Introduction
Prostate cancer is the second leading cause of
cancer-related mortality in American males (1). The androgen receptor (AR) is a ligand
activated steroid hormone receptor and a key regulator of normal
prostate development and function (2). The AR has a critical role in prostate
cancer development and progression (3). Consequently, the current therapeutic
strategies for prostate cancer intervention, including androgen
ablation therapy, inhibits AR function (4). An aggressive form of prostate cancer
therapy is based on a combination of androgen synthesis suppression
and AR inhibition (5). Therefore,
identification of chemical agents that inhibit AR signaling by
known or novel mechanisms warrant further investigation for the
development of a novel prostate cancer therapeutic approach.
Curcumin is a non-nutritive yellow pigment found in
the spice turmeric, which is derived from the rhizome of the plant
Curcuma longa Linn. Numerous studies have demonstrated the
anticancer activity of curcumin and curcumin analogues in animal
models (6–11), as well as growth inhibition and
apoptosis-inductive effects in a variety of cancer cell lines in
vitro (12–20). However, the clinical efficacy of
curcumin is limited, which is likely due to its low bioavailability
(21–23).
Our previous study reported on the synthesis and
evaluation of 61 curcumin-related compounds for the inhibitory
effects on cultured prostate cancer PC-3 cells, pancreas cancer
Panc-1 cells and colon cancer HT-29 cells (24). Five of these curcumin analogues
with different linker groups but identical symmetrical aromatic
rings (as revealed in Fig. 1) were
selected for further study. These compounds included
(2E,6E)-2,6-bis(3,4,5-trimethoxybenzylidene) cyclohexanone (A10),
(2E,5E)-2,5-bis(3,4,5-trimethoxybenzylidene) cyclopentanone (B10),
(1E,4E)-1,5-bis(3,4,5-trimethoxyphenyl) penta-1,4-dien-3-one (C10),
(3E,5E)-3,5-bis (3,4,5-trimethoxybenzyli-dene)
tetrahydropyran-4-one (E10), (3E,5E)-3,5-bis
(3,4,5-trimethoxybenzylidene) tetrahydrothiopyran-4-one (F10).
Compounds with a heteroatom linker (compounds E10 and F10)
demonstrated a stronger inhibitory effect than those without a
heteroatom linker (compounds A10, B10 and C10) on the growth of
human prostate cancer cells, although C10 had intermediate
activity. It was also demonstrated that E10 and F10 more potently
inhibited AR activity and testosterone (TT)- or dihydrotestosterone
(DHT)-induced prostate specific antigen expression than A10, B10
and curcumin in CWR-22Rv1 cells.
Materials and methods
Chemistry
Several curcumin analogues (A10, B10, C10, E10, F10
as demonstrated in Fig. 1) with
different linker groups were synthesized by coupling the
appropriate substituted benzaldehyde with cyclohexanone,
cyclopentanone, acetone, tetrahydropyran-4-ones or
tetrahydrothiopyran-4-one as previously described (24). Characterization of the compounds,
(2E,6E)-2,6-bis(3,4,5-trimethoxy-benzylidene) cyclohexanone (A10),
(2E,5E)-2,5-bis(3,4,5-trimethoxybenzylidene) cyclopentanone (B10),
(1E,4E)-1,5-bis(3,4,5-trimethoxyphenyl) penta-1,4-dien-3-one (C10),
(3E,5E)-3,5-bis(3,4,5-trime thoxybenzylidene) tetrahydropyran-4-one
(E10) and (3E,5E)-3,5-bis(3,4,5-trime-thoxybenzylidene) tetrahydro
thiopyran-4-one (F10), was previously described in detail (24).
Cell culture and reagents
CWR-22Rv1 and LNCaP cells were obtained from the
American Type Culture Collection (ATCC; Rockville, MD, USA).
RPMI-1640 tissue culture medium, penicillin-streptomycin,
L-glutamine and fetal bovine serum (FBS) were purchased from Gibco
(Grand Island, NY, USA). CWR-22Rv1 cells were maintained in
RPMI-1640 culture medium. RPMI-1640 medium was supplemented with
10% FBS, penicillin (100 units/ml)-streptomycin (100 μg/ml) and
L-glutamine (300 μg/ml). The cultured cells were grown at 37°C in a
humidified atmosphere of 5% CO2 and were passaged twice
a week. Curcumin analogues were dissolved in dimethyl sulfoxide and
the final concentration of DMSO was 0.1% in all experiments.
MTT, trypan blue and apoptosis
assays
For the MTT assay, CWR-22Rv1 and LNCaP cells were
seeded at a density of 2×104 cells/ml of medium in a
96-well plate (0.2 ml/well) and incubated for 24 h. The cells were
then treated with various concentrations (0.5–30 μM) of curcumin
analogues for 72 h. Following treatment,
3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazoliumbromide was
added to each well of the plate and incubated for 1 h. After
careful removal of the medium, 0.1 ml DMSO was added to each well
and absorbance at 550 nm was recorded on a microplate reader. For
the trypan blue exclusion assay, the CWR-22Rv1 cells were seeded at
a density of 2×104 cells/ml of medium in 35 mm tissue
culture dishes and incubated for 24 h. The cells were then treated
with curcumin analogues for 96 h. The number of viable cells
following each treatment was determined using a hemocytometer under
a light microscope (Nikon Optiphot; Nikon, Tokyo, Japan). The cell
viability was determined by the trypan blue exclusion assay, which
was performed by mixing 80 μl of the cell suspension and 20 μl of
0.4% trypan blue stain solution for 2 min. The blue cells were
counted as dead cells and the cells that did not absorb dye were
counted as live cells. Apoptosis was determined by morphological
assessment in the cells stained with propidium iodide (25). Apoptotic cells were identified by
classical morphological features, including nuclear condensation,
cell shrinkage and the formation of apoptotic bodies. At least 200
cells were counted in each sample and the percentage of apoptotic
cells was determined.
AR luciferase reporter assay
AR transcriptional activity was measured by an
AR-luciferase reporter gene expression assay. An AR luciferase
construct was stably transfected into CWR-22Rv1 cells and a single
stable clone, CWR-22Rv1/AR, was used in the present study.
CWR22-Rv-1 cells cultured in 10% FBS RPMI-1640 medium were infected
with a lentivirus carrying the Cignal Lenti AR reporter
(luciferase; Qiagen, Valencia, CA, USA) in the medium containing 8
μg/ml Polybrene (Sigma, St. Louis, MO, USA). At 6 h following
infection, the culture medium was replaced with fresh 10% RPMI-1640
medium. To establish the cells expressing stable AR-luciferase
reporter, cells were selected using puromycin (5 μg/ml) on day 3
following infection for one week. The selected cells were then used
for the reporter assay for AR activity.
The CWR-22Rv1/AR cells were treated with curcumin
and its analogues for 24 h, and the luciferase activities were
measured using luciferase assay kits from Promega Corporation
(Madison, WI, USA). Following treatment, the cells were washed with
ice-cold phosphate-buffered saline (PBS) and harvested in a
reporter lysis buffer. After centrifugation, 10 μl aliquots of the
supernatants were used for measuring the luciferase activity with a
luminometer from Turner Designs Instruments (Sunnyvale, CA, USA).
The luciferase activity was normalized against protein
concentration and expressed as the percentage of luciferase
activity in the control cells, which were treated with DMSO
solvent. The protein level was determined by Bio-Rad protein assay
kits (Bio-Rad, Hercules, CA, USA) according to the manufacturer’s
instructions.
Western blot analysis
Following treatment with curcumin, A10, B10, C10,
E10 and F10 for 24 h, the CWR-22Rv1 and LNCaP cells were washed
with ice-cold PBS and lysed with 800 μl of lysis buffer (10 mm
Tris-HCl, pH 8.0, 10 mm EDTA, 150 mm sodium chloride, 1% NP-40,
0.5% SDS, in deionized water). The homogenates were centrifuged at
12,000 × g for 15 min at 4°C. The protein concentration of whole
cell lysates was determined with a Bio-Rad protein assay kit
(Bio-Rad). Equal amounts (50 μg) of protein were then resolved on a
10% Criterion Precast Gel (Bio-Rad) and transferred onto a PVDF
membrane using a semi-dry transfer system. The membrane was then
probed with anti-PSA (Santa Cruz Biotechnology, Inc., Santa Cruz,
CA, USA) primary antibody. Following hybridization with primary
antibody, the membrane was washed with Tris-buffered saline (TBS)
three times, then incubated with horseradish peroxidase-conjugated
secondary antibody (Santa Cruz Biotechnology, Inc.) and washed with
TBS three times. Final detection was performed with enhanced
chemiluminescent reagents. The extent of protein loading was
determined by blotting for β-actin. The membrane was incubated in
stripping buffer (100 mm β-mercaptoethanol, 2% SDS and 62.5 mm
Tris-HCl at pH 6.7) at 50°C for 30 min with occasional agitation
prior to incubating in blocking buffer and re-probing using
anti-β-actin (Santa Cruz Biotechnology, Inc.).
Statistical analyses
The analyses of differences among curcumin and its
analogues on the TT- or DHT-induced activation of AR were based on
a repeated measurement model. The effects of the treatments were
assessed by comparing the rates of change over time between the
treatment groups (i.e., comparing the slopes between the treatment
groups). The analysis of variance (ANOVA) method with the
Tukey-Kramer test was used for the comparison of effects among the
different treatment groups at the end of the study. P<0.05 was
considered to indicate a statistically significant difference.
Results
Effects of curcumin and its analogues on
CWR-22Rv1 and LNCaP cells
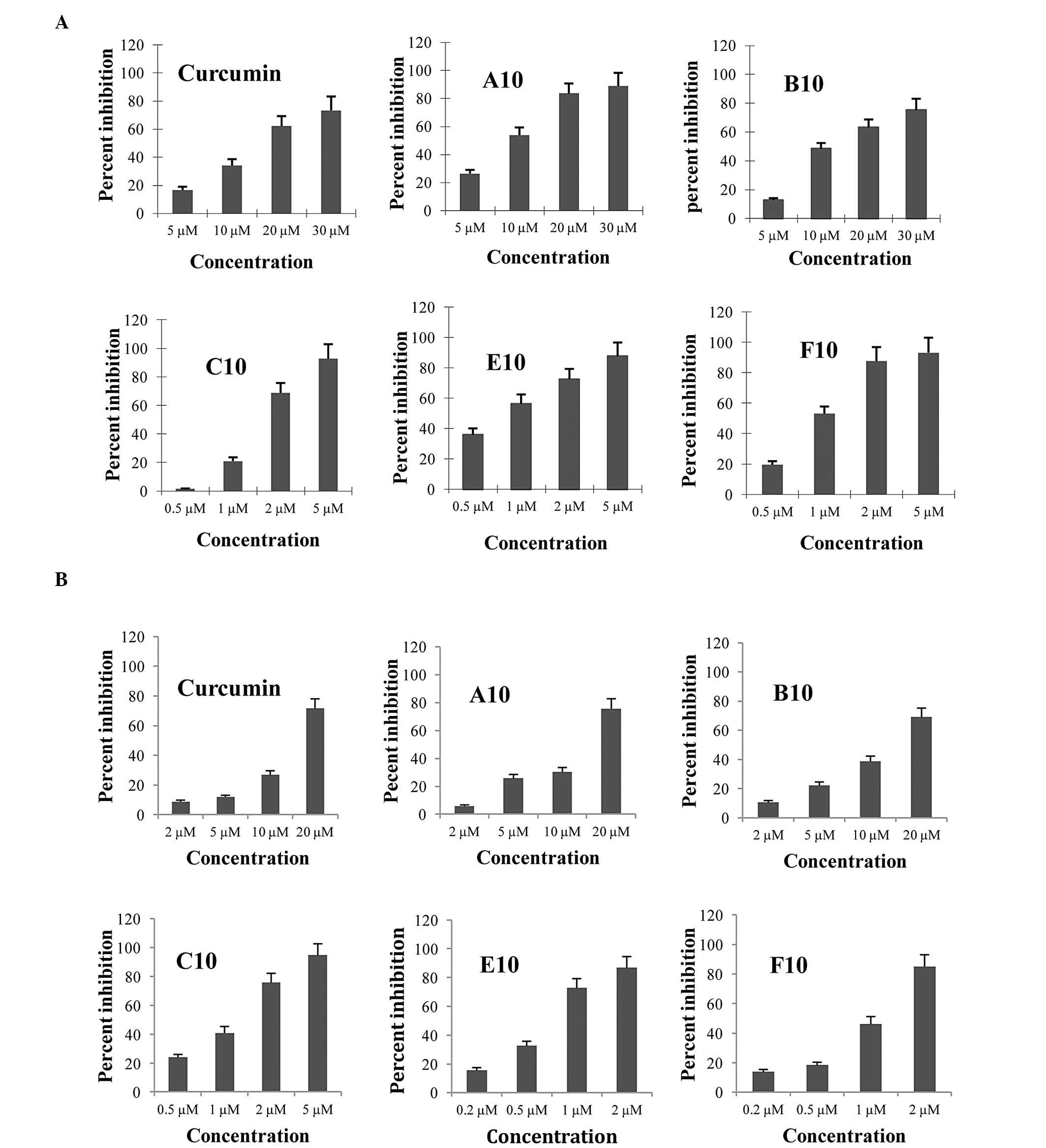
The inhibitory effects of curcumin analogues on the
growth of cultured CWR-22Rv1 and LNCaP cells were determined by
using MTT and trypan blue exclusion assays. For each incubation,
curcumin was examined as a positive control. The inhibitory effects
of different concentrations of curcumin and its analogues in
cultured CWR-22Rv1 and LNCaP cells are presented in Fig. 2. All of the compounds had stronger
inhibitory effects than curcumin as determined by the MTT assay.
Among the five curcumin analogues tested in the present study,
compounds E10, F10 and C10 exhibited the most potent inhibitory
effects on the growth of cultured CWR-22Rv1 and LNCaP cells. The
IC50 values for E10 and F10 were lower than 1 μM in the
CWR-22Rv1 and LNCaP cells, indicating that these compounds were
~20-fold more active than curcumin (IC50=16.99 μM). As
demonstrated in Table I, the
IC50 values of the five curcumin analogues ranged from
0.82 to 13.62 μM. The numbers of viable and dead cells were
determined by the trypan blue exclusion assay following treatment
of the CWR-22Rv1 cells with curcumin and its analogues for 96 h. As
demonstrated in Table II, a
reduction in the number of viable cells were observed. Compared
with the control group, the numbers of viable cells in the various
groups treated with curcumin and its analogues were decreased by
15.4% to 95.4% (Table II). The
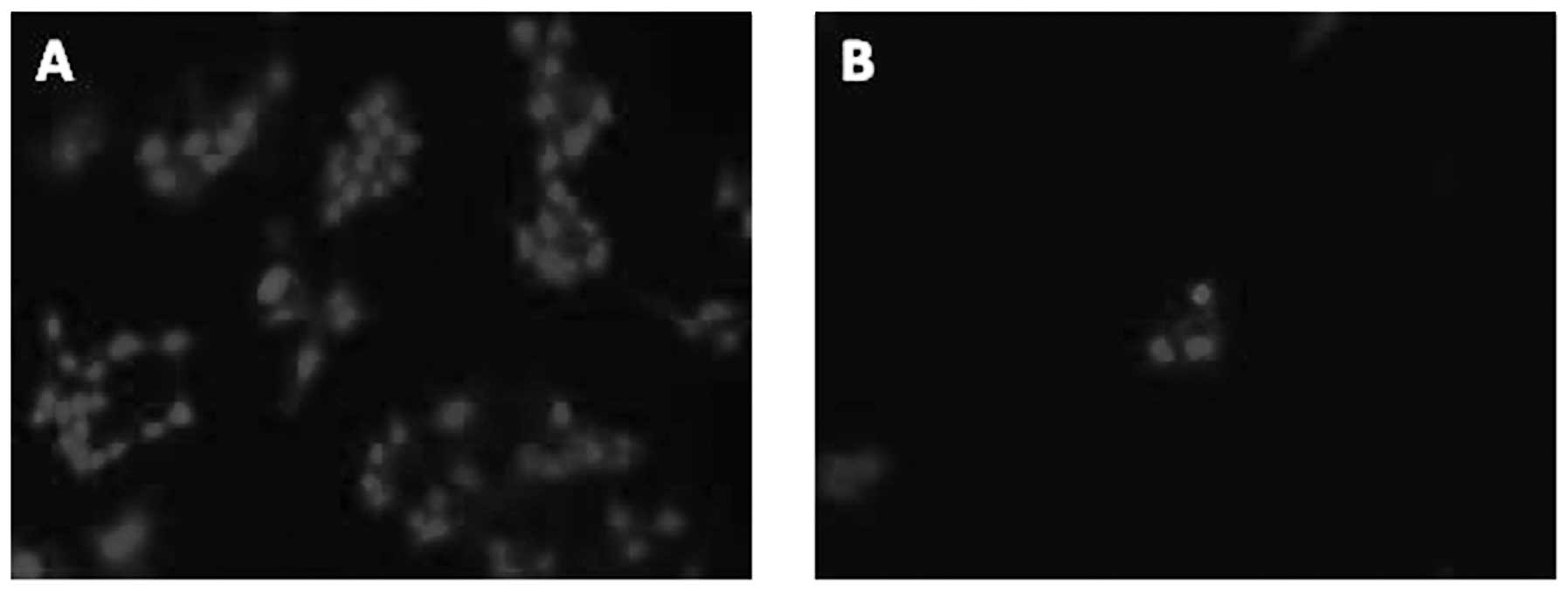
effects of curcumin and its analogues on apoptosis of CWR-22Rv1
cells were determined by morphological assessment in the propidium
iodide stained cells. In these studies, the CWR-22Rv1 cells were
treated with curcumin and its analogues for 96 h. As demonstrated
in Table II, weak inhibitory
effects on growth and weak stimulatory effects on the induction of
apoptosis of CWR-22Rv1 cells were observed by treatment with A10 (1
μM) and B10 (1 μM), while more evident effects were observed by
treatment with E10 (0.5 μM) and F10 (0.5 μM). C10 (1 μM) had a
moderate inhibitory effect on growth and a moderate stimulatory
effect on apoptosis.
 | Table IInhibitory effects of curcumin and its
analogues on the growth of CWR-22Rv1 cells. |
Table I
Inhibitory effects of curcumin and its
analogues on the growth of CWR-22Rv1 cells.
| IC50
(μM) |
|---|
|
|
|---|
| Compound | CWR-22Rv1 | LNCaP |
|---|
| Curcumin | 16.99±3.0 | 13.59±1.8 |
| A10 | 8.76±0.5 | 9.8±1.2 |
| B10 | 13.62±2.0 | 11.4±1.5 |
| C10 | 1.78±0.2 | 1.07±0.2 |
| E10 | 0.82±0.1 | 0.65±0.2 |
| F10 | 0.96±0.1 | 0.81±0.2 |
| Table IIEffects of curcumin and its analogues
on the growth of CWR-22Rv1 cells. |
Table II
Effects of curcumin and its analogues
on the growth of CWR-22Rv1 cells.
| Treatment | No. of viable cells
(1×10−4) | Percent apoptotic
cells |
|---|
| Control | 43.1±1.4 | 1.7±0.2 |
| Curcumin | 36.4±1.5 | 2.3±0.3 |
| A10 | 30.5±1.3 | 4.7±0.9 |
| B10 | 33.0±1.0 | 3.1±0.3 |
| C10 | 12.4±0.9 | 7.6±0.5 |
| E10 | 8.5±0.8 | 22.8±1.0 |
| F10 | 2.0±0.3 | 29.4±2.1 |
Effects of curcumin and its analogues on
AR activity in CWR-22Rv1/AR cells
An AR-luciferase reporter gene expression assay in
CWR-22Rv1/AR cells was utilized to determine the effect of curcumin
and its analogues on the TT- or DHT-induced activation of AR.
Cultured CWR-22Rv1/AR cells were treated with TT in combination
with curcumin analogues (1 μM) for 24 h. As demonstrated in
Fig. 3, a marginal inhibitory
effect on the TT- or DHT-induced increase in AR activity was
observed in the cultured CWR-22Rv1/AR cells treated with curcumin
(1 μM), A10 (1 μM) or B10 (1 μM), while more evident inhibitory
effects were observed in the CWR-22Rv1/AR cells treated with C10 (1
μM), E10 (1 μM) and F10 (1 μM). Statistical analysis using ANOVA
with the Tukey’s multiple comparison tests demonstrated that AR
activity was significantly lower in the cells treated with group
C10, E10 and F10 compounds than in the cells treated with curcumin
or A10 and B10. P<0.001 for control vs. TT (DHT) + CUR, TT (DHT)
+ A10 or TT (DHT) + B10; P<0.001 for TT (DHT) vs. TT (DHT) +
A10, TT (DHT) + B10, TT (DHT) + C10, TT (DHT) + E10 or TT (DHT) +
F10; P<0.001 for TT (DHT) + CUR vs. TT (DHT) + C10, TT (DHT) +
E10 or TT (DHT) + F10; P<0.001 for TT (DHT) + A10 vs. TT (DHT) +
C10, TT (DHT) + E10 or TT (DHT) + F10; P<0.001 for TT (DHT) +
B10 vs. TT (DHT) + C10, TT (DHT) + E10 or TT (DHT) + F10.
Effects of curcumin and its analogues on
the expression of PSA in CWR-22Rv1 and LNCaP cells
The levels of PSA were evaluated by western blot
analysis using an anti-PSA antibody. The cultured CWR-22Rv1 and
LNCaP cells were treated with TT, DHT and curcumin analogues A10,
B10, C10, E10 or F10 for 24 h, and the expression of PSA was
analyzed by western blotting. As demonstrated in Fig. 4, treatment of CWR-22Rv1 and LNCaP
cells with F10 and E10 resulted in a marked decrease in the level
of PSA while the other compounds (curcumin, A10, B10 and C10) were
less active. The results indicate that the effects of E10, F10, A10
and C10 on CWR-22Rv1 and LNCaP cells were all associated with a
decrease in PSA in TT- and DHT-induced cells. B10 was inactive.
 | Figure 4Effect of curcumin and its analogues
on TT- and DHT-induced increase in PSA formation in the CWR-22Rv1
cells. (A) TT and (B) DHT. Effect of curcumin and its analogues on
the activation of AR in CWR-22Rv1 and LNCaP cells. (C) CWR-22Rv1
and (D) LNCaP. The CWR-22Rv1 and LNCaP cells were seeded at a
density of 1×104 cells/ml of medium in 100 mm culture
dishes (10 ml/dish) and incubated for 24 h. The medium was changed
to RPMI without fetal bovine serum, and the cells were then treated
with vehicle, (A) 100 nm TT or (B) 10 nm alone or together with 1
μM curcumin, A10, B10, C10, E10 or F10 for 24 h. PSA was determined
by western blot analysis with anti-PSA antibody. The extent of
protein loading was determined by blotting for β-actin, and the
levels of PSA in the western blots were analyzed by optical density
measurements and normalized for β-actin to obtain the RD for the
various samples. Representative blots from three experiments are
demonstrated. TT, testosterone; DHT, dihydrotestosterone; PSA,
prostate specific antigen; RD, relative optical density; CON,
control; CUR, curcumin; AR, androgen receptor. |
Discussion
In the present study, it was identified that several
curcumin analogues (A10, B10, C10, E10 and F10) exhibited stronger
anticancer activities than curcumin in cultured human prostate
cancer CWR-22Rv1 and LNCaP cells. Among the curcumin analogues,
compounds F10 and E10 demonstrated a more potent inhibitory effect
on the growth of CWR-22Rv1 and LNCaP cells than any of the other
curcumin analogues, and they also had higher stimulatory effects on
apoptosis in CWR-22Rv1 cells compared with the other compounds
(Fig. 5). In addition, it was
identified that all curcumin analogues examined (except for B10)
were more potent inhibitors of AR in LNCaP and CWR-22Rv1 cells than
curcumin. E10 and F10 were the most potent compounds among the five
curcumin analogues examined for inhibiting the activation of
AR.
The natural product curcumin (diferuloylmethane) has
been demonstrated to inhibit numerous targets in prostate
epithelial cells with an importance in cancer formation and
progression. Among these targets are transcription factors,
receptors, intracellular kinases, cytokines and growth factors
(26). The effect of curcumin on
the AR and on its target PSA has been demonstrated by several
independent investigators using both endogenously expressed AR in
LNCaP cells and ectopically expressed AR in PC-3 cells (27,28).
However, in these studies, curcumin was used at relatively high
concentrations, typically at >20 μM. It has previously been
reported that curcumin has poor bioavailability, which has been
determined in animal models and humans (29). This limitation has led researchers
to generate a variety of synthetic analogues of curcumin and to
investigate their capability to affect a number of molecular
pathways implicated in tumorigenesis and cancer progression
(30–33). Typical structure modifications
include the introduction of substituents on the phenyl rings and
modifications of the length of the linker between the phenyl rings.
A specific group of such analogues has been exploited for their
ability to inhibit AR function (34), and a number of these agents have
been demonstrated to inhibit the expression of AR (35).
The present study determined the inhibitory effects
of different curcumin analogues on TT- and DHT-induced activation
of AR in CWR-22Rv1 cells. The results indicate that the different
curcumin analogues had a similar effect on TT- or DHT-induced AR
activation.
Based on the analysis of the association between the
structures of curcumin-related compounds and their ability to
inhibit the growth of cultured cancer cells, the presence of groups
on the linker between the same aromatic rings was found to have a
key role in determining the anticancer activity of the various
analogues. Among the different series of curcumin-related
compounds, linear or cyclic linkers between the two aromatic rings
of curcumin-related compounds demonstrated different activity
trends. In general, the compounds with a tetrahydrothiopyran-4-one
(F10) or a tetrahydropyran-4-one (E10) linker exhibited the
strongest activity, and compounds with an acetone linker (C10)
exhibited moderate activity whereas compounds with a cyclohexanone
linker (A10) or a cyclopentanone linker (B10), were less active.
The activities of compounds with a heteroatom linker (E10 and F10)
demonstrated improved effects compared with those without a
heteroatom linker (A10, B10 and C10), which suggests that
flexibility of curcumin-like compounds may enhance their antitumor
activities by having interactions with the DNA of cancer cells and
disrupting the activity of transcription factors, such as AR.
Compounds with a small and highly rigid linker should be less
active as previously described (36,37).
The results indicate that one of the potential mechanisms for the
anticancer effect of curcumin analogues was inhibition of AR
pathways in human prostate cancer cells.
Acknowledgements
The present study was supported by the 2011
Guangdong Province Leadership Grant, China National Science
Foundation Grants (grant no. 81272452 and 21272043), and by
department funds from the Department of Chemical Biology in the
Ernest Mario School of Pharmacy at Rutgers University (Piscataway,
NJ, USA).
References
|
1
|
Siegel R, Ward E, Brawley O and Jemal A:
Cancer statistics, 2011: the impact of eliminating socioeconomic
and racial disparities on premature cancer deaths. CA Cancer J
Clin. 61:212–236. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gao W, Bohl CE and Dalton JT: Chemistry
and structural biology of androgen receptor. Chem Rev.
105:3352–3370. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Heinlein CA and Chang C: Androgen receptor
in prostate cancer. Endocr Rev. 25:276–308. 2004. View Article : Google Scholar
|
|
4
|
Klotz L: Hormone therapy for patients with
prostate carcinoma. Cancer. 88(Suppl): 3009–3014. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Simmons MN and Klein EA: Combined androgen
blockade revisited: Emerging options for the treatment of
castration-resistant prostate cancer. Urology. 73:697–705. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kuttan R, Bhanumathy P, Nirmala K, et al:
Potential anticancer activity of turmeric (Curcuma longa). Cancer
Lett. 29:197–202. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Huang MT, Smart RC, Wong CQ, et al:
Inhibitory effect of curcumin, chlorogenic acid, caffeic acid, and
ferulic acid on tumor promotion in mouse skin by
12-O-tetradecanoylphorbol-13-acetate. Cancer Res. 48:5941–5946.
1988.
|
|
8
|
Huang MT, Wang ZY, Georgiadis CA, et al:
Inhibitory effects of curcumin on tumor initiation by
benzo[a]pyrene and 7, 12-dimethylbenz[a]anthracene.
Carcinogenesis. 13:2183–2186. 1992.
|
|
9
|
Huang MT, Lou YR, Ma W, et al: Inhibitory
effects of dietary curcumin on forestomach duodenal, and colon
carcinogenesis in mice. Cancer Res. 54:5841–5847. 1994.PubMed/NCBI
|
|
10
|
Rao CV, Rivenson A, Simi B, et al:
Chemoprevention of colon carcinogenesis by dietary curcumin, a
naturally occurring plant phenolic compound. Cancer Res.
55:259–266. 1995.PubMed/NCBI
|
|
11
|
Leite KR, Chade DC, Sanudo A, et al:
Effects of curcumin in an orthotopic murine bladder tumor model.
Int Braz J Urol. 35:599–606. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Agrawal DK and Mishra PK: Curcumin and its
analogues: potential anticancer agents. Med Res Rev. 30:818–860.
2010.PubMed/NCBI
|
|
13
|
Kunnumakkara AB, Anand P and Aggarwal BB:
Curcumin inhibits proliferation, invasion, angiogenesis and
metastasis of different cancers through interaction with multiple
cell signaling proteins. Cancer Lett. 269:199–225. 2008. View Article : Google Scholar
|
|
14
|
Teiten MH, Gaascht F, Eifes S, et al:
Chemopreventive potential of curcumin in prostate cancer. Genes
Nutr. 5:61–74. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bill MA, Bakan C, Benson DM Jr, et al:
Curcumin induces proapoptotic effects against human melanoma cells
and modulates the cellular response to immunotherapeutic cytokines.
Mol Cancer Ther. 8:2726–2735. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Johnson SM, Gulhati P, Arrieta I, et al:
Curcumin inhibits proliferation of colorectal carcinoma by
modulating Akt/mTOR signaling. Anticancer Res. 29:3185–3190.
2009.PubMed/NCBI
|
|
17
|
Piantino CB, Salvadori FA, Ayres PP, et
al: An evaluation of the anti-neoplastic activity of curcumin in
prostate cancer cell lines. Int Braz J Urol. 35:354–360. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kuo CT, Chen BC, Yu CC, et al: Apoptosis
signal-regulating kinase 1 mediates denbinobin-induced apoptosis in
human lung adenocarcinoma cells. J Biomed Sci. 16:432009.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sahu RP, Batra S and Srivastava SK:
Activation of ATM/Chk1 by curcumin causes cell cycle arrest and
apoptosis in human pancreatic cancer cells. Br J Cancer.
100:1425–1433. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Thangapazham RL, Sharma A and Maheshwari
RK: Multiple molecular targets in cancer chemoprevention by
curcumin. AAPS J. 8:E443–E449. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Anand P, Sundaram C, Jhurani S, et al:
Curcumin and cancer: an ‘old-age’ disease with an ‘age-old’
solution. Cancer Lett. 267:133–164. 2008.
|
|
22
|
Dhillon N, Aggarwal BB, Newman RA, et al:
Phase II trial of curcumin in patients with advanced pancreatic
cancer. Clin Cancer Res. 14:4491–4499. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cheng AL, Hsu CH, Lin JK, et al: Phase I
clinical trial of curcumin, a chemopreventive agent, in patients
with high-risk or pre-malignant lesions. Anticancer Res.
21:2895–2900. 2001.PubMed/NCBI
|
|
24
|
Wei X, Du ZY, Zheng X, et al: Synthesis
and evaluation of curcumin-related compounds for anticancer
activity. Eur J Med Chem. 53:235–245. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zheng X, Chang RL, Cui XX, et al:
Inhibitory effect of 12-O-tetradecanoylphorbol-13-acetate alone or
in combination with all-trans-retinoic acid on the growth of LNCaP
prostate tumors in immunodeficient mice. Cancer Res. 64:1811–1820.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Aggarwal BB: Prostate cancer and curcumin:
add spice to your life. Cancer Biol Ther. 7:1436–1440. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Nakamura K, Yasunaga Y, Segawa T, et al:
Curcumin down-regulates AR gene expression and activation in
prostate cancer cell lines. Int J Oncol. 21:825–830.
2002.PubMed/NCBI
|
|
28
|
Tsui KH, Feng TH, Lin CM, et al: Curcumin
blocks the activation of androgen and interlukin-6 on prostate
specific antigen expression in human prostatic carcinoma cells. J
Androl. 29:661–668. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Anand P, Kunnumakkara AB, Newman RA, et
al: Bioavailability of curcumin: Problems and promises. Mol Pharm.
4:807–818. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Adams BK, Ferstl EM, Davis MC, et al:
Synthesis and biological evaluation of novel curcumin analogs as
anti-cancer and antiangiogenesis agents. Bioorg Med Chem.
12:3871–3883. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Basile V, Ferrari E, Lazzari S, et al:
Curcumin derivatives: molecular basis of their anti-cancer
activity. Biochem Pharmacol. 78:1305–1315. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ishida J, Ohtsu H, Tachibana Y, et al:
Antitumor agents. Part 214: synthesis and evaluation of curcumin
analogues as cytotoxic agents. Bioorg Med Chem. 10:3481–3487. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ohori H, Yamakoshi H, Tomizawa M, et al:
Synthesis and biological analysis of new curcumin analogues bearing
an enhanced potential for the medicinal treatment of cancer. Mol
Cancer Ther. 5:2563–2571. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhou J, Geng G, Shi Q, et al: Design and
synthesis of androgen receptor antagonists with bulky side chains
for overcoming antiandrogen resistance. J Med Chem. 52:5546–5550.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Shi Q, Shih CC and Lee KH: Novel
anti-prostate cancer curcumin analogues that enhance androgen
receptor degradation activity. Anticancer Agents Med Chem.
9:904–912. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zsila F, Bikadi Z and Simonyi M: Circular
dichroism spectroscopic studies reveal pH dependent binding of
curcumin in the minor groove of natural and synthetic nucleic
acids. Org Biomol Chem. 2:2902–2910. 2004. View Article : Google Scholar
|
|
37
|
Caruso F, Rossi M, Benson A, et al:
Ruthenium-arene complexes of curcumin: X-ray and density functional
theory structure, synthesis, and spectroscopic characterization, in
vitro antitumor activity, and DNA docking studies of (p-Cymene) Ru
(curcuminato) chloro. J Med Chem. 55:1072–1081. 2012. View Article : Google Scholar
|