human
eccrine sweat gland epithelial cells (hESGECs) cultured in Matrigel
(4); ii) regeneration of
functional sweat gland-like structures following the implantation
of bone mesenchymal stem cells co-cultured with sweat gland cells
(5); iii) constitution of
engineered skin constructs with sweat glands by incorporating sweat
gland cell-microsphere complexes in vitro (6). As a consequence of this research
effort, sweat gland epithelial cells (SGECs) have been identified
as essential for the reconstitution of sweat glands in
vitro, as well as for their regeneration in situ. The
aim of the present study was to improve the methods for the
isolation and culture of hSGECs, in order to explore the possible
presence of sweat gland-derived stem cells and thereby provide an
experimental basis for the reconstruction of sweat glands and the
creation of appendage-containing engineered skin models.
In a previous study, a serum-containing medium was
used to culture hSGECs (5).
However, when we repeated these experiments, we found that there
were technical difficulties with this method, including the
keratinization of cultured cells and fibroblast contamination.
To address these issues, we used enzymatic digestion
methods and serum-free medium to maintain the undifferentiated
state and harvest optimal numbers of hSGE (stem) cells. SGE (stem)
cells were verified by the identification of markers specific for
sweat glands. In the present study, we present a systematic
isolation procedure and modified culture conditions for the
isolation and growth of hSGE (stem) cells.
Materials and methods
Isolation of human sweat glands
Full thickness skin samples were obtained from the
abdomen and the upper arm area of surgical patients who were
between 15 to 43 years of age. Following the removal of
subcutaneous fat under aseptic conditions, the skin was rinsed with
phosphate-buffered saline (PBS) containing 100 μ/ml penicillin and
100 μg/ml streptomycin (HyClone Laboratories, Inc., Logan, UT,
USA). A part of the sample was fixed in 10% formalin for the
preparation of paraffin sections and the remainder was minced into
1-mm3 sections and digested overnight with 0.1%
collagenase type I (Gibco, Grand Island, NY, USA) in a 37°C/5%
CO2 incubator. On the following day, the sweat glands
were isolated with a pipettor under a clean ultraviolet-sterilized
phase contrast microscope.
Primary culture of intact glands
Firstly, the glands were transferred to DMEM/F-12
(1:1) medium (Gibco) containing 5% fetal bovine serum (FBS;
HyClone). They were then cultured in sweat gland culture medium
consisting of DMEM/F-12 (1:1) medium supplemented with 5% (v/v)
FBS, 100 μ/ml penicillin and 100 μg/ml streptomycin, 10 ng/ml
endothelial growth factor (EGF; Promega Corp., Madison, WI, USA), 2
mM L-glutamine (Sigma, St. Louis, MO, USA), 1 ml/100 ml
insulin-transferring-sodium (Sigma), 2 nM/ml triiodothyronine
(Sigma) and 0.4 g/ml hydrocortisone 21-hemisuccinate (Sigma).
Primary serum-containing culture and
purification
The primary SGECs were cultured in sweat gland
culture medium and the purification was performed as follows: the
medium was removed, the cells were rinsed with PBS twice, then
digested with 0.25% trypsin (Invitrogen Life Technologies,
Carlsbad, CA, USA) and 0.02% EDTA (Tong Zheng, Beijing, China).
When most of the fibroblasts had retracted and the SGECs remained
adherent, an equal volume of culture medium containing 10% (v/v)
FBS was added to terminate the digestion. Fibroblasts were detached
with a pipettor and removed by washing with PBS. Fresh sweat gland
culture medium was added for further culture.
Primary serum-free culture and cell
passaging
When the sweat gland tissues had adhered to the
bottom of the culture well and a few cells had grown out from the
explants, serum-free keratinocyte medium containing 50 μg/ml bovine
pituitary extract (BPE) and 5 ng/ml EGF (all from Invitrogen Life
Technologies) were added to replace the sweat gland culture medium.
The SGECs were then cultured in a 37°C/5% CO2 incubator.
The medium was changed every 2–3 days.
The primary SGECs were passaged when they reached
approximately 60–80% confluence. The cells were rinsed twice with
PBS, and were then digested with 0.25% trypsin and 0.02% EDTA in a
37°C/5% CO2 incubator for 5–8 min. An equal volume of
culture medium containing 10% (v/v) FBS was added to terminate the
digestion. The liquid was transferred into a centrifuge tube,
centrifuged at 1,000 rpm/min for 5 min, and the cell pellets were
collected and resuspended in serum-free keratinocyte medium
containing 50 μg/ml BPE, 5 ng/ml EGF and 1% FBS.
Immunofluorescent histochemical
staining
After dewaxing and hybration, the sectioned samples
were blocked with 10% FBS. The sections were incubated with primary
antibodies at 4°C overnight. The antibodies used were anti-CD7,
anti-CD8, anti-CD14, anti-CD15, anti-CD18, anti-CD19 and
anti-carcinoembryonic antigen (CEA) (Abcam, Cambridge, MA, USA).
The sections were then incubated with Alexa Fluor 488-conjujated
anti-mouse/anti-rabbit secondary antibodies (Cell Signaling
Technology, Inc., Danvers, MA, USA), for 1 h at room temperature.
Nuclei were stained with Hoechst 33342 (Invitrogen Life
Technologies).
Immunohistochemical staining
After dewaxing and hydration, the sectioned samples
were treated with 3% H2O2 for 10 min to block
endogenous peroxidase activity. Subsequently, 10% normal goat serum
was used to block non-specific binding. The sections were incubated
with primary antibody against leucine-rich repeat-containing G
protein-coupled receptor 5 (LGR5) at 4°C overnight, followed by
incubation with goat anti-mouse/rabbit secondary antibodies for 10
min at room temperature. The Ultra-Sensitive™ S-P detection system
kit (Maixin, Fuzhou, China) was used with 3-amino-9-ethylarbazole
(AEC) (Boster Biological Technology, Ltd., Wuhan, China) as the
chromogenic substrate for visualization. Nuclei were counterstained
with hematoxylin.
Immunofluorescent cytochemical
staining
The SGECs on coverslips were fixed in 4%
paraformaldehyde for 10 min, permeabilized with 0.1% Triton X-100
(Sigma) for 20 min at room temperature. Subsequently, 10% FBS was
used to block non-specific binding. The SGECs were then incubated
with primary antibodies at 4°C overnight. The antibodies that were
used were the following: anti-CD7, anti-CD8, anti-CD14, anti-CD15,
anti-CD18, anti-CD19, anti-CEA and anti-LGR5 (Abcam). The cells
were then incubated with Alexa Fluor 488-conjujated
anti-mouse/anti-rabbit secondary antibodies (Cell Signaling
Technology, Inc.) for 1 h at room temperature. Nuclei were stained
with Hoechst 33342.
Reverse transcription-polymerase chain
reaction (RT-PCR)
Total RNA from the cells was isolated using TRIzol
(Invitrogen Life Technologies) reagent according to the
manufacturer’s instructions. cDNA was synthesized from 500 ng total
RNA using the Takara RT-PCR AMV 3.0 kit. PCR was carried out with 1
μl cDNA in a 20 μl reaction volume using a PCR kit (Kangweishiji
Biotech Co., Ltd., Beijing, China). A negative control was
established by using H2O as the template. PCR products
were detected by 1.5% agarose gel electrophoresis. The primers used
were as follows: CK7 (forward, 5′-GCATCAT CGCTCAGGTCAA-3′ and
reverse, 5′-TCACGGCTCCCA CTCCAT-3′); CK8 (forward,
5′-TGACCGACGAGATAAA CTTCC-3′ and reverse,
5′-CTTGGCGTTGGCATCCTTA-3′); CK14 (forward,
5′-TGAGCCGCATTCTGAACGAG-3′ and reverse,
5′-GATGACTGCGATCCAGAGGA-3′); CK15 (forward,
5′-TCTGCTAGGTTTGTCTCTTCAGG-3′ and reverse, 5′-CCA
GGGCACGTACCTTGTC-3′); CK18 (forward, 5′-TGGTCACC ACACAGTCTGCT-3′
and reverse, 5′-CCAAGGCATCACCAA GATTA-3′); CK19 (forward,
5′-AGGTGGATTCCGCTCCG GGCA-3′ and reverse,
5′-ATCTTCCTGTCCCTCGAGCA-3′); CEA (forward,
5′-GACGCAAGAGCCTATGTATG-3′ and reverse, 5′-GGCATAGGTCCCGTTATTA-3′);
LGR5 (forward, 5′-CTCTTCCTCAAACCGTCTGC-3′ and reverse, 5′-CACT
CCAAATGCACAGCACT-3′); and GAPDH (forward, 5′-TGT
TGCCATCAATGACCCCTT-3′ and reverse, 5′-CTCCACGA CGTACTCAGCG-3′).
Western blot analysis
Proteins (20 μg/lane) were fractionated by SDS-PAGE
and electrotransferred onto polyvinylidene difluoride membranes.
The blots were first incubated for 1 h in a blocking buffer
consisting of 0.1% Tween-20 (Invitrogen Life Technologies) and 5%
non-fat powdered milk, then incubated with a primary antibody at
4°C overnight. The antibodies that were used were as follows:
anti-CD7, anti-CD8, anti-CD14, anti-CD15, anti-CD18, anti-CD19
(Abcam) and anti-β-actin (Cell Signaling Technology, Inc.). A
horseradish peroxidase-conjugated, goat anti-mouse/anti-rabbit
secondary antibody (Zhongshan Jinqiao Biotechnology Co., Ltd.,
Beijing, China) then was used and antigen-antibody complexes were
detected by chemiluminescence using the BeyoECL Plus kit (BiYunTian
Biotechnology Research Laboratory, Haimen, China).
Results
Isolation of human sweat glands and
cultivation of SGECs
Following overnight digestion with 0.1% collagenase
type I, the sweat glands were dissociated from adjacent connective
tissue. Preparations of simple branched tubular glands mainly
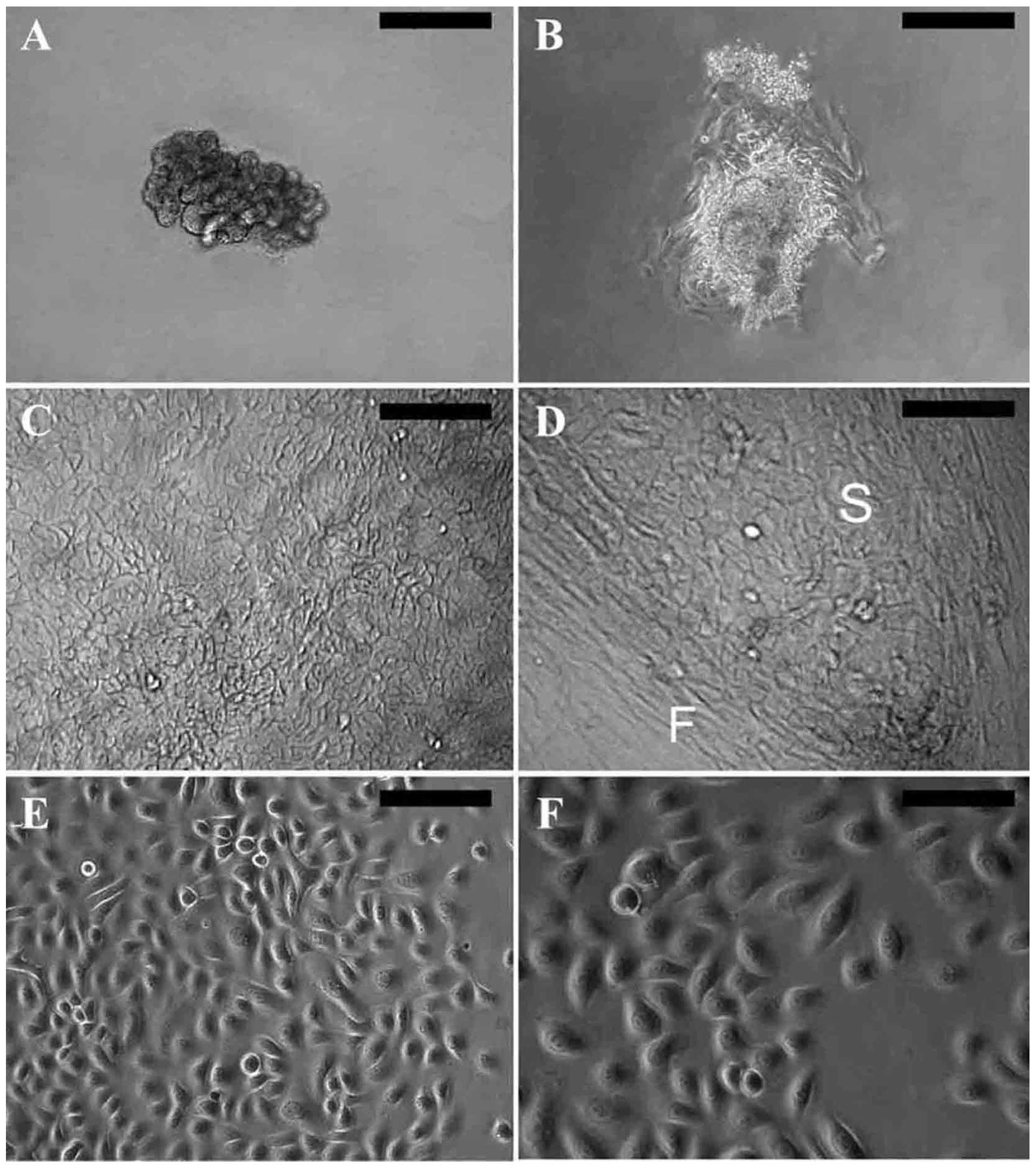
consisted of the secretory portion (Fig. 1A). Following the culture of the
whole glands in sweat gland culture medium for 2–5 days, typical
epithelial cells grew out from the explants, assuming a cobblestone
morphology (Fig. 1B). After
continued growth in sweat gland culture medium, the primary SGECs
resembled keratinized cells (Fig.
1C) and were usually contaminated by fibroblasts (Fig. 1D).
The primary SGECs assumed a good growth state after
replacing the sweat gland culture medium with keratinocyte
serum-free medium. They displayed an epithelial-like morphology
with a rounded cell shape and a large nucleus (Fig. 1E), and grew rapidly; a confluent
monolayer was formed approximately 1 week later. Few fibroblasts
survived under serum-free conditions. After one passage, cell
propagation was performed in keratinocyte serum-free medium with 1%
FBS, and the cells still showed prominent proliferative activity
(Fig. 1F).
Immunofluorescent histochemical
staining
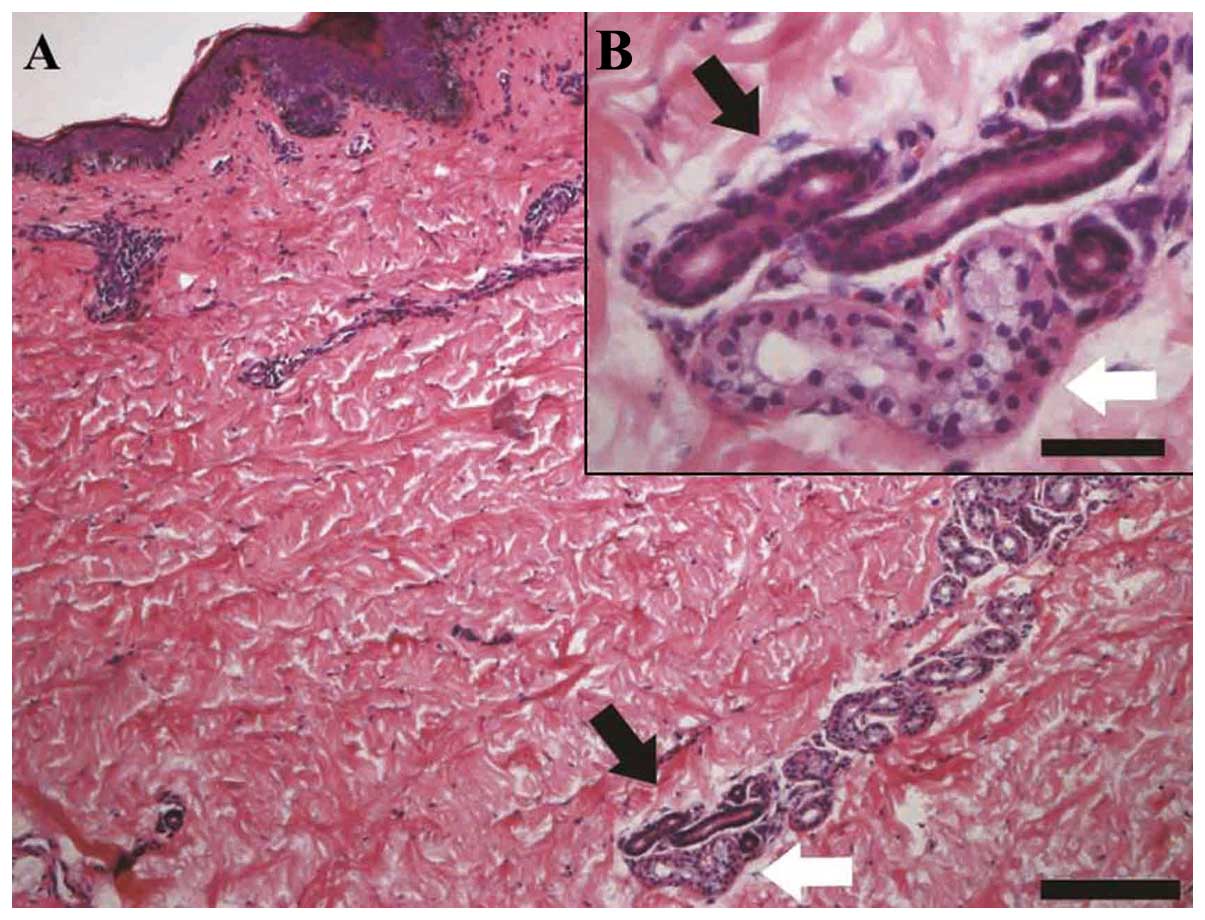
Hematoxylin and eosin (H&E) staining of adult
skin paraffin sections revealed the anatomic characteristics of
eccrine sweat glands. The secretory portion was located in the
dermis and subcutaneous tissue, with a distinct lumen (Fig. 2, white arrows). The duct consists
of 2 layers of cuboidal cells (Fig.
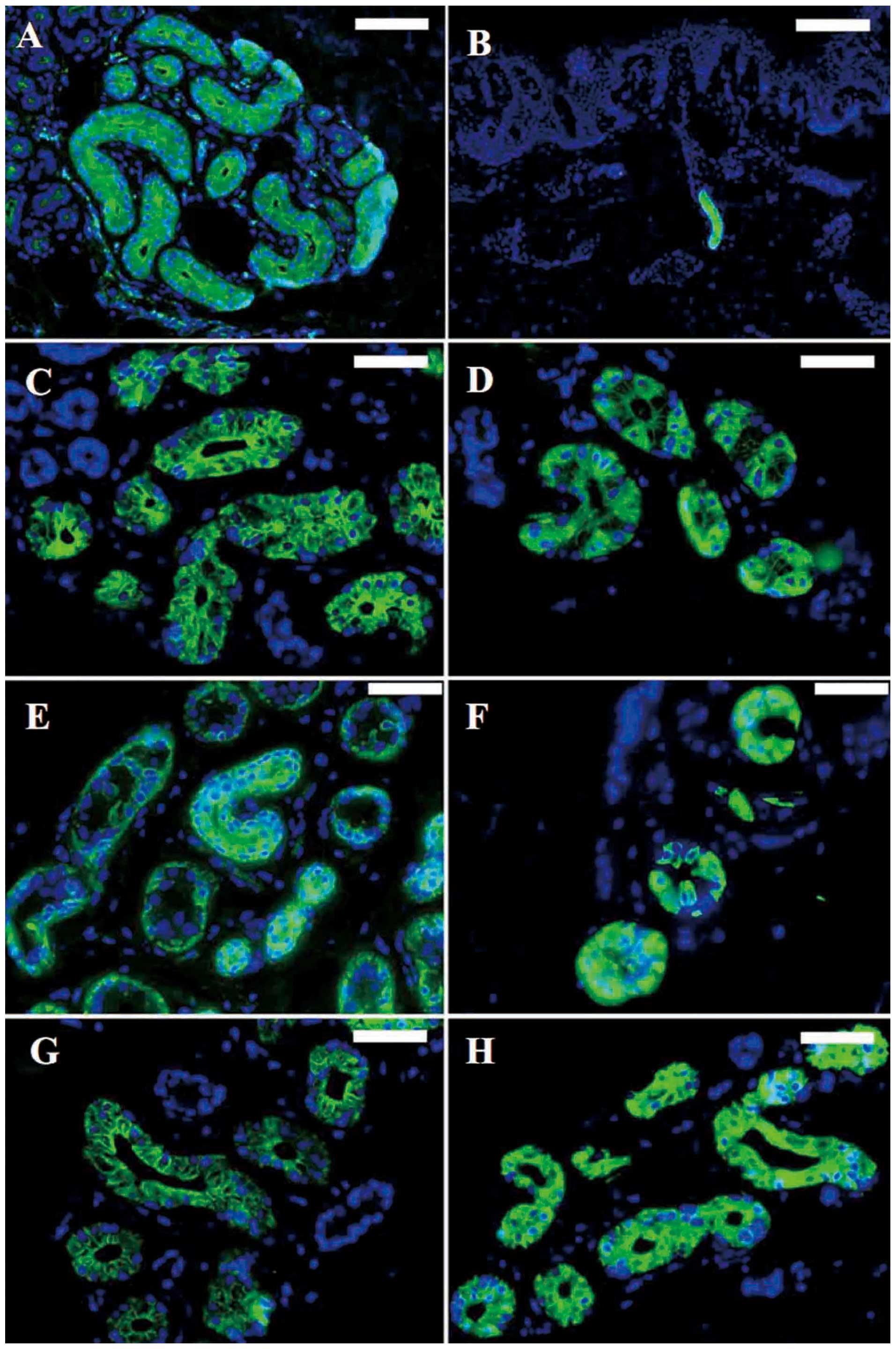
2, black arrows). Immunofluorescent histochemical staining
revealed that CEA (Fig. 3A and B)
and CK14 (Fig. 3E) were expressed
in both the secretory and the ductal portion, and that CK7
(Fig. 3C), CK8 (Fig. 3D), CK15 (Fig. 3F), CK18 (Fig. 3G) and CK19 (Fig. 3H) were expressed in the secretory
portion.
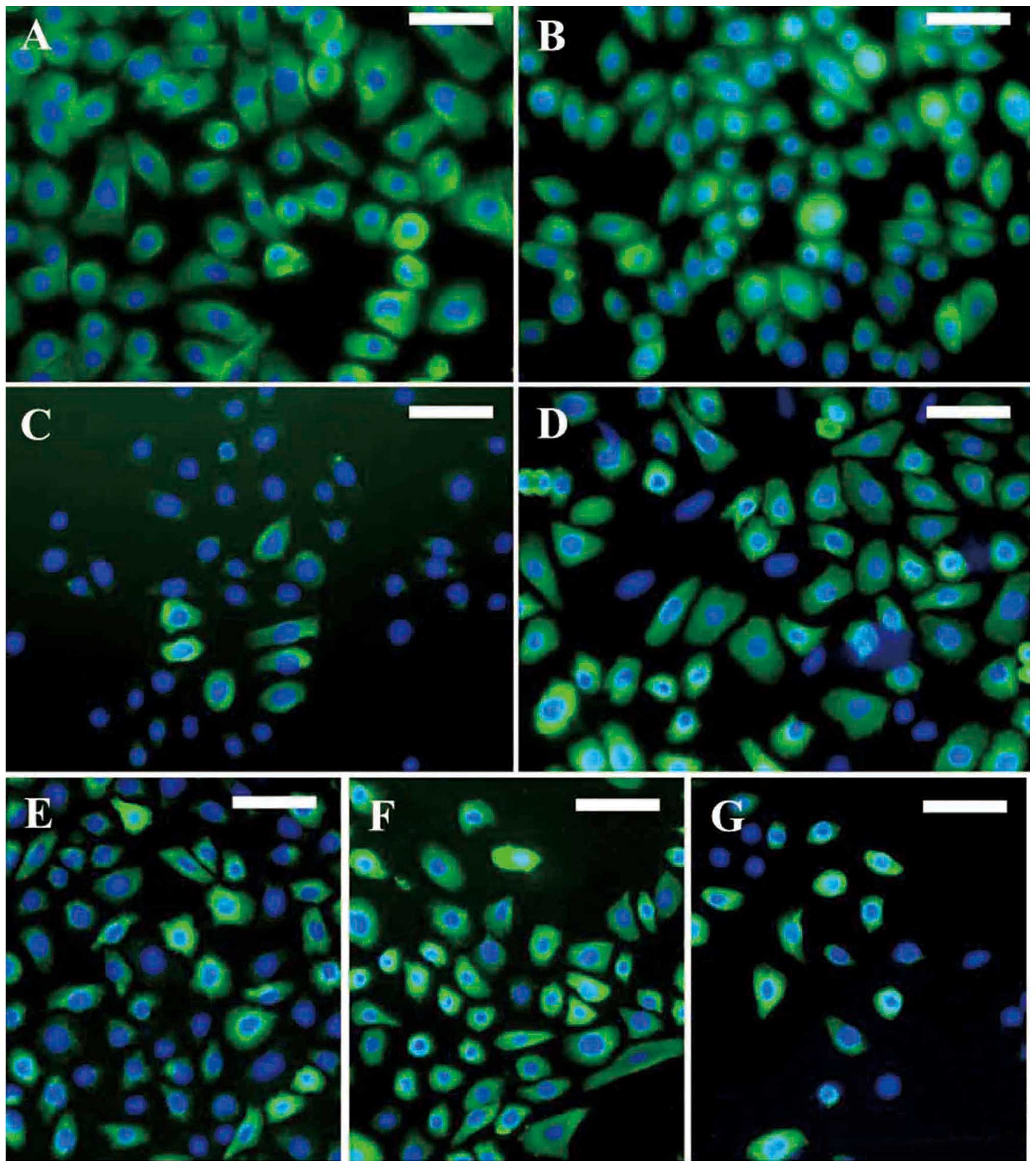
Immunofluorescent staining of SGECs
Immunofluorescent staining revealed that the SGECs
were positive for CEA (Fig. 4A),
CK7 (Fig. 4B), CK8 (Fig. 4C), CK14 (Fig. 4D), CK15 (Fig. 4E), CK18 (Fig. 4F) and CK19 (Fig. 4G), in accordance with the skin
sections.
RT-PCR and western blot analysis
The markers specific for sweat glands were
identified by RT-PCR. CEA, CK7, CK8, CK14, CK15, CK18 and CK19 were
detected at the mRNA level (Fig.
5A). Western blot analysis with keratin subunit-specific
monoclonal antibodies confirmed the expression of CK7, CK8, CK14,
CK15, CKl8 and CKl9 in the SGECs. The expression of these specific
markers distinguished these cells from fibroblasts (Fig. 5B).
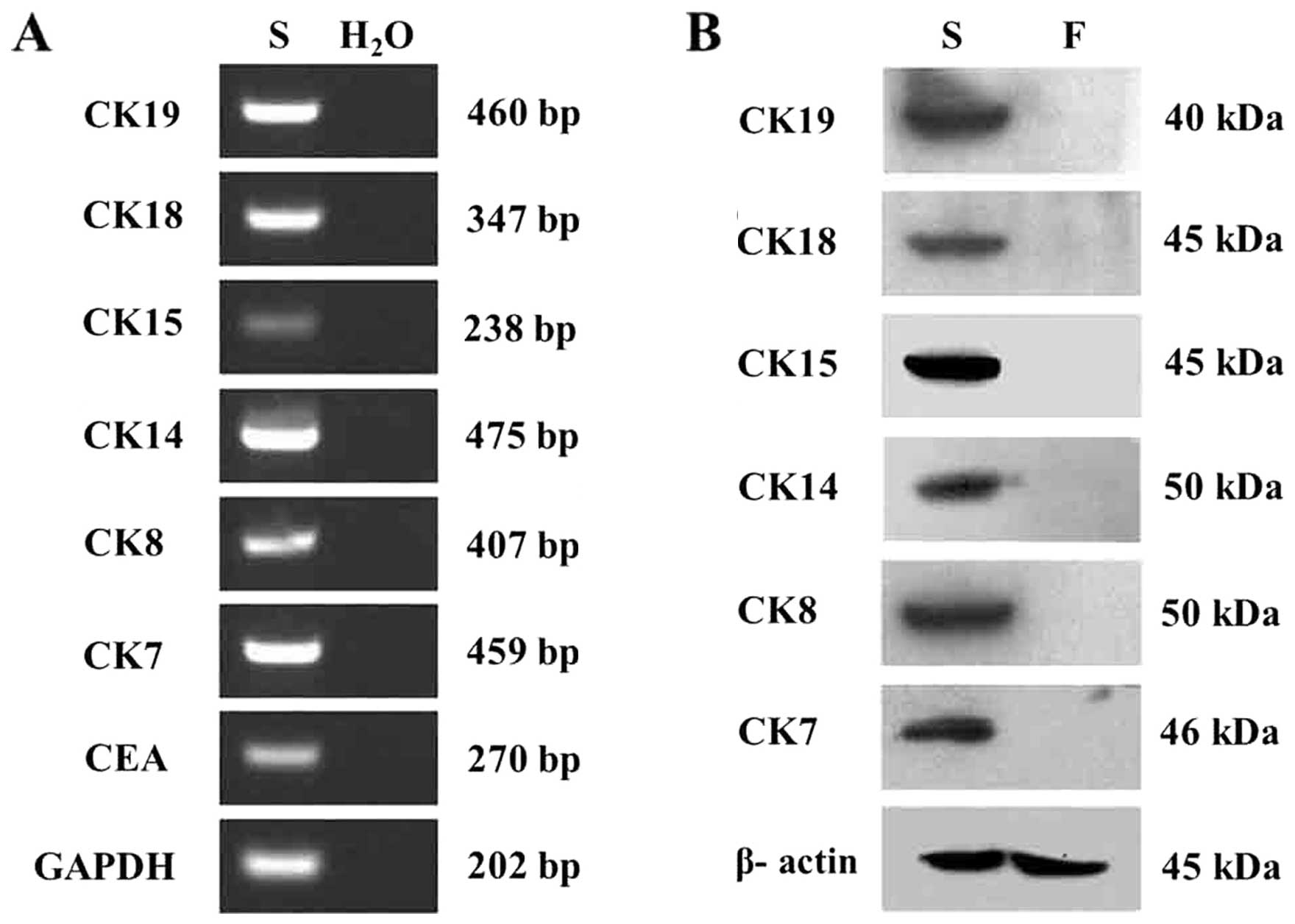 | Figure 5Identification of of markers specific
for sweat glands in human sweat gland epithelial cells (hSGECs) by
reverse transcription-polymerase chain reaction (RT-PCR) and
western blot analysis. (A) Carcinoembryonic antigen (CEA), CK7,
CK8, CK14, CK15, CK18 and CK19 were detected in SGECs (S) at the
mRNA level; a negative control was established by using
H2O as the template. (B) CK7, CK8, CKl4, CKl5, CKl8,
CKl9 were detected in the SGECs (S) by western blot analysis,
compared with the fibroblasts (F) in which these markers were not
found. |
Expression of LGR5 in SGECs
We found that the SGECs expressed LGR5, as shown by
RT-PCR (Fig. 6A) and
immunocytochemistry (Fig. 6B and
C), known as the stem cell marker of intestinal cells. To
confirm our results, we stained the sweat gland tissue in the skin.
Consistent with the immunocytochemistry results, the epithelial
cells in the sweat glands of the skin tissue also expressed LGR5
(Fig. 6D and E).
Discussion
hSGECs are essential for the reconstitution of sweat
glands in vitro (4), as
well as for their regeneration in situ (5). Moreover, sweat gland cells can
reconstitute a functional, stratified epidermis (7) and participate in the construction of
engineered skin constructs with sweat glands (6). Therefore, an optimized method for
isolating and culturing hSGE (stem) cells is important for the
reconstruction of sweat glands and the creation of skin
appendage-containing engineered skin models.
In the present study, intact sweat glands were
isolated from small sections of skin by pipetting under a phase
contrast microscope following overnight digestion with 0.1%
collagenase type I. Subsequently, the intact glands were cultured,
making it possible to determine the source of cells for primary
culture. Initially, we tried a traditional serum-containing culture
medium (5); however, the primary
hSGECs resembled keratinized cells and were usually contaminated by
fibroblasts. Consequently, we established a serum-free culture
method and obtained primary hSGECs with a good growth state. After
one passage, the cells cultured in keratinocyte serum-free medium
with 1% FBS still showed proliferative activity. The SGECs formed a
‘dome-like’ structure after growing into a confluent monolayer that
corresponded with the biological characteristics of glandular
epithelial cells which may paly a role in ion transmembrane
transportation (8,9). Immunofluorescence microscopy, RT-PCR
and western blot analysis demonstrated the expression of specific
phenotypic surface markers, including CEA, CK7, CK8, CK14, CK15,
CK18, and CK19 in cultured SGECs. We concluded that serum-free
culture provides an optimal culture condition for hSGECs.
Sweat glands are detected in embryos at 14–16 weeks
and they reach maturation in 24–week embryos (10). It is generally accepted that there
is no sweat gland formation after birth. The markers expressed in
human sweat glands are also different at each developmental stage
of embryogenesis. In normal adult skin tissue, CK7, CK8, CK14,
CK15, CK18 and CK19 are expressed in the secretory portion
(5,11).
CEA is an important marker for the diagnosis of
colorectal cancer and other types of cancer, bu also shows limited
tissue expression in normal adult tissues, including columnar
epithelial cells and goblet cells in the colon, mucous neck cells
and pyloric mucous cells in the stomach, secretory epithelia and
duct cells of sweat glands (12).
That means CEA is only expressed in sweat glands in normal adult
skin tissue and is therefore considered as a relatively specific
marker for the identification of sweat gland cells.
Thus, the high expression of CEA, CK7, CK8, CK14,
CK15, CK18 and CK19 may be considered as an index for identifying
adult SGECs (5,11–12). According to cell morphology and
the expression of tissue-specific markers, we came to the
conclusion that SGECs were successfully isolated and passaged.
It has been proven that there are multipotent stem
cells in the hair follicle bulge areas and that they contribute to
epidermal regeneration following skin injuries (13). Human glabrous skin, which
completely lacks hair follicles but contains abundant sweat glands,
such as the skin area of the palms, also has epidermal regenerative
potential (7), suggesting the
possible presence of stem cells in human sweat glands.
Sweat glands contain 4 segments: an intraepidermal
duct, an intradermal ‘straight’ duct, an intraglandular ‘coiled’
duct and a secretory portion (14). Label-retaining cells are located
in the intraglandular ‘coiled’ duct and secretory segments. These
cells express CK15, a positive marker for stem cells residing in
the hair follicle bulge areas (15,16). In this study, the cultured SGECs
were mainly derived from the secretory portion and we found clear
evidence that they strongly expressed CK15. To the best of our
knowledge, for the first time, the expression of LGR5 was detected
by RT-PCR and immunohistochemistry in human sweat glands. This is a
novel marker that specifically identifies intestinal epithelial
stem cells and hair follicle stem cells (17,18). The expression of CK15 and LGR5
indicates that CK15 and LGR5 may also represent markers of human
sweat gland-derived stem cells.
In conclusion, SGE (stem) cells were successfully
isolated by collagenase digestion and harvested in culture in
serum-free culture medium. The present study provides an
experimental basis for the reconstruction of sweat glands and the
creation of skin appendage-containing engineered skin models.
Acknowledgements
The present study was supported by grants from the
State Key Development Program of Basic Research of China (no.
211CB606200), the National Natural Science Foundation of China (no.
30930026/C100101), the Science and Technology Planning Project of
Jilin Province, China (nos. 20100947 and 20120935), the Frontier
Interdiscipline Program of Norman Bethune Health Science Center of
Jilin University (2013101007) and the Specialized Research Fund for
the Doctoral Program of Higher Education (20130061110077).
References
|
1
|
Bar-Meir E, Mendes D and Winkler E: Skin
substitutes. Isr Med Assoc J. 8:188–191. 2006.
|
|
2
|
Norbury WB, Jeschke MG and Herndon DN:
Tissue engineered fetal skin constructs for pediatric burns. Crit
Care. 9:533–534. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Shibasaki M and Crandall CG: Mechanisms
and controllers of eccrine sweating in humans. Front Biosci (Schol
Ed). 2:685–696. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lei X, Liu B, Wu J, Lu Y and Yang Y:
Matrigel-induced tubular morphogenesis of human eccrine sweat gland
epithelial cells. Anat Rec (Hoboken). 294:1525–1531. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sheng Z, Fu X, Cai S, Lei Y, Sun T, Bai X
and Chen M: Regeneration of functional sweat gland-like structures
by transplanted differentiated bone marrow mesenchymal stem cells.
Wound Repair Regen. 17:427–435. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Huang S, Xu Y, Wu C, Sha D and Fu X: In
vitro constitution and in vivo implantation of engineered skin
constructs with sweat glands. Biomaterials. 31:5520–5525. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Biedermann T, Pontiggia L,
Böttcher-Haberzeth S, et al: Human eccrine sweat gland cells can
reconstitute a stratified epidermis. J Invest Dermatol.
130:1996–2009. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Buchanan JA, Yeger H, Tabcharani JA, et
al: Transformed sweat gland and nasal epithelial cell lines from
control and cystic fibrosis individuals. J Cell Sci. 95:109–123.
1990.PubMed/NCBI
|
|
9
|
Xia L: Biological characters of human
eccrine sweat gland epithelial cells and three-dimensional
reconstruction of eccrine sweat glands in vitro. Third Military
Medical University; Chong Qing, China: 2006
|
|
10
|
Li J, Fu X and Sheng Z: Study on the
relationship between epidermal stem cells and the developing
process of sweat gland in human fetal skin. Zhonghua Shao Shang Za
Zhi. 18:369–371. 2002.(In Chinese).
|
|
11
|
Li HH, Zhou G, Fu XB and Zhang L: Antigen
expression of human eccrine sweat glands. J Cutan Pathol.
36:318–324. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hammarström S: The carcinoembryonic
antigen (CEA) family: structures, suggested functions and
expression in normal and malignant tissues. Semin Cancer Biol.
9:67–81. 1999.PubMed/NCBI
|
|
13
|
Ito M, Liu Y, Yang Z, Nguyen J, Liang F,
Morris RJ and Cotsarelis G: Stem cells in the hair follicle bulge
contribute to wound repair but not to homeostasis of the epidermis.
Nat Med. 11:1351–1354. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Langbein L, Rogers MA, Praetzel S, Cribier
B, Peltre B, Gassler N and Schweizer J: Characterization of a novel
human type II epithelial keratin K1b, specifically expressed in
eccrine sweat glands. J Invest Dermatol. 125:428–444. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nakamura M and Tokura Y: The Localization
of label-retaining cells in eccrine glands. J Invest Dermatol.
129:2077–2078. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ohyama M, Terunuma A, Tock CL, et al:
Characterization and isolation of stem cell-enriched huma hair
follicle bulge cells. J Clin Invest. 116:249–260. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Barker N, van Es JH, Kuipers J, et al:
Identification of stem cells in small intestine and colon by marker
gene Lgr5. Nature. 449:1003–1007. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Haegebarth A and Clevers H: Wnt signaling,
lgr5, and stem cells in the intestine and skin. Am J Pathol.
174:715–721. 2009. View Article : Google Scholar : PubMed/NCBI
|