Introduction
Atherosclerosis, a chronic inflammatory disease, is
an important pathological process involved in peripheral artery
disease, coronary artery disease and stroke, and has been
attracting research interest during recent years (1,2).
A substantial number of studies have indicated that the NLR family
pyrin domain containing 3 (NLRP3) inflammasome activation is
implicated in the vascular inflammatory response driving the
development and progression of atherosclerosis (3,4).
Upon activation, NLRP3 interacts with adapter apoptosis-associated
speck-like protein containing a C-terminal caspase recruitment
domain (ASC) to form the NLRP3 inflammasome, resulting in caspase-1
activation and the release of proinflammatory cytokines [i.e.,
interleukin (IL)-1β and IL-18], a process referred to as NLRP3
inflammasome activation-mediated pyroptosis (5-7),
which has been reported as an important event in endothelial injury
(8,9) and the pathophysiological process of
atherosclerosis (10,11). Therefore, mitigation of NLRP3
inflammasome activation-mediated pyroptosis may be a promising
therapeutic strategy for reversing endothelial injury in
atherosclerosis.
Oxymatrine, a quinolizidine alkaloid isolated from
the traditional Chinese herb Sophora flavescens Aiton,
possesses multiple pharmacological properties, such as
anti-inflammatory, antioxidant, antifibrotic and cardiovascular
protective effects (12-14). To date, oxymatrine has been
extensively discussed for its protective effects against heart
injury induced by doxorubicin (14), chronic heart failure (15), aldosterone (16) and hypertension (17). Recent studies have claimed that
oxymatrine inhibits homocysteine-induced human umbilical vein
endothelial cell (HUVEC) apoptosis/death, attenuating endothelial
cell dysfunction (a hallmark of atherosclerosis) (18,19), thereby providing new insight into
the role of oxymatrine in the prevention of atherosclerosis;
however, the precise mechanisms have yet to be fully elucidated.
Hence, the present study was undertaken to investigate the
protective effect of oxymatrine against oxidized low-density
lipoprotein (ox-LDL)-induced HUVEC injury, a typical in
vitro model of atherosclerosis (20), and further demonstrated the role
of NLRP3 inflammasome activation-mediated pyroptosis in this
process.
Sirtuin (SIRT) 1, a member of the sirtuin family of
NAD+-dependent deacetylases, has long been considered as
an anti-atherosclerosis factor (21-23). SIRT1 can activate the nuclear
factor-erythroid 2-related factor 2 (Nrf2) signaling pathway, and
the activation of the Nrf2 pathway has been shown to attenuate
endothelial cell injury, inhibit inflammation and exert
anti-atherosclerotic effects (24,25). Once activated, the Nrf2 signaling
pathway can also inhibit NLRP3-mediated pyroptosis (26,27). However, the exact effect of the
SIRT1/Nrf2 signaling pathway on NLRP3 inflammasome
activation-mediated pyroptosis in atherosclerosis has yet to be
reported. Notably, it has been confirmed that oxymatrine can
activate the SIRT1 signaling pathway (28,29). Hence, the present study further
explored the role of the SIRT1/Nrf2 signaling pathway in the
protective effect of oxymatrine against ox-LDL injury and the
inhibitory effect of oxymatrine on NLRP3 inflammasome
activation-mediated pyroptosis in HUVECs.
The results of the present study may improve our
understanding of the protective properties of oxymatrine agaisnt
ox-LDL injury, indicating potential novel approaches to the
treatment of atherosclerosis.
Materials and methods
Materials
ox-LDL, 2′,7′-dichlorofluorescein diacetate
(DCFH-DA) and dimethyl sulfoxide (DMSO) were purchased from
Sigma-Aldrich; Merck KGaA. Oxymatrine (purity, >98%) was
obtained from Green Valley Pharmaceutical Co., Ltd. A stock
suspension of oxymatrine (50 mg/ml) was prepared by dilution with
PBS and stored at 4°C. Dulbecco’s modified Eagle’s medium (DMEM;
cat. no. SH30022.01B) was purchased from HyClone; Cytiva. Fetal
bovine serum (FBS; cat. no. 16140071), doxorubicin and MTT assay
kit (cat. no. V13154) were purchased from Gibco; Thermo Fisher
Scientific, Inc. Annexin V-FITC/PI Apoptosis Detection kit was
obtained from Shanghai Yisheng Technology Co., Ltd. (cat. no.
40302ES50). TNF-α assay kit (cat. no. H052), IL-1β assay kit (cat.
no. H002), IL-6 assay kit (cat. no. H007) and IL-10 assay kit (cat.
no. H009) were obtained from Nanjing Jiangcheng Bioengineering
Institute. Anti-Bax (product no. 2772), anti-Bcl-2 (product no.
15071), anti-NLRP3 (product no. 13158), anti-cleaved caspase-1
(product no. 89332), anti-caspase-1 (product no. 24232) anti-IL-1β
(product no. 12703), anti-IL-18 (product no. 54943), anti-SIRT1
(product no. 8469), anti-Nrf2 (product no. 12721), anti-heme
oxygenase (HO)-1 (product no. 43966), anti-cleaved caspase-3
(product no. 9661), anti-caspase-3 (product no. 9662), anti-GAPDH
(product no. 5174) and anti-histone H3 (product no. 4499)
antibodies were purchased from Cell Signaling Technology, Inc.
Small interfering (si)RNA for NLRP3 (siNLRP3; cat. no. sc-45469)
and control siRNA-B (siNC; cat. no. sc-44230), siRNA for SIRT1
(siSIRT1; cat. no. sc-40986) and control siRNA-A (siCon; cat. no.
sc-37007) were purchased from Santa Cruz Biotechnology, Inc.
HUVEC culture and treatment
HUVECs were purchased from the American Type Culture
Collection and cultured in DMEM supplemented with 10% (v/v) FBS and
1% penicillin/streptomycin at 37°C in a humidified 5%
CO2 incubator. The culture medium was replaced every 1-2
days. HUVECs were treated with ox-LDL (100 µg/ml) for 24 h
at 37°C to mimic atherosclerotic conditions in vitro
(30). To detect the possible
protective effects of oxymatrine against ox-LDL injury, HUVECs were
pre-treated with oxymatrine (2, 4 or 8 µM) for 1 h prior to
ox-LDL (100 µg/ml) challenge for 24 h at 37°C. To
investigate the role of NLRP3 inflammasome activation-mediated
pyroptosis in the protective effect of oxymatrine against
ox-LDL-induced injury, HUVECs were transfected with siRNAs for
NLRP3 (siNLRP3) followed by oxymatrine (4 µM) for 1 h prior
to ox-LDL (100 µg/ml) challenge for 24 h. To demonstrate the
function of the SIRT1/Nrf2 signaling pathway in the protective
effect of oxymatrine against ox-LDL injury, HUVECs were transfected
with siRNAs for SIRT1 (siSIRT1) followed by oxymatrine (4
µM) for 1 h prior to ox-LDL (100 µg/ml) challenge for
24 h. The dose of oxymatrine selected was according to our pilot
study (unpublished data).
siRNA transfection
HUVECs were incubated in 6-well plates at 37°C
overnight at a density of 1×105 cells/well, and then
transfected with siRNAs for NLRP3 (siNLRP3, 50 nM) or siRNA for
SIRT1 (siSIRT1, 50 nM), as well as a respective negative control
(siNC, 50 nM) using Lipofectamine® 2000 reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) following the
manufacturer’s protocol. The transfection sequences were as
follows: siNLRP3 sense, 5′-GCU UCA GCC ACA UGA CUU UTT-3′ and
antisense, 5′-AAA GUC AUG UGG CUG AAG CTT-3′; siNC sense, 5′-UUC
UCC GAA CGU GUC ACG UTT-3′ and antisense, 5′-ACG UGA CAC GUU CGG
AGA ATT-3′; siSIRT1 sense, 5′-UUG GGA UUC ACG CAU AGG AGC ACUG-3′
and antisense, 5′-CAG UGC UCC UAU GCG GAA UCC CAA-3′; and siRNA for
control (siCon), sense 5′-UUC UCC GAA CGU GUC ACG UdT dT-3′ and
antisense 5′-ACG UGA CAC GUU CGG AGA AdT dT-3′. In brief, siNLRP3
(50 nM), siSIRT1 (50 nM), siNC (50 nM) or siCon (50 nM) was mixed
with Lipofectamine® 2000 transfection reagent, and the
mixture was added to the cells and incubated for 6 h at 37°C. Then
the medium was replaced with complete medium and the cells were
cultured for 24 h, following which the cells were collected for
subsequent experiments. The transfection efficiency of siNLRP3 and
siSIRT1 in HUVECs was detected at 24 h after transfection by
western blotting.
Cell viability assay
The viability of HUVECs was measured by Cell
Counting Kit-8 (CCK-8) assay. In brief, HUVECs were grown in
96-well plates and treated as aforementioned. CCK-8 solution (10
µl; cat. no. C0038; Beyotime Biotechnology) was added to
each well and co-incubated for 3 h at 37°C. The optical density
(OD) at 450 nm was measured using a microplate reader (Thermo
Fisher Scientific, Inc.).
Lactate dehydrogenase (LDH) assay
The cell death was further detected using an LDH
Cytotoxicity Assay kit (cat. no. C0017; Beyotime Biotechnology)
according to the manufacturer’s protocol. The LDH release was
measured at 490 nm using a microplate reader (Bio-Rad Laboratories,
Inc.).
Apoptosis assay
The apoptosis ratios of HUVECs were measured using
Annexin V-FITC/PI Apoptosis Detection kit by flow cytometry
according to manufacturer’s instructions. Briefly, after treatment,
HUVECs were collected, washed twice with PBS, and incubated with
Annexin V-FITC (10 µl) and PI (5 µl) for 20 min at
room temperature in the dark. The mixture was detected by flow
cytometric analysis using a flow cytometer (FACSCalibur™; BD
Biosciences), and the data were analyzed using FlowJo 7.6 software
(Tree Star, Inc.).
Determination of reactive oxygen species
(ROS) production
The production of intracellular ROS was detected by
the DCFH-DA fluorescence probe according to the manufacturer’s
protocol. In brief, the treated HUVECs were harvested and washed
twice with PBS, and then incubated with DCFH-DA (10 µM) for
20 min in the dark at 37°C. After washing again with PBS, images
were captured under a fluorescence microscope (80i; Nikon
Corporation). In addition, the mean fluorescence intensity of ROS
was analyzed and averaged using flow cytometry (FACSCalibur™), and
the data were analyzed by FlowJo 7.6 software (Tree Star,
Inc.).
Determination of malondialdehyde (MDA)
content, super-oxide dismutase (SOD), catalase (CAT) and
glutathione peroxidase (GSH-Px) activities
After treatment, HUVECs were harvested, washed with
ice-cold PBS, and incubated with the lysis buffer provided in the
assay kits for 30 min on ice. Subsequently, cell lysates were
centrifuged at 10,000 × g for 10 min at 4°C, the supernatant was
collected and the protein content was measured using bicinchoninic
acid (BCA) protein assay kit (Beyotime Biotechnology). The MDA
content (cat. no. A003-1-2), SOD (cat. no. A001-3-2), CAT (cat. no.
A007-1-1) and GSH-Px (cat. no. A005-1-2) activities were determined
by commercial kits from Nanjing Jiancheng Bioengineering Institute
according to the manufacturers’ protocol.
Measurement of mitochondrial membrane
potential (MMP)
The MMP was monitored using the JC-1 dye kit
according to the manufacturers’ protocol (cat. no. C2006; Beyotime
Biotechnology). Briefly, HUVECs were harvested, washed twice with
PBS, and then incubated with JC-1 for 20 min at 37°C in the dark.
The data were analyzed and averaged using flow cytometry
(FACSCalibur™), and the data were analyzed by FlowJo 7.6 software
(Tree Star, Inc.).
Western blot analysis
Total protein from HUVECs was isolated using RIPA
lysis buffer (Beyotime Biotechnology) containing 1% (v/v) protease
inhibitor cocktail (Roche Diagnostics). The cytoplasmic and nuclear
proteins of HUVECs were extracted with a cytoplasmic and nuclear
extraction kit (cat. no. CW0199), according to the manufacturer’s
instructions (CWBIO). The protein concentration was measured with a
BCA protein assay kit. Equal amounts of protein (30 µg) were
separated by 12% SDS-PAGE and then transferred onto PVDF membranes.
After blocking with 5% (w/v) skimmed dry milk powder dissolved in
Tris-buffered saline containing 0.1% (v/v) Tween-20 for 2 h at room
temperature, the membranes were incubated with primary antibodies
against Bax (1:2,000), Bcl-2 (1:2,000), NLPR3 (1:1,000), ASC
(1:1,000), cleaved caspase-1 (1:1,000), caspase-1 (1:2,000) IL-1β
(1:1,000), IL-18 (1:1,000), SIRT1 (1:2,000), Nrf2 (1:1,000), HO-1
(1:1,000), cleaved caspase-3 (1:2,000), caspase-3 (1:1,000), GAPDH
(1:2,000) and histone H3 (1:2,000) overnight at 4°C. GAPDH was used
as an internal control. Then, the membranes were incubated with
HRP-conjugated secondary antibodies (1:5,000; product no. 7074;
Cell Signaling Technology, Inc.) for 2 h at room temperature and
the probed protein was visualized by Beyo ECL Plus (cat. no. P0018,
Beyotime Biotechnology). The densitometric analysis was
semi-quantified with ImageJ 1.43 software (National Institutes of
Health).
Statistical analysis
Statistical analysis was performed by GraphPad Prism
5 (GraphPad Software, Inc.) using one-way analysis of variance
(ANOVA) followed by Tukey’s post hoc test. All the data are
presented as the mean ± SD. All experiments were repeated at least
three times. P<0.05 was considered to indicate a statistically
significant difference.
Results
Oxymatrine alleviates ox-LDL-induced
HUVEC injury
To assess the protective effect of oxymatrine
against ox-LDL-induced injury in HUVECs, cells were pre-treated
with varying concentrations of oxymatrine in the presence or
absence of ox-LDL (100 µg/ml). The results revealed that
oxymatrine (2, 4 or 8 µM) significantly reversed the
ox-LDL-induced decrease in cell viability (Fig. 1A). Treatment with oxymatrine
alone (2, 4 or 8 µM) did not affect cell viability compared
with the control group. The results revealed that oxymatrine (4
µM) obviously increased the viability of HUVECs treated with
ox-LDL (P<0.01), which was selected as an optimal dose for
interference with HUVECs treated with ox-LDL in a follow-up
experiment. Subsequently, the result confirmed that oxymatrine (4
µM) significantly attenuated the ox-LDL-induced increase in
LDH release in HUVECs (Fig. 1B).
Flow cytometric assay results further demonstrated that oxymatrine
pretreatment also suppressed the ox-LDL-induced upregulation of the
apoptotic percentage in HUVECs (Fig.
1C). Bcl-2 and Bax are important members of the Bcl-2 family
and they play a key role in the mitochondrial-mediated apoptotic
pathway (31). The results of
western blot analysis (Fig. 1D)
further demonstrated that ox-LDL treatment resulted in an increase
in the expression of the pro-apoptotic protein Bax (Fig. 1E) and a decrease in the
expression of the anti-apoptotic protein Bcl-2 (Fig. 1F), while these effects were
significantly blocked by oxymatrine. These data indicated that
oxymatrine protected HUVECs against ox-LDL-induced injury.
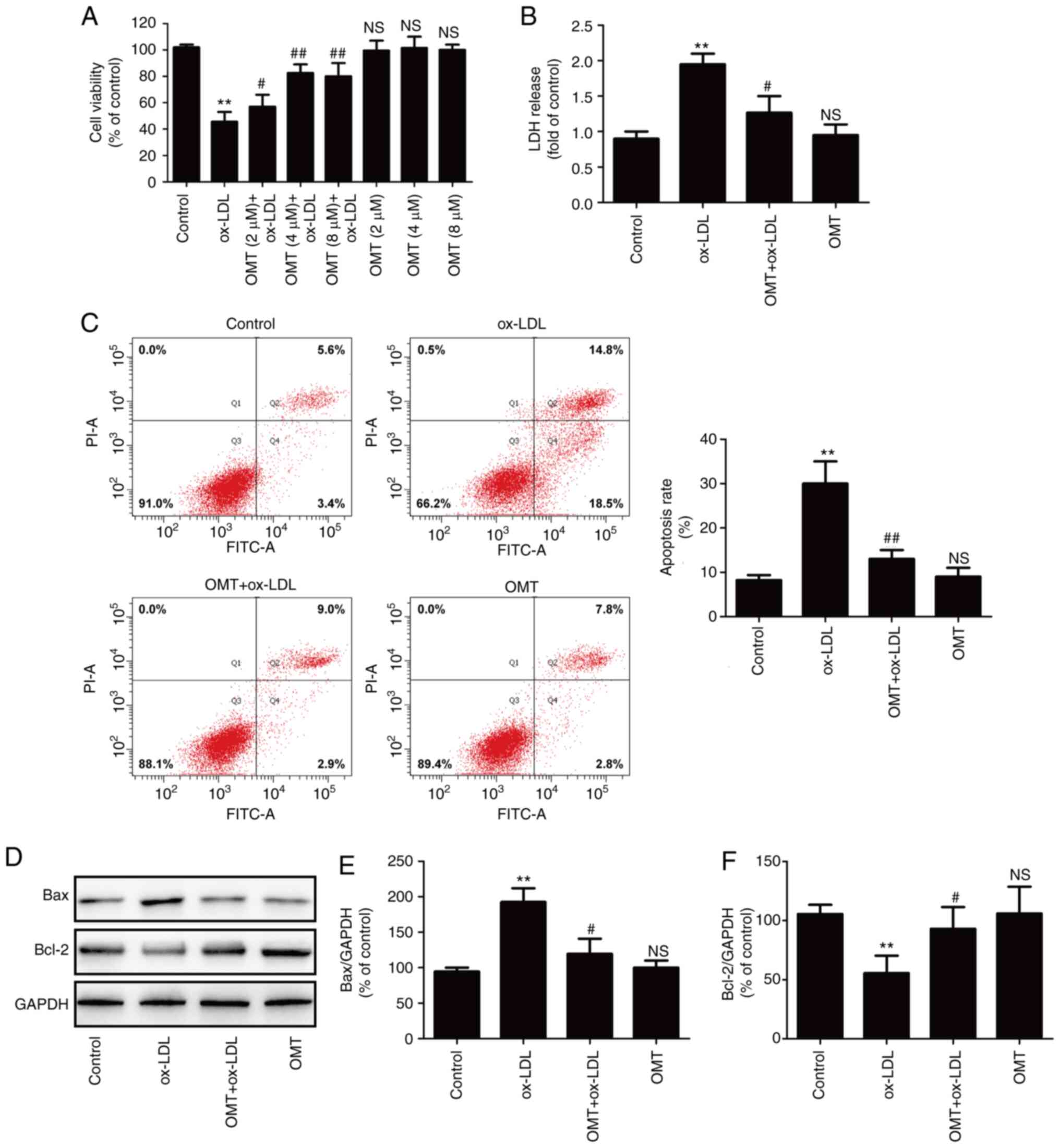 | Figure 1Effects of oxymatrine on cytotoxicity
and apoptosis in ox-LDL-stimulated HUVECs. (A) HUVECs were
pretreated with oxymatrine (2, 4 or 8 µM) for 2 h prior to
treatment with ox-LDL (100 µg/ml) for 24 h, followed by the
evaluation of cell viability. Data are expressed as the mean ± SD,
n=3. **P<0.01 and NSP>0.05 vs. the
control group; #P<0.05 and ##P<0.01 vs.
the ox-LDL group. Cells were pretreated with oxymatrine (4
µM) for 2 h prior to treatment with ox-LDL (100
µg/ml) for 24 h, followed by the determination of (B) LDH
release, (C) apoptosis rate and (D) Bax and Bcl-2 expression.
Quantitative analysis for (E) Bax/GAPDH and (F) Bcl-2/GAPDH. Data
are expressed as the mean ± SD, n=3. **P<0.01 and
NSP>0.05 vs. the control group; #P<0.05
and ##P<0.01 vs. the ox-LDL group. OMT, oxymatrine;
ox-LDL, oxidized low-density lipoprotein; NS, no statistical
significance; HUVECs, human umbilical vein endothelial cells; LDH,
lactate dehydrogenase. |
Oxymatrine inhibits NLRP3
inflammasome-mediated pyroptosis in HUVECs subjected to ox-LDL
treatment
Several studies have revealed the important role of
NLRP3 inflammasome in atherosclerosis (4,32). The present study further
investigated whether oxymatrine has the ability to reduce NLRP3
inflammasome activation during atherosclerosis. As shown in
Fig. 2A, compared with the
ox-LDL group, pretreatment with oxymatrine significantly decreased
the expression of NLRP3 (Fig.
2B) and ASC (Fig. 2C) in
HUVECs. The research confirmed that NLRP3 inflammasome activation
leads to caspase-1 activation, thereby leading to maturation of
effector pro-inflammatory cytokines, such as IL-1β and IL-18; thus,
activated caspase-1 indicates the presence of pyroptosis (7). The results of western blot analysis
(Fig. 2D) further demonstrated
that ox-LDL resulted in the upregulation of the ratio of cleaved
caspase-1/caspase-1 (Fig. 2E),
IL-1β (Fig. 2F) and the
expression of IL-18 (Fig. 2G) in
HUVECs, while these effects were blocked by oxymatrine. These
results indicated that oxymatrine attenuated NLRP3
inflammasome-mediated pyroptosis under conditions of ox-LDL
stimulation.
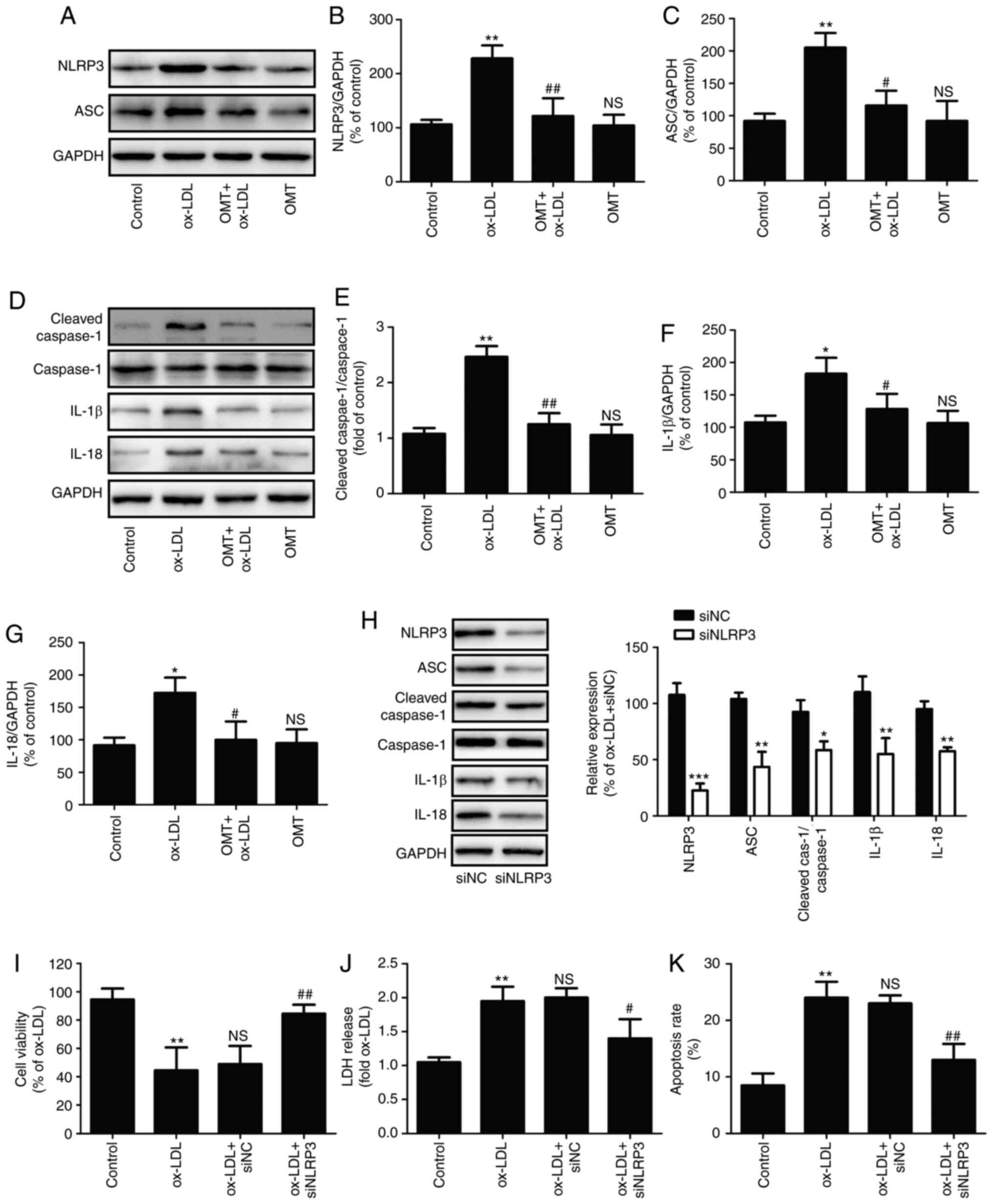 | Figure 2Effects of oxymatrine on NLRP3
inflammasome-mediated pyroptosis in ox-LDL-stimulated HUVECs.
HUVECs were pretreated with oxymatrine (4 µM) for 2 h prior
to treatment with ox-LDL (100 µg/ml) for 24 h. (A) Western
blot analysis for NLRP3 and ASC protein expression. Quantitative
analysis for (B) NLRP3/GAPDH and (C) ASC/GAPDH. (D) Western blot
analysis for cleaved caspase-1, caspase-1, IL-1β and IL-18 protein
expression levels. Quantitative analysis for (E) cleaved
caspase-1/caspase-1, (F) IL-1β/GAPDH and (G) IL-18/GAPDH. Data are
expressed as the mean ± SD, n=3. *P<0.05,
**P<0.01 and NSP>0.05 vs. the control
group; #P<0.05 and ##P<0.01 vs. the
ox-LDL group. HUVECs were transfected with siNLRP3 or siNC for 24
h, followed by measurement of (H) NLRP3, ASC, cleaved caspase-1,
caspase-1, IL-1β and IL-18 protein expression. Data are expressed
as the mean ± SD, n=3. *P<0.05,
**P<0.01, ***P<0.001 vs. siNC group. Cells were
transfected with siNLRP3 or siNC before treatment with oxymatrine
(4 µM) for 2 h prior to treatment with ox-LDL (100
µg/ml) for 24 h. Determination of (I) cell viability, (J)
LDH release and (K) apoptosis rate. Data are expressed as the mean
± SD, n=3. **P<0.01 vs. the control group;
#P<0.05 and ##P<0.01 vs. the ox-LDL +
siNC group. OMT, oxymatrine; ox-LDL, oxidized low-density
lipoprotein; NS, no statistical significance; siNLRP3, small
interfering ribonucleic acid (siRNA) for NLRP3; siNC, siRNA for
negative control; HUVECs, human umbilical vein endothelial cells;
LDH, lactate dehydrogenase; NLRP3, NLR family pyrin domain
containing 3; ASC, apoptosis-associated speck-like protein
containing a C-terminal caspase recruitment domain; Cleaved cas-1,
Cleaved caspase-1. |
Furthermore, to investigate the exact role of NLRP3
inflammasome-mediated pyroptosis in the protection of oxymatrine
against ox-LDL injury, HUVECs were transfected with siNLRP3. The
results demonstrated that siNLRP3 transfection successfully
decreased the expression of NLRP3, ASC, cleaved caspase-1, IL-1β
and IL-18 protein (Fig. 2H) in
HUVECs. Moreover, siNLRP3 transfection also mitigated the
ox-LDL-induced decrease in cell viability (Fig. 2I), the increase in LDH release
(Fig. 2J) and apoptosis rate
(Fig. 2K) of HUVECs, indicating
that inhibition of NLRP3 inflammasome activation-mediated
pyroptosis attenuated ox-LDL-induced HUVEC injury. Collectively,
these findings indicated that oxymatrine protected HUVECs against
ox-LDL-induced injury through inhibition of NLRP3 inflammasome
activation-mediated pyroptosis.
Oxymatrine promotes SIRT1/Nrf2 signaling
pathway activation in HUVECs subjected to ox-LDL treatment
To further investigate the mechanism underlying
oxymatrine-induced endothelial protection against ox-LDL injury,
the effects of oxymatrine on the SIRT1/Nrf2 signaling pathway in
HUVECs were investigated. As revealed in Fig. 3, ox-LDL treatment resulted in the
downregulation of SIRT1 expression in HUVECs, while oxymatrine
pretreatment led to the upregulation of SIRT expression (Fig. 3A and B) in ox-LDL-treated HUVECs.
In addition, oxymatrine blocked the ox-LDL-induced increase in
cytoplasmic Nrf2 (C-Nrf2) expression (Fig. 3A and C) and decrease in nuclear
Nrf2 (N-Nrf2) expression (Fig. 3E
and F), indicating that oxymatrine may promote Nrf2 nuclear
translocation. In addition, the ox-LDL-induced downregulation of
the expression of the Nrf2-related antioxidant defense enzyme HO-1
was also reversed by oxymatrine (Fig. 3A and D). These results indicated
that oxymatrine promoted the activation of the SIRT1/Nrf2 signaling
pathway under conditions of ox-LDL stimulation in HUVECs.
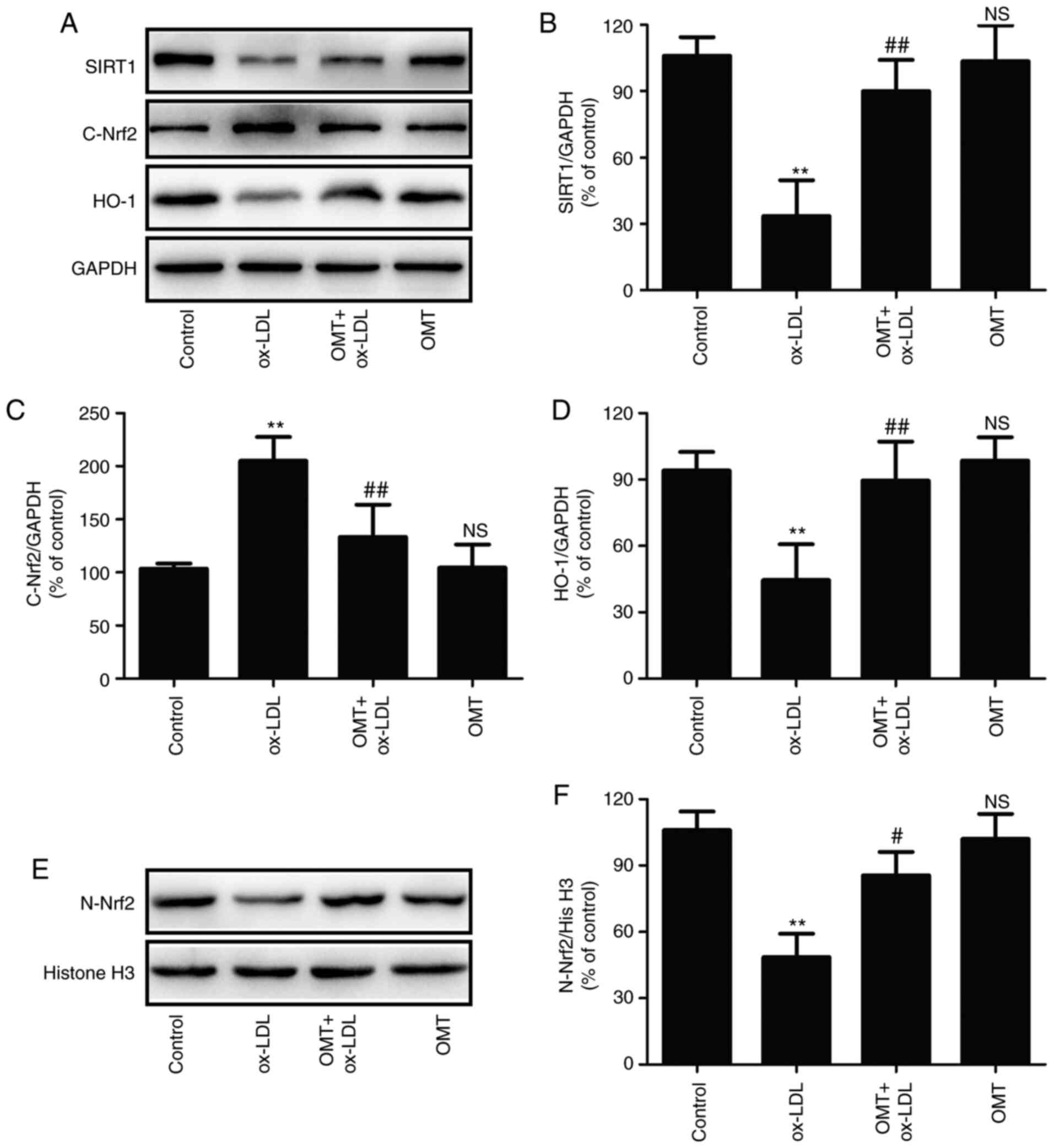 | Figure 3Effects of oxymatrine on the
SIRT1/Nrf2 signaling pathway in ox-LDL-treated HUVECs. HUVECs were
pretreated with oxymatrine (4 µM) for 2 h prior to treatment
with ox-LDL (100 µg/ml) for 24 h. (A) Western blot analysis
for SIRT1, C-Nrf2 and HO-1 protein expression. Quantitative
analysis for (B) SIRT1, (C) C-Nrf2 and (D) HO-1 relative to GAPDH.
(E) Western blot analysis for N-Nrf2 protein expression. (F)
Quantitative analysis for Nrf2 relative to histone H3. Data are
expressed as the mean ± SD, n=3. **P<0.01 and
NSP>0.05 vs. the control group; #P<0.05
and ##P<0.01 vs. the ox-LDL group. OMT, oxymatrine;
ox-LDL, oxidized low-density lipoprotein; NS, no statistical
significance; HUVECs, human umbilical vein endothelial cells; SIRT,
sirtuin; Nrf2, nuclear factor-erythroid 2-related factor 2; C-Nrf2,
cytoplasmic Nrf2; N-Nrf2, nuclear Nrf2; HO-1, heme oxygenase 1. |
Depletion of the SIRT1 signaling pathway
blocks the protective effect of oxymatrine against ox-LDL-induced
injury in HUVECs
To further explore the pathophysiological
significance of oxymatrine-induced SIRT1/Nrf2 signaling pathway
activation, SIRT1 knockdown was performed by transfection of
siSIRT1 to further investigate the effect of oxymatrine against
ox-LDL-induced injury. The results (Fig. 4A) demonstrated that siSIRT1
transfection markedly reduced the expression of SIRT1 compared with
siCon transfection in HUVECs treated with oxymatrine and ox-LDL
(Fig. 4B). siSIRT1 transfection
also decreased the expression of HO-1 (Fig. 4C) and N-Nrf2 (Fig. 4D) in comparison with siCon
transfection, indicating that siSIRT1 transfection successfully led
to the deficiency of the SIRT1/Nrf2 signaling pathway. On this
basis, the results further revealed that siSIRT transfection
inhibited the oxymatrine-induced increase in cell viability
(Fig. 4E) and the decrease in
LDH release (Fig. 4F) in
ox-LDL-treated HUVECs compared with siCon transfection.
Furthermore, the inhibitory effect of oxymatrine on apoptosis rate
was also alleviated by siSIRT1 transfection in comparison with
siCon transfection (Fig. 4G).
Caspase-3 is the main executioner of apoptosis, and activated
caspase-3 can induce the cells to undergo apoptosis (33). Oxymatrine reduced the
ox-LDL-induced increase in the ratio of cleaved
caspase-3/caspase-3, which was also reversed by siSIRT1 (Fig. 4H and I). Collectively, these data
indicated that the SIRT1/Nrf2 signaling pathway may contribute to
the protection of oxymatrine against ox-LDL injury in HUVECs.
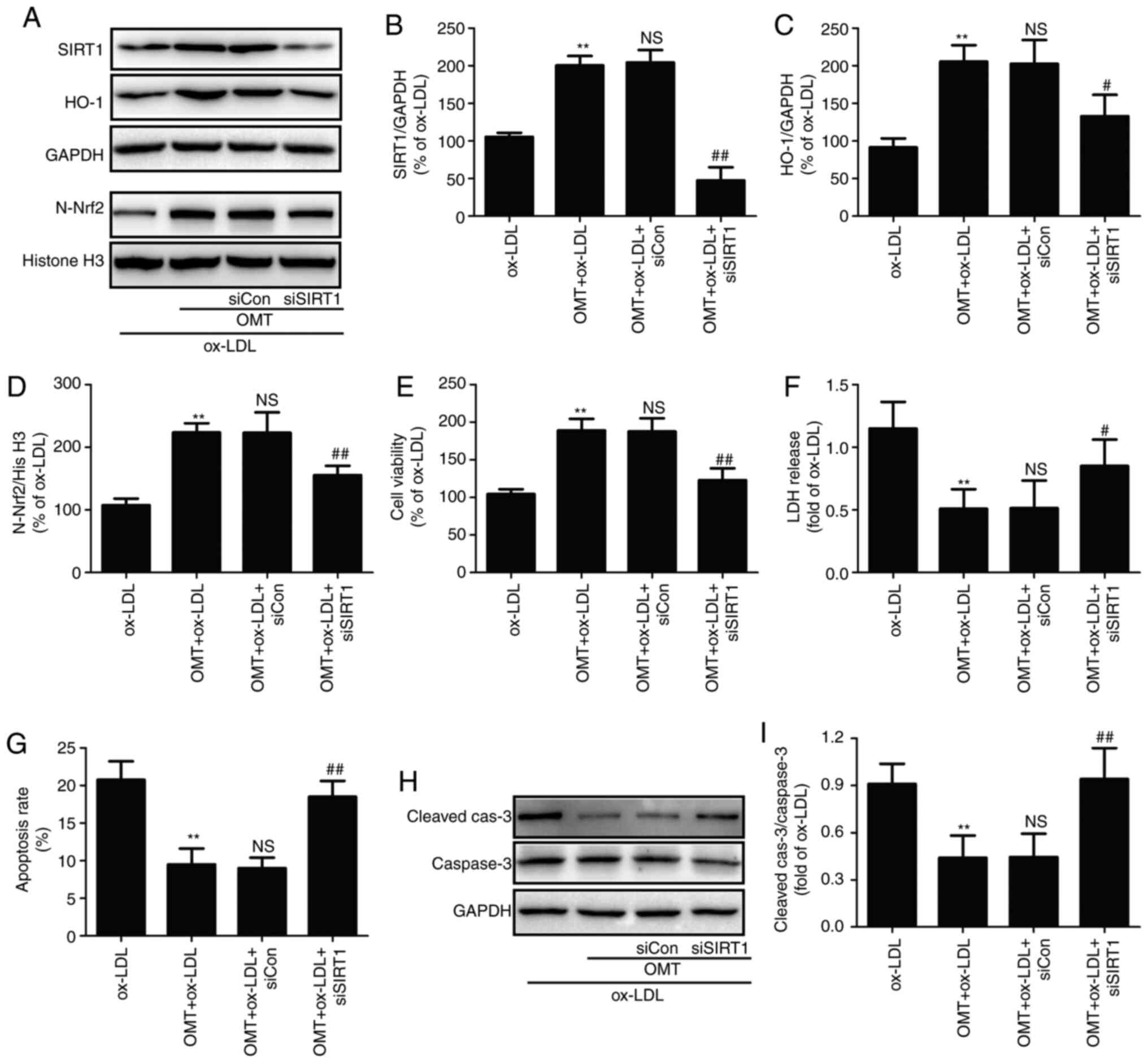 | Figure 4Effects of siSIRT1 transfection on
the protective effects of oxymatrine against ox-LDL-induced injury
in HUVECs. HUVECs transfected with siSIRT1 or siCon were treated
with oxymatrine (4 µM) for 2 h prior to treatment with
ox-LDL (100 µg/ml) for 24 h. (A) Western blot analysis for
protein expression. Quantitative analysis for (B) SIRT1 and (C)
HO-1 relative to GAPDH. (D) Quantitative analysis for N-Nrf2
relative to histone H3. Determination of (E) cell viability, (F)
LDH release and (G) apoptosis rate. (H) Western blot analysis for
cleaved caspase-3 and caspase-3 protein expression. (I)
Quantitative analysis for cleaved caspase-3 relative to caspase-3.
Data are expressed as the mean ± SD, n=3. **P<0.01
vs. the ox-LDL group; NSP>0.05 vs. the OMT + ox-LDL
group; #P<0.05 and ##P<0.01 vs. the OMT
+ ox-LDL + siCon group. OMT, oxymatrine; ox-LDL, oxidized
low-density lipoprotein; NS, no statistical significance; siSIRT1,
siRNAs for SIRT1; siCon, siRNAs for negative control; HUVECs, human
umbilical vein endothelial cells; LDH, lactate dehydrogenase; SIRT,
sirtuin; Nrf2, nuclear factor-erythroid 2-related factor 2; C-Nrf2,
cytoplasmic Nrf2; N-Nrf2, nuclear Nrf2; HO-1, heme oxygenase 1. |
Inhibition of the SIRT1/Nrf2 signaling
pathway eliminates the oxymatrine-induced protective effect against
oxidative stress in ox-LDL-stimulated HUVECs
The present study further investigated the role of
the SIRT1/Nrf2 signaling pathway in the antioxidant activity of
oxymatrine in atherosclerosis. As revealed in Fig. 5, oxymatrine pretreatment
significantly reversed the ox-LDL-induced increase in endogenous
ROS generation (Fig. 5A and B)
and MDA content (Fig. 5C), a
major marker of lipid peroxidation. However, these effects of
oxymatrine were both blocked by siSIRT1 transfection. In addition,
oxymatrine resulted in an increase in MMP in HUVECs subjected to
ox-LDL, which was also reversed by siSIRT1 transfection (Fig. 5D). Excessive ROS production
and/or failure of antioxidant defense systems lead to oxidative
stress and cardiomyocyte injury (34). Hence, the effects of oxymatrine
on antioxidant enzymes inducing SOD, CAT and GSH-Px activity and
the role of the SIRT1/Nrf2 signaling pathway in this process were
explored. The results demonstrated that oxymatrine pretreatment
obviously attenuated the ox-LDL-induced decrease in SOD (Fig. 5E), GSH-Px (Fig. 5F) and CAT (Fig. 5G) activities in HUVECs. However,
oxymatrine-induced increases in SOD (Fig. 5E), GSH-Px (Fig. 5F) and CAT (Fig. 5G) activities were blocked by
siSIRT1 transfection. Collectively, these results indicated that
the SIRT1/Nrf2 signaling pathway mediated the protective effects of
oxymatrine against ox-LDL-induced oxidative damage in HUVECs.
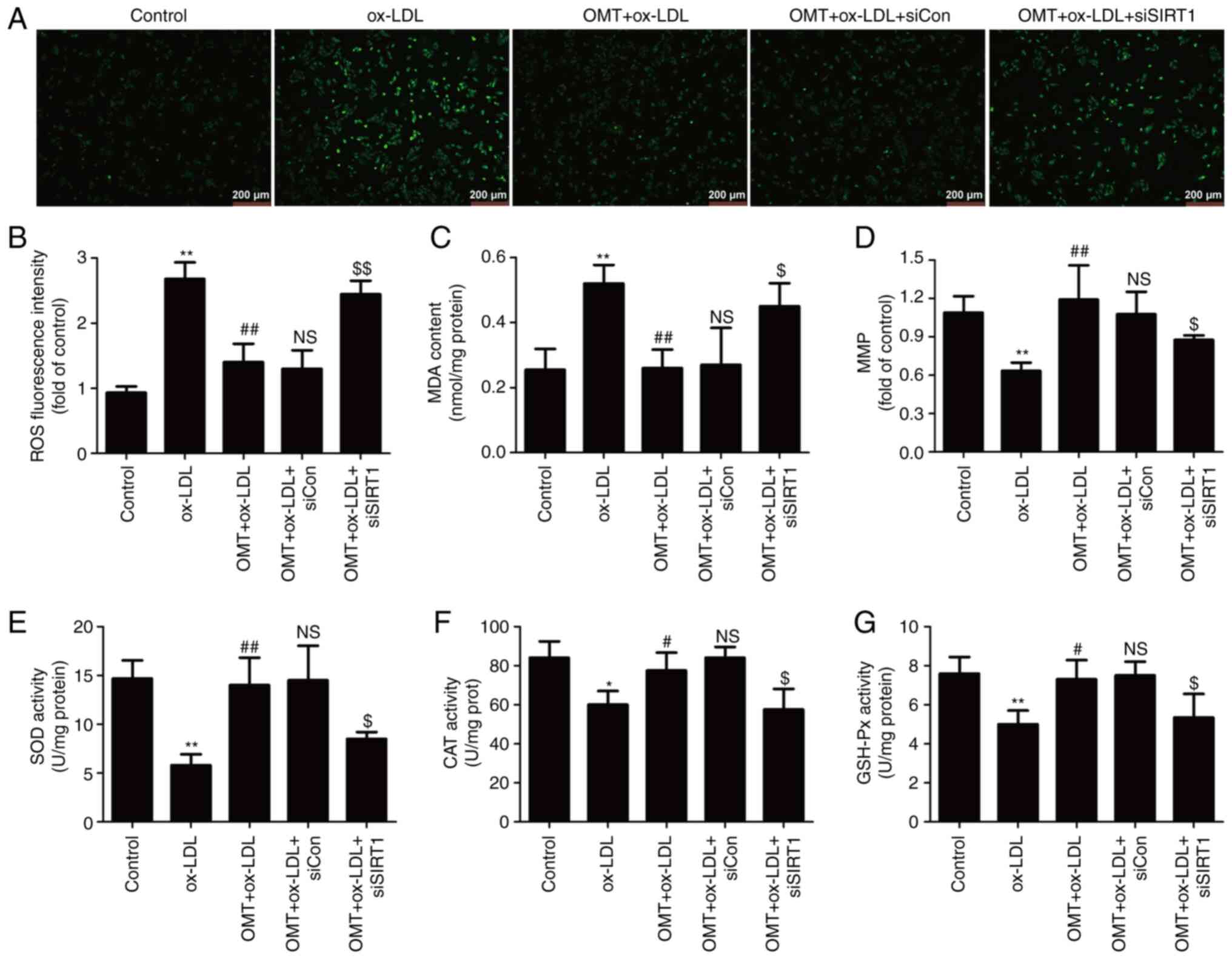 | Figure 5Effects of siSIRT transfection on the
inhibitory effect of oxymatrine on oxidative stress in
ox-LDL-treated HUVECs. HUVECs transfected with siSIRT1 or siCon
were treated with oxymatrine (4 µM) for 2 h prior to
treatment with ox-LDL (100 µg/ml) for 24 h. (A) DCFH-DA
staining for ROS. Scale bar, 200 µm. (B) Quantitative
analysis for ROS generation by flow cytometry. (C) Cell MDA assay
kit for measuring MDA content. (D) JC-1 kit for measuring
mitochondrial membrane potential. (E) SOD assay kit for measuring
SOD activity. (F) CAT assay kit for measuring CAT activity. (G)
GSH-Px assay kit for measuring GSH-Px activity. Data are expressed
as the mean ± SD, n=3. *P<0.05 and
**P<0.01 vs. the control group; #P<0.05
and ##P<0.01 vs. the ox-LDL group;
NSP>0.05 vs. the OMT + ox-LDL group;
$P<0.05 and $$P<0.01 vs. the OMT +
ox-LDL + siCon group. OMT, oxymatrine; ox-LDL, oxidized low-density
lipoprotein; NS, no statistical significance; siSIRT1, siRNAs for
SIRT1; siCon, siRNAs for negative control; HUVECs, human umbilical
vein endothelial cells; SIRT, sirtuin; MDA, malondialdehyde; SOD,
superoxide dismutase; CAT, catalase; GSH-Px, glutathione
peroxidase; ROS, reactive oxygen species; DCFH-DA,
2′,7′-dichlorofluorescein diacetate. |
Inhibition of the SIRT1/Nrf2 signaling
pathway alleviates oxymatrine-reduced NLRP3 inflammasome-mediated
pyroptosis in ox-LDL-stimulated HUVECs
Research has confirmed that SIRT1 signaling inhibits
NLRP3 inflammasome activation in vascular endothelial cells
(35). The present study was
undertaken to further explore whether oxymatrine attenuates NLRP3
inflammasome activation-mediated pyroptosis in ox-LDL-stimulated
HUVECs via promoting SIRT1/Nrf2 signaling. The findings (Fig. 6A) revealed that inactivation of
the SIRT1/Nrf2 signaling pathway induced by siSIRT1 transfection
weakened the oxymatrine-induced decrease in the expression of NLRP3
(Fig. 6B) and ASC (Fig. 6C) in ox-LDL-treated HUVECs.
Transfection with siSIRT1 also inhibited the oxymatrine-induced
decrease in the ratio of cleaved caspase-1/caspase-1 (Fig. 6D and E) and the expression of
IL-1β (Fig. 6D and F) and IL-18
(Fig. 6D and G) in
ox-LDL-treated HUVECs. These results indicated that the SIRT1/Nrf2
signaling pathway mediated the inhibitory effect of oxymatrine on
NLRP3 inflammasome-mediated pyroptosis in ox-LDL-treated
HUVECs.
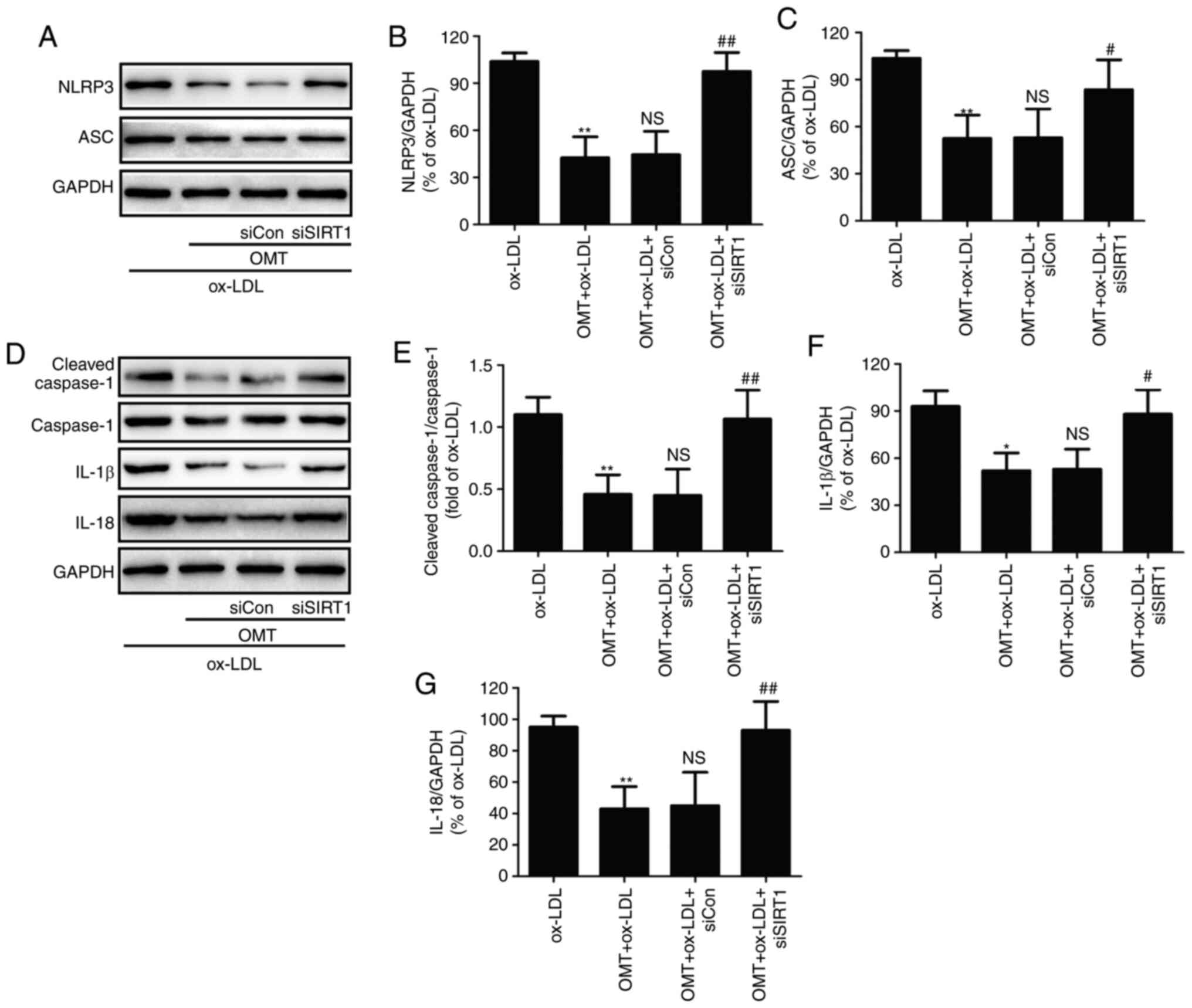 | Figure 6Effects of siSIRT transfection on the
inhibitory effect of oxymatrine against NLRP3 inflammasome-mediated
pyroptosis in ox-LDL-treated HUVECs. HUVECs transfected with
siSIRT1 or siCon were treated with oxymatrine (4 µM) for 2 h
prior to treatment with ox-LDL (100 µg/ml) for 24 h. (A)
Western blot analysis for protein expression. Quantitative analysis
for (B) NLRP3 and (C) ASC relative to GAPDH. (D) Western blot
analysis for protein expression. Quantitative analysis for (E)
cleaved caspase-1/caspase-1, (F) IL-1β and (G) IL-18 relative to
GAPDH. Data are expressed as the mean ± SD, n=3.
**P<0.01 vs. the ox-LDL group; NSP>0.05
vs. the OMT + ox-LDL group; #P<0.05 and
##P<0.01 vs. the OMT + ox-LDL + siCon group. OMT,
oxymatrine; ox-LDL, oxidized low-density lipoprotein; NS, no
statistical significance; siSIRT1, siRNAs for SIRT1; siCon, siRNAs
for negative control; HUVECs, human umbilical vein endothelial
cells; NLRP3, NLR family pyrin domain containing 3; SIRT, sirtuin;
ASC, apoptosis-associated speck-like protein containing a
C-terminal caspase recruitment domain. |
Discussion
The present study investigated the protective
effects of oxymatrine against ox-LDL-induced injury in HUVECs, and
further demonstrated the underlying mechanisms, focusing on NLRP3
inflammasome-mediated pyroptosis and SIRT1/Nrf2 signaling. The
results revealed that oxymatrine attenuated ox-LDL-induced
cytotoxicity and apoptosis via inhibiting NLRP3
inflammasome-mediated pyroptosis. In addition, the SIRT1/Nrf2
signaling pathway mediated the protective effects of oxymatrine
against ox-LDL injury and oxidative stress, and contributed to the
inhibitory effect of oxymatrine on ox-LDL-induced NLRP3
inflammasome-mediated pyroptosis in HUVECs. These results indicated
that oxymatrine may mitigate ox-LDL injury by inhibiting NLRP3
inflammasome-mediated pyroptosis via activating the Sirt1/Nrf2
signaling pathway, providing new insight into a functional role of
oxymatrine in atherosclerosis.
Atherosclerosis is a multifactorial, chronic
inflammatory disease that causes injury of endothelial cells, which
are the first to be exposed to the effects of stimuli and
physiological factors from their surroundings (2,36). Oxymatrine, a natural bioactive
compound, has been demonstrated to possess anti-inflammatory,
antioxidant, antiviral, antifibrotic and cardioprotective
properties (19,37). Recently, Zhang et al
(19) revealed that oxymatrine
may inhibit homocysteine-induced endothelial dysfunction, a
hallmark of atherosclerosis, by attenuating autophagy-activated
HUVEC apoptosis/death. Another study also demonstrated that
oxymatrine prevented homocysteine-induced endothelial injury via
attenuating mitochondrial-dependent apoptosis (18). Consistent with these studies, the
present study also demonstrated that oxymatrine alleviated
ox-LDL-induced cytotoxicity and apoptosis in HUVECs. Bcl-2 family
proteins are involved in the mitochondria-dependent apoptosis
pathway (38). The results of
the present study further demonstrated that oxymatrine reversed the
ox-LDL-induced increase in Bax expression and the decrease in Bcl-2
expression in HUVECs. Collectively, these results indicated that
oxymatrine protected HUVECs against ox-LDL injury via inhibiting
the mitochondria-dependent apoptosis pathway.
Inflammation is an important driver of
atherosclerosis, and therapeutic targeting of inflammatory pathways
is suggested to ameliorate atherosclerosis (39,40). NLRP3 inflammasomes are
intracellular complexes involved in the inflammatory response that
increases the levels of mature IL-1β and IL-18 and initiates
pyroptosis via cleaving procaspase-1, playing a decisive role in
atherosclerosis (3,41). A number of studies have revealed
that oxymatrine plays a markedly beneficial role in multiple
diseases due to its profound anti-inflammatory effects (37,42,43). However, to the best of our
knowledge, the impact of oxymatrine on NLRP3 inflammasomes and
pyroptosis has not been explored in atherosclerosis. The present
study demonstrated that oxymatrine significantly inhibited the
ox-LDL-induced increase in the expression of NLRP3 and ASC, as well
as the expression of cleaved caspase-1, IL-1β and IL-18 in HUVECs,
indicating that oxymatrine attenuated NLRP3 inflammasome
activation-mediated pyroptosis under conditions of ox-LDL
stimulation. In addition, the results further revealed that
inhibition of NLRP3 inflammasome activation-mediated pyroptosis
induced by siNLRP3 transfection inhibited the protective effects of
oxymatrine against ox-LDL-induced HUVEC injury. Collectively, these
results indicated that oxymatrine protected HUVECs against
ox-LD-induced injury via inhibiting NLRP3 inflammasome-mediated
pyroptosis.
SIRT1 was previously revealed to be involved in
multiple cellular processes, and exerts marked protective effects
against endothelial cell injury, vascular aging, inflammation and
atherosclerotic plaque development (22,23). It was recently reported that
oxymatrine plays a beneficial role in ischemic stroke (29) and non-alcoholic fatty liver
disease (29) via promoting SRT1
signaling. However, the effect of oxymatrine on the SIRT1 signaling
pathway in atherosclerosis remains unknown. In the present study,
the results revealed that oxymatrine pretreatment markedly
increased the expression of SIRT1 in ox-LDL-treated HUVECs. Nrf2, a
key antioxidant sensor for cellular defense mechanisms, is
considered as an important downstream target of SIRT1 and increases
resistance to oxidative damage; it is also closely associated with
atherosclerosis development (25). Under normal conditions, Nrf2
binds to its negative regulator, Kelch-like ECH-associated protein
1 (Keap1), which leads to its degradation via ubiquitination. In
response to oxidative stress, Nrf2 is dissociated from Keap1 and
translocates to the nucleus, thereby activating antioxidant
responses by upregulating the expression of several antioxidant
enzymes, particularly HO-1 (44,45). Consistent with these previous
findings, the present study demonstrated that ox-LDL increased the
expression of C-Nrf2, decreased the expression of N-Nrf2, and
enhanced the expression of HO-1, which were also attenuated by
oxymatrine. These results indicated that oxymatrine activated the
SIRT1/Nrf2 signaling pathway in ox-LDL-treated HUVECs. Furthermore,
the present study demonstrated that inhibition of SIRT1/Nrf2
signaling induced by siSIRT1 transfection mitigated the protective
effects of oxymatrine against ox-LDL injury in HUVECs.
Collectively, these results indicated that the SIRT1/Nrf2 signaling
pathway mediated the cardioprotective function of oxymatrine in
atherosclerosis.
An increasing number of studies have revealed that
the mechanisms underlying the protective effects of oxymatrine are
mainly associated with its anti-inflammatory and antioxidant
properties (13,37,42). Similarly, the present results
demonstrated that oxymatrine markedly attenuated ox-LDL-induced
oxidative stress, as evidenced by the decrease in ROS generation
and MDA content, and the increase in MMP and the activities of the
antioxidant enzymes SOD, CAT and GSH-Px in HUVECs. Previous studies
have reported that SIRT1/Nrf2 signaling dysregulation is involved
in the ability to scavenge oxygen free radicals through the action
of SOD, CAT and GSH-Px (46,47). Consistent with these studies, the
present results further revealed that the inhibition of SIRT1/Nrf2
signaling pathway through siSIRT1 transfection eliminated the
antioxidant properties of oxymatrine under conditions of ox-LDL
stimulation. These results indicated that oxymatrine attenuated
ox-LDL-induced injury by attenuating oxidative stress via promoting
SIRT1/Nrf2 signaling in HUVECs.
An increasing number of studies have reported that
activating SIRT1/Nrf2 signaling may reduce NLRP3 inflammasome
activation (48) and pyroptosis
(49). However, the role of the
SIRT1/Nrf2 signaling pathway in the inhibitory effect of oxymatrine
on NLRP3 inflammasome-mediated pyroptosis has not been reported to
date. In the present study, the results demonstrated that siSIRT1
also abolished the inhibitory effect of oxymatrine on NLRP3
inflammasome-mediated pyroptosis, which was similar to the findings
of previous studies reporting that NLRP3 inflammasome activation
was associated with Nrf2 and SIRT1 activation and can be reversed
by Nrf2 siRNA and SIRT1 inhibitor treatment (48,50). These results indicated that
oxymatrine reduced NLRP3 inflammasome-mediated pyroptosis via
enhancing SIRT1/Nrf2 signaling in ox-LDL-treated HUVECs.
Some shortcomings must be acknowledged in this
study. Firstly, no specific immunofluorescence markers for
pyroptosis in HUVECs were tested. Secondly, our experiments are
limited to HUVECs, other cells or an in vivo experimental
model have not yet been considered.
Collectively, the findings of the present study
demonstrated that oxymatrine protected HUVECs against
ox-LDL-induced injury by inhibiting NLRP3 inflammasome-mediated
pyroptosis via activation of the SIRT1/Nrf2 signaling pathway. The
results indicated that oxymatrine may serve as a potential
therapeutic candidate for the treatment of atherosclerosis, and
suggested a new strategy for targeting SIRT1/Nrf2 signaling and
NLRP3 inflammasome-mediated pyroptosis.
Availability of data and materials
The datasets generated and/or analyzed in the
present study are available from the corresponding author upon
reasonable request.
Authors’ contributions
XJ and BW conceived, designed the experiments, and
confirmed the authenticity of the raw data. XJ, WF and JZ performed
the experiments and analyzed the data. NS and YY collected and
analyzed the data. XJ, YY and BW wrote and revised the manuscript.
All the authors have read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
Funding
The present study was supported by the Hunan Provincial Natural
Science Foundation of China (grant nos. 2018JJ3465 and
2019JJ50551).
References
|
1
|
Kobiyama K and Ley K: Atherosclerosis.
Circ Res. 123:1118–1120. 2018. View Article : Google Scholar
|
|
2
|
Libby P, Bornfeldt KE and Tall AR:
Atherosclerosis: Successes, surprises, and future challenges. Circ
Res. 118:531–534. 2016. View Article : Google Scholar
|
|
3
|
Baldrighi M, Mallat Z and Li X: NLRP3
inflammasome pathways in atherosclerosis. Atherosclerosis.
267:127–138. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Paramel Varghese G, Folkersen L,
Strawbridge RJ, Halvorsen B, Yndestad A, Ranheim T, Krohg-Sørensen
K, Skjelland M, Espevik T, Aukrust P, et al: NLRP3 inflammasome
expression and activation in human atherosclerosis. J Am Heart
Assoc. 5:e0030312016. View Article : Google Scholar :
|
|
5
|
Jiang C and Jiang L, Li Q, Liu X, Zhang T,
Dong L, Liu T, Liu L, Hu G, Sun X and Jiang L: Acrolein induces
NLRP3 inflammasome-mediated pyroptosis and suppresses migration via
ROS-dependent autophagy in vascular endothelial cells. Toxicology.
410:26–40. 2018. View Article : Google Scholar
|
|
6
|
Qiu Z, Lei S, Zhao B, Wu Y, Su W, Liu M,
Meng Q, Zhou B, Leng Y and Xia ZY: NLRP3 inflammasome
activation-mediated pyroptosis aggravates myocardial
ischemia/reperfusion injury in diabetic rats. Oxid Med Cell Longev.
2017:97432802017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sutterwala FS, Haasken S and Cassel SL:
Mechanism of NLRP3 inflammasome activation. Ann NY Acad Sci.
1319:82–95. 2014. View Article : Google Scholar
|
|
8
|
Jia C, Zhang J, Chen H, Zhuge Y, Chen H,
Qian F, Zhou K, Niu C, Wang F, Qiu H, et al: Endothelial cell
pyroptosis plays an important role in Kawasaki disease via
HMGB1/RAGE/cathespin B signaling pathway and NLRP3 inflammasome
activation. Cell Death Dis. 10:7782019. View Article : Google Scholar
|
|
9
|
Li P, Zhong X, Li J, Liu H, Ma X, He R and
Zhao Y: MicroRNA-30c-5p inhibits NLRP3 inflammasome-mediated
endothelial cell pyroptosis through FOXO3 down-regulation in
atherosclerosis. Biochem Biophys Res Commun. 503:2833–2840. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zahid A, Li B, Kombe AJK, Jin T and Tao J:
Pharmacological Inhibitors of the NLRP3 Inflammasome. Front
Immunol. 10:25382019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang L, Yuan M, Zhang L, Wu B and Sun X:
Adiponectin alleviates NLRP3-inflammasome-mediated pyroptosis of
aortic endothelial cells by inhibiting FoxO4 in arteriosclerosis.
Biochem Biophys Res Commun. 514:266–272. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu Y, Wang H, Liu N, Du J, Lan X, Qi X,
Zhuang C, Sun T, Li Y and Yu J: Oxymatrine protects neonatal rat
against hypoxic-ischemic brain damage via PI3K/Akt/GSK3β pathway.
Life Sci. 254:1164442020. View Article : Google Scholar
|
|
13
|
Liang J, Chang B, Huang M, Huang W, Ma W,
Liu Y, Tai W, Long Y and Lu Y: Oxymatrine prevents synovial
inflammation and migration via blocking NF-κB activation in
rheumatoid fibroblast-like synoviocytes. Int Immunopharmacol.
55:105–111. 2018. View Article : Google Scholar
|
|
14
|
Zhang YY, Yi M and Huang YP: Oxymatrine
ameliorates doxorubicin-induced cardiotoxicity in rats. Cell
Physiol Biochem. 43:626–635. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hu ST, Tang Y, Shen YF, Ao HH, Bai J, Wang
YL and Yang YJ: Protective effect of oxymatrine on chronic rat
heart failure. J Physiol Sci. 61:363–372. 2011. View Article : Google Scholar
|
|
16
|
Yang Y, Chen S, Tao L, Gan S, Luo H, Xu Y
and Shen X: Inhibitory effects of oxymatrine on
transdifferentiation of neonatal rat cardiac fibroblasts to
myofibroblasts induced by aldosterone via Keap1/Nrf2 signaling
pathways in vitro. Med Sci Monit. 25:5375–5388. 2019. View Article : Google Scholar
|
|
17
|
Huang XY and Chen CX: Effect of
oxymatrine, the active component from Radix Sophorae flavescentis
(Kushen), on ventricular remodeling in spontaneously hypertensive
rats. Phytomedicine. 20:202–212. 2013. View Article : Google Scholar
|
|
18
|
Wu B, Yue H, Zhou GH, Zhu YY, Wu TH, Wen
JF, Cho KW and Jin SN: Protective effects of oxymatrine on
homocysteine-induced endothelial injury: Involvement of
mitochondria-dependent apoptosis and Akt-eNOS-NO signaling
pathways. Eur J Pharmacol. 864:1727172019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang Y, Zhang Y, Tang J, Zhao S, Li C,
Huang YP and Yi M: Oxymatrine inhibits homocysteine-mediated
autophagy via MIF/mTOR signaling in human umbilical vein
endothelial cells. Cell Physiol Biochem. 45:1893–1903. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Khatana C, Saini NK, Chakrabarti S, Saini
V, Sharma A, Saini RV and Saini AK: Mechanistic insights into the
oxidized low-density lipoprotein-induced atherosclerosis. Oxid Med
Cell Longev. 2020:52453082020. View Article : Google Scholar :
|
|
21
|
Kitada M, Ogura Y and Koya D: The
protective role of Sirt1 in vascular tissue: Its relationship to
vascular aging and atherosclerosis. Aging (Albany NY). 8:2290–2307.
2016. View Article : Google Scholar
|
|
22
|
Zhang MJ, Zhou Y, Chen L, Wang X, Long CY,
Pi Y, Gao CY, Li JC and Zhang LL: SIRT1 improves VSMC functions in
atherosclerosis. Prog Biophys Mol Biol. 121:11–15. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Stein S and Matter CM: Protective roles of
SIRT1 in atherosclerosis. Cell Cycle. 10:640–647. 2011. View Article : Google Scholar
|
|
24
|
Lazaro I, Lopez-Sanz L, Bernal S, Oguiza
A, Recio C, Melgar A, Jimenez-Castilla L, Egido J, Madrigal-Matute
J and Gomez-Guerrero C: Nrf2 activation provides atheroprotection
in diabetic mice through concerted upregulation of antioxidant,
anti-inflammatory, and autophagy mechanisms. Front Pharmacol.
9:8192018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mimura J and Itoh K: Role of Nrf2 in the
pathogenesis of atherosclerosis. Free Radic Biol Med. 88:221–232.
2015. View Article : Google Scholar
|
|
26
|
Gao Z, Sui J, Fan R, Qu W, Dong X and Sun
D: Emodin protects against acute pancreatitis-associated lung
injury by inhibiting NLPR3 inflammasome activation via Nrf2/HO-1
signaling. Drug Des Devel Ther. 14:1971–1982. 2020. View Article : Google Scholar :
|
|
27
|
Hou Y, Wang Y, He Q, Li L, Xie H, Zhao Y
and Zhao J: Nrf2 inhibits NLRP3 inflammasome activation through
regulating Trx1/TXNIP complex in cerebral ischemia reperfusion
injury. Behav Brain Res. 336:32–39. 2018. View Article : Google Scholar
|
|
28
|
Xu H, Chen GF, Ma YS, Zhang HW, Zhou Y,
Liu GH, Chen DY, Ping J, Liu YH, Mou X and Fu D: Hepatic proteomic
changes and Sirt1/AMPK signaling activation by oxymatrine treatment
in rats with non-alcoholic steatosis. Front Pharmacol. 11:2162020.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhou S, Qiao B, Chu X and Kong Q:
Oxymatrine attenuates cognitive deficits through SIRT1-mediated
autophagy in ischemic stroke. J Neuroimmunol. 323:136–142. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhu Z, Li J and Zhang X: Salidroside
protects against ox-LDL-induced endothelial injury by enhancing
autophagy mediated by SIRT1-FoxO1 pathway. BMC Complement Altern
Med. 19:1112019. View Article : Google Scholar :
|
|
31
|
Edlich F: BCL-2 proteins and apoptosis:
Recent insights and unknowns. Biochem Biophys Res Commun.
500:26–34. 2018. View Article : Google Scholar
|
|
32
|
Grebe A, Hoss F and Latz E: NLRP3
inflammasome and the IL-1 pathway in atherosclerosis. Circ Res.
122:1722–1740. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Esteban-Fernández de Ávila B,
Ramírez-Herrera DE, Campuzano S, Angsantikul P, Zhang L and Wang J:
Nanomotor-enabled pH-responsive intracellular delivery of
caspase-3: Toward rapid cell apoptosis. CS Nano. 11:5367–5374.
2017.
|
|
34
|
Dubois-Deruy E, Peugnet V, Turkieh A and
Pinet F: Oxidative stress in cardiovascular diseases. Antioxidants
(Basel). 9:8642020. View Article : Google Scholar
|
|
35
|
Li Y, Wang P, Yang X, Wang W, Zhang J, He
Y, Zhang W, Jing T, Wang B and Lin R: SIRT1 inhibits inflammatory
response partly through regulation of NLRP3 inflammasome in
vascular endothelial cells. Mol Immunol. 77:148–156. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Falk E: Pathogenesis of atherosclerosis. J
Am Coll Cardiol. 47(Suppl 8): C7–C12. 2006. View Article : Google Scholar
|
|
37
|
Lan X, Zhao J, Zhang Y, Chen Y, Liu Y and
Xu F: Oxymatrine exerts organ- and tissue-protective effects by
regulating inflammation, oxidative stress, apoptosis, and fibrosis:
From bench to bedside. Pharmacol Res. 151:1045412020. View Article : Google Scholar
|
|
38
|
King KL and Cidlowski JA: Cell cycle
regulation and apoptosis. Annu Rev Physiol. 60:601–617. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Bäck M, Yurdagul A Jr, Tabas I, Öörni K
and Kovanen PT: Inflammation and its resolution in atherosclerosis:
Mediators and therapeutic opportunities. Nat Rev Cardiol.
16:389–406. 2019.PubMed/NCBI
|
|
40
|
Nasonov EL and Popkova TV:
Atherosclerosis: Perspectives of anti-inflammatory therapy. Ter
Arkh. 90:4–12. 2018.
|
|
41
|
Hoseini Z, Sepahvand F, Rashidi B,
Sahebkar A, Masoudifar A and Mirzaei H: NLRP3 inflammasome: Its
regulation and involvement in atherosclerosis. J Cell Physiol.
233:2116–2132. 2018. View Article : Google Scholar
|
|
42
|
Dong P, Ji X, Han W and Han H: Oxymatrine
exhibits anti-neuroinflammatory effects on Aβ142-induced
primary microglia cells by inhibiting NF-κB and MAPK signaling
pathways. Int Immunopharmacol. 74:1056862019. View Article : Google Scholar
|
|
43
|
Jiang Y, Sang W, Wang C, Lu H, Zhang T,
Wang Z, Liu Y, Xue B, Xue S, Cai Z, et al: Oxymatrine exerts
protective effects on osteoarthritis via modulating chondrocyte
homoeostasis and suppressing osteoclastogenesis. J Cell Mol Med.
22:3941–3954. 2018. View Article : Google Scholar :
|
|
44
|
Bellezza I, Giambanco I, Minelli A and
Donato R: Nrf2-Keap1 signaling in oxidative and reductive stress.
Biochim Biophys Acta Mol Cell Res. 1865:721–733. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Chen QM and Maltagliati AJ: Nrf2 at the
heart of oxidative stress and cardiac protection. Physiol Genomics.
50:77–97. 2018. View Article : Google Scholar :
|
|
46
|
Chen XJ, Wu WJ, Zhou Q, Jie JP, Chen X,
Wang F and Gong XH: Advanced glycation end-products induce
oxidative stress through the Sirt1/Nrf2 axis by interacting with
the receptor of AGEs under diabetic conditions. J Cell Biochem. Oct
15–2018.Epub ahead of print. View Article : Google Scholar
|
|
47
|
Zhang B, Zhai M, Li B, Liu Z, Li K, Jiang
L, Zhang M, Yi W, Yang J, Yi D, et al: Honokiol ameliorates
myocardial ischemia/reperfusion injury in type 1 diabetic rats by
reducing oxidative stress and apoptosis through activating the
SIRT1-Nrf2 signaling pathway. Oxid Med Cell Longev.
2018:31598012018.
|
|
48
|
Arioz BI, Tastan B, Tarakcioglu E, Tufekci
KU, Olcum M, Ersoy N, Bagriyanik A, Genc K and Genc S: Melatonin
attenuates LPS-induced acute depressive-like behaviors and
microglial NLRP3 inflammasome activation through the SIRT1/Nrf2
pathway. Front Immunol. 10:15112019. View Article : Google Scholar :
|
|
49
|
Yin Y, Wu X, Peng B, Zou H, Li S, Wang J
and Cao J: Curcumin improves necrotising microscopic colitis and
cell pyroptosis by activating SIRT1/NRF2 and inhibiting the TLR4
signalling pathway in newborn rats. Innate Immun. 26:609–617. 2020.
View Article : Google Scholar
|
|
50
|
Zhang S, Jiang L, Che F, Lu Y, Xie Z and
Wang H: Arctigenin attenuates ischemic stroke via SIRT1-dependent
inhibition of NLRP3 inflammasome. Biochem Biophys Res Commun.
493:821–826. 2017. View Article : Google Scholar : PubMed/NCBI
|














