Introduction
Atherosclerotic cardiovascular disease is one of the
most common causes of mortality, and is considered a major burden
on the healthcare systems of developed countries (1,2).
Inflammation has a central role in the progression of atherogenesis
(3,4). Previous studies have reported an
association between inflammation and the early stages of
atherosclerosis, including foam cell formation, monocyte adhesion
and migration (5–7). Toll-like receptor 4 (TLR4) is a type
of pattern recognition receptor, which elicits inflammation when
activated by endogenous risk factors with pathogen-associated
molecular patterns, including minimally modified low-density
lipoprotein and lipopolysaccharide (LPS). TLR4 is able to induce
activation and nuclear translocation of the transcription factor
nuclear factor-κB (NF-κB), resulting in the expression of
interleukin (IL)-1β, IL-6, IL-18, tumor necrosis factor-α (TNF-α)
and other inflammatory mediators (8). Numerous studies have confirmed the
importance of TLR4 and NF-κB in the suppression of atherosclerosis
(9,10); therefore, regulation of the
TLR4/NF-κB signaling pathway may have considerable potential in the
treatment of atherogenesis. CLI-095, also known as resatorvid or
TAK-242, is a small-molecule inhibitor of TLR4 signaling that is
used for the treatment of septic shock, which acts by binding to
the intracellular domain of TLR4. CLI-095 potently suppresses both
ligand-dependent and -independent signaling of TLR4 (11). To determine the efficacy of CLI-095
for suppressing atherogenesis, the effects and mechanisms of
CLI-095 were investigated in vitro and in vivo in the
present study. The results demonstrated that CLI-095 was able to
alleviate atherosclerotic plaque development in apolipoprotein
E-deficient (ApoE−/−) mice by reducing foam cell
formation.
Materials and methods
Chemicals and antibodies
The TLR4-specific inhibitor CLI-095 was purchased
from InvivoGen (San Diego, CA, USA) and was dissolved in dimethyl
sulfoxide (DMSO) to obtain a stock solution of 100 μg/ml.
The following antibodies: Rabbit polyclonal anti-CD68 (cat. no.
ab125212), rat monoclonal anti-α-anti-smooth muscle actin (SMA;
cat. no. ab7817), rabbit polyclonal anti-lectin-like oxidized
low-density lipoprotein receptor-1 (Lox-1; cat. no. ab60178),
rabbit polyclonal anti-ATP-binding cassette transporter A1 (ABCA1;
cat. no. ab7360), rabbit polyclonal anti-acyl-coenzyme
A:cholesterol acyltransferase-1 (ACAT-1; cat. no. ab71407), rabbit
polyclonal anti-NF-κB P65 (cat. no. ab16502) and mouse monoclonal
anti-TLR4 (cat. no. ab30667) were purchased from Abcam (Cambridge,
UK). Rabbit monoclonal anti-phosphorylated-NF-κB P65 (cat. no.
3033) and rabbit monoclonal anti-GAPDH (cat. no. 5174) were
purchased from Cell Signaling Technology, Inc. (Danvers, MA,
USA).
Cholesterol, triglyceride and cholesteryl
ester (CE) measurements
Total cholesterol and triglyceride commercial kits
were obtained from the Jiancheng Bioengineering Institute (Nanjing,
China). Cholesteryl ester (CE) enzyme-linked immunosorbent assay
kit was obtained from Nanjing Anpei Electro-Mechanics Equipment
Co., Ltd. (Nanjing, China). To measure total cholesterol and
triglyceride, serum was separated from hemocytes by centrifugation
at 13,523 × g at 4°C for 5 min and commercial kits were used
according to the manufacturer's protocol. To conduct the CE ELISA,
cells were harvested and washed in cold phosphate-buffered saline
(PBS; Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA) and
lipids were extracted by resuspending the sample in 200 μl
chloroform (Sigma-Aldrich, St. Louis, MO, USA), isopropanol
(Amresco, LLC, Solon, Ohio) and NP-40 (Beyotime Institute of
Biotechnology, Inc., Haimen, China) at a ratio of 7:11:0.1 in a
Bullet Blender Storm microhomogenizer (Midwest Scientific, Valley
Park, MO, USA) at room temperature. The extract was spun for 5–10
min at 15,000 × g and the ELISA kit was used according to the
manufacturer's instructions.
Animal protocol
Male ApoE−/− mice (n=15; age, 6 weeks)
were purchased from Beijing HFK Bioscience Co., Ltd. (Beijing,
China) and maintained under a specific pathogen-free environment in
micro-isolator cages. They were kept at 18–23°C at humidity 40–60%
under a 12-h light/dark cycle. The mice were separated into the
control, vehicle and treatment groups, with six mice in each group.
The mice in the control group were fed a chow diet, whereas the
mice in the vehicle and treatment groups were fed a high-fat diet
(21% fat, 0.15% cholesterol; Mediscience Diets Co., Ltd., Yangzhou,
China). In addition, the mice in the treatment group received a
daily intraperitoneal injection of CLI-095 at a dose of 3 mg/kg/day
(mouse body weight) for 10 weeks. The mice in the vehicle group
were administered a daily intraperitoneal injection of 20% DMSO/PBS
(volume, 0.1 ml) Mice were sacrificed at 16 weeks of age. Following
sacrifice by inhalation of CO2 (Sigma-Aldrich), blood
and heart tissue samples were collected. All animal procedures were
approved by the Animal Ethics Committee of the Dalian Medical
University (Dalian, China). All in vivo experiments were
performed in accordance with national legislation and institutional
guidelines.
Atherosclerotic lesion analysis
The lesion area in the aortic root sections was
measured following Oil-red-O and hematoxylin-eosin (H&E)
staining, using computer-assisted image quantification with Image
Pro Plus 6.0 (Media Cybernetics, Inc., Rockville, MD, USA). Images
were captured using an Olympus fluorescent microscope (DP80;
Olympus Corp., Tokyo, Japan). Collagen fibers were stained with
Masson's trichrome stain. All staining solutions were obtained from
BASO Precision Optics Ltd. (Taiwan, China).
Immunohistochemistry and
immunocytochemistry
Frozen sections of the aortic root were fixed in
methanol (Sigma-Aldrich), incubated with 3%
H2O2 (ZSGB-BIO, Beijing, China), air-dried
and incubated with 10% goat serum (ZSGB-BIO), for 15–30 min. The
frozen sections were incubated with anti-CD68, anti-α-SMA and
anti-Lox-1 antibodies overnight at 4°C. Fluorophore-conjugated
secondary antibodies were used for immunofluorescence. Macrophages
were extracted from the rat by cutting the outer skin of the
peritoneum and exposing the inner skin lining the peritoneal
cavity, 5 ml PBS [with 3% fetal bovine serum (FBS)] was injected
into the peritoneal cavity using a 27 g needle. Following
injection, the peritoneum was gently massaged to dislodge attached
cells and the fluid was collected with a 25 g needle. The collected
cell suspension was centrifuged at 211 × g for 8 min, the
supernatant was discarded and the cells harvested. Macrophages
extracted from the mice were fixed and stained with anti-Lox-1
antibody and 4′,6-diamidino-2-phenylindole.
Cell culture
Thioglycolate-elicited peritoneal macrophages were
maintained in RPMI 1640 media (Gibco; Thermo Fisher Scientific,
Inc.) supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo
Fisher Scientific, Inc.) and 100 U/ml penicillin-streptomycin
(Gibco; Thermo Fisher Scientific, Inc.) at 37°C in an atmosphere
containing 5% CO2.
Lipoprotein uptake assay
Thioglycolate-elicited peritoneal macrophages were
seeded in serum-free medium. Following overnight fasting, the
macrophages were washed with PBS and cultured in medium with or
without oxidized low-density lipoprotein (Ox-LDL; 100 μg/ml;
Guangzhou Yiyuan Biotechnology Co., Ltd., Guangzhou, China).
Subsequently, the cells were treated with CLI-095 (1 μM) for
36 h at 37°C in a CO2 incubator. Macrophages were fixed
and stained with Oil-red-O and observed under a DP80 fluorescent
microscope. Experiments were repeated in triplicate in each
group.
Western blot analysis
Cell total proteins were extracted using a total
protein extraction kit from Nanjing KeyGen Biotech. Co., Ltd.
(Nanjing, China), which contains a lysis buffer, proteinase
inhibitor, phosphorylase inhibitor and phenylmethylsulfonyl
fluoride. The protein concentration was determined using a Pierce™
BCA Protein assay kit (Thermo Fisher Scientific, Inc.). Proteins
(30–50 μg) were separated by 12% sodium-dodecyl
sulfate-polyacrylamide gel electrophoresis for 1.5 h at 120 V in
electrophoretic buffer solution. The proteins were then transferred
to polyvinylidene difluoride membranes (EMD Millipore, Billerica,
MA, USA). Following blocking with 5% milk/Tris-buffered saline
(w/v; Sigma-Aldrich) at 25°C for 1 h, the membranes were incubated
with primary antibodies at 37°C for 4 h, including anti-Lox-1
(dilution, 1:1,000), anti-GAPDH (1:1,000), anti-TLR4 (1:500),
anti-T-P65 (1:1,000), anti-ABCA1 (1:1,000) and anti-ACAT1
(1:1,000). The membranes were then washed and incubated with
horseradish peroxidase-conjugated goat anti-rabbit IgG (cat. no.
sc-2004; dilution, 1:10,000) and horseradish peroxidase-conjugated
goat anti-mouse IgM (cat. no. sc-2064; dilution, 10,000) secondary
antibodies obtained from Santa Cruz Biotechnology, Inc., Dallas,
TX, USA), and Texas Red-X conjugated goat anti-rat IgG secondary
antibody (dilution, 1:200; Thermo Fisher Scientific, Inc.; cat. no.
T-6392) for 1 h at room temperature. The blots were treated with
WesternBright ECL kit (Advansta Inc., Menlo Park, CA, USA)
visualized using Bio-Rad imaging system (Bio-Rad Laboratories,
Inc., Hercules, CA, USA) and analyzed using Image Pro Plus 6.0.
Statistical analysis
All data are presented as the mean ± standard
deviation. Comparisons between different groups were analyzed using
two-tailed Student's t-test. Statistical analysis was performed
using SPSS version 13.0 statistical software (SPSS Inc., Chicago,
IL, USA). P<0.05 was considered to indicate a statistically
significant difference.
Results
CLI-095 potently attenuates the
development of atherosclerosis
To address the role of TLR4 in the development of
atherosclerosis, ApoE−/− mice were fed an atherogenic
high-fat diet and were treated with or without CLI-095 via a daily
intraperitoneal injection for a period of 10 weeks (dose, 3
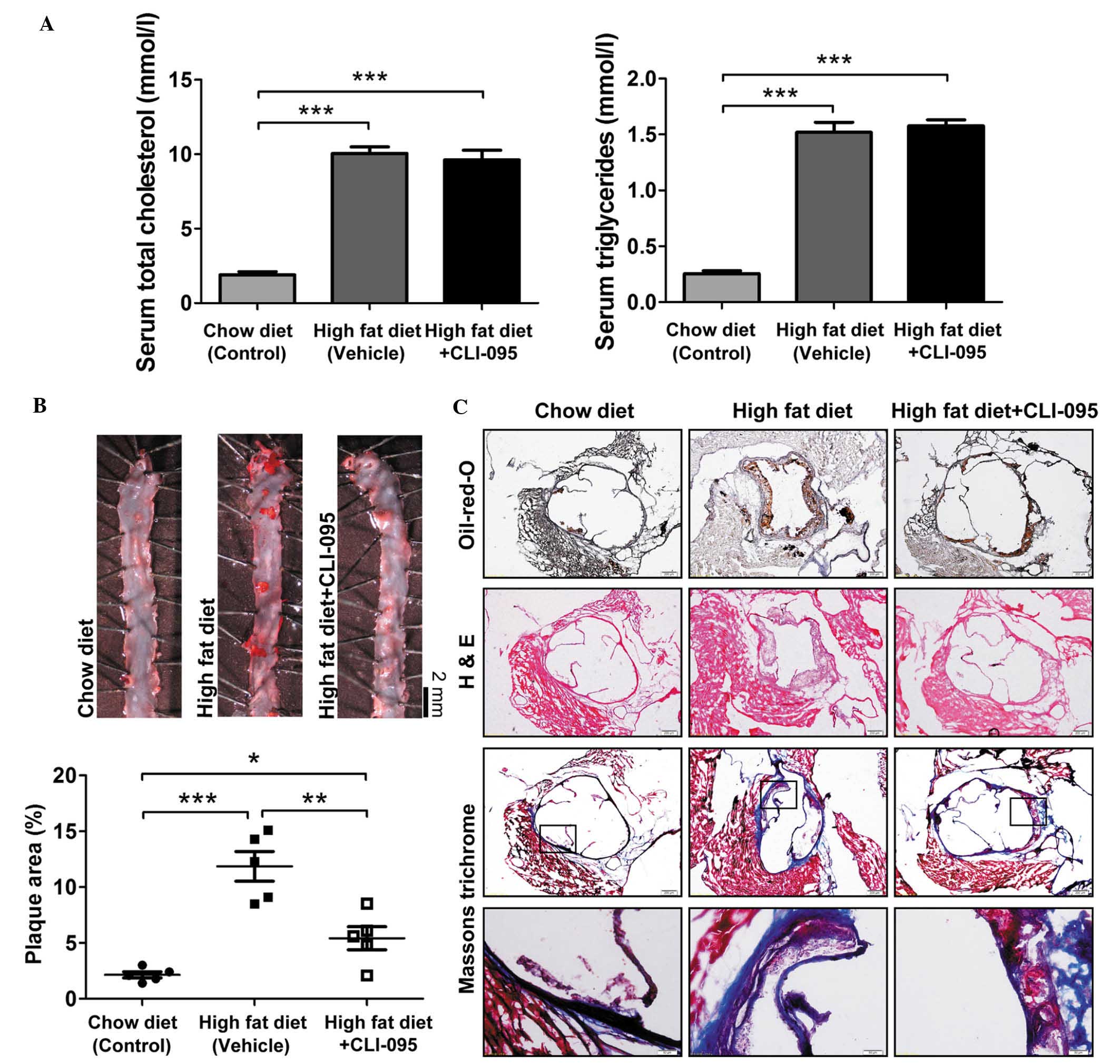
mg/kg/day). As expected, ApoE−/− mice fed an atherogenic
high-fat diet exhibited a significant increase in cholesterol and
triglyceride levels, as compared with the chow diet-fed mice. In
addition, ApoE−/− mice fed an atherogenic diet and
treated with CLI-095 did not exhibit any variation in lipid
metabolism, as compared with the vehicle-treated group (Fig. 1A). Subsequently, en face Oil-red-O
staining was performed on the thoracic aorta. Mice treated with the
TLR4 inhibitor CLI-095 exhibited markedly reduced atherosclerotic
plaque size (Fig. 1B), suggesting
that CLI-095 exerts a protective effect on atherosclerosis.
Oil-red-O and H&E staining of serial cross sections of aortic
roots also revealed fewer atherosclerotic lesions in the
CLI-095-treated mice, as compared with the vehicle-treated mice;
however, Masson's trichrome staining revealed that the total
collagen content did not differ between the CLI-095- and
vehicle-treated groups (Fig. 1C).
These results indicate that CLI-095 may protect against
atherosclerosis in ApoE−/− mice.
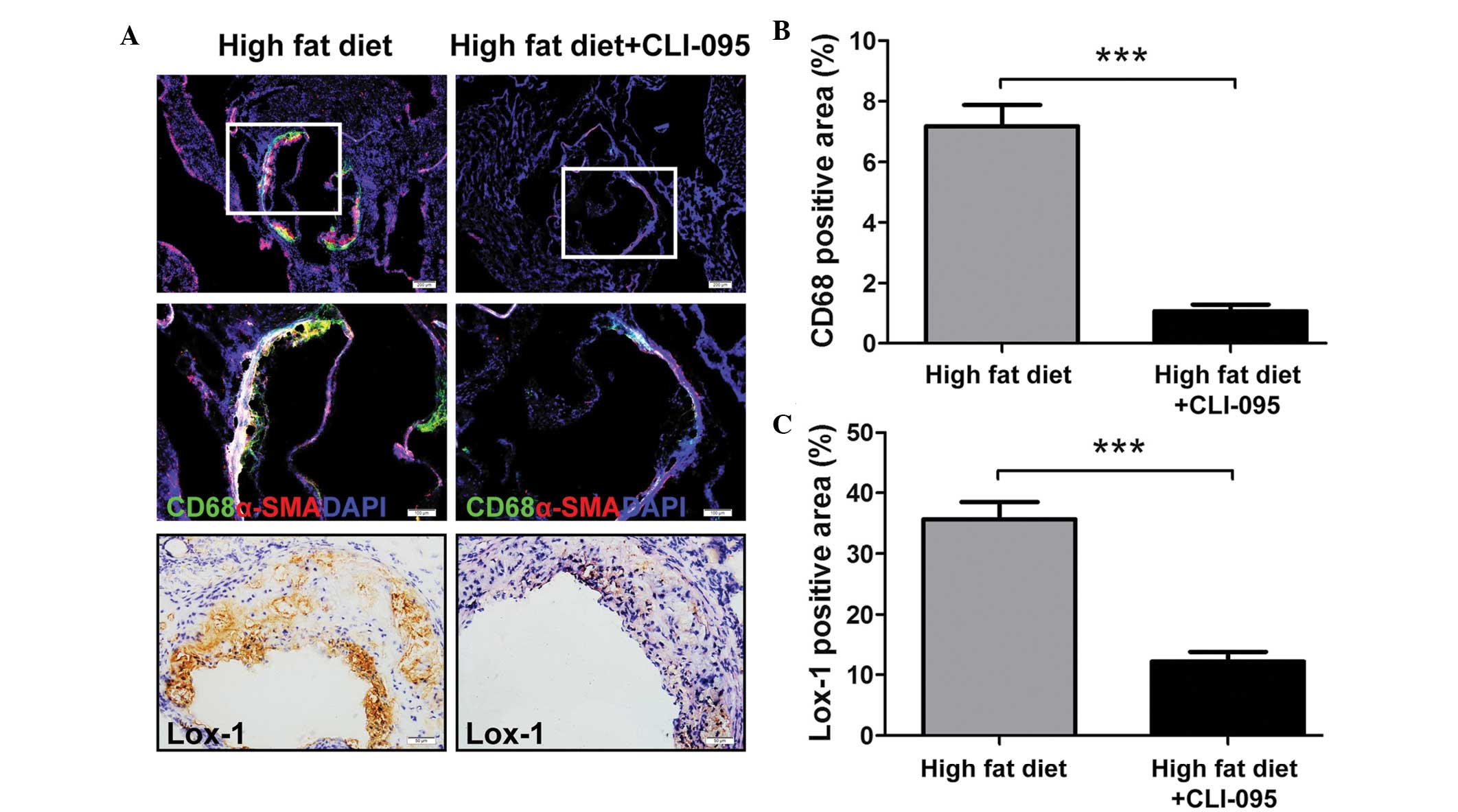
CLI-095 reduces macrophage recruitment to
atherosclerotic plaques
The recruitment of macrophages to the subintimal
space is considered to have a key role in the progression of
atherosclerosis (12). The present
study revealed that the vehicle-treated ApoE−/− mice
displayed clear macrophage recruitment to the atherosclerotic
plaques, whereas only a limited number of macrophages were retained
in the CLI-095-treated mice, as determined by immunofluorescent
staining (Fig. 2A). The relative
CD68-positive areas in the CLI-095-treated mice (n=5) were
significantly smaller than those in the vehicle-treated mice (n=5)
(Fig. 2B). In addition, α-SMA
positive areas were markedly increased in the atherosclerotic
plaques of the vehicle-treated group (Fig. 2A).
A previous study demonstrated that overexpression of
Lox-1, a macrophage and endotheliocyte receptor for Ox-LDL
(13), in ApoE−/− mice
upregulated the endothelial expression of vascular cell adhesion
molecule-1 and increased the accumulation of CD68-positive cells in
the plaques (14). In order to
investigate whether CLI-095 modulates Lox-1 expression in plaque
formation, the expression of Lox-1 was examined.
Immunohistochemical staining of cross sections revealed a decreased
expression of Lox-1 in CLI-095-treated mice, as compared with the
vehicle-treated controls (Fig. 2A and
C). These results suggest that CLI-095 is sufficient to reduce
atherosclerotic plaque formation, possibly via the modulation of
Lox-1 expression.
CLI-095 is sufficient to reduce murine
peritoneal macrophage (MPM) foam cell formation in vitro
Lipoprotein uptake by macrophages promotes the
formation of foam cells, which, in turn, contribute to the
progression of atherosclerosis (15). To determine whether TLR4 inhibition
is sufficient to regulate foam cell formation in
thioglycolate-elicited MPMs in vitro, MPMs were treated with
or without CLI-095, and foam cell formation was induced by Ox-LDL.
Notably, the accumulation of cytoplasmic lipid droplets decreased
in the CLI-095-treated MPMs, as compared with the vehicle-treated
MPMs, as determined by Oil-red-O staining, and CLI-095 treatment
resulted in a significant suppression of MPM foam cell formation
(Fig. 3A). The upregulation of
Lox-1 has been shown to promote lipoprotein uptake in macrophages
(16), and the present study
demonstrated that CLI-095 could suppress Lox-1 expression in cross
sections from ApoE−/− mice (Fig. 2A). Subsequently, the expression of
Lox-1 was determined using immunofluorescent staining and western
blot analysis. The data revealed that stimulation of MPMs with
Ox-LDL resulted in a significant increase in Lox-1 protein
expression (vehicle vs. control), whereas CLI-095 treatment
significantly decreased Lox-1 expression (Fig. 3B). These results suggest that
CLI-095 exerts its protective effects on lipoprotein uptake by
downregulating the expression of Lox-1.
CLI-095 decreases the quantity of CE in
MPMs by differentially regulating the expression of ABCA1 and
ACAT-1
The accumulation of CE in macrophages has a critical
role in foam cell formation. The possible effects of CLI-095 on
macrophage CE accumulation in vitro were examined in the
present study. Treatment with CLI-095 resulted in a marked decrease
in the abundance of cellular CE in MPMs, as compared with the
vehicle group (Fig. 4A).
Furthermore, CLI-095 resulted in a marked reduction in the ratio of
cellular CE to total cholesterol in MPMs (Fig. 4B). During the process of foam cell
formation, excess cellular free cholesterol is converted to CE by
the enzyme ACAT-1, or is removed from the cell by ABCA1-dependent
cholesterol efflux (17–19). In addition, activation of NF-κB can
suppress ABCA1 and enhance ACAT-1 expression to promote CE-laden
cell formation (20,21). In the present study, Ox-LDL
stimulation resulted in enhanced TLR4 expression as previously
reported (22,23); however, the expression of TLR4 was
not altered in the CLI-095-treated MPMs, as compared with the
vehicle-treated MPMs (Fig. 4C).
Notably, treatment with TLR4 inhibitor CLI-095 significantly
reduced Ox-LDL-induced phosphorylation of NF-κB P65 (Fig. 4D), suggesting that CLI-095 may
inhibit TLR4 signaling by affecting its adaptor proteins but
without downregulating its expression. Furthermore, it was observed
that CLI-095 markedly promoted ABCA1 expression and attenuated
ACAT-1 expression (Fig. 4E and F).
These data strongly indicate that CLI-095 may exert its vascular
protective function by restricting CE synthesis and enhancing
cholesterol efflux in macrophages.
Discussion
TLR4 is a member of the TLR family (TLR1-TLR13),
which regulates the innate immune response. The TLR4/NF-κB pathway
is one of the most studied signaling pathways in atherosclerosis;
it is well known that some atherogenic factors, including LPS and
Ox-LDL, are important ligands of TLR4. Following ligand binding to
TLR4, the intracellular toll/interleukin-1 receptor (TIR) domains
of TLR4 recruit the following signaling adaptor proteins: TIR
domain-containing adaptor protein, myeloid differentiation factor
88, TIR-domain-containing adaptor-inducing interferon-β (TRIF) and
TRIF-related adaptor molecule. Subsequently, numerous kinases and
ubiquitin ligases, such as IL-1 receptor-associated kinase
(IRAK)-1, IRAK-4, tumor necrosis factor receptor-associated factor
6 and transforming growth factor-β-activated kinase 1, are
recruited and activated, culminating in the nuclear translocation
of NF-κB (8). The activated NF-κB
can subsequently induce the production of inflammatory mediators
(8). Over the past decade, several
TLR4 inhibitors have been developed to treat inflammatory diseases
in animals (24,25). CLI-095, a selective inhibitor of
TLR4 signaling, has been shown to potently suppress the activation
of NF-κB, as well as the production of TNF-α, which is induced by
TLR4-specific ligands interfering with the interactions between
TLR4 intracellular domain and its adaptors in macrophages (11,26).
The present study demonstrated that decreased NF-κB activation in
CLI-095-treated mouse macrophages was not dependent on TLR4
expression. Some of these findings are consistent with those of
previous studies (27,28).
Atherosclerosis is a chronic inflammatory disease.
TLR4 has been shown to promote atherosclerosis by increasing the
secretion of inflammatory mediators, foam cell formation and
monocyte adhesion in ApoE−/− mice fed a high-fat diet
(9,22,29).
In addition, NF-κB has been demonstrated to augment foam cell
formation by interfering with cellular lipid metabolism (30–32).
In the present study, the molecular mechanisms underlying
CLI-095-induced reductions in atherosclerotic lesions were
determined. The results suggested that CLI-095 was able to inhibit
the TLR4/NF-κB signaling cascade, in order to promote cholesterol
efflux, and suppress lipoprotein uptake and CE synthesis in
macrophages.
Foam cell formation has a critical role in
atherosclerosis and its mechanisms include uptake of atherogenic
lipoproteins, impaired cellular cholesterol efflux and disturbed
intracellular cholesterol processing. Notably, Lox-1, ABCA1 and
ACAT-1 have an important role in these three mechanisms
respectively. The present data suggested that CLI-095 may
upregulate the expression of ABCA1 and downregulate that of Lox-1
and ACAT-1 in vitro. These findings were consistent with
those from previous studies; both ABCA1 overexpression and Lox-1
deficiency in mice have been shown to lead to decreased
atherosclerotic lesions (14,33–35).
However, previous studies have demonstrated that loss of ACAT-1
expression may lead to severe atherosclerosis (36,37).
A possible explanation for the controversy between previous studies
and the present results is that global ACAT-1 knockout causes
monocytosis in ApoE−/− mice during the development of
atherosclerosis; however, there is no evidence to demonstrate that
the inhibition of TLR4 signaling causes monocytosis.
Both the present and previous studies demonstrated
that TLR4 regulates cholesterol biosynthesis in vitro;
however, whether TLR4 is able to regulate lipid metabolism in
vivo remains controversial. The results of the present study
revealed that CLI-095 did not reduce increased serum cholesterol
and triglyceride levels in mice receiving a high-fat diet. In
addition, Higashimori et al (9) reported that TLR4 deficiency was not
associated with reduced levels of cholesterol and triglycerides,
whereas Aspichueta et al (38) reported that endotoxic rats
exhibited increased levels of serum very low-density
lipoprotein-apoB, -triglyceride, and -cholesterol. In addition, Lu
et al (39) reported that
Rs-LPS, a TLR4 antagonist, decreased the serum levels of
cholesterol and triglycerides in non-diabetic mice; therefore,
further research is required to determine whether and how TLR4
affects lipid metabolism in animals.
In conclusion, the results of the present study
demonstrated that the TLR4 inhibitor CLI-095 was able to
effectively reduce atherosclerosis in ApoE−/− mice by
suppressing foam cell formation. The present study also provides
novel insights into the protective effects of TLR4 inhibition on
enhancing cholesterol efflux by upregulating the expression of
ABCA1, and reducing CE biosynthesis by downregulating the
expression of ACAT-1, which is mediated by inhibiting the
TLR4/NF-κB signaling pathway. These results suggested that TLR4 may
be considered a potential therapeutic target for the prevention of
atherosclerotic progression.
Acknowledgments
The authors of the present study would like to thank
Dr Dan He and Dr Kuang Peng for their helpful discussions and
technical assistance. This study was supported by funds from the
Second Affiliated Hospital of Dalian Medical University and the
National Natural Science Foundation of China (grant no.
81372853).
References
|
1
|
Yang Z and Hall AG: The financial burden
of overweight and obesity among elderly Americans: The dynamics of
weight, longevity, and health care cost. Health Serv Res.
43:849–868. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hansson GK: Inflammation, atherosclerosis,
and coronary artery disease. N Engl J Med. 352:1685–1695. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Libby P, Ridker PM and Hansson GK:
Progress and challenges in translating the biology of
atherosclerosis. Nature. 473:317–325. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hansson GK and Libby P: The immune
response in atherosclerosis: A double-edged sword. Nat Rev Immunol.
6:508–519. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhuang J, Peng W, Li H, Lu Y, Wang K, Fan
F, Li S and Xu Y: Inhibitory effects of vinpocetine on the
progression of atherosclerosis are mediated by Akt/NF-κB dependent
mechanisms in apoE−/− mice. PLoS One. 8:e825092013.
View Article : Google Scholar
|
|
6
|
Wu S, Xu H, Peng J, Wang C, Jin Y, Liu K,
Sun H and Qin J: Potent anti-inflammatory effect of dioscin
mediated by suppression of TNF-α-induced VCAM-1, ICAM-1 and EL
expression via the NF-κB pathway. Biochimie. 110:62–72. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jehs T, Faber C, Juel HB, Bronkhorst IH,
Jager MJ and Nissen MH: Inflammation-induced chemokine expression
in uveal melanoma cell lines stimulates monocyte chemotaxis. Invest
Ophthalmol Vis Sci. 55:5169–5175. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lim KH and Staudt LM: Toll-like receptor
signaling. Cold Spring Harb Perspect Biol. 5:a0112472013.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Higashimori M, Tatro JB, Moore KJ,
Mendelsohn ME, Galper JB and Beasley D: Role of toll-like receptor
4 in intimal foam cell accumulation in apolipoprotein E-deficient
mice. Arterioscler Thromb Vasc Biol. 31:50–57. 2011. View Article : Google Scholar :
|
|
10
|
Gareus R, Kotsaki E, Xanthoulea S, van der
Made I, Gijbels MJ, Kardakaris R, Polykratis A, Kollias G, de
Winther MP and Pasparakis M: Endothelial cell-specific NF-kappaB
inhibition protects mice from atherosclerosis. Cell Metab.
8:372–383. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kawamoto T, Ii M, Kitazaki T, Iizawa Y and
Kimura H: TAK-242 selectively suppresses Toll-like receptor
4-signaling mediated by the intracellular domain. Eur J Pharmacol.
584:40–48. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Libby P: Inflammation in atherosclerosis.
Nature. 420:868–874. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sawamura T, Kume N, Aoyama T, Moriwaki H,
Hoshikawa H, Aiba Y, Tanaka T, Miwa S, Katsura Y, Kita T and Masaki
T: An endothelial receptor for oxidized low-density lipoprotein.
Nature. 386:73–77. 1997. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Akhmedov A, Rozenberg I, Paneni F, Camici
GG, Shi Y, Doerries C, Sledzinska A, Mocharla P, Breitenstein A,
Lohmann C, et al: Endothelial overexpression of LOX-1 increases
plaque formation and promotes atherosclerosis in vivo. Eur Heart J.
35:2839–2848. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Itabe H: Oxidized low-density
lipoproteins: What is understood and what remains to be clarified.
Biol Pharm Bull. 26:1–9. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hossain E, Ota A, Karnan S, Takahashi M,
Mannan SB, Konishi H and Hosokawa Y: Lipopolysaccharide augments
the uptake of oxidized LDL by up-regulating lectin-like oxidized
LDL receptor-1 in macrophages. Mol Cell Biochem. 400:29–40. 2015.
View Article : Google Scholar
|
|
17
|
Chang TY, Chang CC, Ohgami N and Yamauchi
Y: Cholesterol sensing, trafficking, and esterification. Annu Rev
Cell Dev Biol. 22:129–157. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sekiya M, Osuga J, Igarashi M, Okazaki H
and Ishibashi S: The role of neutral cholesterol ester hydrolysis
in macrophage foam cells. J Atheroscler Thromb. 18:359–364. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shao B, Tang C, Sinha A, Mayer PS,
Davenport GD, Brot N, Oda MN, Zhao XQ and Heinecke JW: Humans with
atherosclerosis have impaired ABCA1 cholesterol efflux and enhanced
high-density lipoprotein oxidation by myeloperoxidase. Circ Res.
114:1733–1742. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lei L, Xiong Y, Chen J, Yang JB, Wang Y,
Yang XY, Chang CC, Song BL, Chang TY and Li BL: TNF-alpha
stimulates the ACAT1 expression in differentiating monocytes to
promote the CE-laden cell formation. J Lipid Res. 50:1057–1067.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhao GJ, Tang SL, Lv YC, Ouyang XP, He PP,
Yao F, Chen WJ, Lu Q, Tang YY, Zhang M, et al: Antagonism of
betulinic acid on LPS-mediated inhibition of ABCA1 and cholesterol
efflux through inhibiting nuclear factor-kappaB signaling pathway
and miR-33 expression. PLoS One. 8:e747822013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yin YW, Liao SQ, Zhang MJ, Liu Y, Li BH,
Zhou Y, Chen L, Gao CY, Li JC and Zhang LL: TLR4-mediated
inflammation promotes foam cell formation of vascular smooth muscle
cell by upregulating ACAT1 expression. Cell Death Dis. 5:e15742014.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yang K, Zhang XJ, Cao LJ, Liu XH, Liu ZH,
Wang XQ, Chen QJ, Lu L, Shen WF and Liu Y: Toll-like receptor 4
mediates inflammatory cytokine secretion in smooth muscle cells
induced by oxidized low-density lipoprotein. PLoS One.
9:e959352014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sun Y and Pearlman E: Inhibition of
corneal inflammation by the TLR4 antagonist Eritoran tetrasodium
(E5564). Invest Ophthalmol Vis Sci. 50:1247–1254. 2009. View Article : Google Scholar
|
|
25
|
Fenhammar J, Rundgren M, Forestier J,
Kalman S, Eriksson S and Frithiof R: Toll-like receptor 4 inhibitor
TAK-242 attenuates acute kidney injury in endotoxemic sheep.
Anesthesiology. 114:1130–1137. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Matsunaga N, Tsuchimori N, Matsumoto T and
Ii M: TAK-242 (resatorvid), a small-molecule inhibitor of Toll-like
receptor (TLR) 4 signaling, binds selectively to TLR4 and
interferes with interactions between TLR4 and its adaptor
molecules. Mol Pharmacol. 79:34–41. 2011. View Article : Google Scholar
|
|
27
|
Gárate I, García-Bueno B1, Madrigal JL,
Caso JR, Alou L, Gómez-Lus ML and Leza JC: Toll-like 4 receptor
inhibitor TAK-242 decreases neuroinflammation in rat brain frontal
cortex after stress. J Neuroinflammation. 11:82014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sun M, Deng B, Zhao X, Gao C, Yang L, Zhao
H, Yu D, Zhang F, Xu L, Chen L and Sun X: Isoflurane
preconditioning provides neuroprotection against stroke by
regulating the expression of the TLR4 signalling pathway to
alleviate microglial activation. Sci Rep. 5:114452015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lee SJ, Choi EK, Seo KW, Bae JU, Park SY
and Kim CD: TLR4-mediated expression of Mac-1 in monocytes plays a
pivotal role in monocyte adhesion to vascular endothelium. PLoS
One. 9:e1045882014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ferreira V, van Dijk KW, Groen AK, Vos RM,
van der Kaa J, Gijbels MJ, Havekes LM and Pannekoek H:
Macrophage-specific inhibition of NF-kappaB activation reduces
foam-cell formation. Atherosclerosis. 192:283–290. 2007. View Article : Google Scholar
|
|
31
|
Wang Y, Wu JF, Tang YY, Zhang M, Li Y,
Chen K, Zeng MY, Yao F, Xie W, Zheng XL, et al: Urotensin II
increases foam cell formation by repressing ABCA1 expression
through the ERK/NF-κB pathway in THP-1 macrophages. Biochem Biophys
Res Commun. 452:998–1003. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lee HY, Kim SD, Baek SH, Choi JH, Cho KH,
Zabel BA and Bae YS: Serum amyloid A stimulates macrophage foam
cell formation via lectin-like oxidized low-density lipoprotein
receptor 1 upregulation. Biochem Biophys Res Commun. 433:18–23.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Brunham LR, Singaraja RR, Duong M, Timmins
JM, Fievet C, Bissada N, Kang MH, Samra A, Fruchart JC, McManus B,
et al: Tissue-specific roles of ABCA1 influence susceptibility to
atherosclerosis. Arterioscler Thromb Vasc Biol. 29:548–554. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hu C, Dandapat A, Sun L, Chen J, Marwali
MR, Romeo F, Sawamura T and Mehta JL: LOX-1 deletion decreases
collagen accumulation in atherosclerotic plaque in low-density
lipoprotein receptor knockout mice fed a high-cholesterol diet.
Cardiovasc Res. 79:287–293. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Westerterp M, Murphy AJ, Wang M, Pagler
TA, Vengrenyuk Y, Kappus MS, Gorman DJ, Nagareddy PR, Zhu X,
Abramowicz S, et al: Deficiency of ATP-binding cassette
transporters A1 and G1 in macrophages increases inflammation and
accelerates atherosclerosis in mice. Circ Res. 112:1456–1465. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Fazio S, Major AS, Swift LL, Gleaves LA,
Accad M, Linton MF and Farese RV Jr: Increased atherosclerosis in
LDL receptor-null mice lacking ACAT1 in macrophages. J Clin Invest.
107:163–171. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
37
|
Huang LH, Gui J, Artinger E, Craig R,
Berwin BL, Ernst PA, Chang CC and Chang TY: Acat1 gene ablation in
mice increases hematopoietic progenitor cell proliferation in bone
marrow and causes leukocytosis. Arterioscler Thromb Vasc Biol.
33:2081–2087. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Aspichueta P, Pérez-Agote B, Pérez S,
Ochoa B and Fresnedo O: Impaired response of VLDL lipid and apoB
secretion to endotoxin in the fasted rat liver. J Endotoxin Res.
12:181–192. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lu Z, Zhang X, Li Y, Jin J and Huang Y:
TLR4 antagonist reduces early-stage atherosclerosis in diabetic
apolipoprotein E-deficient mice. J Endocrinol. 216:61–71. 2013.
View Article : Google Scholar
|