Introduction
Endothelial dysfunction has similar early
pathological alterations and risk factors to those in vascular
diseases, such as cardiovascular disease and diabetic vascular
complications (1).
Endothelium-dependent damage arises from metabolic abnormalities in
the glucose metabolism that lead to vascular dysfunction (2). Carbonyl compound-induced stress and
pro-inflammatory responses lead to the formation of advanced
glycation end products (AGEs) (3).
Methylglyoxal (MGO), a key precursor for AGEs and a reactive
dicarbonyl compound, is suggested to be an intermediate derived
from the metabolism of glucose (4). Numerous previous studies support a
role for MGO in triggering two distinct signaling cascades leading
to oxidative damage and pro-inflammatory responses in HUVECs
(5,6). The trapping of dicarbonyl compounds
like MGO represents an effective strategy for attenuating carbonyl
stress-induced endothelial injury (3).
Curcumin (Cur) is the most active component of the
curcuminoids extracted from Curcuma longa L. and has been
demonstrated to protect against AGE-induced cellular inflammatory
responses and oxidative stress in vascular complications (7,8).
Increasing evidence has suggested that Cur possesses a potential
protective effect against MGO-induced endothelial dysfunction via
scavenging reactive oxygen species (ROS) and attenuating the levels
of inflammatory mediators (9).
Previous studies have indicated that Cur may prevent MGO-induced
endothelial dysfunction by directly trapping MGO to form a
curcumin-MGO adduct at the electron-dense carbon atom (C10) between
the two keto carbon groups (10).
However, Cur-MGO adducts have not been studied in depth at present
and it remains to be investigated whether Cur-MGO adducts are able
to attenuate cytotoxicity in HUVECs.
In the present study, the trapping capacity of Cur
was investigated through examining the optimal reaction parameters
and analyzing the Cur-MGO adducts by high-performance liquid
chromatography-diode-array detection (HPLC-DAD) coupled with liquid
chromatography-electrospray ionization-tandem mass spectrometry
(LC-ESI-MS/MS). Furthermore, the differences in the levels of
oxidative damage and pro-inflammatory cytokines as a result of the
formation of AGEs in the presence or absence of Cur were compared
in HUVECs to investigate the protective mechanisms of Cur on
endothelial dysfunction.
Materials and methods
Chemicals and materials
MGO, 40% aqueous solution), aminoguanidine
hydrochloride (AG; purity ≥98%),
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT),
1,2-diaminobenzene (DB) and 2-methylquinoxaline were purchased from
Sigma-Aldrich (St. Louis, MO, USA). Human serum albumin (HSA) was
obtained from Amresco LLC (Solon, OH, USA). Human AGEs ELISA kit,
SABC kit, transforming growth factor-β1 (TGF-β1; cat. no. sc-1672;
1:200) and intercellular adhesion molecule-1 (ICAM-1; cat. no.
sc-1506; 1:200) antibodies were obtained from Santa Cruz
Biotechnology, Inc., (Dallas, TX, USA). The ROS kit was purchased
from Beyotime Institute of Biotechnology (Nantong, China). Curcumin
(purity ≥98%) was obtained from the National Institute for the
Control of Pharmaceutical and Biological Products (Beijing, China).
HPLC-grade methanol was purchased from Tedia (Fairfield, OH, USA).
HPLC-grade water was prepared using a Millipore Milli-Q
purification system (EMD Millipore, Billerica, MA, USA). Other
reagents were analytical grade and from Nanjing Chemical Reagent
Co., Ltd. (Nanjing, China).
Cell culture
Human umbilical vein endothelial cells (HUVECs),
were selected to model endothelial disease in vitro and were
purchased from the American Type Culture Collection (Manassas, VA,
USA). HUVECs were cultured in low-glucose Dulbecco's modified
Eagle's medium supplemented with 10% fetal bovine serum (FBS; Gibco
Life Technologies, Carlsbad, CA, USA), 80 U/ml penicillin and 80
U/ml streptomycin. The cells were maintained in a humidified
incubator at 37°C containing 5% CO2. The culture medium
was replaced every 2 days. HUVECs used in the study were passaged
3–5 times prior to use.
Reaction of Cur or DB with MGO
DB (1 mM) and MGO (1 mM) were incubated in 2 ml
phosphate-buffered saline (PBS; 0.2 M, pH 7.4; Nanjing KeyGen
Biotech Co., Ltd., Nanjing, China). The mixtures were agitated at
40 rpm at 37°C for 30, 90, 240 and 720 min. A PBS solution was used
as the blank control. Cur (1 mM) and MGO (1 mM) were incubated in
0.2 M PBS. The mixtures were agitated at 40 rpm at 37°C for 720
min. MGO alone (1 mM) or Cur (1 mM) was used as the control.
Following the reaction, the samples were dried using high purity
nitrogen overnight. The residue was redissolved in 1 ml HPLC-grade
methanol and vortexed for 30 sec. The sample solutions were
filtered through a 0.45 µm microporous membrane prior to
HPLC analysis.
Kinetic study of the trapping of MGO by
Cur
In order to optimize the incubation ratio of Cur and
MGO, Cur (0.2, 0.33, 1, 3 and 5 mM) was incubated with 1 mM MGO in
3 ml PBS (0.2 M, pH 7.4) at 37°C and agitated at 40 rpm.
Subsequently, 200 µl reaction mixture was collected at each
time point, 1 µl acetic acid (Nanjing Chemical Reagent Co.,
Ltd.) added to stop the reaction and 100 mM DB (14.6 mmol) added to
react with the remaining MGO, as previously described (11).
HPLC analysis of the reaction mixture of
Cur or DB with MGO
HPLC-DAD was conducted to analyze the reaction
mixture of Cur or DB with MGO. This analysis was conducted on an
Agilent 1200 system (Agilent Technologies, Inc., Santa Clara, CA,
USA) which was equipped with a quaternary pump, DAD detector, an
autosampler and Agilent ChemStation B.0401 software. The analysis
conditions were as follows: An Alltima C18 (4.6×150 mm, 5
µm) column was used for this analysis; the flow rate was set
at 1.0 ml/min and the column temperature maintained at 30°C. For
the reaction of Cur and MGO, the mobile phase consisted of methanol
(eluent A) and water (eluent B) with a gradient program of 0–45
min, 2–80% A, conducted for component separation with a linear
gradient elution, and the determination wavelength set at 425 nm.
For the reaction of DB and MGO, methanol (eluent A) and water
(eluent B), (0–9 min, 5–50% A) was used for the mobile phase with a
wavelength of 280 nm set. In total, a 20 µl sample was
injected into the HPLC system.
LC-ESI-MS/MS analysis
An Agilent 1200 HPLC system combined with an
LCQ-Fleet Ion Trap Mass Spectrometer (Thermo Fisher Scientific,
Waltham, MA, USA) was used to identify the chemical structure of
the Cur-MGO adducts. The detailed conditions and parameters were as
follows: The ionization was achieved using the electrospray in
positive mode; helium was used as the collision gas and nitrogen
(N2) as the nebulizing gas; spray voltage was set at 4.5
kV and capillary voltage at 5 V; the capillary temperature was kept
at 300°C; N2 was selected as the sheath gas and its
pressure was set at 90 arbitrary units. In addition, the isolation
width of the precursor ions was set at 1.5 Th. The mass range of
compounds scanned was from 50–1,200 m/z.
MGO derived-AGEs preparation in the
presence or absence of Cur
MGO-AGEs were prepared with a modified reaction
quantity of HSA and MGO as previously described (12). Briefly, 4 mg HSA and 50 mM MGO were
incubated in 1 ml PBS (0.2 M, pH 7.4) in the presence or absence of
Cur (10−7, 10−6 and 10−5 M) under
sterile conditions and maintained in 95% air/5% CO2 at
37°C. For the kinetic study, the mixture was agitated for 0, 4, 12,
24, 48, 72, 96, 120, 144 and 168 h. Similarly, HSA in the absence
of MGO was incubated in the same conditions. AG (10−6 M)
was selected as the positive control. The fluorescent intensity of
MGO-AGEs was determined using a fluorescent microplate reader with
the excitation/emission wavelength set at 370/440 nm (Gemini EM™;
Molecular Devices, LLC, Sunnyvale, CA, USA) (13). In addition, the level of MGO-AGEs
was measured using an AGEs ELISA kit and a SpectraMax 190
microplate reader at 450 nm (Molecular Devices, LLC). The obtained
MGO-AGEs were stored at 4°C for further experimental use. MGO-AGEs
containing 1 mM MGO were used for the cell experiments.
ELISA assay for MGO-AGEs
In order to measure the inhibitory effect of Cur on
the formation of MGO-AGEs, an Human AGE ELISA kit (cat. no.
sc-1609; Santa Cruz Biotechnology, Inc.) was used to measure the
level of MGO-AGEs, according to the manufacturer's instructions.
The optical density (OD) value of samples was measured using an
MK-3 microplate reader (Thermo Fisher Scientific, Inc.) at 450
nm.
Dihydroethidium (DHE) staining and flow
cytometry for the level of ROS
MGO-AGEs may induce intracellular ROS generation and
lead to endothelial injury. Therefore, in the current study, DHE
(Nanjing KeyGen Biotech Co., Ltd.) staining was conducted to
measure the level of ROS as previously described (14). Cells were incubated with 5
µM DHE at 37°C for 30 min according to the manufacturer's
instructions. The free DHE molecules were then removed by washing
with PBS. Fluorescent images of ROS were captured by fluorescence
microscopy (Olympus IX73; Olympus Corporation, Tokyo, Japan). In
addition, the level of intracellular ROS was measured using flow
cytometery (FACSCalibur; BD Biosciences, Franklin Lakes, NJ, USA)
at an excitation/emission wavelength of 488/530 nm (15). The fluorescence intensity of DHE
was analyzed in 10,000 cells using CellQuest software (BD
Biosciences).
MTT assay for cell viability
To investigate the cytotoxicity of the Cur-MGO
adducts, an MTT assay was used to measure the cell viability of
HUVECs. Cells (1×105 cells/well) were seeded into
96-well plates and incubated for 24 h. Once 80% confluent, cells
were stimulated with Cur-MGO adducts for 48 h. Subsequently MTT (5
mg/ml, 10 µl) was added in FBS-free media to each well to
replace the original media and then incubated for 4 h at 37°C in an
incubator. Following this, the supernatant was removed and the
formazan crystals were dissolved in 100 µl dimethyl
sulfoxide (Nanjing Chemical Reagent Co., Ltd.). Following agitation
for 10 min, the OD of each sample was measured using the microplate
reader at 550 nm. Cell viability was calculated according to the
relative difference in OD value.
Immunocytochemistry for ICA M-1 and
TGF-β1
Immunocytochemistry was conducted to investigate the
effect of Cur-MGO adducts on the expression levels of ICAM-1 and
TGF-β1 in HUVECs (16). Cells were
grown on glass coverslips in 24-well plates. Following 48 h
stimulation with Cur-MGO adducts, HUVECs were fixed in fresh 4%
formaldehyde (Nanjing Chemical Reagent Co., Ltd.) and washed three
times in PBS. Cells were treated with 3% H2O2
for 10 min to block endogenous peroxidase activity. Subsequently,
cells were blocked with 5% bovine serum albumin (BSA; Roche
Diagnostics, Basel, Switzerland) for 30 min and then incubated with
ICAM-1 (1:500) or TGF-β1 (1:500) antibodies for 120 min at 37°C.
Cells were then incubated with biotin-conjugated secondary
antibodies ICAM1 and TGF-β1 for 120 min at room temperature, and
the staining was visualized using 3,3-diaminobenzidine (Nanjing
KeyGen Biotech Co., Ltd.). The cells were counterstained with
hematoxylin (Nanjing KeyGen Biotech Co., Ltd.) and images were
captured using a microscope (Olympus IX71; Olympus Corporation) and
Image-Pro Plus 6.0 software (Media Cybernetics, Inc., Rockville,
MD, USA). As a control, the primary antibodies were replaced with
PBS.
Western blot analysis for ICAM-1 and
TGF-β1 levels
Following treatment with Cur-MGO adducts, protein
was extracted from HUVECs using lysis buffer containing 1% Triton
X-100 (Nanjing Chemical Reagent Co., Ltd.), 150 mM NaCl (Nanjing
Chemical Reagent Co., Ltd.), 1 mM EDTA (Nanjing KeyGen Biotech Co.,
Ltd.), 20 mM Tris-HCl (Nanjing KeyGen Biotech Co., Ltd.), 5
µg/ml pepstatin A (Nanjing KeyGen Biotech Co., Ltd.), 2 mM
diisopropyl fluorophosphate (Nanjing Chemical Reagent Co., Ltd.)
and 1 mM phenylmethylsulfonyl fluoride (Nanjing Chemical Reagent
Co., Ltd.). Equal amounts of protein (50 µg) were loaded and
then separated using 10% SDS-PAGE (Nanjing KeyGen Biotech Co.,
Ltd.). Subsequently, the proteins were transferred to
polyvinylidene difluoride membranes (EMD Millipore). Following
blocking with 5% BSA in Tris-buffered saline containing 0.1%
Tween-20 (Nanjing KeyGen Biotech Co., Ltd.) for 1 h, the membranes
were incubated with primary antibodies against ICAM-1 (1:500) or
TGF-β1 (1:500) at 37°C for 2 h. Following three washes with PBS,
the membranes were incubated with horseradish peroxide-conjugated
ICAM1 and TGF-β1 secondary antibodies (1:1,000) for 2 h at 37°C.
Bands were visualized using enhanced chemiluminescence (Nanjing
KeyGen Biotech Co., Ltd.) and imaged using a minicamera (Olympus
IX71). The experiment was repeated a minimum of three times.
Statistical analysis
All data were taken from three individual
experiments and presented as the mean ± standard deviation. SPSS
software, version 16.0 (SPSS, Inc., Chicago, IL, USA) was used to
conduct statistical analyses using a one-way analysis of variance
and Tukey's test. P<0.05 was considered to indicate a
statistical significance difference.
Results
The reliability of the reaction
system
To ensure the reliability of the reaction system, DB
was reacted with MGO. The aldehyde groups of MGO and the amino
groups of DB form −CH=N- through a Schiff base reaction, forming
2-methylquinoxaline (Fig. 1A).
Following the reaction, the 2-methylquinoxaline generated by the
reaction system was analyzed by HPLC-DAD at 280 nm. As presented in
Fig. 1B, the reaction product
matched the 2-methylquinoxaline standard according to the UV
spectra and retention times. In order to investigate the optimal
reaction time, 30, 90, 240 and 720 min were selected as reaction
time points. The HPLC-DAD indicated that the level of
2-methylquinoxaline in the reaction system increased gradually and
that the content reached the maximum level at 720 min (content at
later time points was measured in preliminary experiments).
Therefore, 720 min was selected as the optimal reaction time. The
results indicated that the reaction system was suitable for
trapping dicarbonyl compounds.
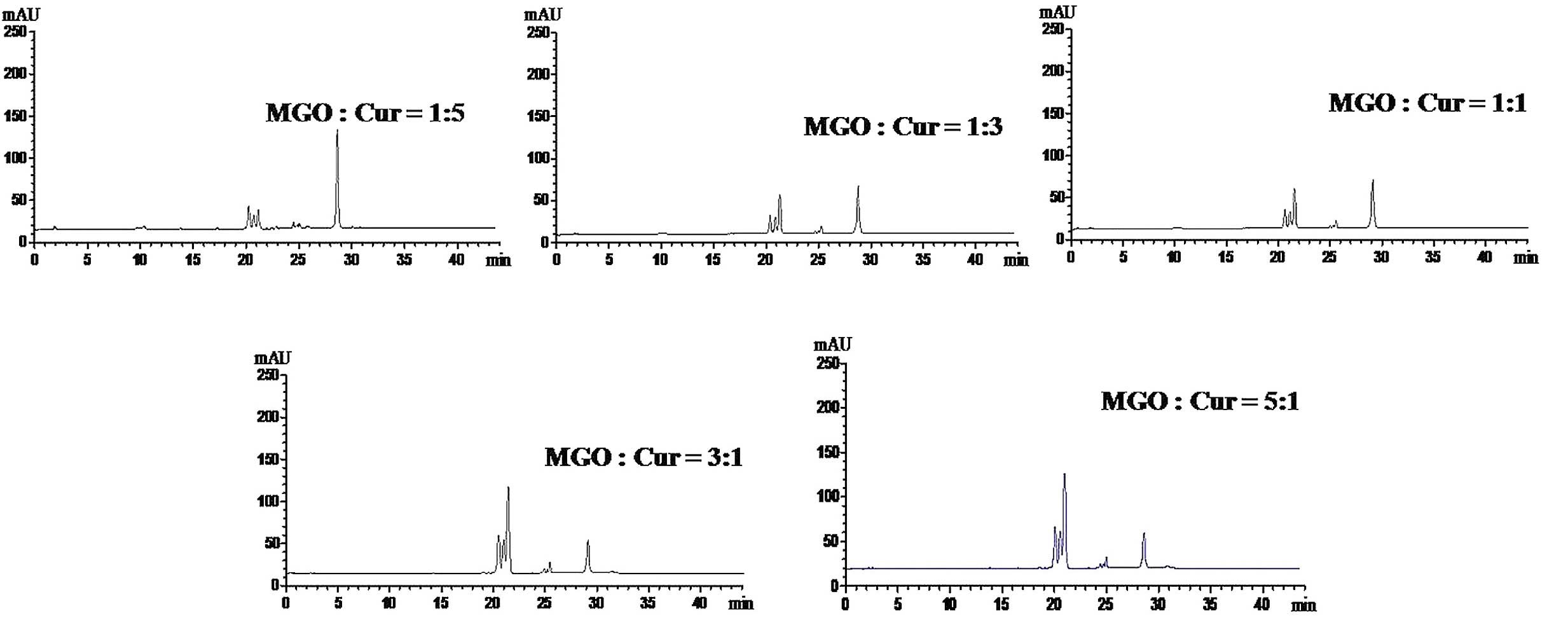
The optimal reaction ratio for the
trapping of MGO by Cur
In order to identify the optimal reaction
conditions, the ratio of MGO:Cur (1:5, 1:3, 1:1, 3:1 and 5:1) was
investigated. As presented in Fig.
2, the content of Cur in the reaction system was reduced and
three novel peaks were increased with the increasing MGO:Cur ratio.
When the ratio of MGO:Cur was 1:1, the content of Cur was not
further reduced while the formation of MGO-Cur adducts was not
increased. This indicated that the 1:1 ratio of MGO:Cur resulted in
a complete chemical reaction.
Identification of Cur-MGO adducts by
HPLC/electrospray ionization tandem mass spectrometry
(HPLC-ESI-MS/MS)
The ability of Cur to capture the carbonyl compound
MGO was investigated in the reaction system. MGO (1 mM) and Cur (1
mM) were reacted together for 720 min. As presented in Fig. 3A, compared with Cur alone or MGO
alone, three new peaks were formed on the chromatogram along with a
reduction in the Cur peak. The retention time of Cur was 29.16 min
while the three Cur-MGO adducts were 20.17, 20.94 and 21.37 min.
These results demonstrated that Cur is able to trap MGO to form
three Cur-MGO adducts.
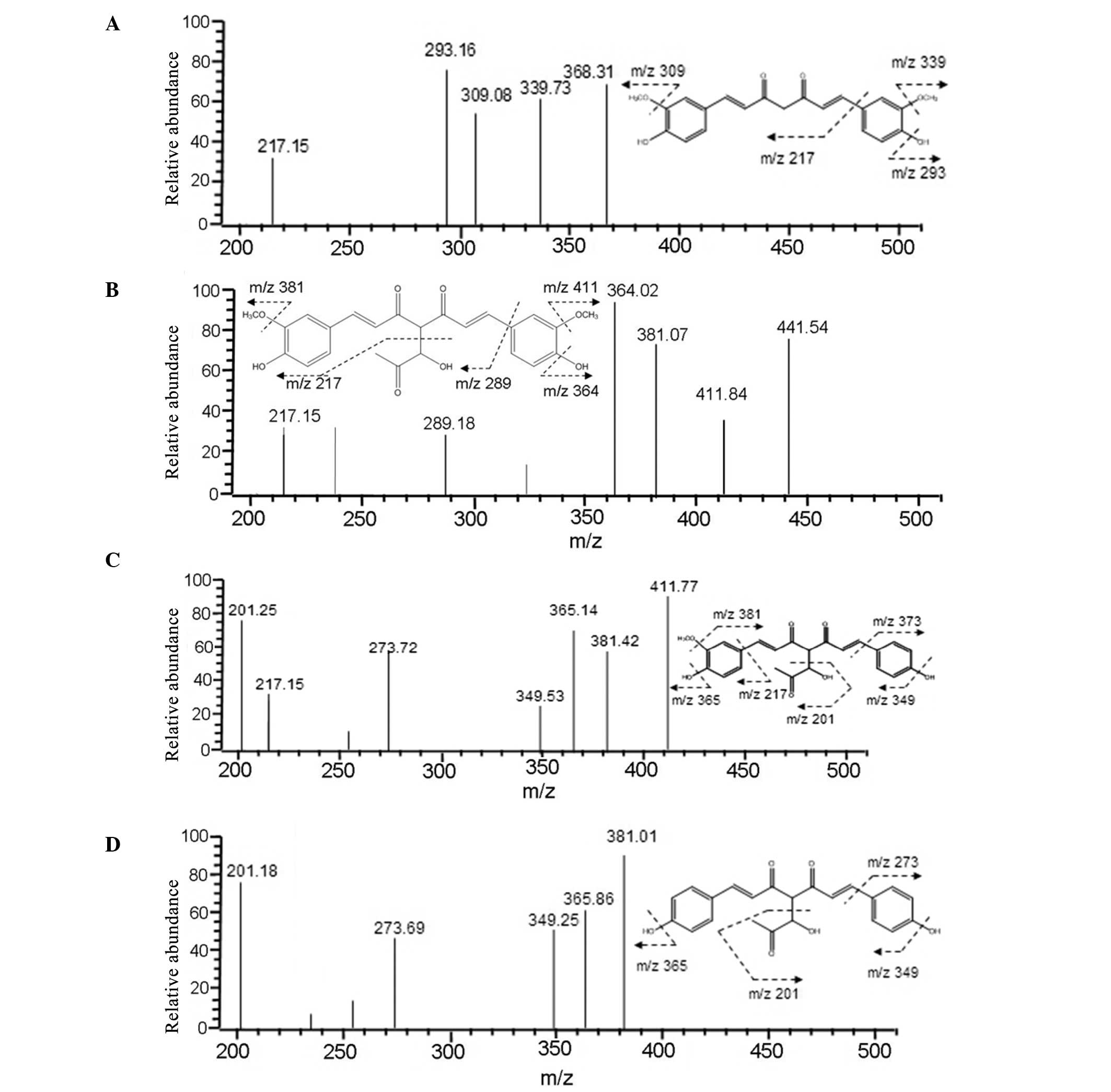
To identify the Cur-MGO adducts, HPLC-ESI-MS/MS was
conducted to analyze the molecular ion composition. As presented in
Fig. 4, Cur-MGO adducts exhibited
similar retention times of pure Cur and the molecular ion mass to
charge ratios (m/z) of 339, 309, 293 and 217 [(M+H)−]. Peak 1 was
identified as Cur-MGO adduct 1 according to the fragment ion m/z of
411, 381, 364, 289 and 217 [(M+H)−]. Adduct 1 is the same as a
previously reported adduct, with a β-hydrogen shift to the double
bond of one of the diketones of deprotonated Cur (10). In addition, the current study
identified two novel Cur-MGO adducts, adducts 2 and 3. Based on the
fragment ion m/z of 411, 381, 365, 349, 273 and 217, and 381, 365,
349, 273 and 201, adducts 2 and 3 were suggested to be present due
to the observation of the loss of one or two −OCH3
groups, respectively. The formation of these adducts may be
associated with the structural instability of Cur and Cur-MGO
adducts. The potential formation pathway under physiological
conditions of Cur-MGO adducts is presented in Fig. 3B (pH 7.4, 37°C).
Inhibitory effect of Cur on the formation
of AGEs
In order to further investigate the inhibitory
effect of Cur on the formation of AGEs through the trapping of
dicarbonyl compounds, Cur was incubated with MGO and HSA and the
reaction kinetics were measured. As presented in Fig. 5A, there was a reduction in the
formation of MGO-AGEs with increasing concentrations of Cur
(10−7–10−5 M). Following incubation for 24 h,
the relative fluorescence units of the samples in the presence or
absence of Cur were stable. Furthermore, AG, the positive control
(10−6 M), demonstrated a significant inhibition of the
formation of MGO-AGEs. In addition, the results of the ELISA
demonstrated that Cur was able to significantly inhibit the
formation of MGO-AGEs in a concentration-dependent manner, compared
with the MGO and MGO + HSA groups (P<0.01; Fig. 5B and C). These results indicate
that Cur inhibits the formation of MGO-AGEs in the reaction
system.
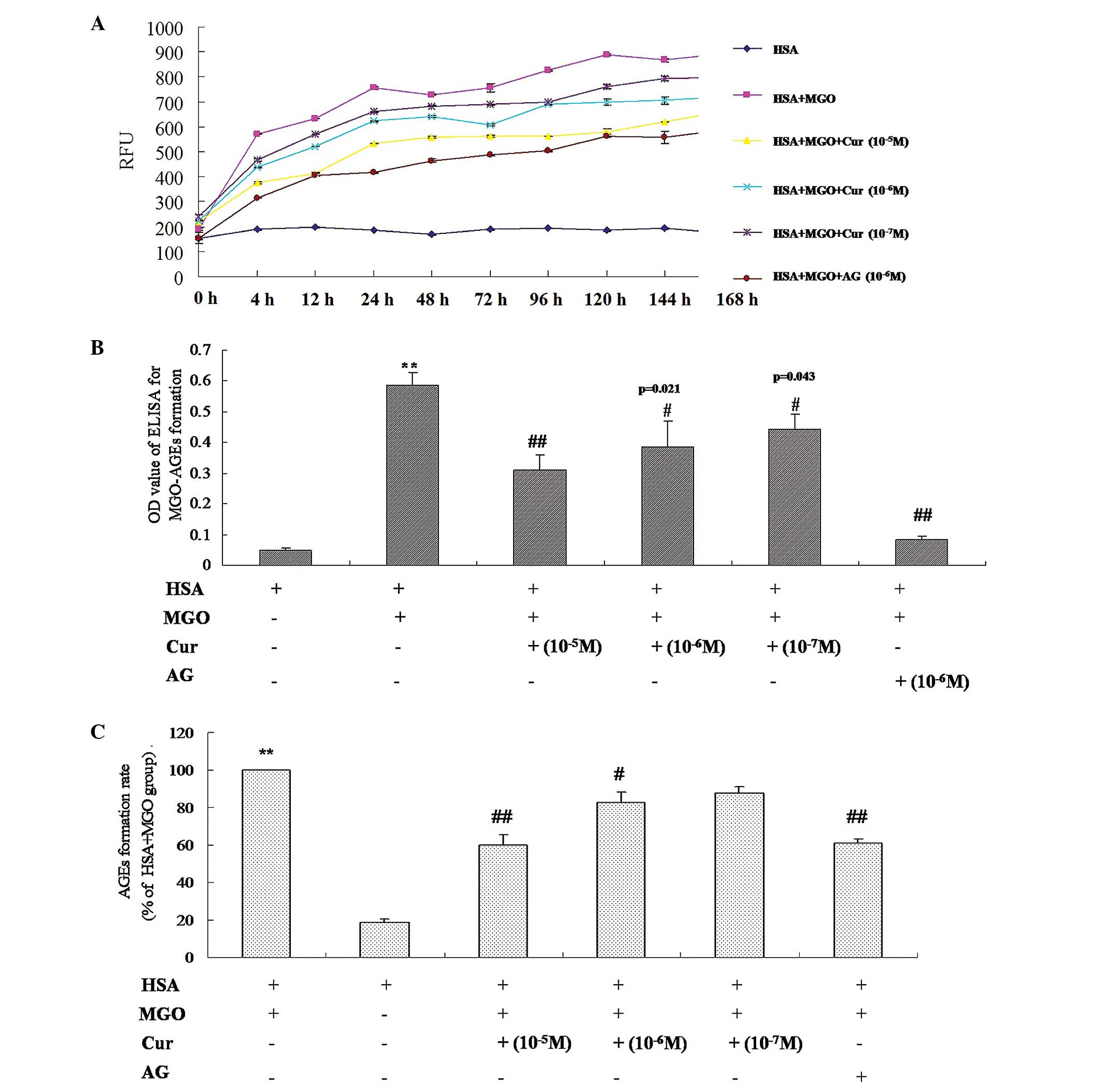 | Figure 5Inhibitory effect of Cur on the
formation of AGEs in the MGO and HSA reaction system. Cur
(10−7, 10−6 and 10−5 M) was
co-incubated with MGO (50 mM) and HSA (4 mg) in 1 ml
phosphate-buffered saline solution for 0, 4, 12, 24, 48, 74, 96,
120, 144 and 168 h. (A) The fluorescent intensity of MGO-AGEs was
determined at the excitation/emission wavelengths of 370/440 nm.
(B) The level of MGO-AGEs was measured using ELISA and (C) the
relative formation rate of MGO-AGEs was calculated. The formation
rate of MGO-AGEs was calculated according to the following formula:
The formation rate= [(OD MGO & HSA - OD MGO & HSA &
Cur)/OD MGO & HSA] × 100. AG (10−6 M) was used as
the positive control. The data are presented as the mean ± standard
deviation (n=6). **P<0.01, vs. HSA or vs. MGO + MGO;
#P<0.01, ##P<0.01, vs. MGO + HSA. Cur,
curcumin; AGEs, advanced glycation end products; MGO, methylgloxal;
HSA, human serum albumin; OD, optical density; AG, aminoguanidine
hydrochloride; RFU, relative fluorescence units. |
Attenuation of ROS generation by Cur-MGO
adducts in HUVECs
The level of ROS in HUVECs was measured using a ROS
kit and flow cytometry. As presented in Fig. 6A, the production of ROS was
significantly induced by MGO or MGO + HSA (P<0.01). However, the
generation of ROS was inhibited following incubation with Cur-MGO
adducts in a concentration-dependent manner. Quantification of the
flow cytometry results demonstrated that the Cur-MGO adducts
resulted in a significant reduction in the generation of ROS
(P<0.05 or P<0.01) (Fig. 6B and
C). These data demonstrate that Cur may attenuate MGO-induced
endothelial dysfunction via the trapping of MGO.
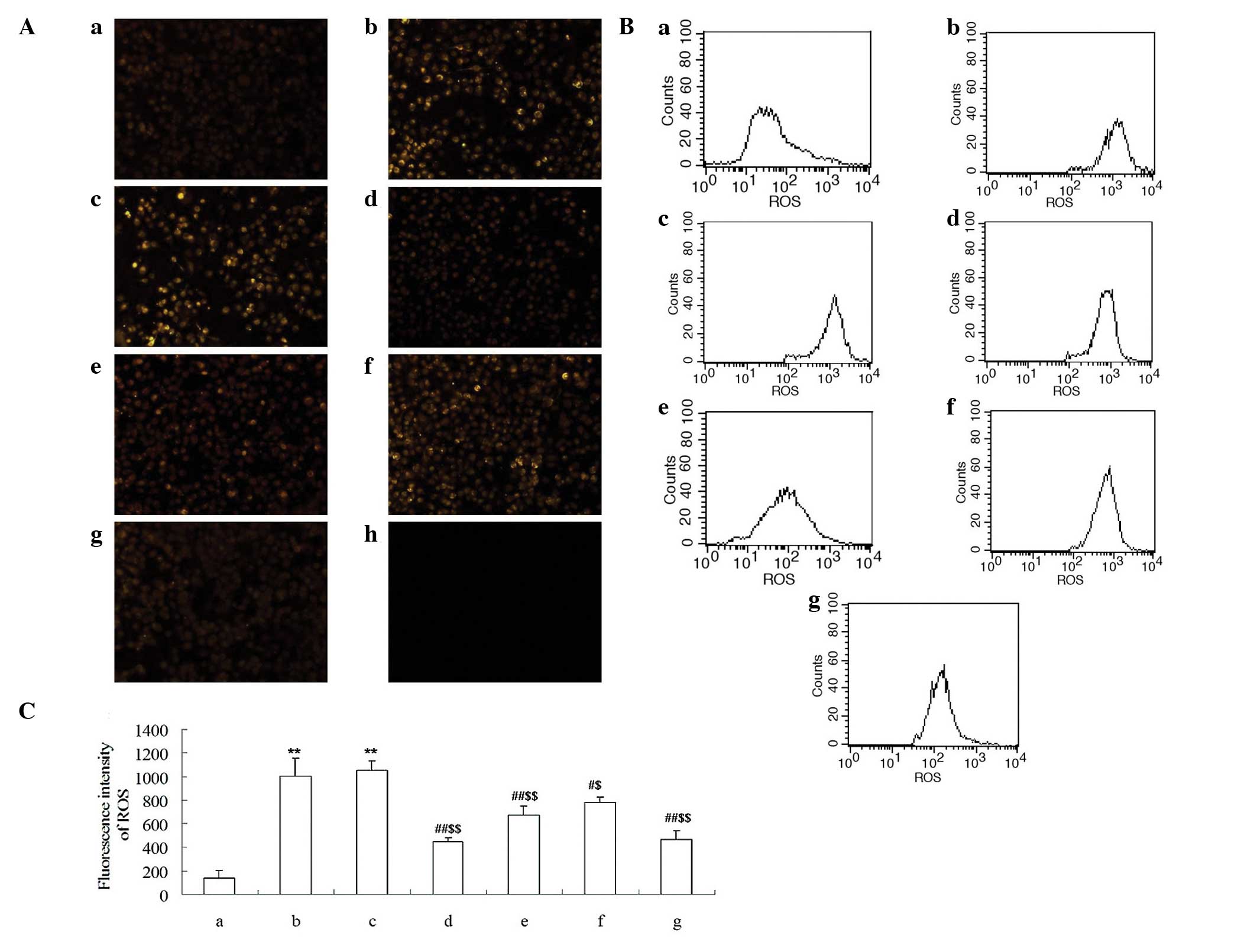 | Figure 6Effect of Cur-MGO adducts on ROS
generation in HUVECs. (A) Intracellular ROS was stained with 5
µM DHE and imaged using fluorescence microscopy. (B) The
level of intracellular ROS was measured using flow cytometry and
(C) the fluorescence intensity was calculated. a, HSA; b, MGO; c,
MGO + HSA; d, MGO + HSA + Cur (10−7 M); e, MGO + HSA +
Cur (10−6 M); f, MGO + HSA + Cur (10−5 M); g,
MGO + HSA + AG (10−6 M); h, negative control. The
experiments were repeated three times and the data are presented as
the mean ± standard deviation (n=3). **P<0.01, vs.
HSA; #P<0.05, ##P<0.01, vs. MGO;
$$P<0.01, vs. HSA + MGO. Cur, curcumin; MGO,
methylgloxal; ROS, reactive oxygen species; HUVECs, human umbilical
vein endothelial cells; DHE, dihydroethidium; HSA, human serum
albumin; AG, aminoguanidine hydrochloride. |
Reduced cytotoxicity of HUVECs treated
with Cur-MGO adducts
An MTT assay was conducted to measure the cell
viability of HUVECs in order to investigate the role of Cur in the
protection against MGO-induced cell injury. Following exposure to
MGO or MGO + HSA, the cell viability of HUVECs was reduced compared
with HSA treatment only (P<0.05). Notably, this reduction was
restored to normal levels by Cur-MGOs (10−7,
10−6 and 10−5 M). In addition, the Cur-MGO
adducts resulted in reduced cytotoxicity compared with MGO + HSA.
Furthermore, the effect of Cur was similar to that of the AG
postive control (10−6 M; Fig. 7). These data suggest that Cur may
attenuate MGO-induced cell damage and this effect may be associated
with its ability to trap MGO.
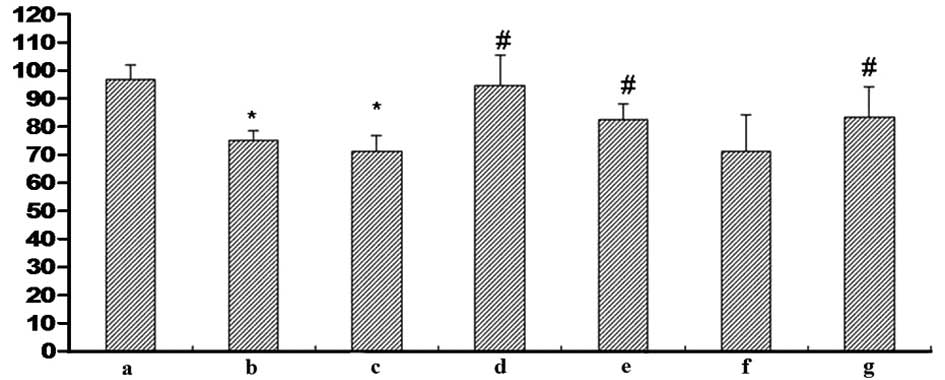 | Figure 7Effect of Cur-MGO adducts on the cell
viability of HUVECs. Cells were treated with the reaction solution
and the optical density value was determined using an MTT assay. a,
HSA; b, MGO; c, MGO + HSA; d, MGO + HSA + Cur (10−7 M);
e, MGO + HSA + Cur (10−6 M); f, MGO + HSA + Cur
(10−5 M); g, MGO + HSA + AG (10−6 M). The
data are presented as the mean ± standard deviation (n=3).
*P<0.01, vs. HSA; #P<0.05, vs. MGO or
MGO + HSA. Cur, curcumin; MGO, methylgloxal; HUVECs, human
umbilical vein endothelial cells; HSA, human serum albumin; AG,
aminoguanidine hydrochloride. |
Reduced upregulation of the expression of
ICAM-1 and TGF-β1 with Cur-MGO adducts
The expression levels of ICAM-1 and TGF-β1 in HUVECs
were measured by immunocytochemistry and western blot analysis. As
presented in Figs. 8 and 9, MGO (1 mM) or MGO + HSA upregulated the
expression levels of ICAM-1 and TGF-β1 in HUVECs following
stimulation for 48 h. In addition, incubation with Cur-MGO adducts
(10−7, 10−6 and 10−5 M) resulted
in reduced upregulation in the expression levels of ICAM-1 and
TGF-β1 in HUVECs compared with MGO or MGO + HSA (P<0.05 or
P<0.01). These data indicate that Cur is able to attenuate the
effect of MGO on the expression of ICAM-1 and TGF-β1 in HUVECs. The
effect of Cur may be associated with its ability to trap the
carbonyl compound MGO.
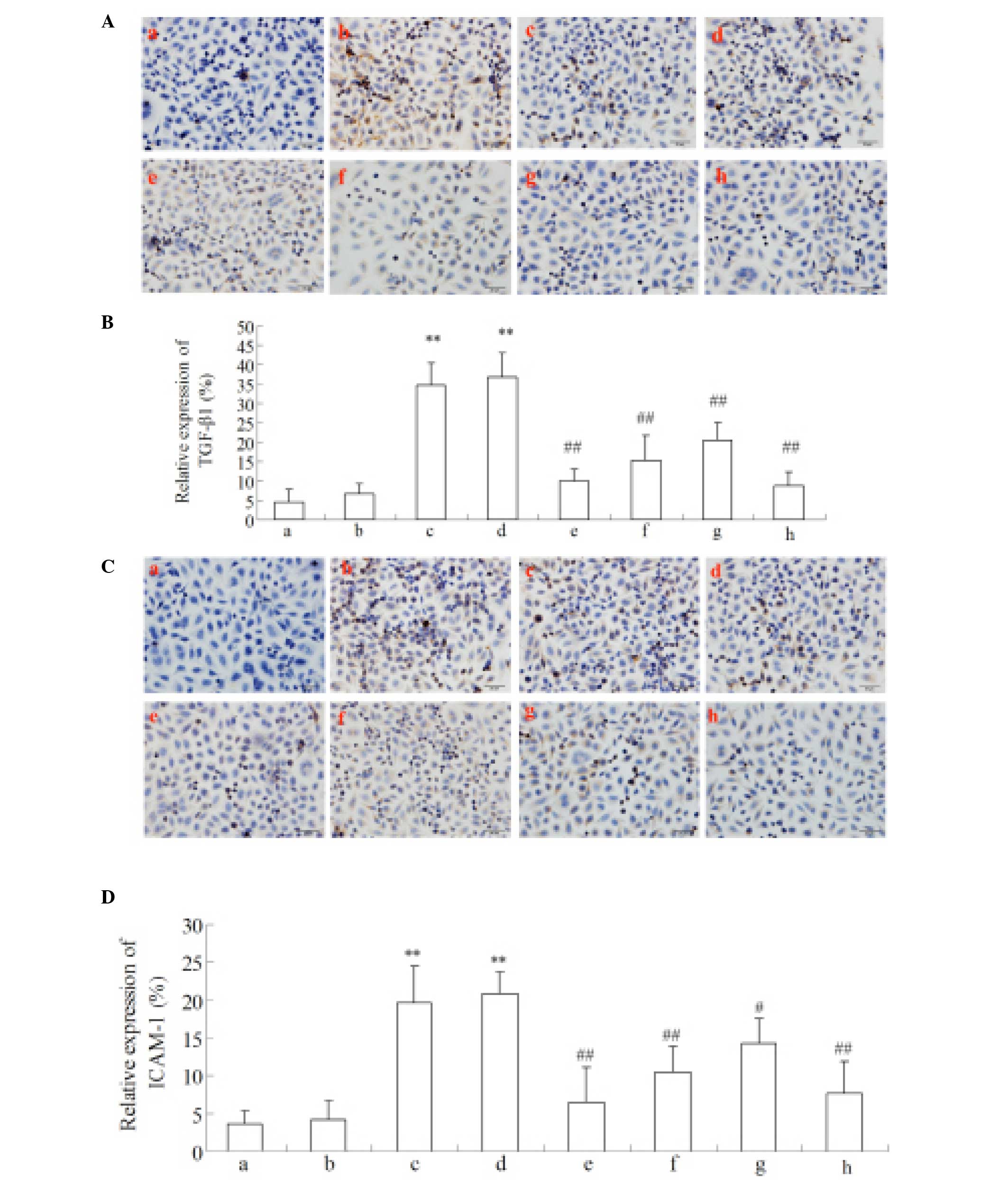 | Figure 8Regulation of Cur-MGO adducts on
inflammatory cytokines (A and B) ICAM-1 and (C and D) TGF-β1
expression in HUVECs by immunocytochemistry. a, Control; b, HSA; c,
MGO; d, MGO + HSA; e-g, MGO + HSA in the presence of
10−7, 10−6 and 10−5 M Cur,
respectively; h, MGO + HSA in the presence of 10−6 M AG.
This experiment was performed three times. The data are presented
as the mean ± standard deviation (n=3). **P<0.01, vs.
HSA; #P<0.05 and ##P<0.01, vs. MGO and
HAS+MGO. Cur, curcumin; MGO, methylgloxal; ICAM-1, intercellular
adhesion molecule-1; TGF-β1, transforming growth factor-β1; HUVECs,
human umbilical vein endothelial cells; HSA, human serum albumin;
AG, aminoguanidine hydrochloride. |
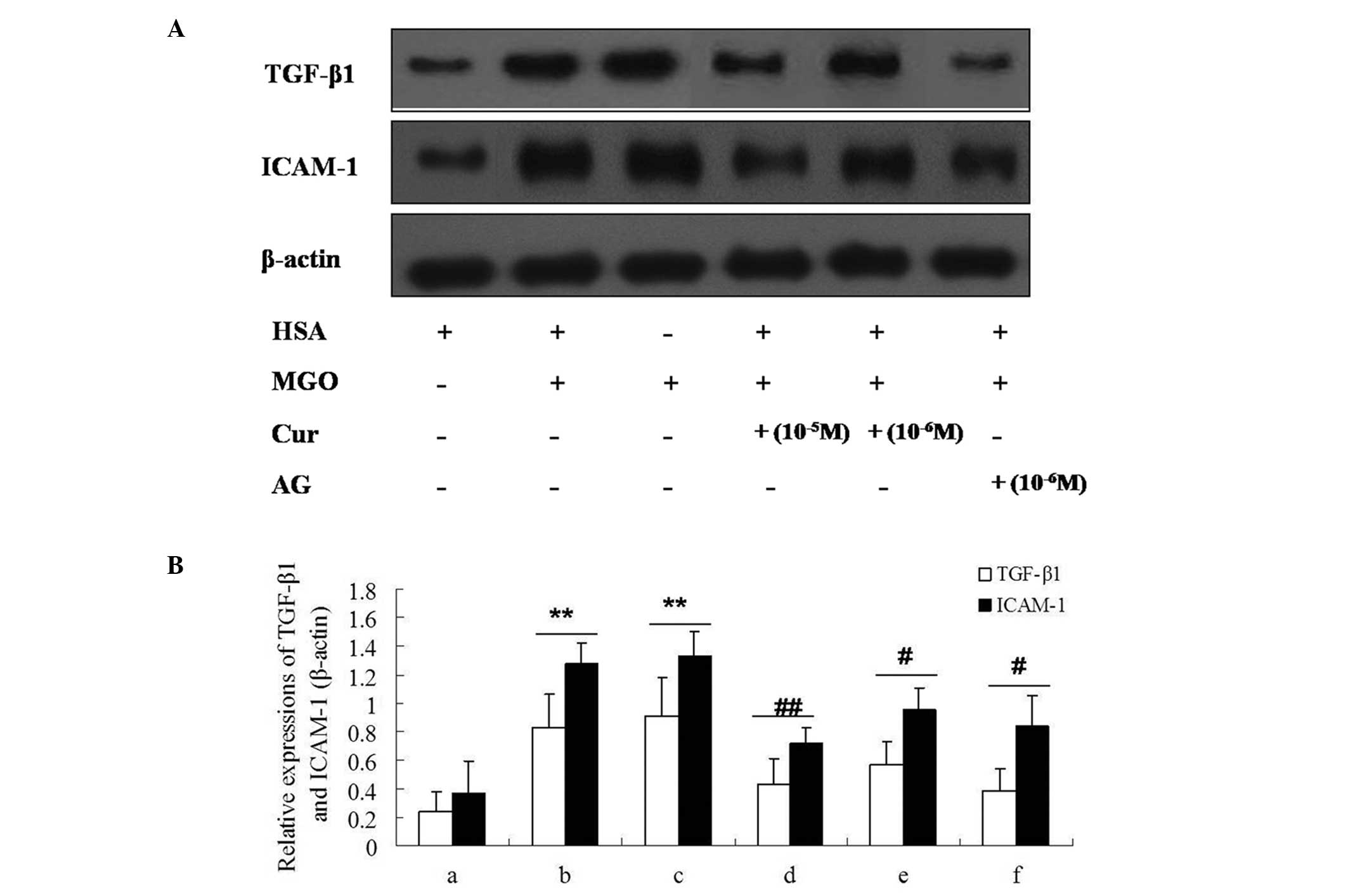 | Figure 9Effect of Cur-MGO adducts on the
expression of the inflammatory cytokines ICAM-1 and TGF-β1 in
HUVECs by western blot analysis. (A) The bands of ICAM-1 and
TGF-β1; (B) the relative expression levels of ICAM-1 and TGF-β1. a,
HSA; b, MGO; c, MGO + HSA; d, MGO + HSA + Cur (10−6 M);
e, MGO + HSA + Cur (10−5 M); f, MGO + HSA + AG
(10−6 M). Data are presented as the mean ± standard
deviation (n=3). **P<0.01, vs. HSA;
#P<0.05, ##P<0.01, vs. MGO or MGO +
HSA. Cur, curcumin; MGO, methylgloxal; ICAM-1, intercellular
adhesion molecule-1; TGF-β1, transforming growth factor-β1; HUVECs,
human umbilical vein endothelial cells; HSA, human serum albumin;
AG, aminoguanidine hydrochloride. |
Discussion
Carbonyl stress-induced oxidative damage and
inflammatory responses have been suggested as a potential mechanism
contributing to endothelial cell dysfunction (5). Under hyper-glycemic conditions, the
glucose metabolism may accelerate the abnormal accumulation of the
reactive dicarbonyl compound MGO (2). This induces oxidative stress and
inflammatory responses, resulting in the initiation and development
of diabetic vascular complications (17). Previous studies have demonstrated
that natural products are able to attenuate endothelial damage via
the capture of carbonyl compounds (3,7,10).
Cur, a major active component of Curcuma longa L., has been
demonstrated to possess beneficial effects on carbonyl
stress-induced endothelial damage (18). Although the trapping ability of Cur
on MGO is known, the effects of Cur-MGO adducts remain to be fully
elucidated. In the present study, due to the instability of the
chemical structure of Cur, additional Cur-MGO adducts were
identified by LC-ESI-MS/MS. In addition, the Cur-MGO adducts were
demonstrated to result in a reduction in cell injury compared with
MGO alone. These data demonstrate that Cur may attenuate
MGO-induced endothelial damage via the trapping of MGO to form a
number of Cur-MGO adducts.
Cur is a highly active and unstable compound which
is easily oxidized. The rate of degradation of curcuminoids
increases with increasing pH in media (18). In the current study, in order to
ensure the reliability of the reaction system, DB was used to
investigate the ability of Cur to trap the dicarbonyl compound,
MGO. These data demonstrate that the reaction product of DB and MGO
matched the 2-meth-ylquinoxaline standard, according to the UV
spectra and retention time. These results demonstrate that the
reaction system is suitable for trapping the dicarbonyl compound
MGO using natural products.
The present study investigated the optimal reaction
ratio of MGO and Cur. The reaction of MGO and Cur at the ratios of
1:5, 1:3, 1:1, 3:1 and 5:1 were analyzed using HPLC-DAD, which
indicated that a 1:1 ratio of MGO and Cur was best suited for the
reaction. The structure of the Cur-MGO adducts which were derived
from the condensation of one molecule Cur and one molecule MGO
confirmed this was the optimal reaction ratio. Although the
reaction time of Cur and MGO was not investigated, the reaction was
maintained for 720 min according to the investigation of the
reaction time of DB and MGO. These conditions were established to
be suitable for the reaction of Cur and MGO.
The −OH and −OCH3 groups on the phenyl
ring of Cur possess strong electron-donating abilities and increase
the electron density of the benzene ring, resulting in an increase
in the electrophilic reactivity of the carbon chain (19). However, the electrophilic
reactivity of the carbon chain of Cur is reduced by the loss of the
−OCH3 group on the benzene ring with a reduction in
electronic capacity. Following the reaction with MGO under
experimental conditions, adduct 1 was identified in the reaction
system based on the fragment ions. The condensation reaction
between Cur and the aldehyde group of MGO resulted in the loss of
one molecule of H2O. The results of the current study
are in agreement with previous studies (6,7,10,18,19).
In addition, the current study identified two further compounds,
adducts 2 and 3. Analysis using HPLC/ESI-MS-MS demonstrated that
these two compounds have lost two −OCH3 groups, with
441, 411 and 381 m/z fragments ions. These data provide evidence
that adduct 1 was unstable and easily formed adducts 2 and 3 in the
reaction system. However, it remains unclear whether Cur
condensates with MGO prior to or following the loss of the
−OCH3 groups.
The formation and accumulation of AGEs has been
suggested to contribute to endothelial dysfunction. Trapping the
dicarbonyl compound MGO has been demonstrated to inhibit the
formation of AGEs to attenuate carbonyl stress-induced cell injury
(20). In the current study, the
inhibition of the formation of AGEs by Cur (10−7,
10−6, 10−5 M) was investigated from 0–168 h.
The kinetic curves of the formation of AGEs indicated that the
formation was stable from 24–168 h. Furthermore, the rate of AGE
formation was reduced by the Cur-MGO adducts in a
concentration-dependent manner, further supported by the ELISA
results. The present study provides further evidence for the effect
of Cur on the attenuation of endothelial dysfunction via the
trapping of MGO to inhibit the formation of AGEs.
In order to investigate the effect of Cur on the
inhibition of the formation of MGO-AGEs by the trapping of MGO, the
oxidative damage and inflammatory responses of the reaction product
of MGO and HSA in the presence or absence of Cur were compared in
the current study. The data collected demonstrate that the
cytotoxicity of MGO + HSA in presence of Cur is reduced compared
with MGO + HSA alone. The present study provides further evidence
for the Cur-mediated attenuation of the pathogenesis and
development of endothelial damage via MGO trapping.
Carbonyl stress may induce oxidative stress which is
associated with endothelial cell dysfunction. The oxidative
modification of HSA by MGO is a causative factor in oxidative
injury (21). Increasing evidence
demonstrates that MGO stimulated super-oxide production from
mitochondria and partially stimulated ROS generation in HUVECs
(22,23). In the current study, MGO was
modified by Cur to form Cur-MGO adducts which were observed to
reduce the cytotoxicity of MGO. Notably, there was a significant
reduction in the oxidative damage resulting from incubation with
Cur-MGO compared with MGO or MGO + HSA. DHE staining and flow
cytometry indicated that the trapping of MGO by Cur is a potential
mechanism by which carbonyl stress-induced endothelial damage may
be ameliorated.
ICAM-1 and TGF-β1 serve important roles in the
systemic inflammatory response in generalized endothelial
dysfunction (24). In the current
study, following exposure of HUVECs to MGO or MGO + HSA, the
expression levels of ICAM-1 and TGF-β1 were significantly
increased, compared with HSA alone. However, following stimulation
with Cur-MGO, the expression levels of ICAM-1 and TGF-β1 were
reduced compared with MGO + HSA without Cur. These data indicate
that the cytotoxicity of MGO was attenuated due to the trapping of
MGO by Cur.
Taken together, these data provide evidence
demonstrating that Cur attenuates the cytotoxicity of the
dicarbonyl compound MGO in endothelial damage through the trapping
of MGO. In the current study, the coreaction of Cur with MGO
resulted in the formation of Cur-MGO adducts, two of which have not
previously been described. The results of LC-ESI-MS/MS indicated
that the two novel adducts may be formed due to the instability of
adduct 1 or Cur. The differences in oxidative damage and the
inflammatory responses to Cur-MGO and MGO + HSA indicate that the
attenuation of endothelial damage by Cur may be associated with its
ability to trap dicarbonyl compounds, such as MGO. The current
study provides further evidence for the protective role of Cur
against carbonyl stress-induced endothelial damage.
Acknowledgments
The current study was supported by grants from the
National Natural Science Foundation of China (grant no. 81202906)
and the Education Department of Shaanxi Province (grant no.
12JK1019).
References
|
1
|
Sabayan B, Westendorp RG, Grond J, Stott
DJ, Sattar N, van Osch MJ, van Buchem MA and de Craen AJ: Markers
of endothelial dysfunction and cerebral blood flow in older adults.
Neurobiol Aging. 35:373–377. 2014. View Article : Google Scholar
|
|
2
|
Žižek B, Žižek D, Bedenčič K, Jerin A and
Poredoš P: Effect of metabolic abnormalities on endothelial
dysfunction in normotensive offspring of subject with hypertension.
Int Angiol. 32:386–393. 2013.PubMed/NCBI
|
|
3
|
Lo CY, Li S, Tan D, Pan MH, Sang S and Ho
CT: Trapping reactions of reactive carbonyl species with tea
polyphenols in simulated physiological conditions. Mol Nutr Food
Res. 50:1118–1128. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
van Eupen MG, Schram MT, Colhoun HM,
Hanssen NM, Niessen HW, Tarnow L, Parving HH, Rossing P, Stehouwer
CD and Schalkwijk CG: The methylglyoxal-derived AGE
tetrahy-dropyrimidine is increased in plasma of individuals with
type 1 diabetes mellitus and in atherosclerotic lesions and is
associated with sVCAM-1. Diabetologia. 56:1845–1855. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Akhand AA, Hossain K, Mitsui H, Kato M,
Miyata T, Inagi R, Du J, Takeda K, Kawamoto Y, Suzuki H, et al:
Glyoxal and methylglyoxal trigger distinct signals for map family
kinases and caspase activation in human endothelial cells. Free
Radic Biol Med. 31:20–30. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Santel T, Pflug G, Hemdan NY, Schäfer A,
Hollenbach M, Buchold M, Hintersdorf A, Lindner I, Otto A, Bigl M,
et al: Curcumin inhibits glyoxalase 1: A possible link to its
anti-inflammatory and anti-tumor activity. PLoS One. 3:e35082008.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liu JP, Feng L, Zhu MM, Wang RS, Zhang MH,
Hu SY, Jia XB and Wu JJ: The in vitro protective effects of
curcumin and demethoxycurcumin in Curcuma longa extract on advanced
glycation end products-induced mesangial cell apoptosis and
oxidative stress. Planta Med. 78:1757–1760. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hu TY, Liu CL, Chen JY and Hu ML: Curcumin
ameliorates methylglyoxal-induced alterations of cellular
morphology and hyperpermeability in human umbilical vein
endothelial cells. J Funct Foods. 5:745–754. 2013. View Article : Google Scholar
|
|
9
|
Lip H, Yang K, MacAllister SL and O'Brien
PJ: Glyoxal and methylglyoxal: Autoxidation from dihydroxyacetone
and polyphenol cytoprotective antioxidant mechanisms. Chem Biol
Interact. 202:267–274. 2013. View Article : Google Scholar
|
|
10
|
Hu TY, Liu CL, Chyau CC and Hu ML:
Trapping of methylglyoxal by curcumin in cell-free systems and in
human umbilical vein endothelial cells. J Agric Food Chem.
60:8190–8196. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lv L, Shao X, Chen H, Ho CT and Sang S:
Genistein inhibits advanced glycation end product formation by
trapping methylglyoxal. Chem Res Toxicol. 24:579–586. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Schmitt A, Bigl K, Meiners I and Schmitt
J: Induction of reactive oxygen species and cell survival in the
presence of advanced glycation end products and similar structures.
Biochim Biophys Acta. 1763:927–936. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Feng L, Zhu M, Zhang M, Gu J, Jia X, Tan
X, Gao C and Zhu Q: The protection of
4,4′-diphenylmethane-bis(methyl) carbamate from Cortex Mori on
advanced glycation end product-induced endothelial dysfunction: Via
inhibiting AGE formation or blocking AGEs-RAGE axis? Fitoterapia.
89:239–249. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Feng L, Zhu MM, Zhang MH, Wang RS, Tan XB,
Song J, Ding SM, Jia XB and Hu SY: Protection of glycyrrhizic acid
against AGEs-induced endothelial dysfunction through inhibiting
RAGE/NF-κB pathway activation in human umbilical vein endothelial
cells. J Ethnopharmacol. 148:27–36. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li Y, Gu JF, Zou X, Wu J, Zhang MH, Jiang
J, Qin D, Zhou JY, Liu BX, Zhu YT, et al: The anti-lung cancer
activities of steroidal saponins of P polyphylla Smith var
chinensis (Franch) Hara through enhanced immunostimulation in
experimental Lewis tumor-bearing C57BL/6 mice and induction of
apoptosis in the A549 cell line. Molecules. 18:12916–12936. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Feng L, Zhu M, Zhang M, Jia X, Cheng X,
Ding S and Zhu Q: Amelioration of compound
4,4′-diphenylmethane-bis(methyl) carbamate on high mobility group
box1-mediated inflammation and oxidant stress responses in human
umbilical vein endothelial cells via RAGE/ERK1/2/NF-κB pathway. Int
Immunopharmacol. 15:206–216. 2013. View Article : Google Scholar
|
|
17
|
Li W, Maloney RE, Circu ML, Alexander JS
and Aw TY: Acute carbonyl stress induces occludin glycation and
brain microvascular endothelial barrier dysfunction: Role for
gluta-thione-dependent metabolism of methylglyoxal. Free Radic Biol
Med. 54:51–61. 2013. View Article : Google Scholar :
|
|
18
|
Singh R, Kristensen S and Tønnesen HH:
Influence of vehicle properties and excipients on hydrolytic and
photochemical stability of curcumin in preparations containing
Pluronics: Studies of curcumin and curcuminoids XLVIII. Pharmazie.
68:160–169. 2013.PubMed/NCBI
|
|
19
|
Deshpande SS and Maru GB: Effects of
curcumin on the formation of benzo[a]pyrene derived DNA adducts in
vitro. Cancer Lett. 96:71–80. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Okouchi M, Okayama N and Aw TY:
Preservation of cellular glutathione status and mitochondrial
membrane potential by N-acetylcysteine and insulin sensitizers
prevent carbonyl stress-induced human brain endothelial cell
apoptosis. Curr Neurovasc Res. 6:267–278. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ciolino HP and Levine RL: Modification of
proteins in endothelial cell death during oxidative stress. Free
Radic Biol Med. 22:1277–1282. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tatsunami R, Oba T, Takahashi K and Tampo
Y: Methylglyoxal causes dysfunction of thioredoxin and thioredoxin
reductase in endothelial cells. J Pharmacol Sci. 111:426–432. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Miyazawa N, Abe M, Souma T, Tanemoto M,
Abe T, Nakayama M and Ito S: Methylglyoxal augments intracellular
oxidative stress in human aortic endothelial cells. Free Radic Res.
44:101–107. 2010. View Article : Google Scholar
|
|
24
|
Szarka A, Rigó J Jr, Lázár L, Beko G and
Molvarec A: Circulating cytokines, chemokines and adhesion
molecules in normal pregnancy and preeclampsia determined by
multiplex suspension array. BMC Immunol. 11:592010. View Article : Google Scholar : PubMed/NCBI
|