Introduction
Resveratrol, a non-flavone polyphenol compound with
a stilbene structure (Fig. 1A), is
highly enriched in several dietary and pharmaceutical sources,
including grapes, peanuts, berries, red wine and Polygonum
cuspidatum (1,2). Previous studies have revealed that it
has unique, beneficial effects on human health, such as
cardiovascular protection, lifespan prolongation, anti-inflammatory
effects, microcirculation improvement and regulation of lipid
metabolism. In addition, a previous study documented that
resveratrol has a chemopreventive and chemotherapeutic effect
against the progression of various types of cancer, including
prostate, breast, liver, skin and lung cancer (3).
Cancer is a disease characterized by loss of control
over cellular growth, which evolves, in part by over-riding the
regulation of cellular proliferation (4). The progress of the cell cycle in
cancer cells is regulated by three protein families: Cyclins,
cyclin-dependent kinases (CDKs) and CDK inhibitors (CDKIs). CDKs
are critical regulators of the cell cycle machinery, which, when
activated, provide a means to progress the cell cycle from one
phase to the next (5). However,
multiple changes occur in cancer cells, including cyclin
amplification, CDK or substrate mutation, as well as inactivation
of inhibitors. This results in abnormal CDK activity, amplification
of positive growth signals, mutation of checkpoint and surveillance
genes, as well as dysregulation of programmed cell death or
apoptotic processes, inducing the selective growth advantage of
cancer cells (6). Therefore,
identifying agents that may induce cell cycle arrest has become a
goal of cancer therapy, including small molecule inhibitors and
gene therapy.
p53, the tumor suppressor gene product, is a key
component in the regulation of cell cycle progression, which is
activated in response to a wide spectrum of stresses and damage
(7). A study demonstrated that p53
negatively regulates cell cycle progression in response to
different cellular stresses (8).
Commonly, when activated by genotoxic stress, p53 may directly
regulate the p21WAF1/CIP1/SDI1 gene, which encodes a
universal inhibitor of CDKs, to inhibit the cell cycle progression
(9).
The present study aimed to investigate the
anti-cancer effects of resveratrol on the A549 lung cancer cell
line in order to confirm the function of the p53-independent
pathway in resveratrol-induced cell cycle arrest in A549 cells.
Materials and methods
Reagents and antibodies
Resveratrol, purchased from Sigma-Aldrich (St.
Louis, MO, USA), was dissolved at a concentration of 50 mmol/l in
dimethysulfoxide (DMSO; MP Biomedicals, LLP, Santa Ana, CA, USA)
stored at −20°C and diluted with Dulbecco’s modified Eagle’s medium
(DMEM; Gibco-BRL, Carlsbad, CA, USA) to the desired working
concentrations. The final concentration of DMSO did not exceed 0.4%
(v/v) throughout the study. Pifithrin-α was purchased from
Sigma-Aldrich and diluted to a final concentration of 5 mg/ml. DAPI
was obtained from Beyotime Institute of Biotechnology (Haimen,
China).
The primary monoclonal human anti-rabbit antibodies
against cyclin D1 (#2926), CDK4 (#2906), CDK6 (#3136), p21 (#2947),
p27 (#2552), p53 (#9282) and GAPDH (#2118) were all obtained from
Cell Signaling Technology, Inc. (Danvers, MA, USA). The horseradish
peroxidase-conjugated anti-rabbit (E030220) and anti-mouse IgG
secondary antibody was purchased from EarthOx (San Francisco, CA,
USA) and the Alexa Fluor 488 labeled anti-rabbit IgG secondary
antibody was obtained from Beyotime Institute of Biotechnology.
Cell culture
A549 cells were obtained from the Cancer Cell
Repository (Shanghai Cell Bank, Shanghai, China). Cells were
maintained in DMEM supplemented with 10% (v/v) heat-inactivated
fetal bovine serum (Gibco-BRL) and antibiotics (100 U/ml penicillin
and 100 U/ml streptomycin; Hyclone Laboratories, Inc., Logan, UT,
USA) at 37°C in a humidified atmosphere of 5% CO2.
Cell proliferation assay
The cells were plated at a density of
5×104 cells/ml in 96-well microtiter plates. Different
concentrations of resveratrol (25, 50, 100 and 150 μm/l) were used
to treat cells for 24, 48 and 72 h. A total of 20 μl MTT solution
(5 mg/ml; Sigma-Aldrich) was transferred to each well. Plates were
incubated for 4 h at 37°C. Following incubation, supernatants were
removed and 100 μl DMSO was added to ensure total solubility of
formazan crystals. After 15 min of agitation, the absorbance was
measured at 570 nm using a plate reader (Perkin-Elmer Inc.,
Waltham, MA, USA). The survival ratio was calculated using the
following equation: Survival ratio (%) = OD treated/OD
control × 100.
Cell cycle analysis
For cell cycle analysis, the cells were plated at a
density of 5×104 cells/ml in 6-well microtiter plates
for 24 h adherence and treated with different concentrations of
resveratrol or without for 48 h. Following treatment, the cells
were harvested by trypsinization and fixed in ice-cold 70% methanol
overnight at −20°C. The cells were subsequently centrifuged at 300
× g for 5 min and incubated with a propidium iodide (PI) working
solution (100 μg/ml PI and 100 μg/ml RNaseA) for 30 min at 37°C.
Cell cycle distribution was analyzed using a FACScan flow cytometer
(FACSCalibur; BD Biosciences, Franklin Lakes, NJ, USA).
Western blot analysis
The A549 cells were treated with various
concentrations of resveratrol (25, 50 and 100 μm/l) for 48 h and
lysed with radioimmunoprecipitation assay lysis buffer (Beyotime
Institute of Biotechnology, Jiangsu, China). The cell lysates were
centrifuged at 13,000 × gfor 10 min and the supernatant was
collected. Protein concentration was determined using the Bradford
protein assay (10). Total protein
(20 μl) was separated on 10–15% sodium-dodecyl sulfate
polyacrylamide gel electrophoresis and transferred onto a 0.45 μm
polyvinylidene difluoride membrane (Millipore, Billerica, MA, USA)
in a buffer containing 25 mmol/l Tris-HCl (pH 8.3), 192 mmol/l
glycine and 20% methanol. The membranes were then blocked with 5%
fat-free dry milk in Tris-buffered saline and Tween-20 containing
0.05% Tween-20 for 2 h and then incubated with primary antibodies
overnight at 4°C. Subsequently, treatment with the appropriate
horseradish peroxidase conjugated secondary antibody was performed
at a dilution of 1:2,000. The immunoreactive bands were detected
using the enhanced chemiluminescence method.
Immunofluorescence staining
Cells were incubated for 48 h with or without
resveratrol. All sloughed and attached cells were harvested, fixed
with ice-cold 4% paraformaldehyde (Beyotime Institute of
Biotechnology) for 10 min and washed with ice-cold
phosphate-buffered saline (PBS). Cells were then permeabilized with
0.3% Triton X-100 (Beyotime Institute of Biotechnology), washed
with ice-cold PBS (Jinuo, Hangzhou, China), stained with antibodies
against P53 and subsequently incubated with Alexa Fluor secondary
antibodies (Invitrogen Life Technologies, Carlsbad, CA, USA). Cell
nuclei were stained with DAPI (Beyotime Institute of Biotechnology)
and the cells were observed using a fluorescence microscope (TCS
SP5II, Germany) with peak excitation wavelengths at 570 nm and 460
nm.
Statistical analysis
Values are presented as the mean ± standard
deviation of at least three independent experiments. Data were
evaluated using the Student’s t-test and GraphPad Prism version 5
(GraphPad Software, Inc., La Jolla, CA, USA). P<0.05, P<0.01
or P<0.001 were considered to indicate statistically significant
differences between values.
Results
Resveratrol inhibits cell proliferation
in A549 cells
To examine the antitumor effects of resveratrol,
A549 cells were treated with different concentrations of
resveratrol for 24, 48 and 72 h and then cell viability was
determined using an MTT assay. As shown in Fig. 1B, resveratrol inhibited the cell
viability of A549 cells in a concentration- and time-dependent
manner. As the microscopy images in Fig. 1C show, following exposure to 25, 50
or 100 μmol/l resveratrol for 48 h, the A549 cells exhibited
morphological features, including shrinkage and distortion; in
addition, certain cells became rounded, while the number of
adherent cells decreased significantly, which was consistent with
the cell proliferation assay.
Resveratrol induces cell cycle arrest in
G0/G1 phase
To demonstrate the possible mechanism of
resveratrol-induced cell growth inhibition in A549 cells, cell
cycle progression was analyzed. A549 cells were treated with 25, 50
or 100 μmol/l resveratrol for 48 h and then the cell cycle phase
was analyzed using flow cytometry. As shown in Fig. 2A, upon treatment with 25 μmol/l
resveratrol, the S phase distribution increased between 33.91 and
60.50%, but with the increase in the concentration of reveratrol,
the percentage of cells in S phase was decreased. While the
G0/G1 phase distribution was increased
between 53.96, 66.84 and 72.30% when treated with 25, 50 and 100
μmol/l resveratrol respectively, this was accompanied by a cell
cycle G0/G1 phase distribution increase and S
phase distribution decrease.
Effects of resveratrol on proteins
involved in G0/G1 phase arrest
In view of the progression of the cell cycle in
vertebrate cells, cells prepare for S phase during the
G1 phase, which is triggered by regulation of cyclin D1,
CDK4 and CDK6 (11,12). The formation of cyclin D1-CDK4/6
complexes regulate the synthesis of DNA to prepare for cell
division, while the CKIs, including p21 and p27 have a negative
regulatory role in this process (13,14).
The western blot analysis (Fig.
2B) revealed that resveratrol treatment reduced the protein
levels of cyclin D1, CDK4 and CDK6 significantly, and resulted in
upregulation of the proteins levels of p21 and p27 in a
concentration-dependent manner.
Resveratrol induces cell cycle arrest,
however not via p53
p53 is a regulator of the cell cycle and the
inactivation of p53 has an important role in tumor formation
(15). However, it remains to be
determined whether p53 is involved in cell cycle arrest induced by
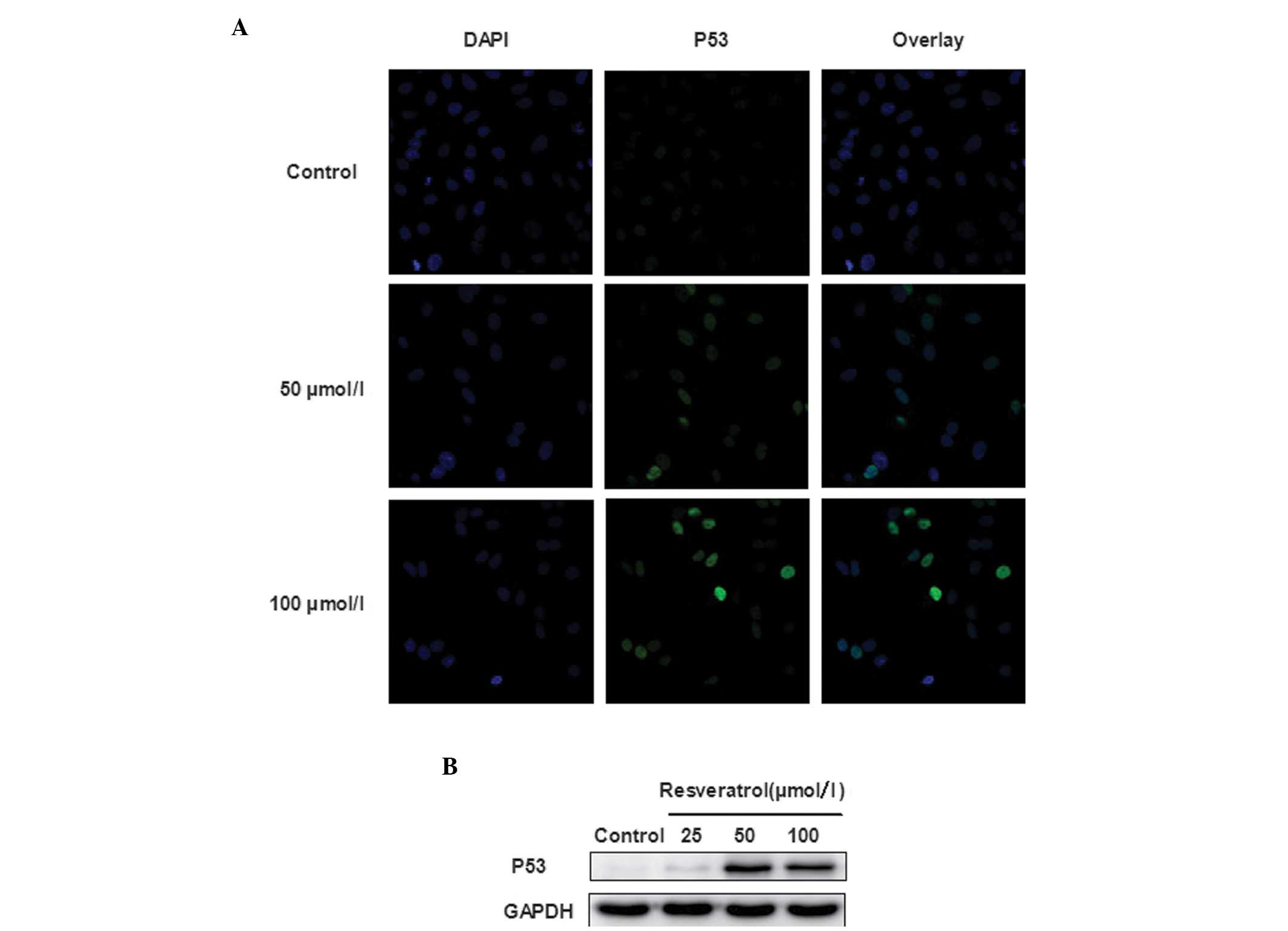
resveratrol. As shown in Fig. 3A and
B, it was demonstrated that resveratrol upregulated the protein
level of p53 significantly. Resveratrol treatment resulted in
nuclear accumulation of p53, inducing apoptosis in A549 cells. To
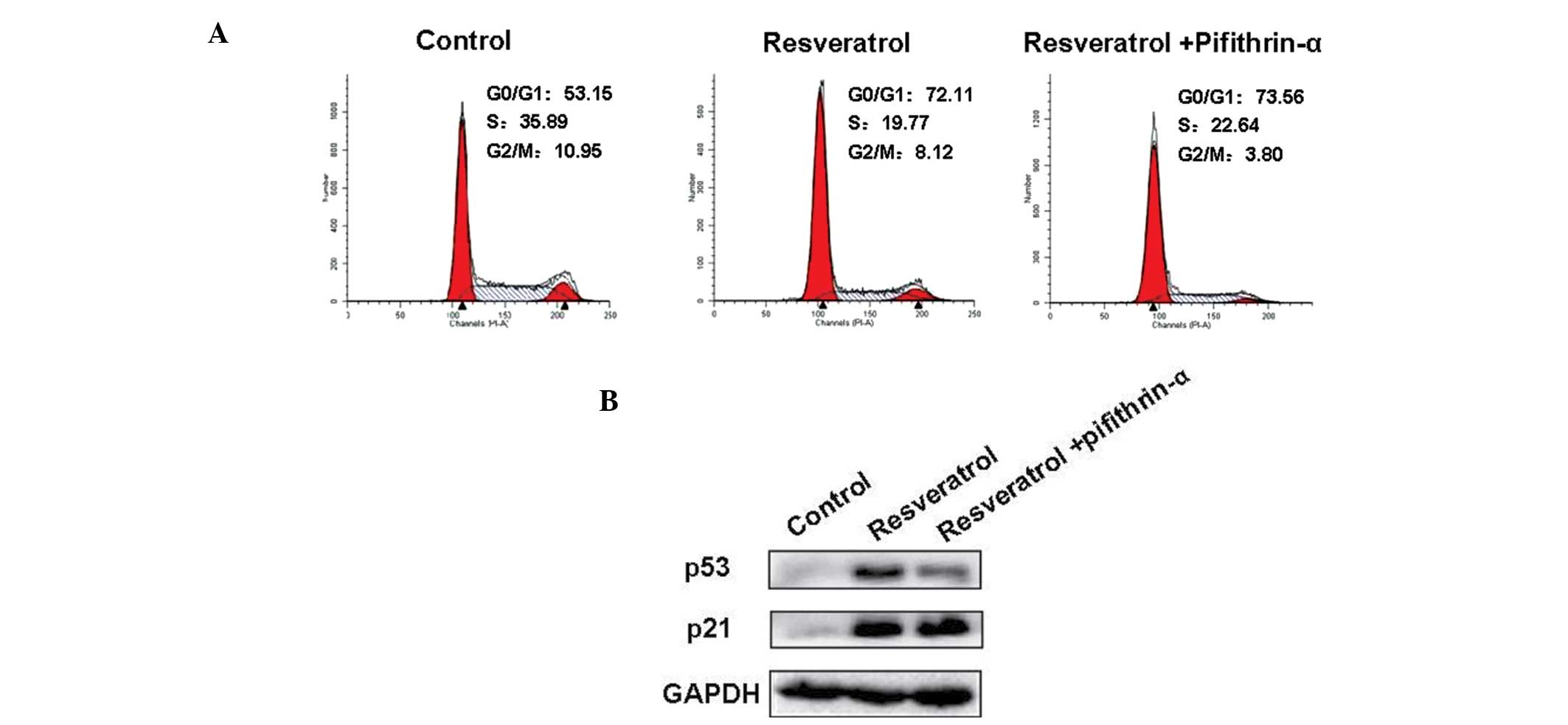
confirm the role of p53 in resveratrol-induced cell cycle arrest,
pifithrin-α, a p53 inhibitor was used to pre-treat A549 cells. The
results demonstrated that p53 downregulation did not induce cell
cycle arrest (Fig. 4A) or regulate
p21 (Fig. 4B) in A549 cells.
Discussion
Resveratrol is currently being evaluated as a
potential cancer chemopreventive compound (16). In the present study, the
chemopreventive/therapeutic potential of resveratrol was evaluated
in the A549 lung cancer cell line, and its underlying mechanism was
examined. Consistent with previous observations, it was identified
that resveratrol may inhibit cell proliferation through the
induction of cell cycle arrest in A549 cells in a time- and
concentration-dependent manner (17). However, an important observation of
the present study is that resveratrol induced cell cycle arrest in
the G0/G1 phase, this was accompanied by p53
upregulation, which may contribute to the antitumor effect of
resveratrol. However, the cell cycle arrest induced by resveratrol
was not reversed following treatment with an inhibitor of p53,
pifithrin-α.
The potent cytotoxic action of resveratrol was
demonstrated by a time- and concentration-dependent decrease in
cell viability of A549 cells, which was concomitant with an
increase in the percentage of cells that underwent cell cycle
arrest at the G0/G1 phase. Different classes
of cyclins and their complex formation with CDKs control the
different phases of cell cycle progression (18). G0/G1 phase
provides the materials for mitosis, and DNA replication occurs
during S phase, which is regulated by cyclin D1, CDK4 and CDK6. In
the present study, flow cytometric analysis revealed that
resveratrol induced A549 cells to arrest in the
G0/G1 phase and it was associated with a
decrease in the protein level of cyclin D1, CDK4 and CDK6.
CDK-cyclin complexes, the motors of the cell cycle, are regulated
by two families of CKIs, the CIP/KIP family (p21WAF/CIP,
p27KIP1 and p57KIP2) and the INK4 family
(p15INK4b, p16INK4a, p18INK4c and p19INK4d) (19). p21WAF1/CIP1, a member of
the CIP/KIP family, binds to CDK/cyclin complexes and prevents
kinase activation, subsequently halting the cell cycle in the
G1 phase. In response to cell cycle arrest induced by
antitumor agents, it was demonstrated that the cell cycle may be
regulated through either p53-dependent or p53-independent pathways
(20,21). In the present study,
p21WAF1/CIP1 was upregulated by resveratrol in A549
cells accompanied with a significant increase in the protein level
of p53. The present results provided a possible mechanism, through
which resveratrol upregulated the expression level of
p21WAF1/CIP1 and thereby led to
G0/G1 phase arrest through a p53-dependent
pathway. However, further investigation revealed that the
inhibition of p53 had no effect on resveratrol-induced cell cycle
arrest and the expression of P21WAF1/CIP1.
In conclusion, the present study may offer a
scientific basis for the further in-depth evaluation of the tumor
suppressor, resveratrol. Resveratrol inhibited cell proliferation
in a concentration- and time-dependent manner in A549 cells by
inducing cell cycle arrest at the G0/G1 phase
via p21 upregulation, accompanied by p53 activation. However, the
resveratrol-induced p21-mediated G0/G1 arrest
was not dependent on the p53 pathway, which indicated that p53 may
not exhibit effects on cell-cycle arrest-induced cell growth in
A549 cells.
References
|
1
|
Chen BY, Kuo CH, Liu YC, et al:
Ultrasonic-assisted extraction of the botanical dietary supplement
resveratrol and other constituents of Polygonum cuspidatum. J Nat
Prod. 75:1810–1813. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Liu P, Wang X, Hu C and Hu T: Inhibition
of proliferation and induction of apoptosis by trimethoxyl stilbene
(TMS) in a lung cancer cell line. Asian Pac J Cancer Prev.
12:2263–2269. 2011.
|
|
3
|
Shukla Y and Singh R: Resveratrol and
cellular mechanisms of cancer prevention. Ann NY Acad Sci.
1215:1–8. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Clarke MF and Fuller M: Stem cells and
cancer: two faces of eve. Cell. 124:1111–1115. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Schwartz GK and Shah MA: Targeting the
cell cycle: a new approach to cancer therapy. J Clin Oncol.
23:9408–9421. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
McDonald E and El-Deiry W: Cell cycle
control as a basis for cancer drug development (Review). Int J
Oncol. 16:8712000.PubMed/NCBI
|
|
7
|
Horn H and Vousden K: Coping with stress:
multiple ways to activate p53. Oncogene. 26:1306–1316. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bourougaa K, Naski N, Boularan C, et al:
Endoplasmic reticulum stress induces G2 cell-cycle arrest via mRNA
translation of the p53 isoform p53/47. Mol Cell. 38:78–88. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Vogelstein B, Lane D and Levine AJ:
Surfing the p53 network. Nature. 408:307–310. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
López J, Imperial S, Valderrama R and
Navarro S: An improved Bradford protein assay for collagen
proteins. Clinica chimica Acta. 220:91–100. 1993. View Article : Google Scholar
|
|
11
|
Aggarwal P, Vaites LP, Kim JK, et al:
Nuclear cyclin D1/CDK4 kinase regulates CUL4 expression and
triggers neoplastic growth via activation of the PRMT5
methyltransferase. Cancer cell. 18:329–340. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bockstaele L, Bisteau X, Paternot S and
Roger PP: Differential regulation of cyclin-dependent kinase 4
(CDK4) and CDK6, evidence that CDK4 might not be activated by CDK7,
and design of a CDK6 activating mutation. Mol Cell Biol.
29:4188–4200. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Roy S, Gu M, Ramasamy K, et al: p21/Cip1
and p27/Kip1 are essential molecular targets of inositol
hexaphosphate for its antitumor efficacy against prostate cancer.
Cancer Res. 69:1166–1173. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Perisanidis C, Perisanidis B, Wrba F, et
al: Evaluation of immunohistochemical expression of p53, p21, p27,
cyclin D1, and Ki67 in oral and oropharyngeal squamous cell
carcinoma. J Oral Pathol Med. 41:40–46. 2012. View Article : Google Scholar
|
|
15
|
Suzuki K and Matsubara H: Recent advances
in p53 research and cancer treatment. J Biomed Bio.
2011:9783122011.
|
|
16
|
Joe AK, Liu H, Suzui M, Vural ME, Xiao D
and Weinstein IB: Resveratrol induces growth inhibition, S-phase
arrest, apoptosis, and changes in biomarker expression in several
human cancer cell lines. Clin Cancer Res. 8:893–903.
2002.PubMed/NCBI
|
|
17
|
Kim Y, Lee WH, Choi TH, Rhee SH, Park KY
and Choi YH: Involvement of p21WAF1/CIP1, pRB, Bax and NF-κB in
induction of growth arrest and apoptosis by resveratrol in human
lung carcinoma A549 cells. Int J Oncol. 23:1143–1149.
2003.PubMed/NCBI
|
|
18
|
Giacinti C and Giordano A: RB and cell
cycle progression. Oncogene. 25:5220–5227. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hwang HJ, Kang YJ, Hossain MA, et al:
Novel dihydrobenzofuro [4, 5-b][1, 8] naphthyridin-6-one
derivative, MHY-449, induces apoptosis and cell cycle arrest in
HCT116 human colon cancer cells. Int J Oncol. 41:2057–2064.
2012.PubMed/NCBI
|
|
20
|
Le NT and Richardson DR: The role of iron
in cell cycle progression and the proliferation of neoplastic
cells. Biochim Biophys Acta. 1603:31–46. 2002.PubMed/NCBI
|
|
21
|
Chatterjee SJ, Datar R, Youssefzadeh D, et
al: Combined effects of p53, p21, and pRb expression in the
progression of bladder transitional cell carcinoma. J Clin Oncol.
22:1007–1013. 2004. View Article : Google Scholar : PubMed/NCBI
|