Introduction
Reactive oxygen species (ROS) are known to have an
important role in atherosclerosis and associated vascular diseases
(1–3). In vascular physiology and
pathophysiology, the majority of previous studies have focused on
the role of the endothelium and vascular smooth muscle cells (VSMC)
in ROS production (4,5). However, increasing evidence has
suggested that vascular adventitia also had an important role in
the process of vascular remodeling (6,7).
Studies have shown that adventitia, rather than the intima or
media, was the main source of ROS in angiotensin II (AngII)-induced
hypertension (8); in addition,
adventitial fibroblasts were reported to participate in
AngII-induced vascular wall inflammation and remodeling (9). ROS production is primarily mediated
via the activation of nicotinamide adenine dinucleotide phosphate
(NADPH) oxidase in adventitia (10). A previous ex vivo study in
adventitial fibroblasts demonstrated that AngII reduced the
expression and activity of catalase, an important anti-oxidase, in
a time- and dose-dependent manner (11). These studies therefore suggested
that the imbalance between the pro-and antioxidant systems in
adventitia may contribute to vascular remodeling following vascular
injury.
Perivascular gene transfer of an adenoviral vector
suspended in pluronic gel was found to be an efficient approach for
the treatment of vascular injuries, which resulted in a minimal
inflammatory response (12,13).
Previous studies have shown that gene transfer of NADPH oxidase
inhibitors into adventitia significantly attenuated vascular
remodeling (8,14); however, there are a limited number
studies which have investigated the role of anti-oxidative enzymes
in the adventitia in vivo (15). In the present study, gene transfer
techniques were used to determine whether overexpression of
catalase through adventitial transfection improved AngII-induced
vascular remodeling in vivo. The present study also
investigated whether the p38 mitogen-activated protein kinase
(MAPK) pathway was involved in the underlying mechanisms of
AngII-induced vascular remodeling.
Materials and methods
Adenovirus (Ad) vector constructs
An adenoviral expression vector,
Ad-cytomegalovirus-human catalase (hCAT)-internal ribosome entry
site (IRES)-enhanced green fluorescent protein (eGFP)
(Ad-CAT-eGFP), was constructed in order to co-express human
catalase and eGFP, using the pAdEasy system (Stratagene, La Jolla,
CA, USA). Therefore, eGFP expression indicated the expression of
catalase. In brief, human catalase complementary DNA was cloned
into the pShuttle-IRES-eGFP vector. The resultant shuttle plasmid
was transformed into the Escherichia coli strain BJ5183
(Biochemistry Laboratory, Shanghai Jiao Tong University School of
Medicine, Shanghai, China) containing the adenoviral backbone
plasmid pAdEasy-1. Recombinant adenoviral plasmids were selected
based on kanamycin (Sangon Biotech Inc., Shanghai, China)
resistance and confirmed using restriction digestion. Virus
particles were obtained through transfection of recombinant
adenoviral plasmids into AD-293 cells (Invitrogen Life
Technologies, Carlsbad, CA, USA). The virus was purified using CsCl
banding (Amresco LLC, Solon, OH, USA) and the titer of the virus
stock was determined using a plaque assay, as previously described
(16). A control adenovirus
Ad-eGFP which expressed eGFP only was also constructed.
Animals and adventitial gene transfer of
Ad-CAT-eGFP or Ad-eGFP
A total of 30 male Sprague-Dawley (SD) rats
(6–8-weeks-old; 150–200 g) were purchased from the Shanghai
Laboratory Animal Center of the Chinese Academy of Sciences
(Shanghai, China) and maintained under environmentally controlled
conditions (temperature, 20±2°C; 12-h light/dark cycle). Rats were
anesthetized by intraperitoneal administration of sodium
pentobarbital (30 mg/kg; Sigma-Aldrich, St. Louis, MO, USA), then
the left common carotid artery was exposed (n=4–6 per group).
Ad-CAT-eGFP or Ad-eGFP suspended in 200 μl pluronic F127 gel (20%
wt/vol; Sigma-Aldrich) with a concentration of 1×109
plaque forming units/ml, was carefully applied to the adventitial
surface of the left common carotid artery (CCA). The incision was
closed and the rats were allowed to recover from the anesthesia
following coagulation of the gel. All protocols in the present
study were approved by the Institutional Animal Care and Use
Committee of Renji Hospital (Shanghai, China) and were consistent
with the Guide for the Care and Use of Laboratory Animals (National
Institutes of Health, Bethesda, MA, USA).
AngII treatment and systolic blood
pressure measurements
Two days post-adenovirus transfer, rats were
anesthetized, as described above, and osmotic minipumps (2002;
Alzet Osmotic Pumps, Cupertino, CA, USA) filled with AngII
(Sigma-Aldrich) were implanted subcutaneously. The rate of AngII
infusion was 0.75 mg/kg/day. Systolic blood pressure was measured
using a computerized tail-cuff system (BP98A; Softron, Tokyo,
Japan). Following 14 days of treatment, all 30 rats were euthanized
by intraperitoneal injection of sodium pentobarbital (50 mg/kg) and
CCAs were perfused with saline and then excised. The middle
portions of vessels were fixed using 4% paraformaldehyde
(Sigma-Aldrich) and embedded in paraffin (Sangon Biotech,
Inc.).
Morphometric and immunohistochemical
analyses
Vessel segments were serially sectioned (5 μm) and
then stained with modified hematoxylin and eosin (HE; Beyotime
Institute of Biotechnology, Jiangsu, China). Cross-sectional images
were captured using a light microscope (AxioSkop 20; Zeiss,
Oberkochen, Germany). The thickness and area of the media and lumen
were measured using Image-pro Plus 6.0 Software (Media Cybernetics,
Inc., Rockville, MD, USA). The collagen content was measured using
Picrosirius red staining (Beyotime Institute of Biotechnology), as
previously described (17). A
color threshold mask was defined in order to detect the red color
by sampling. A negative background (black) was selected for
thresholding and the positive area was measured by subtraction. The
area of positive staining was recorded for each section. Results
were obtained from two separate sections from 4–6 individual rats
in each group. For immunohistochemistry, sections were
deparaffinizated using xylene (Sangon Biotech, Inc.). and then
rehydrated through a graded ethanol series (Beyotime Institute of
Biotechnology), incubated with 0.3% hydrogen peroxide (Beyotime
Institute of Biotechnology) for 20 min and then blocked with 5%
goat serum (Beyotime Institute of Biotechnology), followed by
incubation with the primary antibodies against eGFP (rabbit
polyclonal anti-eGFP; 1:50; Cell Signaling Technologies, Danvers,
MA, USA), α-smooth muscle actin (mouse monoclonal anti-α-SMA;
1:100; Sigma-Aldrich), 4-hydroxy-2-nonenal (rabbit polyclonal
anti-4-HNE; 1:100; Abcam, Cambridge, MA, USA), CD68 (mouse
monoclonal anti-CD68; 1:100; AbD Serotec, Kidlington, UK) and
phosphorylated p38MAPK (rabbit polyclonal anti-phospho-p38MAPK;
1:100; Cell Signaling Technologies), followed by incubation with
horseradish peroxidase-conjugated polyclonal goat anti-rabbit or
goat anti-mouse secondary antibodies (1:200; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA). The reaction was visualized
using diaminobenzidine and sections were counterstained with
hematoxylin. Then the mean optical density (MOD) in each group was
measured using Image-pro Plus Software.
Cell culture
Adventitial fibroblasts (AFs) were isolated from
thoracic aortas of male SD rats as previously described (9). Cells were grown in Dulbecco’s
modified Eagle’s medium (DMEM; Invitrogen Life Technologies)
containing 20% fetal bovine serum (FBS; Invitrogen Life
Technolgies), 100 U/ml penicillin and 100 μg/ml streptomycin
(Sangon Biotech, Inc.). Subconfluent cells were made quiescent by
placing in 0% FBS-DMEM for an additional 24 h prior to
intervention. AFs were then administered AngII (1×10−7
mol/l) or polyethylene glycol-catalase (PEG-catalase; 1,000 U/ml;
Beyotime Institute of Biotechnology) in order to evaluate p38MAPK
phosphorylation levels under different conditions.
Western blot analysis
Following administration with different stimulants,
total protein was extracted from cells using
radioimmunoprecipitation assay lysis buffer in the presence of PMSF
(Beyotime Institute of Biotechnology) and protein concentrations
were measured using a bicinchoninic acid protein assay (Beyotime
Institute of Biotechnology). Protein samples were separated using
10% SDS-PAGE and transferred to nitrocellulose membranes
(Millipore, Bellerica, MA, USA). Following blocking with 5% non-fat
milk or 5% bovine serum albumin, membranes were then incubated with
primary antibodies against phosphorylated p38MAPK (rabbit
monoclonal anti-phospho-p38MAPK; 1:1,000; Cell Signaling
Technologies) and total p38MAPK (rabbit monoclonal anti-p38MAPK;
1:1,000; Cell Signaling Technologies), overnight at 4°C. The
membranes were then washed and incubated with horseradish
peroxidase-conjugated polyclonal goat anti-rabbit IgG secondary
antibodies (1:2,000; Santa Cruz Biotechnology, Inc.) for 1 h.
Western blots were developed using an enhanced chemiluminescence
detection kit (PerkinElmer, Inc., Waltham, MA, USA) and quantified
by densitometry using Gel-Pro Analyzer software 4.0 (Media
Cybernetics, Inc.).
Statistical analysis
Values are expressed as the mean ± standard
deviation. Student’s t-test or one-way analysis of variance were
performed in order to compare differences between two or multiple
groups, respectively. P<0.05 and P<0.01 were considered to
indicate a statistically significant difference between values. All
statistical analyses were performed using SPSS software 16.0 (SPSS
Inc., Chicago, IL, USA).
Results
Detection of adenovirus expression in CCA
in vivo
Immunohistochemical staining using antibodies
specific to eGFP was performed to determine whether Ad-CAT-eGFP was
successfully transfected into CCA following adventitial delivery.
As shown in Fig. 1, the carotid
arteries without gene transfer showed negative HE staining 14 days
following minipump implant (Fig.
1A). In carotid arteries treated with Ad-CAT-eGFP without AngII
infusion, positive staining was detected only in the adventitial
layer (Fig. 1B). However, the
carotid arteries of rats transfected with Ad-CAT-eGFP or Ad-eGFP
with AngII infusion stained positively with HE, predominantly in
media and neointima (Fig. 1C and
D); this therefore indicated that adventitial cells migrated
towards the media and neointima following vascular injury, as
previously described (18).
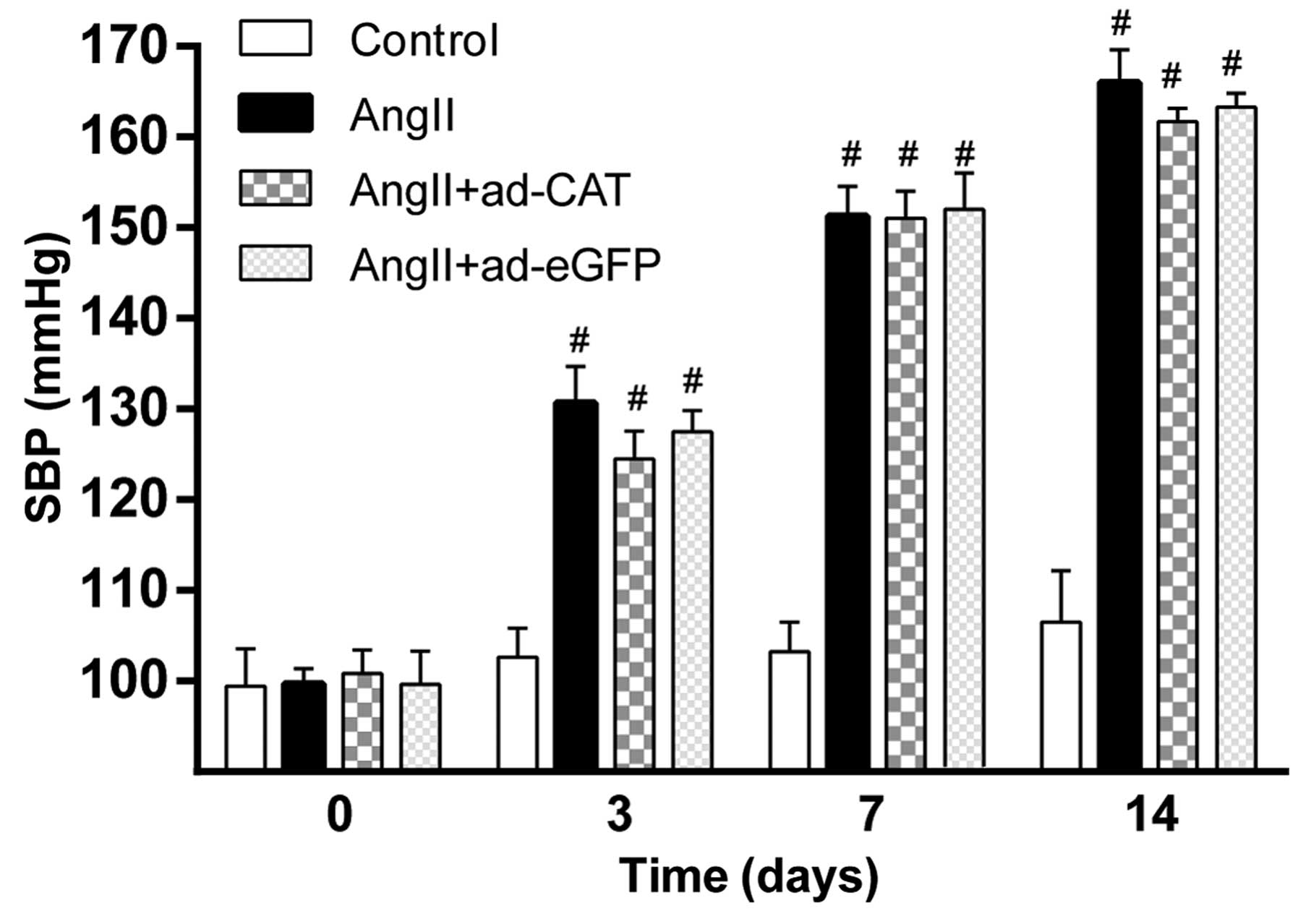
Catalase transfection has not effect on
the hypertensive response to AngII
In order to determine the effect of Ad-CAT-eGFP
transfection on blood pressure, the systolic blood pressure in each
group was measured prior to and following AngII infusion (Fig. 2). Baseline systolic blood pressure
was comparable among each experimental group at each time-point
(P>0.05). In addition, systolic blood pressure was significantly
increased in rats at 3, 7 and 14 days following AngII treatment
compared to that of the control group (P<0.01); however, no
significant differences were observed among the AngII,
AngII+Ad-eGFP and AngII+Ad-CAT-eGFP groups. This therefore
suggested that perivascular gene transfer of catalase has no
obvious impact on systolic blood pressure.
Catalase transfection ameliorates
AngII-induced vascular hypertrophy
In order to determine whether the adventitial gene
transfer of catalase attenuated AngII-induced vascular hypertrophy,
the media thickness, media cross-sectional area (MCSA) and lumen
diameter of the CAA of rats were measured in each group;
comparisons were performed using sections taken from approximately
the same area of the CCA. As shown in Fig. 3, AngII treatment significantly
increased the media thickness, media-to-lumen ratio (M/L) and MCSA
of the CCA compared to those of the untreated control, whereas the
lumen diameter following AngII treatment was found to be decreased.
In addition, transfection of Ad-CAT-eGFP significantly attenuated
AngII-induced vascular hypertrophy (Fig. 3C); however, transfection of the
Ad-eGFP vector did not attenuate vascular hypertrophy (Fig. 3D).
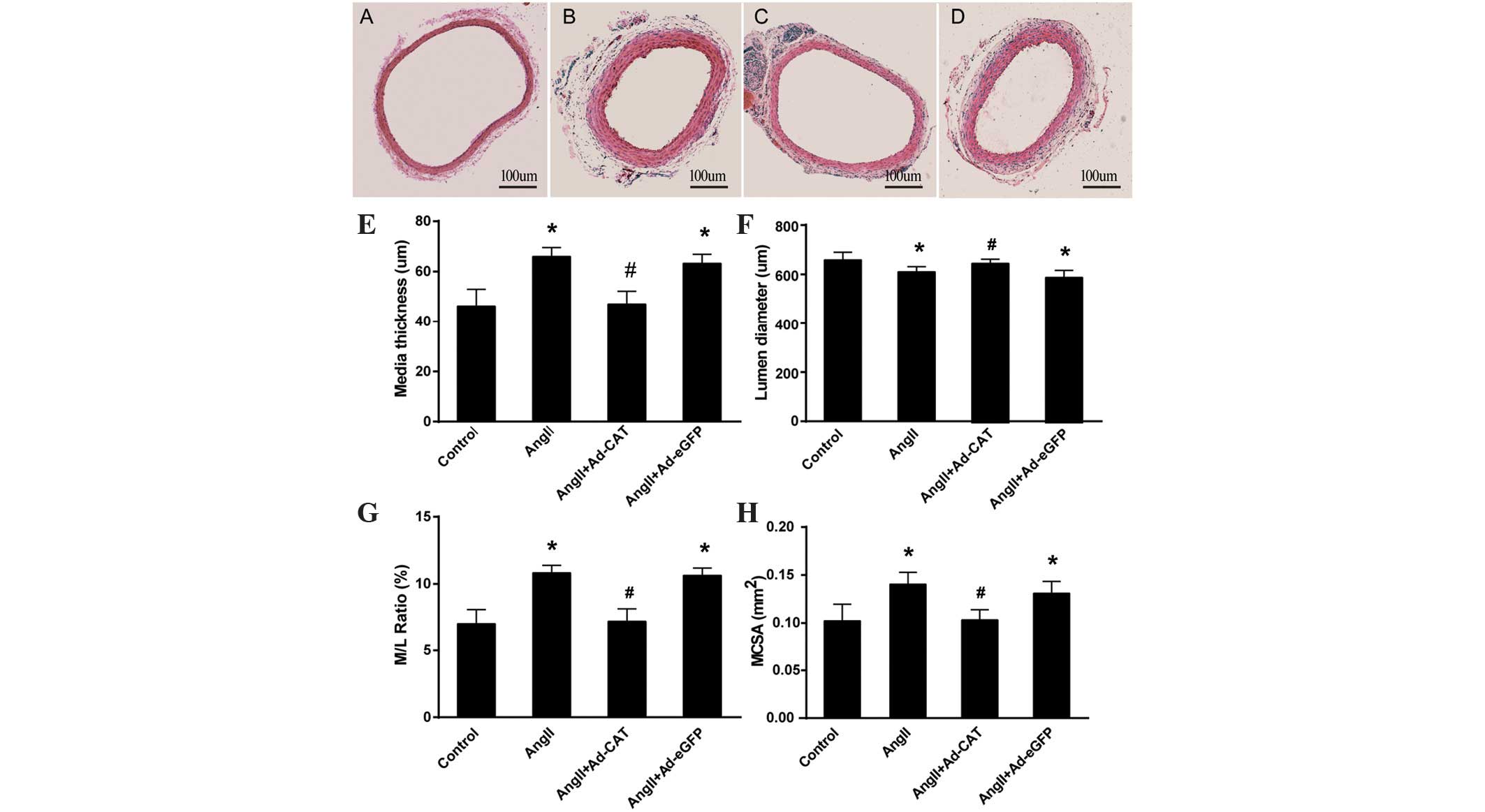 | Figure 3Ad-CAT-eGFP transfection attenuates
AngII-induced characteristics of vascular hypertrophy.
Immunohistochemistry images of section of the left common carotid
artery in each group as follows: (A) Control, (B) AngII, (C)
AngII+Ad-CAT-eGFP and (D) AngII+Ad-eGFP. Quantitative analysis of
vascular hypertrophy characteristics, including (E) media
thickness, (F) lumen diameter, (G) M/L ratio and (H) MCSA. Values
are presented as the mean ± standard deviation (n=4–6).
*P<0.05 vs. control and #P<0.05 vs.
AngII or AngII+Ad-eGFP. eGFP, enhanced green fluorescent protein;
CAT, catalase; ad, adenovirus; AngII, angiotensin II; M/L ratio,
media to lumen ratio; MCSA, media cross-sectional area. |
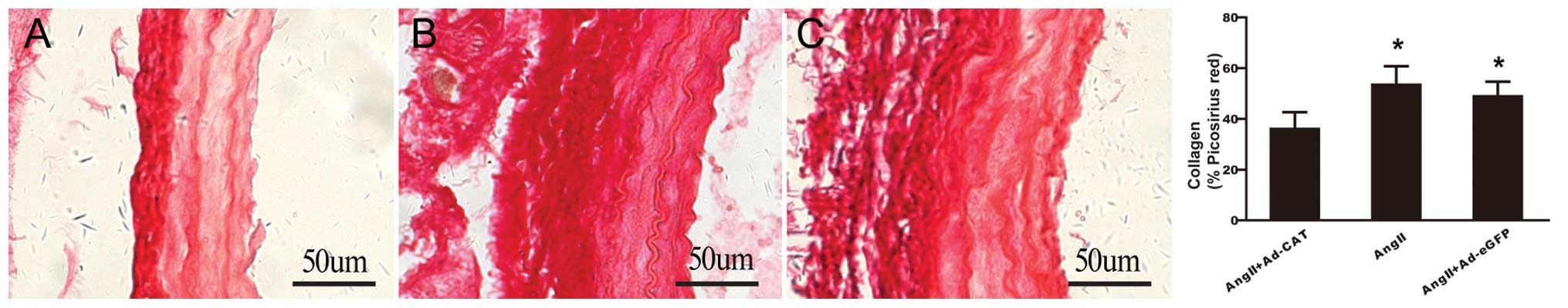
Catalase transfection reduces α-SMA
expression, ROS generation, macrophage infiltration and collagen
deposition in the CCA
In order to investigate whether catalase transfer
improved vascular remodeling, measurements were taken of the
following common markers of vascular remodeling using
immunohistochemistry: α-SMA; 4-HNE, a lipid peroxidation by-product
which reflects ROS generation; and CD68, which reflects macrophage
infiltration. As shown in Fig. 4,
adventitial transfection of Ad-CAT-eGFP significantly attenuated
the expression levels of all of the above markers compared with
those of the AngII and AngII+Ad-eGFP groups. Picrosirius red
staining revealed that catalase overexpression markedly decreased
collagen deposition compared with that in arteries treated with
AngII only and AngII+Ad-eGFP (Fig.
5). These results demonstrated that catalase transfection
significantly attenuated AngII-induced vascular remodeling.
Catalase transfection attenuates
AngII-induced vascular remodeling via inhibition of adventitial
p38MAPK phosphorylation
p38MAPK is a potential critical component of the
redox-sensitive signaling pathway, which is activated by AngII and
ROS in numerous types of tissue and cells (19,20).
In order to determine whether adventitial p38MAPK was involved in
AngII-induced vascular remodeling and whether catalase attenuated
AngII-induced vascular remodeling via inhibition of adventitial
p38MAPK, immunohistochemical and western blot analyses were used to
examine p38MAPK phosphorylation levels in vivo and ex
vivo (Fig. 6). In the in
vivo experiment, AngII infusion was found to significantly
increase phospho-p38MAPK expression in the vascular adventitia,
whereas following catalase transfection, adventitial
phospho-p38MAPK expression was significantly decreased. In the
ex vivo experiment, AFs were cultured and AngII
(1×10−7 mol/l) or PEG-catalase (1,000 U/ml) was
administrated in order to evaluate p38MAPK phosphorylation levels
under different conditions. Western blot analysis revealed that
AngII stimulation increased phospho-p38MAPK expression in AFs,
which peaked at 10 min and then decreased again in a time-dependent
manner. In addition, PEG-catalase treatment was shown to
significantly inhibit AngII-induced phospho-38MAPK expression.
These results therefore suggested that catalase overexpression
attenuated AngII-induced vascular remodeling via inhibition of
adventitial p38MAPK phosphorylation.
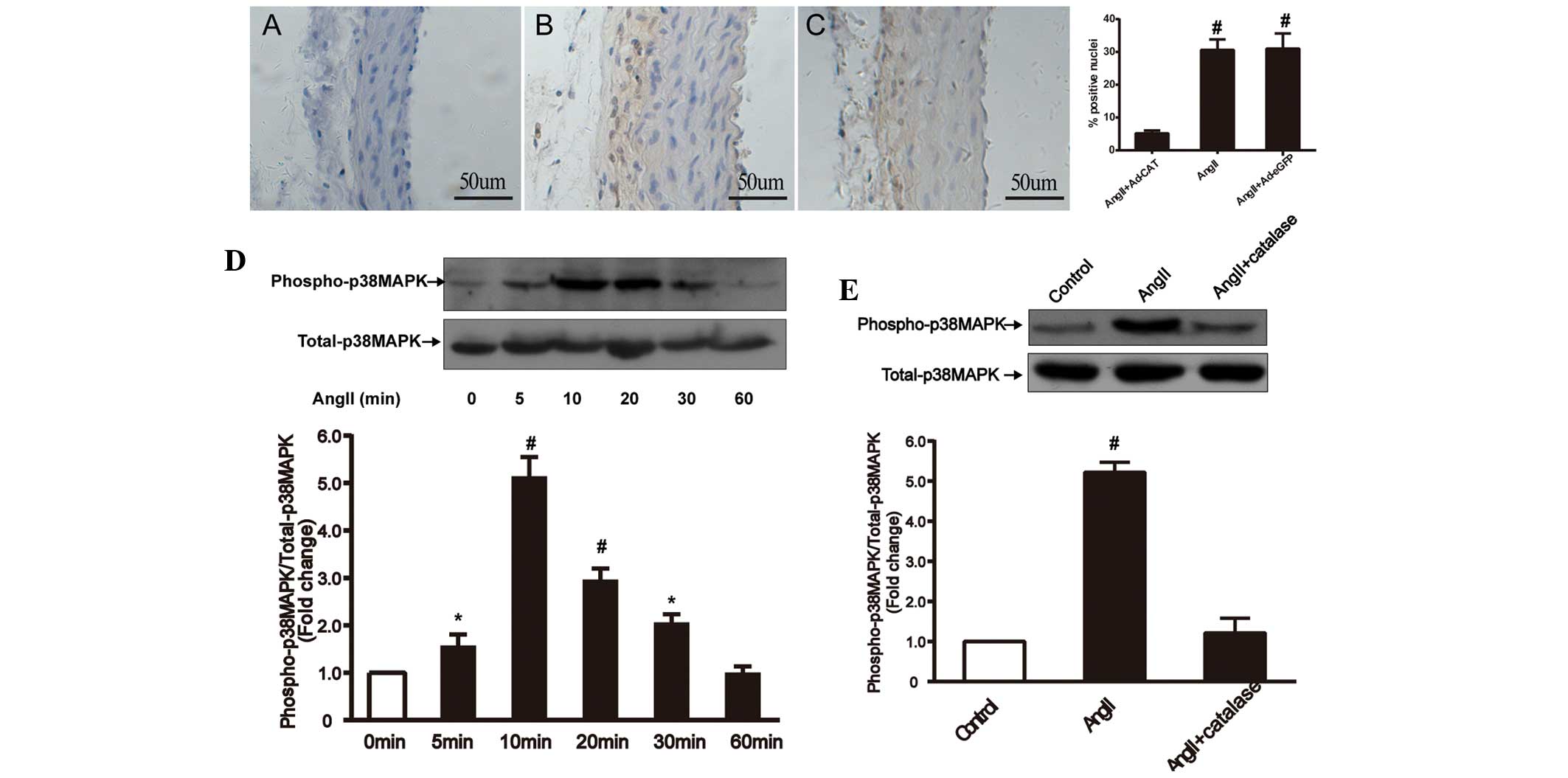 | Figure 6Catalase transfection inhibits
AngII-induced phospho-p38MAPK expression in left common carotid
arteries and adventitial fibroblasts in vivo and ex vivo,
respectively. Immunohistochemical analysis of arteries treated
with (A) AngII+Ad-CAT-eGFP, (B) AngII only and (C) AngII+Ad-eGFP
(magnification, ×200). The bar graph in the panel on the right
shows the percentage of hematoxylin-positive nuclei in each group
(n=4–6). Ex vivo western blot analysis and quantification of
(D) p38MAPK phosphorylation at 0, 5, 10, 20, 30 and 60 min
following AngII treatment (1×10−7 mol/l) and (E) p38MAPK
phosphorylation following pretreatment with polyethylene
glycol-catalase (1,000 U/ml) for 30 min prior to exposure to AngII
(1×10−7 mol/l) for 10 min in adventitial fibroblasts.
Values are presented as the mean ± standard deviation (each
experiment was conducted 6–8 times). *P<0.05 vs.
control and #P<0.01 vs. control or AngII+catalase.
AngII, angiotensin II; MAPK, mitogen-activated protein kinase; Ad,
adenovirus; eGFP, enhanced green fluorescent protein; CAT,
catalase. |
Discussion
A previous ex vivo study demonstrated that
adventitial catalase was downregulated by AngII in a time- and
dose-dependent manner (11). The
results of the present study showed that adventitial gene transfer
of catalase into carotid arteries markedly attenuated
characteristics of AngII-induced vascular remodeling, including
α-SMA expression, ROS generation, macrophage infiltration and
collagen deposition. In addition, the protective effects of
catalase were found to be independent on blood pressure
alterations. Furthermore, the present study indicated that the
attenuation of AngII-induced vascular remodeling by catalase was,
at least in part, mediated via inhibition of adventitia-derived
p38MAPK phosphorylation.
Vascular adventitia is traditionally considered to
provide support to the arterial wall; however, an increasing number
of studies have suggested that adventitia may also have an
important role in the regulation of vessel structure and function
following exposure to a pathophysiological environment (21), including injury, hypoxia and
hypertension (22,23). AFs are an important source of ROS
in a variety of pathological conditions, such as AngII-induced
hypertension (24); in addition,
studies have shown that ROS generation was significantly increased
in adventitia compared to that of the media or intima, when
stimulated by AngII (8,25). Fibroblast-derived ROS, particularly
H2O2 due to its stable and permeable nature,
serve as intercellular signaling messengers, which may induce the
development of vascular remodeling, leading to hypertension and
atherosclerosis. In addition, when stimulated with AngII, AFs
synthesize and release a variety of cytokines, including
platelet-derived growth factor (26), interleukin 6 and monocyte
chemotactic protein 1, which recruit macrophages and further
promote vascular dysfunction (27). These studies therefore indicated
the importance of adventitia in the process of AngII-induced
vascular remodeling.
Adenovirus-mediated arterial gene transfection is a
promising approach for the study of vascular biology and vascular
gene treatment. Gene transfection into vascular adventitia was
demonstrated to be effective and resulted in a minimal inflammatory
response (12). Several studies
have revealed that adventitial gene transfection into adventitial
gene targets significantly ameliorated vascular remodeling under
various conditions (8,14,28).
However, the majority of studies to date have focused on inhibiting
components of the pro-oxidant system, including the
gp91phox and p67phox subunits of NADPH oxide.
In the present study, adventitial gene transfection techniques were
used to determine the effects of adventitial application of
catalase, a crucial anti-oxide enzyme that scavenges
H2O2 in vivo, on AngII-induced
vascular remodeling. The results of the present study demonstrated
that adventitial gene transfection of catalase into carotid
arteries significantly attenuated AngII-induced vascular
remodeling. In addition, the present study provided further
evidence for the participation of vascular adventitia in remodeling
and the prominent defensive value of adventitial anti-oxide
enzymes. In concurrence with previous studies (29,30),
the present study also demonstrated that local perivascular gene
transfection targeting the vascular adventitia was a reliable
method for the treatment of vascular remodeling in animal models,
which may have the potential for use in clinical practice.
Vascular inflammatory responses have traditionally
been regarded as ‘inside-out’ processes; however, increasing
evidence has supported an ‘outside-in’ hypothesis (31), in which vascular adventitia was
considered to be the ‘first-responder’ during the early stages of
vascular disease. Several studies have reported a rapid
infiltration of leukocytes into the adventitia following vascular
injuries (9,32). The present study demonstrated that
macrophage infiltration primarily occurred in adventitia following
AngII infusion; in addition, following catalase transfection, this
AngII-induced adventitial macrophage infiltration was significantly
attenuated. This therefore suggested that catalase inhibited
AngII-induced vascular wall inflammation. Cascino et al
(33) showed that
adventitia-derived H2O2 led to vascular
relaxation dysfunction and that catalase improved AngII-induced
vascular relaxation dysfunction. In concurrence with a previous
ex vivo study (11), the
in vivo experiment in the present study confirmed that
catalase attenuated adventitial fibroblast phenotypic
differentiation and collagen deposition via reducing ROS
generation. In addition, the present study demonstrated that the
protective mechanisms of catalase proceeded, at least in part, via
inhibition of adventitial p38MAPK phosphorylation.
In conclusion, the present study demonstrated that
adventitial gene transfection of catalase significantly attenuated
AngII-induced vascular remodeling via inhibition of p38MAPK
phosphorylation, indicating that adventitia and adventitia-derived
ROS had an important role in pathological vascular remodeling.
Acknowledgements
The present study was supported by a grant from the
National Natural Science Foundation of China (no. 81170301) and a
grant from the Shanghai Natural Science Foundation (no.
11ZR1421500).
References
|
1
|
Raaz U, Toh R, Maegdefessel L, et al:
Hemodynamic regulation of reactive oxygen species: implications for
vascular diseases. Antioxid Redox Signal. 20:914–928. 2014.
View Article : Google Scholar :
|
|
2
|
Juni RP, Duckers HJ, Vanhoutte PM, Virmani
R and Moens AL: Oxidative stress and pathological changes after
coronary artery interventions. J Am Coll Cardiol. 61:1471–1481.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Paravicini TM and Touyz RM: Redox
signaling in hypertension. Cardiovasc Res. 71:247–258. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Clempus RE and Griendling KK: Reactive
oxygen species signaling in vascular smooth muscle cells. Cardiovas
Res. 71:216–225. 2006. View Article : Google Scholar
|
|
5
|
Schulz E, Gori T and Münzel T: Oxidative
stress and endothelial dysfunction in hypertension. Hypertens Res.
34:665–673. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Stenmark KR, Yeager ME, El Kasmi KC, et
al: The adventitia: essential regulator of vascular wall structure
and function. Annu Rev Physiol. 75:23–47. 2013. View Article : Google Scholar :
|
|
7
|
Sartore S, Chiavegato A, Faggin E, et al:
Contribution of adventitial fibroblasts to neointima formation and
vascular remodeling: from innocent bystander to active participant.
Circ Res. 89:1111–1121. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu J, Ormsby A, Oja-Tebbe N and Pagano
PJ: Gene transfer of NAD (P) H oxidase inhibitor to the vascular
adventitia attenuates medial smooth muscle hypertrophy. Circ Res.
95:587–594. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tieu BC, Ju X, Lee C, et al: Aortic
adventitial fibroblasts participate in angiotensin-induced vascular
wall inflammation and remodeling. J Vasc Res. 48:261–272. 2011.
View Article : Google Scholar
|
|
10
|
Pagano PJ, Chanock SJ, Siwik DA, Colucci
WS and Clark JK: Angiotensin II induces p67phox mRNA expression and
NADPH oxidase superoxide generation in rabbit aortic adventitial
fibroblasts. Hypertension. 32:331–337. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yang W, Zhang J, Wang H, et al:
Angiotensin II downregulates catalase expression and activity in
vascular adventitial fibroblasts through an AT1R/ERK1/2-dependent
pathway. Mol Cell Biochem. 358:21–29. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Schneider DB, Sassani AB, Vassalli G,
Driscoll RM and Dichek DA: Adventitial delivery minimizes the
proinflammatory effects of adenoviral vectors. J Vasc Surg.
29:543–550. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Laitinen M and Yla-Herttuala S:
Adventitial gene transfer to arterial wall. Pharmacol Res.
37:251–254. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dourron HM, Jacobson GM, Park JL, et al:
Perivascular gene transfer of NADPH oxidase inhibitor suppresses
angioplasty-induced neointimal proliferation of rat carotid artery.
Am J Physiol Heart Circ Physiol. 288:H946–H953. 2005. View Article : Google Scholar
|
|
15
|
Ozumi K, Tasaki H, Takatsu H, et al:
Extracellular superoxide dismutase overexpression reduces
cuff-induced arterial neointimal formation. Atherosclerosis.
181:55–62. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tollefson AE, Kuppuswamy M, Shashkova EV,
Doronin K and Wold WS: Preparation and titration of CsCl-banded
adenovirus stocks. Methods Mol Med. 130:223–235. 2007.PubMed/NCBI
|
|
17
|
Ducharme A, Frantz S, Aikawa M, et al:
Targeted deletion of matrix metalloproteinase-9 attenuates left
ventricular enlargement and collagen accumulation after
experimental myocardial infarction. J Clin Invest. 106:55–62. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Siow RC, Mallawaarachchi CM and Weissberg
PL: Migration of adventitial myofibroblasts following vascular
balloon injury: insights from in vivo gene transfer to rat carotid
arteries. Cardiovasc Res. 59:212–221. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chan SH, Hsu KS, Huang CC, Wang LL, Ou CC
and Chan JY: NADPH oxidase-derived superoxide anion mediates
angiotensin II-induced pressor effect via activation of p38
mitogen-activated protein kinase in the rostral ventrolateral
medulla. Circ Res. 97:772–780. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ushio-Fukai M, Alexander RW, Akers M and
Griendling KK: P38 mitogen-activated protein kinase is a critical
component of the redox-sensitive signaling pathways activated by
angiotensin II. Role in vascular smooth muscle cell hypertrophy. J
Biol Chem. 273:15022–15029. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Di Wang H, Rätsep MT, Chapman A and Boyd
R: Adventitial fibroblasts in vascular structure and function: the
role of oxidative stress and beyond. Can J Physiol and Pharmacol.
88:177–186. 2010. View
Article : Google Scholar
|
|
22
|
Havelka GE and Kibbe MR: The vascular
adventitia: its role in the arterial injury response. Vasc
Endovascular Surg. 45:381–390. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shi Y, O’Brien JE, Fard A, Mannion JD,
Wang D and Zalewski A: Adventitial myofibroblasts contribute to
neointimal formation in injured porcine coronary arteries.
Circulation. 94:1655–1664. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Haurani MJ and Pagano PJ: Adventitial
fibroblast reactive oxygen species as autacrine and paracrine
mediators of remodeling: bellwether for vascular disease?
Cardiovasc Res. 75:679–689. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Di Wang H, Hope S, Du Y, et al: Paracrine
role of adventitial superoxide anion in mediating spontaneous tone
of the isolated rat aorta in angiotensin II-induced hypertension.
Hypertension. 33:1225–1232. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bishop JE and Lindahl G: Regulation of
cardiovascular collagen synthesis by mechanical load. Cardiovasc
Res. 42:27–44. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Al Ghouleh I, Khoo NK, Knaus UG, et al:
Oxidases and peroxidases in cardiovascular and lung disease: new
concepts in reactive oxygen species signaling. Free Radic Biol Med.
51:1271–1288. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Weaver M, Liu J, Pimentel D, et al:
Adventitial delivery of dominant-negative p67phox attenuates
neointimal hyperplasia of the rat carotid artery. Am J Physiol
Heart Circ Physiol. 290:H1933–H1941. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mallawaarachchi CM, Weissberg PL and Siow
RC: Smad7 gene transfer attenuates adventitial cell migration and
vascular remodeling after balloon injury. Arterioscler Thromb Vasc
Biol. 25:1383–1387. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Siow RC and Churchman AT: Adventitial
growth factor signalling and vascular remodelling: potential of
perivascular gene transfer from the outside-in. Cardiovasc Res.
75:659–668. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Maiellaro K and Taylor WR: The role of the
adventitia in vascular inflammation. Cardiovasc Res. 75:640–648.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Okamoto E, Couse T, De Leon H, et al:
Perivascular inflammation after balloon angioplasty of porcine
coronary arteries. Circulation. 104:2228–2235. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Cascino T, Csanyi G, Al Ghouleh I, et al:
Adventitia-derived hydrogen peroxide impairs relaxation of the rat
carotid artery via smooth muscle cell p38 mitogen-activated protein
kinase. Antioxid Redox Signal. 15:1507–1515. 2011. View Article : Google Scholar :
|