Introduction
Ovarian cancer is one important cause of gynecologic
cancer-associated mortality, with an overall five-year survival
rate of 45% and an overall 10-year survival rate of 35% in the USA
(1,2). Globally, it is estimated that
~283,000 new cases of ovarian cancer are likely to be diagnosed in
the world in 2020, with ~66% of the cases affecting women aged
<65 years (3). At present,
ovarian cancer treatment consists of cytoreductive surgery and
platinum-based chemotherapy. However, as the disease is usually
diagnosed at an advanced stage, its prognosis remains poor
(4,5). Although substantial progress has
been made in the research and development of novel targeted
therapies for tumors or cancer in humans, including human ovarian
cancer, a useful and effective alternative for drug research is the
repurposing of drugs. At present, examples of this type of drug are
in different stages of clinical trials (6), and improved targeted therapeutic
strategies are required to inhibit the progression of ovarian
cancer.
Epidemiological studies have suggested that the
intake of cruciferous vegetables, including broccoli, reduces the
risks for the induction of certain forms of cancer (7). The natural compound, sulforaphane,
was isolated from broccoli in the early 1990s as an inducer of
phase 2 enzymes (xenobiotic metabolism), and numerous studies have
since proposed various anti-neoplastic pharmacological properties
of sulforaphane, thereby suggesting its potential as a promising
candidate in cancer chemoprevention (8–10).
Pharmacokinetic investigations in rats and humans have demonstrated
that sulforaphane can be distributed in the body and reach
micromolar concentrations in the blood. Multiple mechanisms of
action inherent to the anticancer properties of sulforaphane have
been reported (11,12). Dietary consumption of cruciferous
vegetables containing sulforaphane may reduce the risk of several
common types of cancer, including prostate, breast, lung and
colorectal cancer (13–16). For example, it can inhibit the
migration and invasion of glioblastoma cells (17), and has also been considered for
the treatment of cervical cancer cells (18). Additionally, sulforaphane is
involved in the inhibition of breast cancer cell proliferation and
the induction of apoptosis of certain cancer cell lines, including
osteosarcoma (19).
The present study indicated that sulforaphane acted
synergistically with the chemotherapeutic drug cisplatin to inhibit
ovarian cancer cell proliferation and promote apoptosis in
vitro. In the in vivo experiments, sulforaphane
effectively inhibited xenograft tumor growth and progression, at
least partly through inhibiting cell proliferation via
cancer-related signaling pathway regulation. Therefore, these
results indicated that sulforaphane offers potential and may be
repurposed as an anti-human ovarian cancer agent. However, further
investigations are required to examine the anticancer role of
sulforaphane in preclinical and clinical trials in the future.
Materials and methods
Cell culture and treatment
The human ovarian cancer cell lines, A2780 and
OVCAR, were purchased from the American Type Culture Collection
(Manassas, VA, USA) and the Cell Resource Center, Shanghai
Institute of Biochemistry and Cell Bank at the Chinese Academy of
Sciences (Shanghai, China). The cell lines were routinely
authenticated by DNA-fingerprinting and isoenzyme analyses, and
checked for contamination by mycoplasma using Hoechst staining. All
cell lines were maintained in Roswell Park Memorial Institute-1640
(Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA),
Dulbecco's modified Eagle's medium or Minimum Essential Medium,
containing 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.), 100 μg of streptomycin and 100 units of
penicillin. A2780 were cultured in RPMI-1640 medium and OVCAR cells
in DMEM medium. The cells were incubated at 37°C with 5%
CO2. The sulforaphane chemical was obtained from
Hangzhou Lin'an Tianhong Bio-Tech Co., Ltd. (Hangzhou, China).
Colony formation and migration assay
The human ovarian cancer cells were suspended in
0.9% methylcellulose-based semisolid medium MethoCult H4100
(StemCell Technologies, Inc., Beijing, China), with 1,000 cells
seeded in 3.5 cm dishes, and treated with 0, 2.5, 5 or 10 μM
sulforaphane or ethanol control, and maintained at 37°C. After 14
days, individual primary clones (450 cells) were trypsinized and
replated in the same conditions to examine the secondary colony
forming ability for self-renewal. For the Transwell migration
assays, 10×104 cells were planted in the top chamber
with a non-coated membrane. The cells were seeded in serum-free
medium, and a medium containing 10% serum used as a chemoattractant
in the lower chamber. The cells were then incubated for 16 h at
37°C and 5% CO2 in a tissue culture incubator. After 16
h, the non-migrated cells were removed from the upper sides of the
Transwell membrane filter inserts with cotton-tipped swabs. The
migrated cells on the lower sides of the inserts were then stained
with Giemsa, and the cells were counted under a Nikon Eclipse Ti2
inverted microscope (Nikon Corporation, Tokyo, Japan).
Cell viability assay
The viable cancer cells were determined via Trypan
blue exclusion staining analysis. In brief, the A2780 and OVCAR
cells were administrated with sulforaphane at the indicated
concentrations or with the vehicle control. After 48 and 72 h, the
cells were collected through trypsin dissociation, and then stained
with Trypan blue until a final concentration of 0.1% Trypan blue.
The unstained viable cancer cells were then counted under a bright
field microscope. The assay was performed in triplicate.
Cell proliferation assay
The ovarian cancer cell proliferation was measured
using Premixed WST-1 reagent (Roche Diagnostics, Basel,
Switzerland). In brief, the A2780 and OVCAR cells were seeded in
96-well plates, and were treated with sulforaphane and/or other
drugs at the indicated concentrations for 24 or 48 h. WST-1 reagent
(10 μl) was then administrated to each well, which was
followed by incubation at 37°C for 30–60 min. The wells were
finally read at 440 nm using the microplate reader (Thermo Fisher
Scientific, Inc.). The assay was performed in triplicate.
Cell wound healing assay
When the A2780 cells reached 95% confluence, they
were wounded with micro-pipette tips and treated with sulforaphane
at the indicated concentrations. The measurement of the wound gap
was recorded at 0, 24 and 48 h following sulforaphane treatment
under bright field microscopy. The assay was performed in
triplicate.
Analysis of apoptosis using Hoechst 33258
staining
The A2780 and OVCAR cells were treated with the
various concentrations of sulforaphane or ethanol control. After 24
h, the cells were collected, and fixed and stained with Magic
Solution containing 10X stock of 0.5% NP-40, 3.4% formaldehyde and
10 μg/ml Hoechst 33258 in PBS. The apoptotic cells were
determined and recorded using a fluorescence microscope. The assay
was performed in triplicate. The average number of apoptotic
ovarian cancer cells was evaluated by counting the apparent
apoptotic cells in 10 randomly selected fields at ×100
magnification for each condition.
Flow cytometric analysis
A flow cytometric assay was used to clarify the
apoptotic cells and cell cycle arrest following sulforaphane
treatment. Following treatment with sulforaphane for 48 h, the
ovarian cancer cells were collected with trypsinization and then
washed twice with PBS, fixed in cold 80% ethanol, and finally
stored at 4°C overnight. The cells were then washed twice with PBS,
and RNase A (10 mg/ml) was administrated for analysis. Propidium
iodide (PI) was then added at a concentration of 0.05 mg/ml and
incubated for 20 min at 4°C in the dark. The FITC-labeled Annexin
V/PI staining was applied according to the manufacturer's protocol
(Nanjing Keygen Biotech Co., Ltd., Nanjing, China). In brief,
1×106 cells in each well were suspended with buffer
containing FITC-conjugated Annexin V/PI. The samples were then
analyzed using flow cytometry.
Immunofluorescence assay
Following induction with the conditioned culture
medium, the cells were fixed in 4% paraformaldehyde, and
permeabilized with 0.1% Triton X-100 in PBS containing 0.5% BSA
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) for 30 min. The
cells at 70–80% confluence were subsequently incubated with
antibodies against human epidermal growth factor receptor 2 (Her2;
1:200; 4290; Cell Signaling Technology, Inc., Danvers, MA, USA) for
30 min at room temperature, followed by labeling with Alexa Fluor
488-conjugated rabbit anti-mouse IgG antibody at room temperature
in the dark for 30 min (A27023; 1:1,000 Thermo Fisher Scientific,
Inc.). The cells were viewed under a fluorescent microscope.
Immunohistochemical (IHC) assay
The sections (6 μm) of the paraffin-embedded
tumor tissue blocks were deparaffinized and rehydrated. The tumor
tissue samples were incubated with primary monoclonal antibodies
against Her2 and KI67 (1:200; 9449; Cell Signaling Technology,
Inc.) at 4°C overnight. The slide was then gently rinsed with PBS
and developed using the Envision system/HRP for 30 min and
substrate-chromogen for 15 min at room temperature. The nuclei were
counterstained with Mayer's hematoxylin (MHS1-100ML; Sigma-Aldrich;
Merck KGaA) under an Eclipse TE2000E inverted microscope (Nikon
Corporation).
Analysis of apoptosis using terminal
deoxynucleotidyl transferase-mediated dUTP nick end labeling
(TUNEL)
The analysis of apoptosis in the tissue samples was
determined by TUNEL using an in situ cell death detection
kit, Fluorescein (Roche Applied Science, Madison, WI, USA)
according to the manufacturer's protocol. The number of
TUNEL-positive cells was counted under a fluorescence microscope.
The percentages of apoptotic cells were calculated from the ratio
of apoptotic cells to total cells counted. The tissue sections were
counter-stained with hematoxylin, mounted and observed under light
microscopy. The experiment was performed three times independently
for each cell line.
Western blot analysis
Cell proteins from the ovarian cancer cells were
extracted using a T-PER Tissue Protein Extraction Reagent kit
(Thermo Fisher Scientific, Inc.) according to the manufacturer's
protocol. The protein concentrations were determined using a BCA
protein assay kit, and equal quantities of protein (40 μg)
were loaded per well on a 10% sodium dodecyl sulphatepolyacrylamide
gel. Subsequently, the proteins were transferred onto a
polyvinylidene difluoride membrane. The resulting membrane was
blocked with Tris-buffered saline containing 0.05% Tween-20
(TBS-T), supplemented with 5% skimmed milk (Sigma-Aldrich; Merck
KGaA) at room temperature for 2 h on a rotary shaker, and followed
by TBS-T washing. The membrane was incubated with specific primary
antibodies, diluted in TBST, at 4°C overnight. Subsequently, the
membrane was washed with TBS-T followed by incubation with the goat
anti-rabbit peroxidase-conjugated secondary antibody (1662408edu;
1:2,000; Bio-Rad Laboratories, Inc., Hercules, CA, USA) at room
temperature for 1 h. The immunoreactive proteins were detected
using an enhanced chemiluminescence western blotting detection kit.
The western blot bands were observed using the GE Healthcare ECL
Western Blotting Analysis system (GE Healthcare Life Sciences,
Chalfont, UK) and exposed to x-ray film (Kodak, Rochester, NY,
USA). The primary antibodies used were: Rabbit anti-P53 (cat. no.
2527), anti-Bcl-2 (cat. no. 3498), anti-phosphorylated nuclear
factor-κB (p-NF-κB; cat. no. 3033), anti-cMyc (cat. no. 5605),
anti-AKT (cat. no. 4685) and anti-p-AKT (cat. no. 4060) from Cell
Signaling Technology, Inc.; rabbit anti-Bcl-2-associated X protein
(Bax; cat. no. ab32503), anti-NF-κB (cat. no. ab16502),
anti-Caspase-3 (cat. no. 13847), anti-Cyclin-D1 (cat. no. 134175)
and mouse anti-P27 (cat. no. 193379) from Abcam (Cambridge, MA,
USA); and rabbit anti-cytochrome c (cyto-c; cat. no.
sc-13561) and anti-GAPDH (cat. no. sc-47724) from Santa Cruz
Biotechnology, Inc. (Dallas, TX, USA). All antibodies were used at
a dilution of 1:1,000, with the exception of anti-GAPDH
(1:500).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA from the cultured cells and tissue samples
was isolated using the mirVana miRNA isolation kit (Ambion; Thermo
Fisher Scientific, Inc.) based on the manufacturer's protocol. The
cDNA was then synthesized from total RNA with the Taqman miRNA
reverse transcription kit (Applied Biosystems; Thermo Fisher
Scientific, Inc.). The RT-qPCR analysis was performed using the
Applied Biosystems 7500 Sequence Detection system with iQ™
SYBR-Green SuperMix (Bio-Rad Laboratories, Inc.) containing 5 ng
cDNA and 10 pM of each primer. The PCR cycles were 95°C for 5 min,
then 95°C for 20 sec and 60°C for 60 sec for 40 cycles. The
annealing, extension and also the data reading were at 60°C. The
data were normalized to the geometric mean of housekeeping gene
GAPDH. The data were analyzed with 2-ΔΔCq method
(20). The sequences of the
primers are summarized in Table
I.
 | Table ISequences of primers used for reverse
transcription-quantitative polymerase chain reaction in the present
study. |
Table I
Sequences of primers used for reverse
transcription-quantitative polymerase chain reaction in the present
study.
| Gene | Primer (5′→3′) |
|---|
| Bcl-2
(forward) |
TAATACGACTCAATACTGGG |
| Bcl-2
(reverse) |
ACTATTTAGGTGACATAG |
| Bax (forward) |
GACTGTAGCAATGGGAGGTAGA |
| Bax (reverse) |
CCTGTGTGCATATCTTCTTGCA |
| P53 (forward) |
GGCCCTTGCTTTCTCTTCG |
| P53 (reverse) |
ATTGATTTAATAAAGTTATGT |
| P27 (forward) |
CATAAAGTGGTACGCCGCA |
| P27 (reverse) |
ACCACCAGGAAACTCCAGCAGT |
| Cyclin-D1
(forward) |
GCGGTCCAAGAAACCTGTCA |
| Cyclin-D1
(reverse) |
GAACGGCATTATCCATGCC |
| cMyc (forward) |
GAGTAAGCCCGACTTGGTTGA |
| cMyc (reverse) |
CCGACACTTCTCTGTGCTATTTG |
| Her2 (forward) |
GCTTACCGCCTACTTCACC |
| Her2 (reverse) |
GCGAGGTCCACTAGATGA |
| GAPDH
(forward) |
AGGCTGAAGGGGCTCATTTG |
| GAPDH
(reverse) |
AGCATCGGGCAGTCATCCTC |
Xenograft tumor models of human ovarian
cancer cells
The use and care of animals as models were approved
by the Institutional Animal Care and Use Committee of Zaozhuang
City Hospital (Zaozhuang, China). All experimental procedures were
performed according to the approved guidelines. The mouse
experiments were performed in the Animal Laboratory Center at
Zaozhuang City Hospital. The cells (1×107cells) treated
with various concentrations of sulforaphane, described above, were
suspended in 100 μl serum-free medium and injected
subcutaneously into the left flank of 4–6-week-old male BALB/c
nu/nu nude mice (15–18 g), with 8 mice in each groups, 32 mice in
total, were injected with cells. The mice were raised in the SPF
animal facility with sterile food and water ad libitum, at
20°C with a 50% humidity and a 12-h light/dark cycle. Tumor size
was measured with digital calipers and calculated every week. Tumor
volume was measured every 7 days and at the end (~6 weeks) of
treatment, when the mice were sacrificed. The tumors were excised,
weighed, fixed in 10% neutral formalin, and embedded in paraffin
for histological analysis under an Eclipse TE2000E inverted
microscope (Nikon Corporation).
Statistical analysis
The differences of indices are presented as the mean
± standard error of the mean using the data. The treated tissue and
the corresponding controls were compared using GraphPad Prism
(version 6.0; GraphPad Software, Inc., La Jolla, CA, USA) using
one-way analysis of variance with Dunn's least significant
difference test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Sulforaphane effectively suppresses human
ovarian cancer cell proliferation
The present study attempted to examine the effect of
the sulforaphane on the proliferative activity of A2780 and OVCAR
human ovarian cancer lines. The A2780 and OVCAR cells were
inhibited by increasing concentrations of sulforaphane. The crystal
violet staining suggested that sulforaphane effectively suppressed
cell proliferative activity in the A2780 and OVCAR cells at
different concentrations between 2.5 and 10 μM (Fig. 1A and B). The direct cell numbers
following growth of A2780 and OVCAR cells administrated with
different concentrations of sulforaphane (0–20 μM) were also
investigated. The results showed that the number of viable cells
was significantly downregulated when the sulforaphane concentration
increased in the A2780 (Fig. 1C)
and OVCAR (Fig. 1D) cell lines at
48 and 72 h. Further calculation of anti-proliferative effects was
achieved through the WST-1 proliferation assay, which indicated
that significant cell proliferation suppression occurred at varying
concentrations of sulforaphane in the A2780 (Fig. 1E) and OVCAR (Fig. 1F) cell lines. Together, these
results demonstrated that sulforaphane effectively inhibited
ovarian cancer cell proliferation.
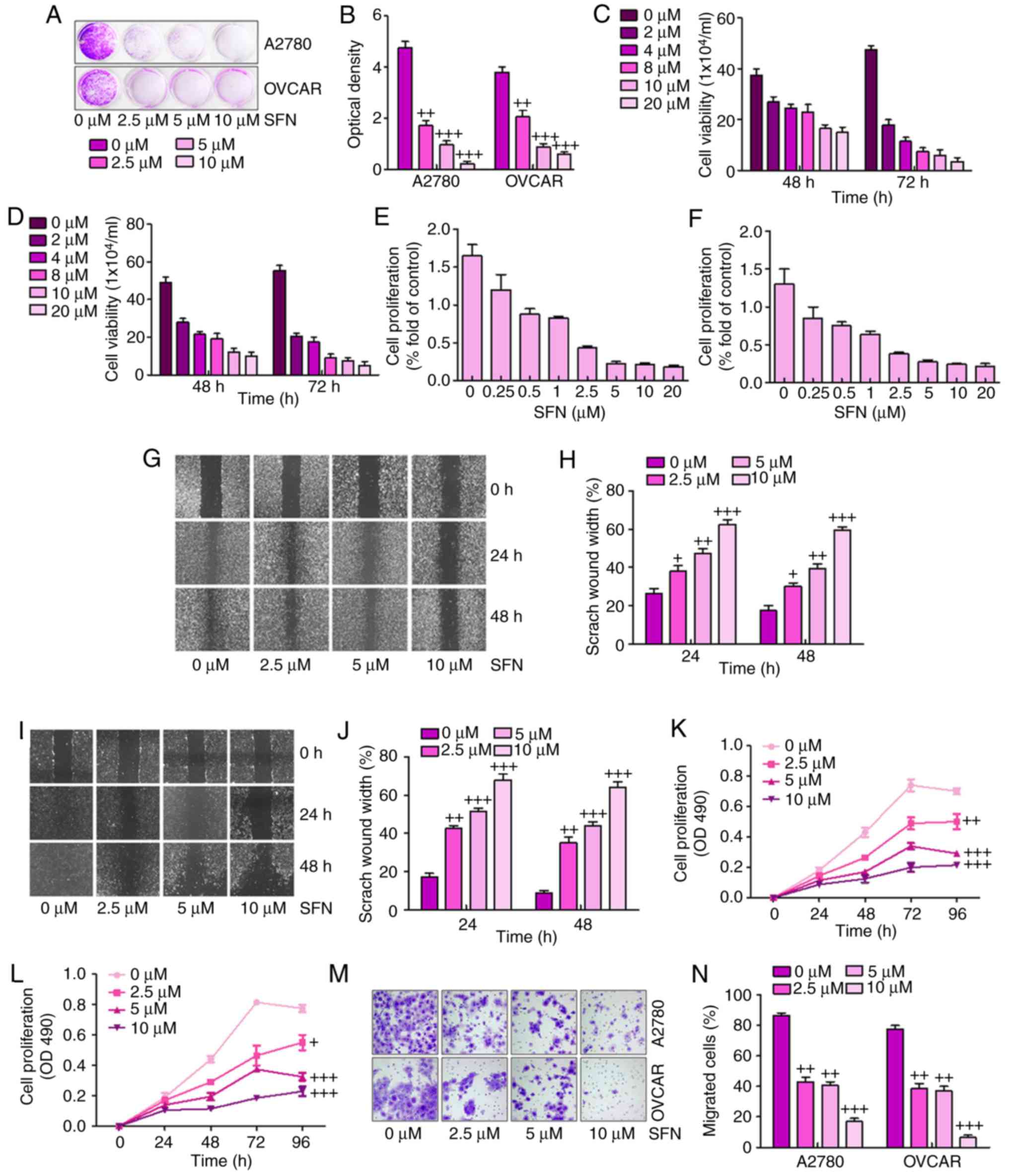 | Figure 1Sulforaphane effectively suppresses
human ovarian cancer cell proliferation. (A) Colony formation
assay. Subconfluent A2780 and OVCAR cells were seeded in plates and
treated with sulforaphane at the concentrations shown. After 48 h,
the cancer cells were fixed and stained with crystal violet for the
colony formation assay. (B) Colony-forming ability of ovarian
cancer cell lines following sulforaphane treatment was quantified
for optical absorbance. Cell viability assay. (C) A2780 and (D)
OVCAR cells were seeded in plates and treated with sulforaphane at
the concentrations shown. After 48 and 72 h, the viable cells were
collected, stained and counted under a bright field microscope.
WST-1 cell proliferation assay. (E) A2780 and (F) OVCAR cancer
cells were seeded in 96-well plates and treated with sulforaphane
at the indicated concentrations. At 24 h post-treatment, WST-1
reagent was added to plates and incubated for 1 h, and absorbance
measurement was performed. All assay conditions were performed in
triplicate. Cell wounding analysis of (G) A2780 cells with (H)
quantification, and (I) OVCAR cells with (J) quantification. Cells
were wounded with the micro-pipette tips and treated with
sulforaphane at the indicated concentrations. The wound gap was
measured at 0, 24 and 48 h post-sulforaphane treatments
(magnification, ×40). The proliferation assay indicated that
sulforaphane inhibited the proliferation of (K) A2780 and (L) OVCAR
ovarian cancer cells. (M) Transwell assay showed the metastasis
capacities were inhibited by sulforaphane in A2780 and OVCAR
ovarian cancer cells (magnification, ×40). (N) Quantification of
the migrated ovarian cancer cells. The values are presented as the
mean ± standard deviation (n=8-10) of the samples.
+P<0.05, ++P<0.01 and
+++P<0.001, vs. 0 μM group. SFN,
sulforaphane. |
Subsequently, the present study examined whether
sulforaphane has any effect on ovarian cancer cell migration or
wound healing. The monolayer cells of A2780 and OVCAR were wounded
and treated with different concentrations of sulforaphane. The
width of the gap of the wound defect, relative to the starting
width, was determined for A2780 cells at 24 and 48 h (Fig. 1G and H). The data suggested that
the scratch wound healing width was increased following 2.5, 5 and
10 μM sulforaphane administration. Similarly, compared with
the 0 h group without sulforaphane treatment, the scratch wound
healing width was altered significantly in the OVCAR cells in the
presence of sulforaphane at 24 and 48 h (Fig. 1I and J). These results indicated
that sulforaphane suppressed ovarian cancer cell migration and
proliferation in a dose-dependent manner.
To further examine the effects of sulforaphane on
ovarian cancer cell lines, MTT and Transwell assays of the
migration of cancer cells were performed for the A2780 and OVCAR
cells. The results suggested that, compared with the cells in the
absence of sulforaphane, the proliferation ability of the A2780
(Fig. 1K) and OVCAR (Fig. 1L) cells were significantly
inhibited by sulforaphane treatment at different concentrations.
The Transwell assays indicated that sulforaphane treatment
significantly suppressed the migrated ovarian cancer cells,
particularly at the highest concentration of sulforaphane (Fig. 1M and N). These results indicated
that administration with sulforaphane was able to inhibit the
proliferation, migration and metastasis of human ovarian cancer
cells.
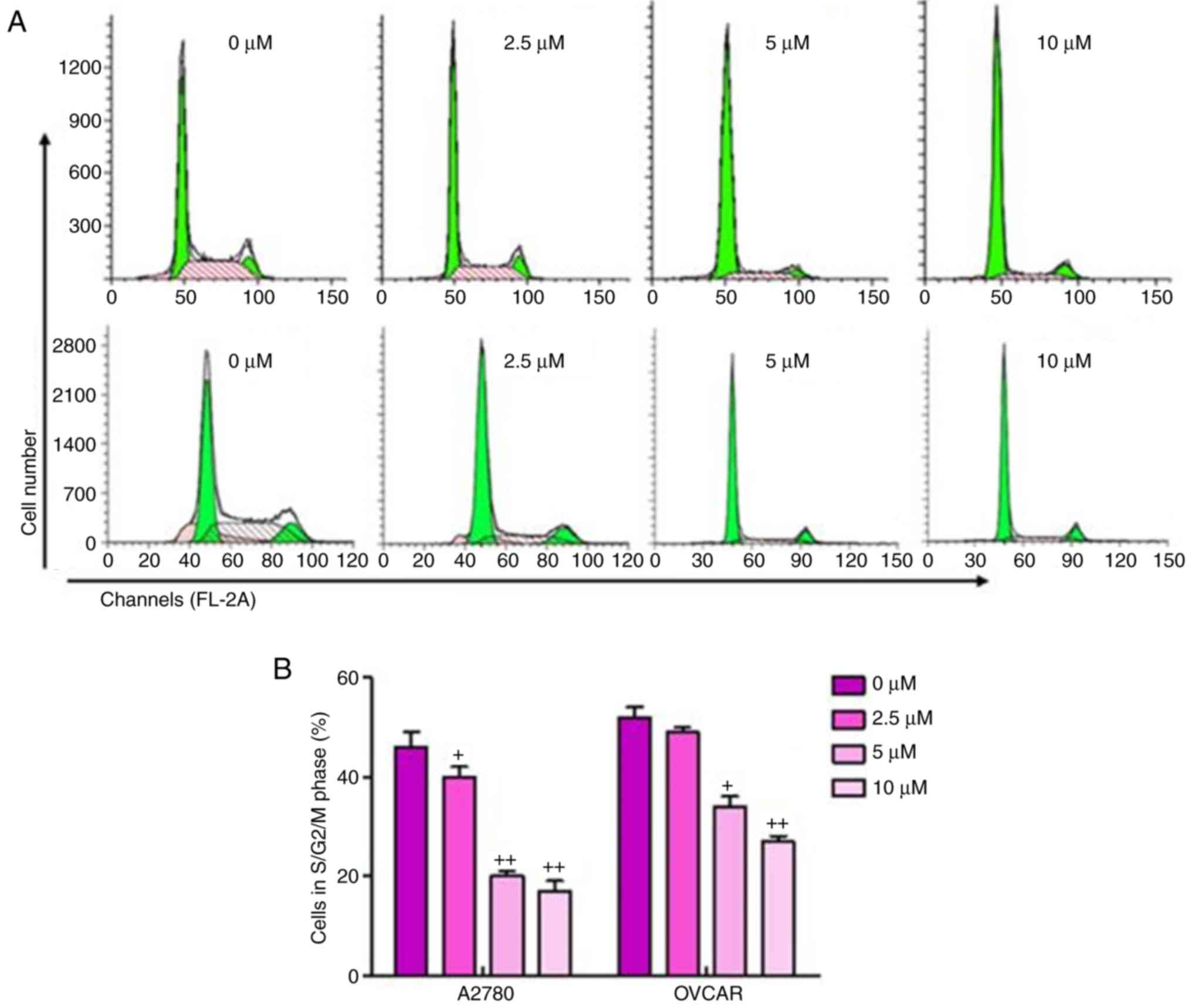
The present study also performed cell cycle analysis
on the sulforaphane-treated ovarian cancer cells and found a
significant upregulation in cancer cells arrested in the G1 phase,
and downregulation of cells in the S/M phase in
sulforaphane-treated A2780 and OVCAR cells, relative to the cells
without sulforaphane administration (Fig. 2A and B). These results suggested
that the inhibitory effect of sulforaphane on ovarian cancer cell
proliferation may be due, at least partly, to the inhibition of
cell cycle development and progression.
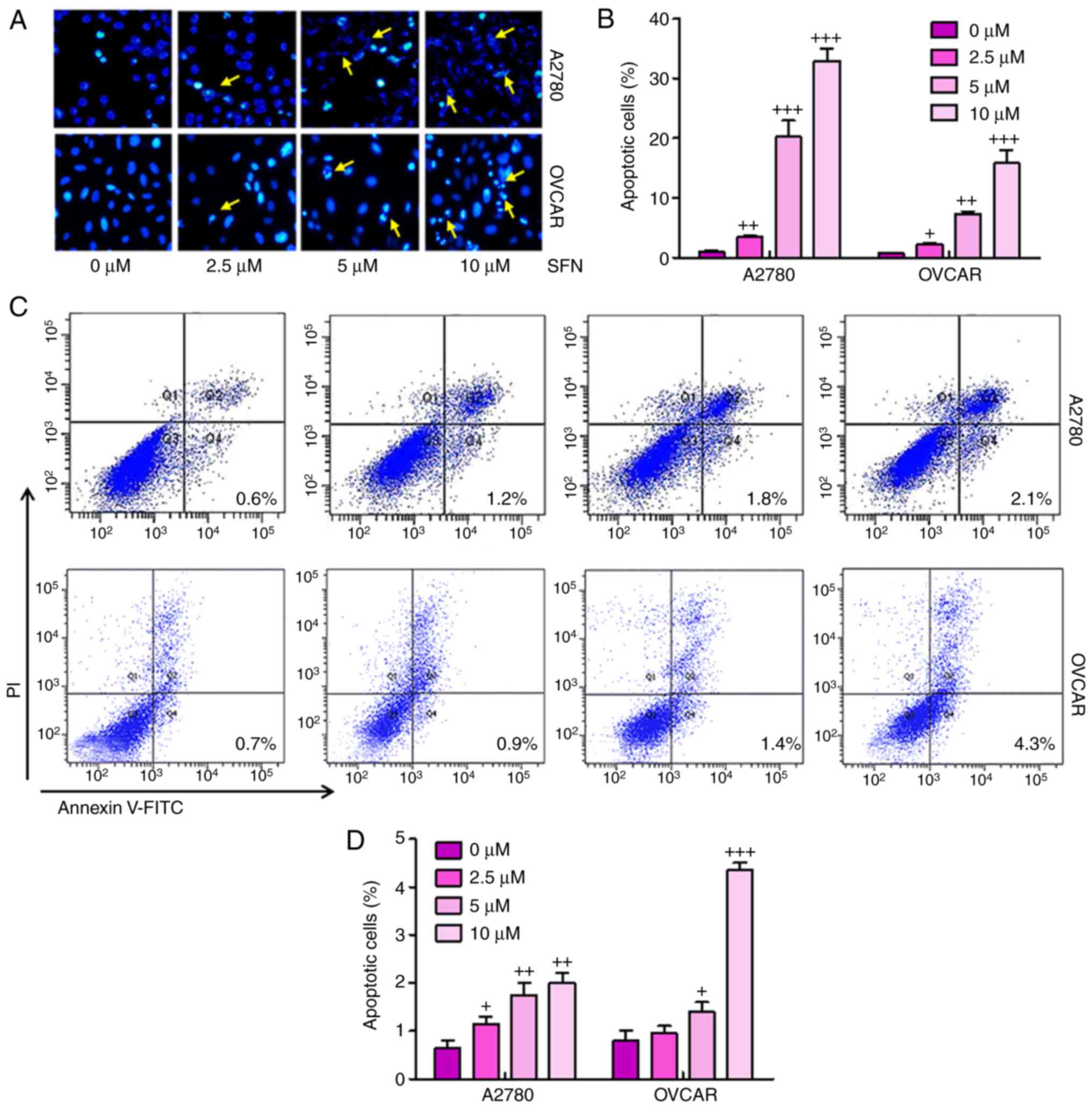
Sulforaphane suppresses ovarian cancer
progression via promotion of apoptosis
To illustrate the possible mechanisms underlying
sulforaphane-induced cell proliferation inhibition of ovarian
cancer, the present study examined whether sulforaphane can induce
apoptosis in ovarian cancer cell lines. When the proliferating
A2780 and OVCAR cells were treated with 2.5, 5 and 10 μM
sulforaphane for 24 h and then stained with Hoechst 33,258, the
numbers of apoptotic cancer cells were significant (Fig. 3A and B). Furthermore, flow
cytometry was used to evaluate the apoptotic levels in A2780 and
OVCAR cells. As shown in Fig. 3C and
D, the number of apoptotic cells were higher in the cells with
sulforaphane administration in a dose-dependent manner, indicating
that sulforaphane may be involved in enhancing apoptosis in ovarian
cancer development. Quantitative analysis of the ovarian cancer
cells indicated that the percentages of apoptotic cells were
significantly upregulated in the sulforaphane-treated A2780 and
OVCAR groups, with a significant difference. These data indicated
that the sulforaphane-induced suppression of A2780 and OVCAR cells
was due to the induction of apoptosis.
Sulforaphane inhibits multiple cancer
development-associated signaling pathways in A2780 cells
The high expression of Her2 is a marker for tumor
progression in the development of several types of cancer according
to previous studies (21). In
order to calculate whether there was a similar effect in ovarian
cancer cells, immunofluorescent assays were used to analyze the
expression levels of Her2 in different groups of A2780 cells. A
marked elevation in Her2 staining was observed in cells without
sulforaphane administration, and this was significantly suppressed
by sulforaphane in a dose-dependent manner (Fig. 4A). These results confirmed that
sulforaphane inhibited the expression of Her2 in ovarian cancer
cells, as previously indicated (22).
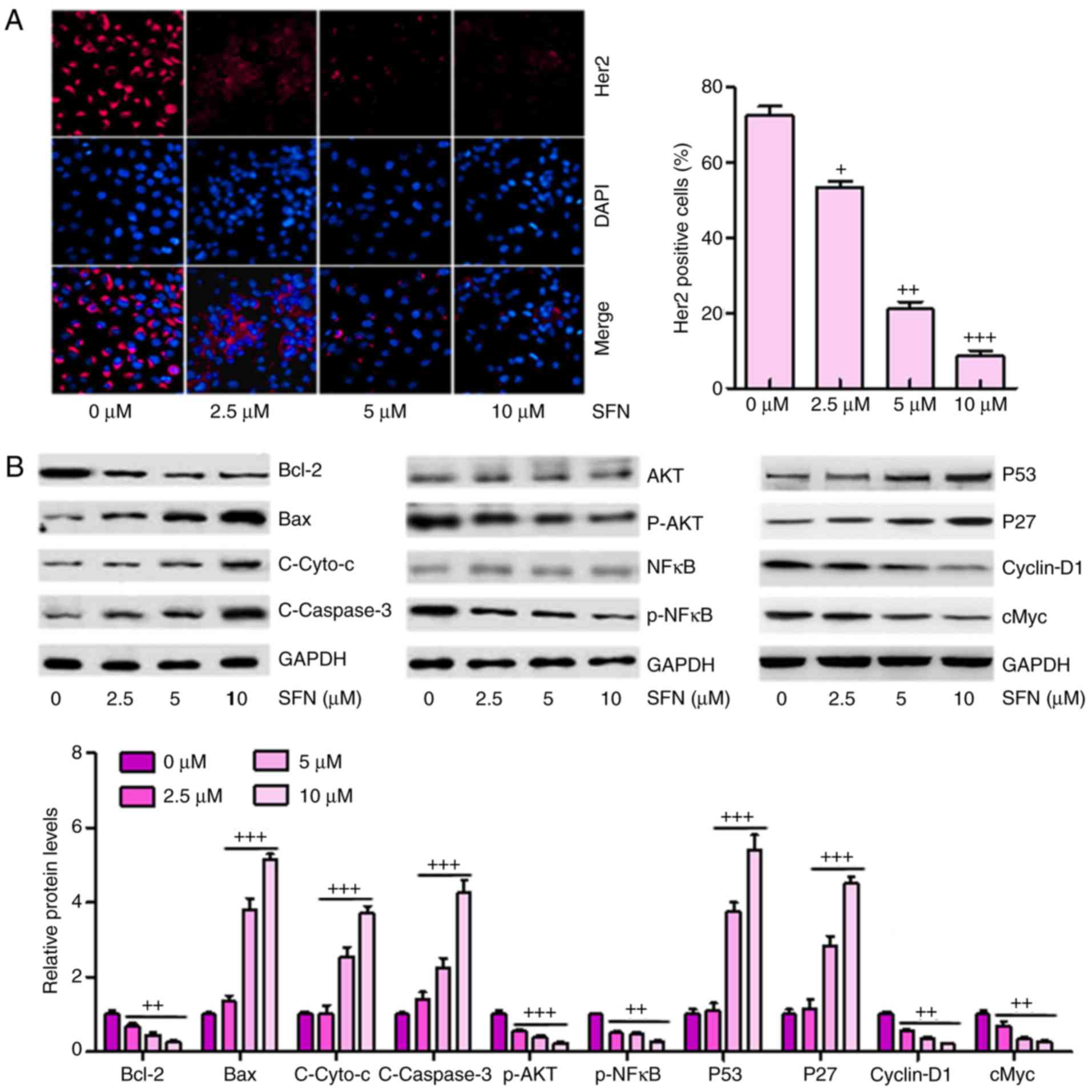 | Figure 4Sulforaphane inhibits multiple cancer
development-associated signaling pathways in A2780 cells. (A)
Effect of sulforaphane on intracellular levels of Her2 through
immunofluorescence assays (magnification, ×100). (B) Western blot
assays were performed to examine the effect of sulforaphane on the
10 cancer-associated signaling pathway reporter activities,
including Bcl-2, Bax, Cyto-c, Caspase-3, p-AKT, p-NF-κB, P53, P27,
cyclin-D1 and cMyc. The values are presented as the mean ± standard
deviation (n=8-10) of the samples. +P<0.05,
++P<0.01 and +++P<0.001, vs. 0
μM group. SFN, sulforaphane; Bcl-2, B-cell lymphoma 2; Bax,
Bcl-2-associated X protein; Cyto-c, cytochrome-c; NF-κB,
nuclear factor-κB; p-, phosphorylated. |
Subsequently, the present study attempted to
determine which, if any, cancer-involved signaling pathways were
regulated by sulforaphane. A total of 10 cancer-associated
signaling pathways were examined via western blot analysis
according to previous studies (23–25). It was found that Bcl-2, an
anti-apoptotic factor, was significantly inhibited under
sulforaphane administration. However, Bax, Cyto-c and Caspase-3
were upregulated in cells in the presence of different
concentrations of sulforaphane in a dose-dependent manner,
contributing to the apoptosis of cancer cells. In addition, AKT and
NF-κB, as important regulators of cell proliferation, were
inactivated in ovarian cancer cells under sulforaphane
administration. P53 and P27 were upregulated following sulforaphane
treatment in the A2780 cells, which suppressed cancer cell cycle,
consistent with a previous study (26). Furthermore, Cyclin-D1 and cMyc
were inhibited following sulforaphane induction at 2.5, 5 and 10
μM (Fig. 4B).
Sulforaphane effectively suppresses tumor
growth and progression in a xenograft model of human ovarian cancer
cells
The present study also examined the anticancer
effect of sulforaphane in an in vivo xenograft tumor model
of human ovarian cancer. A2780 cells in the presence or the absence
of sulforaphane at different concentrations were injected
subcutaneously into the flanks of athymic nude mice. Following
injection, tumor growth was observed and monitored for up to 42
days (Fig. 5A). Smaller tumor
volumes and tumor weights were found in the sulforaphane-treated
groups, compared with those in the groups without sulforaphane
administration (Fig. 5B and C).
Histologic evaluation was also performed on the retrieved tumor
samples. Immunohistochemical staining with the TUNEL, KI67 and Her2
antibodies indicated a significant increase in the number of
TUNEL-positive cells in the sulforaphane treatment groups,
particularly at the highest dose of sulforaphane, compared with the
control group (Fig. 5D). By
contrast, the expression levels of KI67 and Her2 expression were
reduced by sulforaphane treatment, which was consistent with
earlier results and support the hypothesis that Her2 may serve as a
key cellular target for sulforaphane in human ovarian cancer
inhibition. Finally, RT-qPCR analysis was used to confirm the tumor
progression-associated signals. Similar to the previous data, it
was found that Bcl-2, Cyclin-D1, cMyc and Her2 were significantly
downregulated following sulfora-phane administration. Bax, P53 and
P27 were upregulated following sulforaphane treatment.
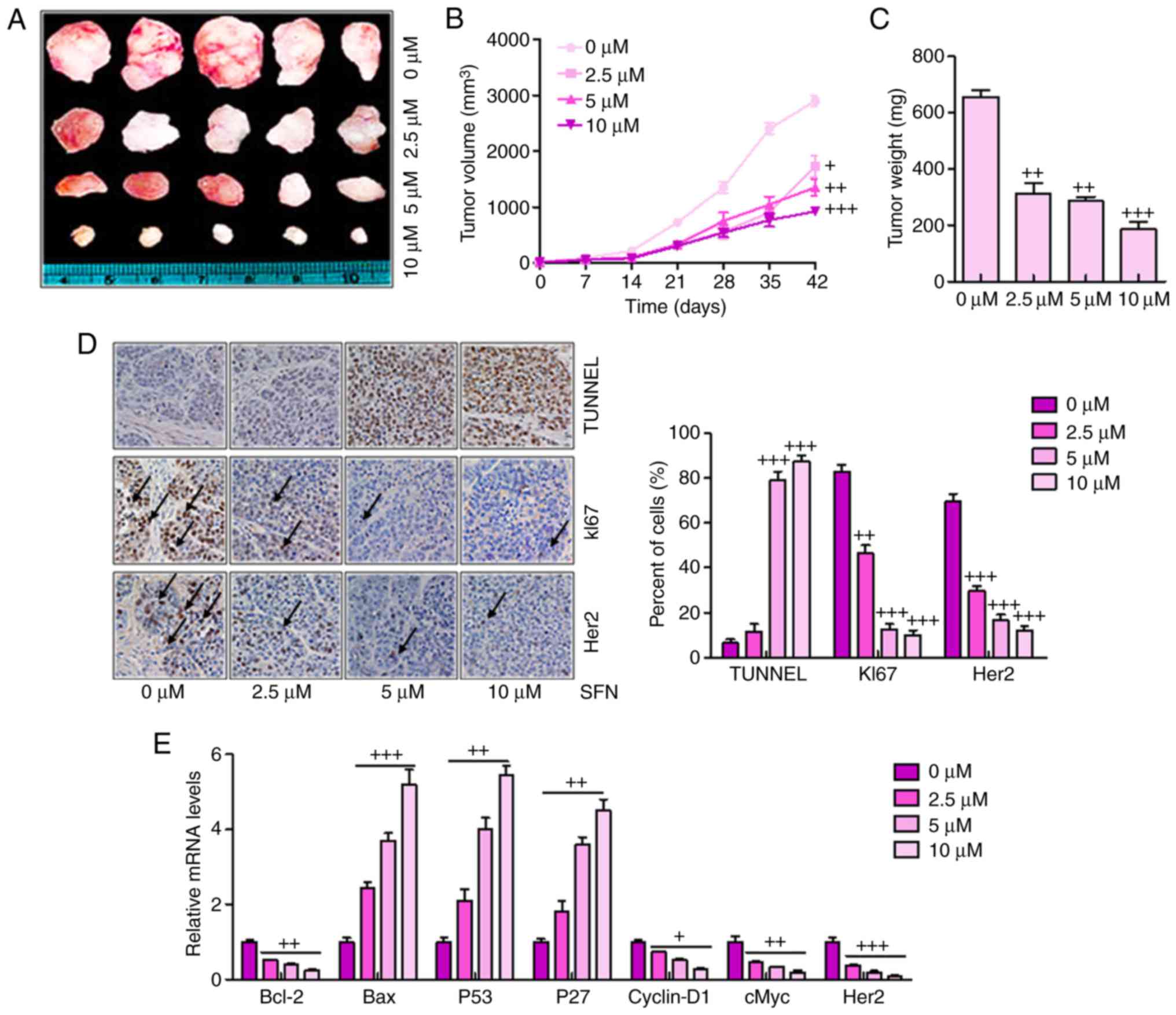 | Figure 5Sulforaphane effectively suppresses
tumor growth and progression in the xenograft model of human
ovarian cancer cells. (A) Tumor xenograft model. The A2780 cells
were injected into the hindlimbs of nude mice; and (B) tumor size
and (C) weight were observed and measured. (D) Analysis of TUNEL,
KI67 and Her2 in cancer tissues by immunohistochemistry. A brown
signal was considered positive staining for TUNEL, KI67 and Her2
(magnification, ×100). Arrows indicate representative positive
cells. (E) Reverse transcription-quantitative polymerase chain
reaction assays were used to examine the effect of sulforaphane on
the seven cancer-associated signaling pathway reporter activities,
including Bcl-2, Bax, P53, P27, Cyclin-D1, cMyc and Her2. The
values are presented as the mean ± standard deviation (n=8-10) of
the samples. +P<0.05, ++P<0.01 and
+++P<0.001, vs. 0 μM group. SFN, sulforaphane;
TUNEL, terminal deoxynucleotidyl transferase-mediated dUTP nick end
labeling; Her2, human epidermal growth factor receptor 2; Bcl-2,
B-cell lymphoma 2; Bax, Bcl-2-associated X protein. |
Sulforaphane synergizes with cisplatin in
suppressing cancer cell proliferation and promoting apoptosis in
ovarian cancer cells
If sulforaphane is to be targeted as an anti-ovarian
cancer agent, it is important that sulforaphane can synergize with
presently-used chemotherapeutic drugs, including cisplatin, in
order to suppress cancer cell proliferation and progression. When
the A2780 cells were administrated with different concentrations of
sulforaphane or cisplatin, ovarian cell proliferation inhibition
was observed in a dose-dependent manner (Fig. 6A), and this synergism was
confirmed by a colony formation assay (Fig. 6B and C). Additionally, western
blot analysis demonstrated that a combination of sulforaphane and
cisplatin further decreased the levels of Bcl-2, Cyclin-D1 and
cMyc, whereas P53 and cleaved Caspase-3 were significantly
upregulated in the group treated with the combination of
sulforaphane and cisplatin, promoting cell death (Fig. 6D and E). Therefore, these results
indicated that sulforaphane synergized with cisplatin by
suppressing proliferation and promoting apoptosis of human ovarian
cancer.
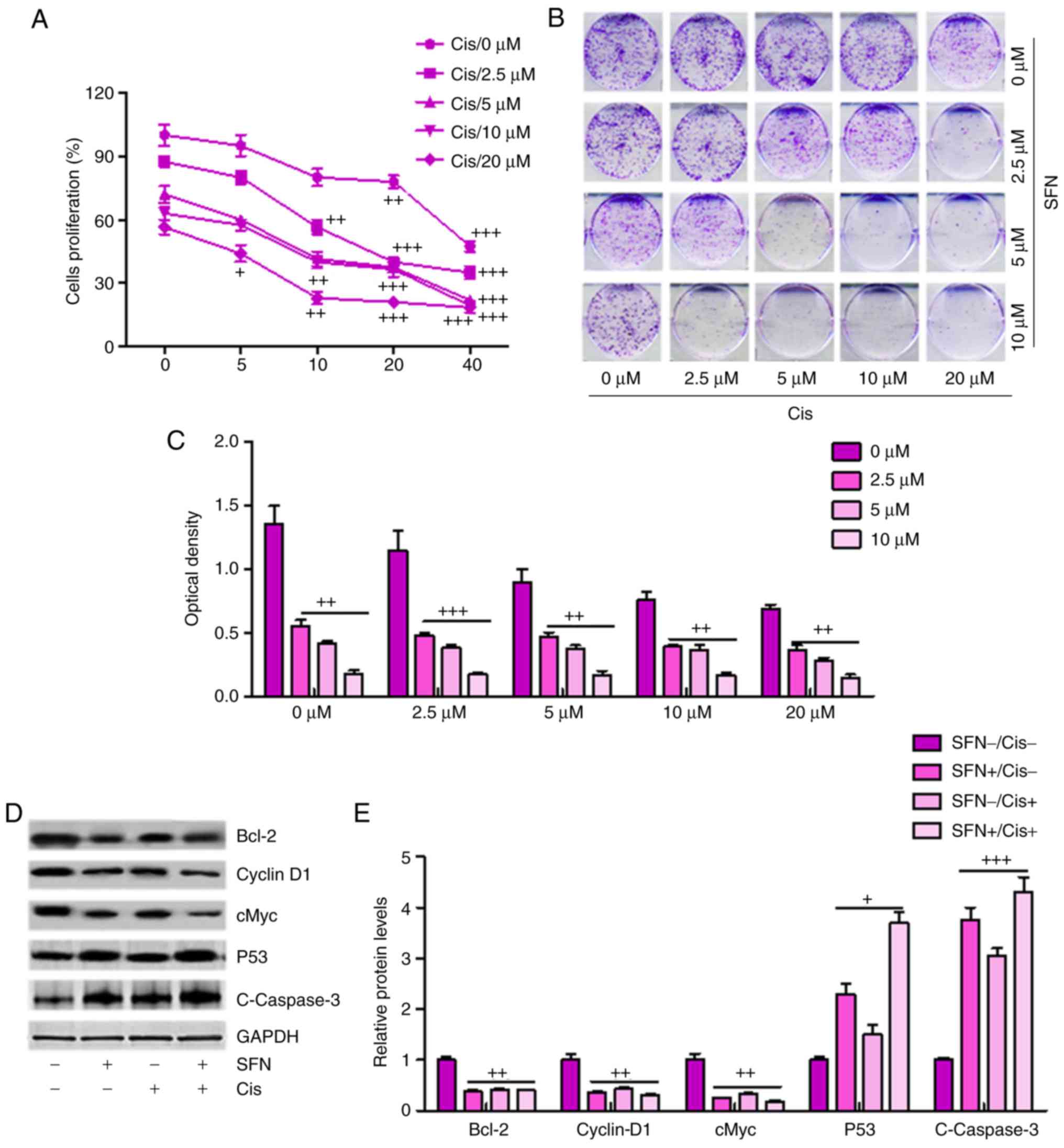 | Figure 6Sulforaphane synergizes with
cisplatin in suppressing cancer cell proliferation and promoting
apoptosis of ovarian cancer cells. (A) Synergism between
sulforaphane and cisplatin. A2780 cells were treated with
sulforaphane and cisplatin at the indicated concentrations. After
24 h, WST-1 reagent was administrated to the culture medium and
then incubated for 1 h. The WST-1 activities were determined at 440
nm. (B) Colony formation assay. Subconfluent A2780 cells were
seeded in plates and treated with sulforaphane and cisplatin at the
indicated concentrations. After 48 h, the cancer cells were then
fixed and stained with crystal violet for the colony formation
assay. (C) Colony-forming ability of ovarian cancer cells following
sulforaphane and cisplatin treatment was quantified for optical
absorbance. (D) Western blot assays with (E) quantification were
performed to determine protein levels of Bcl-2, Caspase-3, P53, and
Cyclin-D1 in A2780 cells treated with or without sulforaphane and
cisplatin. The values are presented as the mean ± standard
deviation (n=8-10) of the samples. +P<0.05,
++P<0.01 and +++P<0.001, vs. 0
μM group. SFN, sulforaphane; Cis, cisplatin; Bcl-2, B-cell
lymphoma 2. |
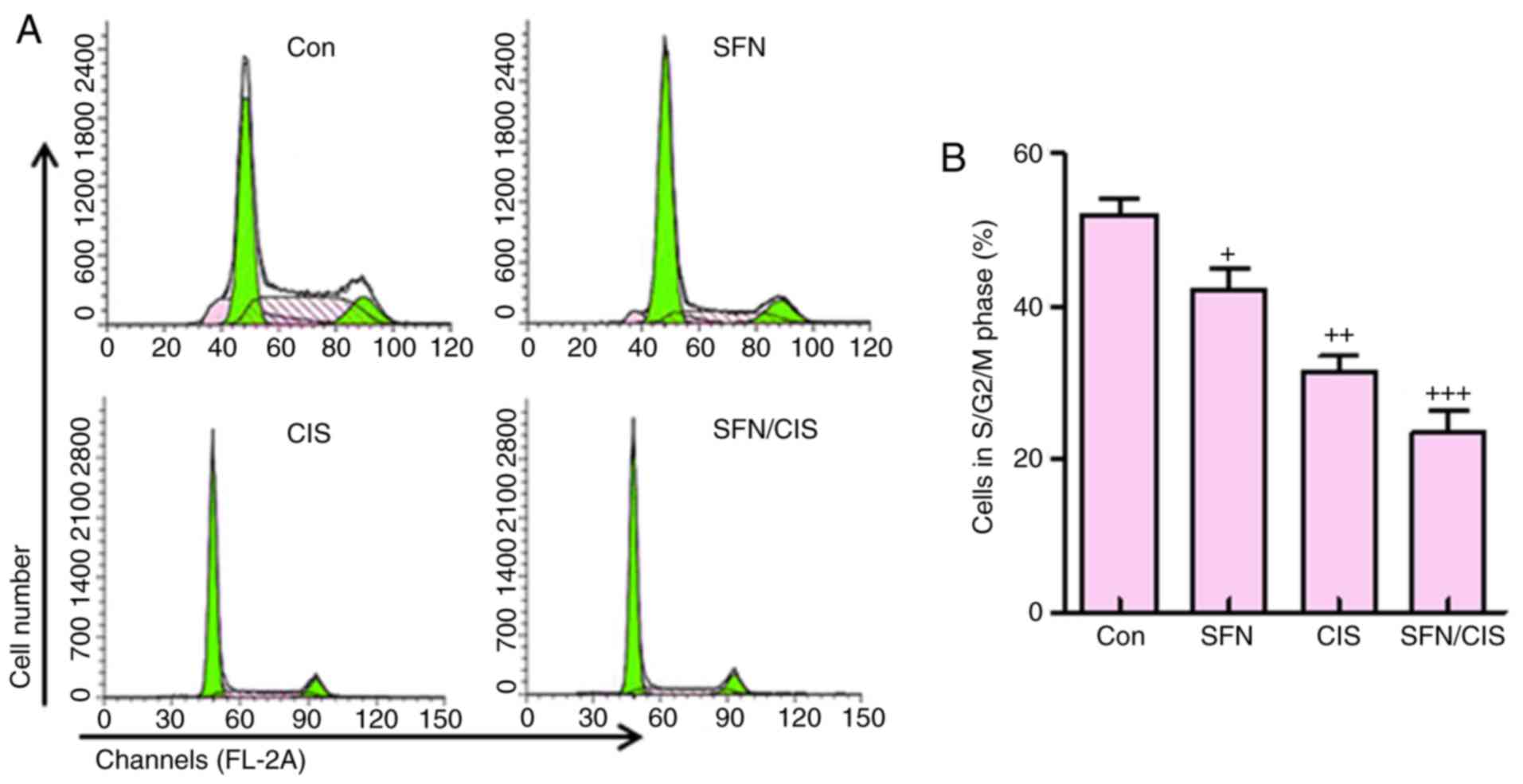
In addition, as shown in Fig. 7A and B, sulforaphane significantly
downregulated the proportion of cells in the S/G2/M phases,
compared with the control group, which was consistent with previous
data (8). Cisplatin further
reduced the number of cells in this phase. Of note, the combination
of sulforaphane and cisplatin exhibited the most marked effect on
the suppression of cells in the S/G2/M phase. These data suggested
that sulforaphane synergized with cisplatin to inhibit ovarian
cancer cell proliferation by regulating cell cycle.
Discussion
Among all types of gynecologic cancer, human ovarian
cancer is reported to be a life-threatening disease and a major
contributor to cancer-associated mortality rates in females across
the world (27,28). Human ovarian cancer is known to be
asymptomatic or exhibit vague symptoms during the early stage;
therefore, a large number of cases of ovarian cancer are detected
at advanced stages, with cancer cells spreading beyond the site of
the primary tumor (29).
Currently, the standard therapeutic treatment for human ovarian
cancer is limited to the combination of chemotherapy and surgery
(30,31). Therefore, understanding the
mechanism of metastasis and identifying novel strategies to inhibit
ovarian cancer is important for improving clinical outcomes.
Sulforaphane has been shown to suppress cell cycle
progression and induce apoptosis in pre-cancerous and tumor cells
of different origin (9,11,13–16). This anticancer agent at a
concentration of 75 μM was shown to cause G1/G2 cell cycle
arrest and induce apoptosis by downregulating the expression of
anti-apoptotic Bcl-2 and increasing the expression of
apoptosis-inducing Bax in colon cancer cells (32,33). In the present study, it was found
that the number of cells in the S/G2/M phase was reduced due to
sulforaphane administration, leading to the cell death in human
ovarian cancer. Additionally, apoptosis was induced as determined
via flow cytometry, and western blot analysis revealed the
downregulation of Bcl-2 and upregulation of Bax and cleaved
Caspase-3, which are important regulators contributing to
apoptosis. Therefore, the data suggested that sulforaphane
inhibited ovarian cancer progression through inducing
apoptosis.
Sulforaphane can also suppress the proliferation of
bladder cancer cells, leading to the inhibition of cell
proliferation and downregulated expression of NF-κB (34,35). In the present study, it was found
that sulforaphane was successful in suppressing ovarian cancer cell
proliferation. P53 and P27 are known essential antitumor factors.
The P53 tumor suppressor gene encodes a nuclear protein, which is
crucial in cell cycle regulation and major early events in cancer
progression (36). P27 is a
member of the Cip/Kip family of cyclin-dependent kinase inhibitors,
which can induce cell cycle arrest and serve as a tumor suppressor
(37). Cyclin-D1, a regulatory
kinase subunit that is selectively associated with cyclin-dependent
kinase 4, is a crucial modulator in the cell cycle of cancer or
tumor formation (38). The
oncogene cMyc is a known target gene, which is involved in cell
proliferation and cell cycle (39,40). In the present study, it was found
that sulforaphane suppressed the progression of ovarian cancer via
the promotion of P53 and P27. By contrast, Cyclin-D1 and cMyc were
downregulated following sulforaphane treatment. These data
suggested that the sulforaphane-inhibited progression of ovarian
cancer was associated with the upregulation of P53 and P27, and the
downregulation of Cyclin-D1 and cMyc. In addition, AKT and NF-κB
were suppressed following sulforaphane administration, contributing
to the apoptosis of ovarian cancer cells (41).
Of note, sulforaphane synergized with cisplatin to
further suppress human ovarian cancer. The results of the present
study indicated that sulforaphane had a potential role in
suppressing human ovarian cancer development via the inhibition of
proliferation and induction of apoptosis. However, following
combination with cisplatin, the cell proliferation was further
suppressed, compared with that following sulforaphane or cisplatin
treatment alone. In addition, western blot analysis indicated that
Bcl-2, Cyclin-D1 and cMyc were inhibited following treatment with
sulforaphane and cisplatin together. The levels of P53 and cleaved
Caspase-3 were significantly upregulated, compared with those in
the cells treated with sulforaphane or cisplatin alone.
Taken together, the present study investigated the
potential effect of repurposing sulforaphane as an essential
anticancer agent for the suppression of human ovarian cancer. The
results indicated that sulforaphane effectively suppressed ovarian
cancer cell proliferation, migration and cell cycle progression,
and induced apoptosis. Sulforaphane was found to target multiple
tumor- or cancer-related signaling pathways, including Bcl-2,
Caspase-3, AKT, NF-κB and P53/P27, and suppressed the expression of
Cyclin-D1 and cMyc in human ovarian cancer cells. Sulforaphane was
also found to act synergistically with the chemotherapeutic drug
cisplatin to suppress ovarian cancer cell proliferation and promote
apoptosis. The in vivo xenograft model experiments further
confirmed that sulforaphane effectively suppressed xenograft tumor
size and weight by inhibiting cancer cell proliferation via
reducing cancer-related signals. Therefore, the results indicated
that sulforaphane may be repurposed as an important anti-ovarian
cancer agent. However, investigations are required to further
examine the anticancer role of sulforaphane in preclinical and
clinical studies in the future.
Funding
Not applicable.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
JW and SK designed and analyzed the data. JW wrote
the manuscript. GS performed the flow cytometry experiments. SK
performed all the other experiments. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
The use and care of animals as models were approved
by the Institutional Animal Care and Use Committee of Zaozhuang
City Hospital (Zaozhuang, China). All experimental procedures were
performed according to the approved guidelines.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Kamieniak MM, Muñoz-Repeto I, Rico D,
Osorio A, Urioste M, García-Donas J, Hernando S, Robles-Díaz L and
Ramón Y: DNA copy number profiling reveals extensive genomic loss
in hereditary BRCA1 and BRCA2 ovarian carcinomas. Br J Cancer.
108:1732–1742. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pruthi S, Gostout BS and Lindor NM:
Identification and management of women with BRCA mutations or
hereditary predisposition for breast and ovarian cancer. Mayo Clin
Proc. 85:1111–1120. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chester C, Dorigo O, Berek JS and Kohrt H:
Immunotherapeutic approaches to ovarian cancer treatment. J
Immunother Cancer. 3:72015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gnjatic S, Ritter E, Büchler MW, Giese NA,
Brors B, Frei C, Murray A, Halama N, Zörnig I, Chen YT, et al:
Seromic profiling of ovarian and pancreatic cancer. Proc Natl Acad
Sci USA. 107:5088–5093. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Vaughan S, Coward JI, Bast RC Jr, Berchuck
A, Berek JS, Brenton JD, Coukos G, Crum CC, Drapkin R,
Etemadmoghadam D, et al: Rethinking ovarian cancer: Recommendations
for improving outcomes. Nat Rev Cancer. 11:719–725. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Frampton JE: Olaparib: A review of its use
as maintenance therapy in patients with ovarian cancer. BioDrugs.
29:143–150. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Okeke TC, Anyaehie UB and Ezenyeaku CC:
Premature menopause. Ann Med Health Sci Res. 3:90–95. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Suppipat K, Park CS, Shen Y, Zhu X and
Lacorazza HD: Sulforaphane induces cell cycle arrest and apoptosis
in acute lymphoblastic leukemia cells. PLoS One. 7:e512512012.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Clarke JD, Dashwood RH and Ho E:
Multi-targeted prevention of cancer by sulforaphane. Cancer Lett.
269:291–304. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Myzak MC, Hardin K, Wang R, Dashwood RH
and Ho E: Sulforaphane inhibits histone deacetylase activity in
BPH-1, LnCaP and PC-3 prostate epithelial cells. Carcinogenesis.
27:811–819. 2006. View Article : Google Scholar
|
|
11
|
Ho E, Clarke JD and Dashwood RH: Dietary
sulforaphane a histone deacetylase inhibitor for cancer prevention.
J Nutr. 139:2393–2396. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Su ZY, Zhang C, Lee JH, Shu L, Wu TY, Khor
TO, Conney AH, Lu YP and Kong AN: Requirement and epigenetics
reprogramming of Nrf2 in suppression of tumor promoter TPA-induced
mouse skin cell transformation by sulforaphane. Cancer Prev Res.
7:319–329. 2014. View Article : Google Scholar
|
|
13
|
Atwell LL, Hsu A, Wong CP, Stevens JF,
Bella D, Yu TW, Pereira CB, Löhr CV, Christensen JM, Dashwood RH,
et al: Absorption and chemopreventive targets of sulforaphane in
humans following consumption of broccoli sprouts or a
myrosinase-treated broccoli sprout extract. Mol Nutr Food Res.
59:424–433. 2015. View Article : Google Scholar :
|
|
14
|
Ho E, Beaver LM, Williams DE and Dashwood
RH: Dietary factors and epigenetic regulation for prostate cancer
prevention. Adv Nutr. 2:497–510. 2011. View Article : Google Scholar :
|
|
15
|
Hsu A, Wong CP, Yu Z, Williams DE,
Dashwood RH and Ho E: Promoter de-methylation of cyclin D2 by
sulforaphane in prostate cancer cells. Clin Epigenet. 3:32011.
View Article : Google Scholar
|
|
16
|
Li C, Zhou Y, Peng X, Du L, Tian H, Yang
G, Niu J and Wu W: Sulforaphane inhibits invasion via activating
ERK1/2 signaling in human glioblastoma U87MG and U373MG cells. PLoS
One. 9:e905202014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fimognari C, Turrini E, Sestili P,
Calcabrini C, Carulli G, Fontanelli G, Rousseau M, Cantelli-Forti G
and Hrelia P: Antileukemic activity of sulforaphane in primary
blasts from patients affected by myelo- and lympho-proliferative
disorders and in hypoxic conditions. PLoS One. 9:e1019912014.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sharma C, Sadrieh L, Priyani A, Ahmed M,
Hassan AH and Hussain A: Anti-carcinogenic effects of sulforaphane
in association with its apoptosis-inducing and anti-inflammatory
properties in human cervical cancer cells. Cancer Epidemiol.
35:272–278. 2011. View Article : Google Scholar
|
|
19
|
Ferreira de Oliveira JM, Remédios C,
Oliveira H, Pinto P, Pinho F, Pinho S, Costa M and Santos C:
Sulforaphane induces DNA damage and mitotic abnormalities in human
osteosarcoma MG-63 cells: Correlation with cell cycle arrest and
apoptosis. Nutr Cancer. 66:325–334. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCT method. Methods. 25:402–408. 2001. View Article : Google Scholar
|
|
21
|
Tuefferd M, Couturier J, Penault-Llorca F,
Vincent-Salomon A, Broët P, Guastalla JP, Allouache D, Combe M,
Weber B, Pujade-Lauraine E, et al: HER2 status in ovarian
carcinomas: A multicenter GINECO study of 320 patients. PLoS One.
2:e11382007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kang HS, Huh YM, Kim S and Lee DK:
Isolation of RNA aptamers targeting HER2-overexpressing breast
cancer cells using cell-SELEX. Bull Korean Chem Soc. 30:1827–1831.
2009. View Article : Google Scholar
|
|
23
|
Chen H, Landen CN, Li Y, Alvarez RD and
Tollefsbol TO: Epigallocatechin gallate and sulforaphane
combination treatment induce apoptosis in paclitaxel-resistant
ovarian cancer cells through hTERT and Bcl-2 down-regulation. Exp
Cell Res. 319:697–706. 2003. View Article : Google Scholar
|
|
24
|
Xu C, Shen G, Chen C, Gélinas C and Kong
AN: Suppression of NF-κB and NF-κB-regulated gene expression by
sulforaphane and PEITC through IjBa, IKK pathway in human prostate
cancer PC-3 cells. Oncogene. 24:4486–4495. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lee WK and Kim SJ: Sulforaphane regulates
differentiation via PI-3K/AKT pathway in human chondrosarcoma cell
line, HTB-94 cells. Cancer Prev Res. 18:26–32. 2013.
|
|
26
|
Lubecka-Pietruszewska K, Kaufman-Szymczyk
A, Stefanska B, Cebula-Obrzut B, Smolewski P and
Fabianowska-Majewska K: Clofarabine, a novel adenosine analogue,
reactivates DNA methylation-silenced tumour suppressor genes and
inhibits cell growth in breast cancer cells. Eur J Pharmacol.
723:276–287. 2014. View Article : Google Scholar
|
|
27
|
Suh DH, Lee KH, Kim K, Kang S and Kim JW:
Major clinical research advances in gynecologic cancer in 2014. J
Gynecol Oncol. 26:156–167. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mahajan N: Fertility preservation in
female cancer patients: An overview. J Hum Reprod Sci. 8:3–13.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kamieniak MM, Muñoz-Repeto I, Rico D,
Osorio A, Urioste M, García-Donas J, Hernando S, Robles-Díaz L,
Ramón Y, Cajal T, Cazorla A, et al: DNA copy number profiling
reveals extensive genomic loss in hereditary BRCA1 and BRCA2
ovarian carcinomas. Br J Cancer. 108:1732–1742. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Moyer V; The U.S. Preventive Services Task
Force: Risk assessment, genetic counseling, and genetic testing for
BRCA-related cancer in women: U.S. Preventive Services Task Force
recommendation statement. Ann Intern Med. 160:271–281. 2014.
|
|
31
|
Jablonska K, Pula B, Zemla A, Kobierzycki
C, Kedzia W, Nowak-Markwitz E, Spaczynski M, Zabel M,
Podhorska-Okolow M and Dziegiel P: Expression of the MT1 melatonin
receptor in ovarian cancer cells. Int J Mol Sci. 15:23074–23089.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Dexheimer TS, Daekyu S and Hurley LH:
Deconvoluting the structural and drug-recognition complexity of the
G-quadruplex-forming region upstream of the bcl-2 P1 promoter. J Am
Chem Soc. 128:5404–5415. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chen T and Wong Y: Selenocystine induces
reactive oxygen species-mediated apoptosis in human cancer cells.
Biomed Pharmacother. 63:105–113. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kallifatidis G, Rausch V, Baumann B, Apel
A, Beckermann BM, Groth A, Mattern J, Li Z, Kolb A, Moldenhauer G,
et al: Sulforaphane targets pancreatic tumour-initiating cells by
NF-κB-induced antiapoptotic signaling. Gut. 58:949–963. 2009.
View Article : Google Scholar
|
|
35
|
Ju W, Wang X, Shi H, Chen W, Belinsky SA
and Lin Y: A critical role of luteolin-induced reactive oxygen
species in blockage of tumor necrosis factor-activated nuclear
factor κB pathway and sensitization of apoptosis in lung cancer
cells. Mol Pharmacol. 71:1381–1388. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bardeesy N, Aguirre AJ, Chu GC, Cheng KH,
Lopez LV, Hezel AF, Feng B, Brennan C, Weissleder R, Mahmood U, et
al: Both p16Ink4a and the p19Arf-p53 pathway
constrain progression of pancreatic adenocarcinoma in the mouse.
Proc Natl Acad Sci USA. 103:5947–5952. 2006. View Article : Google Scholar
|
|
37
|
Psyrri A, Bamias A, Yu Z, Weinberger PM,
Kassar M, Markakis S, Kowalski D, Efstathiou E, Camp RL, Rimm DL,
et al: Subcellular localization and protein levels of
cyclin-dependent kinase inhibitor p27 independently predict for
survival in epithelial ovarian cancer. Clin Cancer Res.
11:8384–8390. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhou X, Zhang Z, Yang X, Chen W and Zhang
P: Inhibition of cyclin D1 expression by cyclin D1 shRNAs in human
oral squamous cell carcinoma cells is associated with increased
cisplatin chemosensitivity. Int J Cancer. 124:483–489. 2009.
View Article : Google Scholar
|
|
39
|
Biliran H Jr, Wang Y, Banerjee S, Xu H,
Heng H, Thakur A, Bollig A, Sarkar FH and Liao JD: Overexpression
of cyclin D1 promotes tumor cell growth and confers resistance to
cisplatin-mediated apoptosis in an elastase-myc
transgene-expressing pancreatic tumor cell line. Clin Cancer Res.
11:6075–6086. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Luo H, Rankin GO, Juliano N, Jiang BH and
Chen YC: Kaempferol inhibits VEGF expression and in vitro
angiogenesis through a novel ERK-NFκB-cMyc-p21 pathway. Food Chem.
130:321–328. 2012. View Article : Google Scholar
|
|
41
|
Chen T and Wong YS: Selenocystine induces
S-phase arrest and apoptosis in human breast adenocarcinoma MCF-7
cells by modulating ERK and Akt phosphorylation. J Agric Food Chem.
56:10574–10581. 2008. View Article : Google Scholar : PubMed/NCBI
|