Introduction
Rheumatic diseases are characterized by inflammation
and autoimmune activation. This is more evident in paediatric
patients, where the pathogenesis of rheumatic diseases appears to
possess intermediate features between two ends of an immunologic
disease continuum, represented by monogenic autoinflammatory and
autoimmune diseases (1).
This complex pathogenesis may account for the
difficulties in completely curing rheumatic diseases, regardless of
the improved therapeutic approaches, which relieve symptoms and
prevent severe complications in the majority of cases. In addition,
the requirement for enduring treatments may have a negative impact
on patient quality of life and upon health costs (2).
Current treatments are based on glucocorticoids,
disease modifying anti-rheumatic drugs (DMARDs) [such as
methotrexate (MTX) and leflunomide] and on biological drugs
targeting inflammatory cytokines [anti-interleukin (IL)-1,
anti-IL-6, anti-tumour necrosis factor (TNF)-α] or targeting immune
cell activation (such as CTLA4Ig). Furthermore, novel molecules,
such as kinase inhibitors [for example, tofacitinib (Tofa) or
baricitinib] have recently been introduced (3–6).
Although the number of available drugs is
increasing, treatment options and schedules remain largely
empirical, and are based on their ability to delay or prevent
tissue damage in clinical trials. Considering the role of
inflammation and autoimmunity in rheumatic disorders, approaches to
treatment include drugs with prevalent anti-inflammatory action
(such as glucocorticoids and anti-cytokine biologics) and
immunosuppressive agents (such as kinase inhibitors, MTX,
cyclosporine, CTLA4Ig and Janus kinase). Different drugs can be
administered in sequential schedules (step-up, commencing with
drugs with lower costs and toxicity; top-down, commencing with
drugs with high potency to induce a remission that can be
maintained using other drugs with higher safety and lower costs) or
combination treatments, to exploit their different mechanisms of
action (7–9).
However, the preclinical data regarding how these
schedules act on the underlying mechanisms of the pathogenesis and
progression of rheumatic diseases (such as activation and spreading
of autoimmunity, while preserving protective immunity) is poor.
An in vitro model, in which the fate of
unstimulated and stimulated cells could be differentially tracked
by flow cytometry, was developed to reproduce an experimental
condition relevant to the pathogenesis of rheumatic diseases, in
which only certain lymphocytes are hypothesized to undergo
antigen-driven activation. This model was used to investigate
differences in action and possible synergisms between the two
DMARDs, MTX and Tofa, by analysing the proliferation, activation
and cytokine secretion of T cells. It was found that the stimulated
lymphocytes and unstimulated cells were affected by the treatment.
Furthermore, resting cells undergo a bystander activation due to
the presence of stimulated lymphocytes and are consequently
affected by the treatment.
Materials and methods
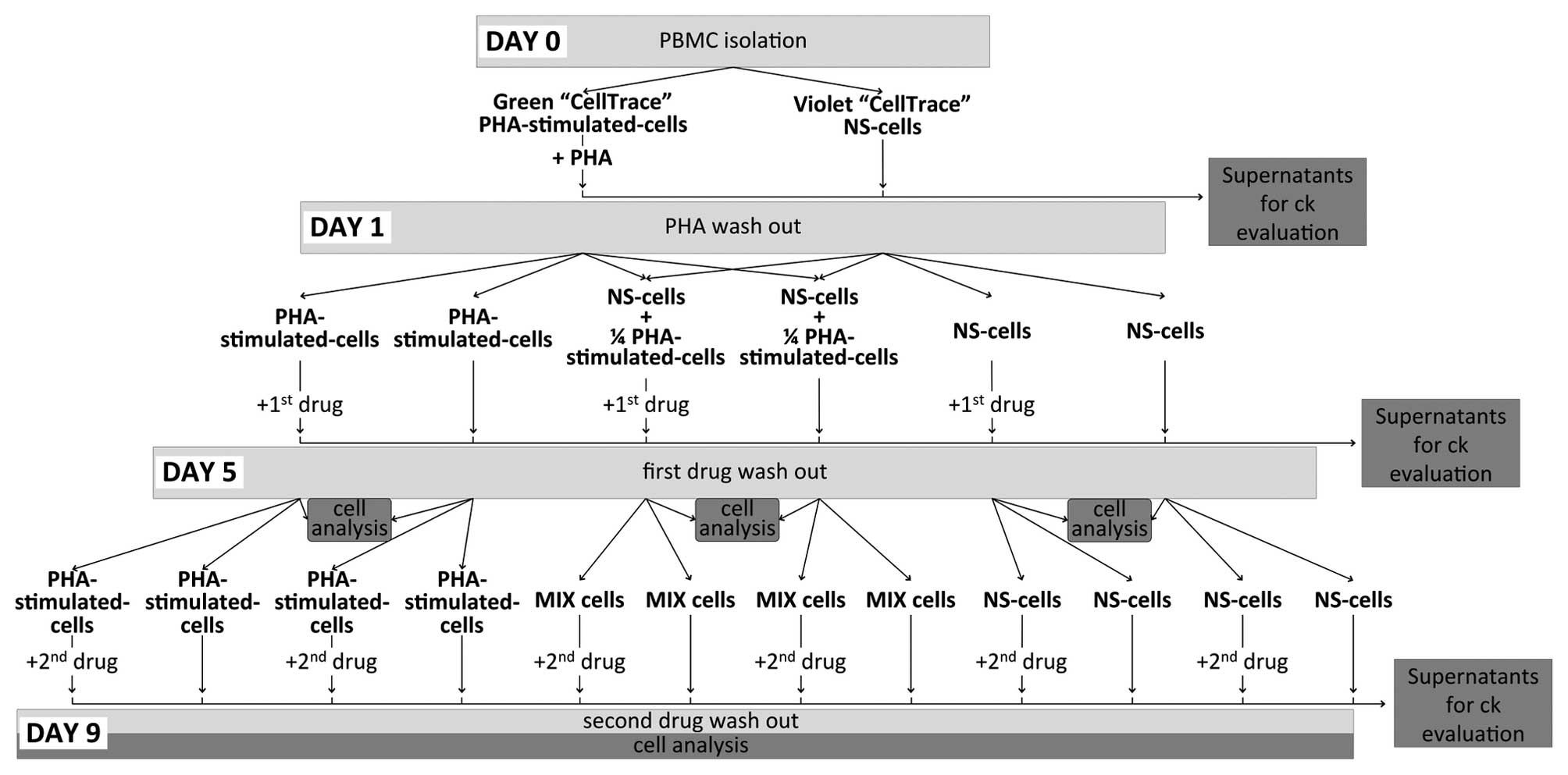
Experimental plan
The present study was approved by the Institutional
Review Board of the Institute for Maternal and Child Health, IRCCS
'Burlo Garofolo', Trieste, Italy (RC 28/2009). Peripheral blood
mononuclear cells (PBMCs) were obtained from the buffy coats of six
healthy donors subsequent to receiving written consent. PBMCs were
cultured for 9 days according to the experimental plan presented in
Fig. 1. Briefly, at day 0, PBMCs
were obtained by density gradient centrifugation (at 500 × g for 30
min without breaking, at room temperature) and divided into two
groups. The first group contained 6×107 cells stained
with 5 µM CellTrace™ Oregon Green® 488 Carboxylic
Acid Diacetate, Succinimidyl Ester (Carboxy-DFFDA, SE; CellTrace™
Green) obtained from Thermo Fisher Scientific, Inc. (Waltham, MA,
USA) and stimulated with 2 µg/ml phytohaemagglutinin (PHA;
Biochrom GmbH, Berlin, Germany), while the second group
(4×107 cells) was stained with 5 µM CellTrace™
Violet Cell Proliferation kit (CellTrace™ Violet; Thermo Fisher
Scientific, Inc.) and maintained in a culture without direct
stimulation (non-stimulated; NS-cells). After 24 h of independent
culture (day 1), the two groups of cells were washed with saline
(Braun Melsungen AG, Melsungen, Germany) to remove the PHA and were
re-plated in the presence of 200 µM MTX (Teva
Pharmaceuticals Industries, Ltd., Milan, Italy) or 10 µM
Tofa (Sigma-Aldrich, St Louis, MO, USA), or in the absence of
drugs. Concentrations of Tofa and MTX were selected on the basis of
previous studies (10,11) to achieve an inhibitory effect on
cell proliferation and activation without a marked effect on cell
viability. To mimic a pathogenic setting containing stimulated and
unstimulated cells, PHA-stimulated cells and NS-cells were plated
together at a 1:4 ratio (designated as MIX). On day 5, the cells
were washed to remove the first drug and were plated in the
presence or absence of the second drug (either Tofa or MTX) for a
further four days (day 9). For all experiments, cells were
incubated at 37°C (atmosphere, 5% CO2) at a
concentration of 106 cells/ml in X-VIVO™ 15 medium
(Lonza Verviers Sprl, Verviers, Belgium) supplemented with 10%
Human AB Serum (Sigma-Aldrich), 100 U/ml penicillin/streptomycin
and 200 mM L-glutamine (EuroClone S.p.A., Milan, Italy).
Flow cytometric analysis of proliferation
and activation
At day 5 and day 9, 2×105 cells were
stained with allophycocyanine/Cy7-conjugated anti-human CD25
monoclonal antibody (final dilution, 2.5 µg/ml; cat. no.
302614; BioLegend, Inc., San Diego, CA, USA) to evaluate
activation. Simultaneously, staining with CellTrace™ was performed
before cells were placed in the culture, to evaluate cell
proliferation, which was assessed using a dye dilution assay
following each cell division (12)
and then expressed as a percentage of proliferated cells.
Activation of cells was evaluated by measuring the median
fluorescence intensity (MFI) of CD25. Data were obtained using a
CyAn™ ADP cytometer (Beckman Coulter, Inc., Brea, CA, USA) and
analysed with FlowJo software version 7.6 (Tree Star, Inc.,
Ashland, OR, USA).
Analysis of cytokines in culture
supernatants
To measure the levels of cytokines released by the
cells, at day 1, 5 and 9, culture supernatants were aspirated,
collected in a vial and stored at −20°C until analysis.
Concentrations were evaluated using the magnetic-beads suspension
assay using Bio-Plex® technology (Bio-Plex Pro™, Human
Cytokine 8-Plex Panel; Bio-Rad Laboratories, Inc., Hercules, CA,
USA). The analysis included eight cytokines and chemokines as
follows: IL-2, -4, -6, -8, -10, granulocyte-macrophage
colony-stimulating factor, interferon-γ (IFN-γ), TNF-α. Multiplex
assay suspension data were acquired using a Bio-Plex®
200 reader (Bio-Rad Laboratories, Inc.), and data output was
analysed using a high-speed digital processor (Bio-Plex Multiplex
System; Bio-Rad Laboratories, Inc.). Concentrations are presented
in units (pg/ml).
Statistical analysis
All results are presented as means of two
independent experiments ± standard deviation. Statistical
significance was calculated using a one-way analysis of variance
and Tukey's multiple comparison post-test. Analysis was performed
using GraphPad Prism software (version 5.0; GraphPad Software,
Inc., La Jolla, CA, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
When administered as the first drug, MTX
blocked proliferation, but Tofa did not exert significant
effects
After the first 24 h of culture in the presence or
absence of PHA, cells were washed, treated with the first drug,
MTX, and incubated for a further 4 days. Cells were then washed and
incubated in the presence of the second drug, Tofa, for a further 4
days. At the end of the incubation periods (days 5 and 9), the
cells were analysed by flow cytometry to assess the activation and
proliferation. At day 5 in the MIX condition, the proliferation of
PHA-stimulated cells was strongly reduced by MTX treatment:
PHAMIX, 83.35±4.65 vs. PHAMIX + MTX,
13.39±9.72% (P<0.0001; Fig.
2A). In addition, NS-cells in the MIX condition demonstrated
low percentages of proliferation that appeared to be marginally
affected by MTX, although the difference was not statistically
significant: NSMIX, 19.25±3.55 vs. NSMIX +
MTX, 8.85±3.65% (Fig. 2A).
Following the washout, at day 9, the antiproliferative effect of
MTX on PHA-stimulated cells in the MIX condition continued to be
evident: PHAMIX, 86.40±7.90 vs. PHAMIX + MTX,
22.65±5.15% (P<0.0001).
 | Figure 2Cell proliferation and activation
following treatment with MTX then Tofa. Percentage of proliferating
cells after (A) 5 and (B) 9 days of incubation with MTX then Tofa.
Activation evaluated as MFI of CD25 at (C) day 5 and (D) day 9 of
drug treatments. NS, NS-cells cultured alone; PHA, PHA-stimulated
cells cultured alone; NSMIX, NS-cells co-cultured in the
presence of PHA-stimulated cells; PHAMIX, PHA-stimulated
cells co-cultured in the presence of NS-cells. To distinguish
between the two populations, NS-cells were stained with CellTrace™
Violet, while PHA-stimulated cells were stained with CellTrace™
Green. *P<0.05 and ***P<0.0001. NS,
non-stimulated; MTX, methotrexate; PHA, phytohaemagglutinin; Tofa,
tofacitinib; MFI, median fluorescence intensity. |
The administration of Tofa on day 5 did not
demonstrate any further antiproliferative effect on PHA-stimulated
cells regardless of a previous treatment with MTX:
PHAMIX, 86.40±7.90 vs. PHAMIX + Tofa,
87.15±7.75%; PHAMIX + MTX, 22.65±5.15 vs.
PHAMIX + MTX / Tofa, 26.40±3.50% (Fig. 2B).
NS-cells in the MIX condition at day 9 showed
marginal proliferation (NSMIX, 31.70±0.10%), which
appeared to be reduced by the addition of MTX (NSMIX +
MTX, 12.50±0.80%) during the first time frame (days 1-5), although
the difference was not statistically significant, and Tofa
treatment alone during the second time frame (days 5–9) did not
exert any effects (NSMIX + MTX-Tofa, 31.20±1.60%).
Furthermore, the addition of Tofa to MTX-treated cells did not
reveal any increase in the antiproliferative effect when compared
with MTX alone: NSMIX + MTX, 12.50±0.80 vs.
NSMIX + MTX-Tofa 15.25±1.55% (Fig. 2B).
Activation was not completely blocked by
MTX
MTX induced a significant decrease in the level of
CD25 expression in PHA-stimulated cells in the MIX condition:
PHAMIX, 18.35±2.45 vs. PHAMIX + MTX,
9.03±1.27 (P<0.05; Fig. 2C).
Whereas no differences in activation was evident in the NS-cells
irrespective of the presence of MTX: NSMIX, 7.86±0.07;
NSMIX + MTX, 6.55±0.01 (Fig. 2C). At day 9, there was a residual
level of CD25 expression that was not influenced by any of the
pharmacologic treatments: PHAMIX, 8.75±0.05;
PHAMIX + MTX, 7.30±0.66; PHAMIX + MTX-Tofa,
7.44±0.52; PHAMIX + Tofa, 8.56±0.64 (Fig. 2D).
Following withdrawal of MTX treatment,
cells underwent a cytokine rebound, which was partially restored by
Tofa
At day 5, following PHA stimulation, the cells
exhibited higher concentrations of all of the cytokines when
compared with the NS condition. In addition, MTX treatment appeared
to reduce the cytokine secretion in PHA-stimulated cells alone and
in the MIX condition, but to a reduced extent. An exception was
observed for the concentration of IL-2, which was marginally
increased after MTX treatment (Fig.
3).
 | Figure 3Cytokine release in supernatants after
sequential treatment with MTX and Tofa. Bar charts represent the
cytokine concentration, expressed as pg/ml, evaluated with
Bio-Plex® technology. NS, supernatants from unstimulated
cells; PHA, supernatants from PHA-stimulated cells; MIX,
supernatants from a co-culture of NS and PHA-stimulated cells. NS,
non-stimulated; IL, interleukin; GM-CSF, granulocyte-macrophage
colony-stimulating factor; IFN-γ, interferon-γ; TNF-α, tumour
necrosis factor-α; PHA, phytohaemagglutinin; MTX, methotrexate;
Tofa, tofacitinib. |
By contrast, at day 9, PHA-stimulated cells that had
initially undergone MTX treatment showed increased production of
all cytokines when compared with the untreated cells.
Treatment with Tofa subsequent to MTX led to a
reduced cytokine secretion when compared with MTX treatment alone,
although the concentrations remained higher than those of the
untreated PHA-stimulated cells. In the MIX condition, the effects
of the drug treatments demonstrated similar trends to those of PHA
stimulation, although the concentrations were generally lower.
Furthermore, higher concentrations of IL-2 were
demonstrated in PHA-stimulated cells treated with MTX + Tofa, when
compared with other conditions (Fig.
3). Notably, no significant difference in IL-8 concentration
was demonstrated regardless of the stimulus and of the drug
treatment at day 1 and 5; conversely, a higher secretion was
observed at day 9 in the culture supernatants of the MIX-condition,
when cells were treated with Tofa alone or in combination with MTX
(Fig. 3).
Tofa demonstrated a low antiproliferative
effect on PHA-stimulated cells, increased by the addition of
MTX
Cells were cultured for 24 h with or without PHA,
and then washed and plated in the presence or absence of Tofa.
After 4 days of culture (day 5), cells were washed and incubated in
the presence or absence of MTX for a further 4 days (day 9). When
Tofa was administered as the first drug in the MIX condition,
proliferation of PHA-stimulated cells at day 5 tended to be
marginally decreased: PHAMIX, 73.60±21.40 vs.
PHAMIX + Tofa, 55.00±25.60%. NS-cells in the MIX
condition displayed an appreciable proliferation rate, which was
partly reduced by Tofa treatment: NSMIX, 15.45±4.15 vs.
NSMIX + Tofa, 8.15±4.26% (Fig. 4A).
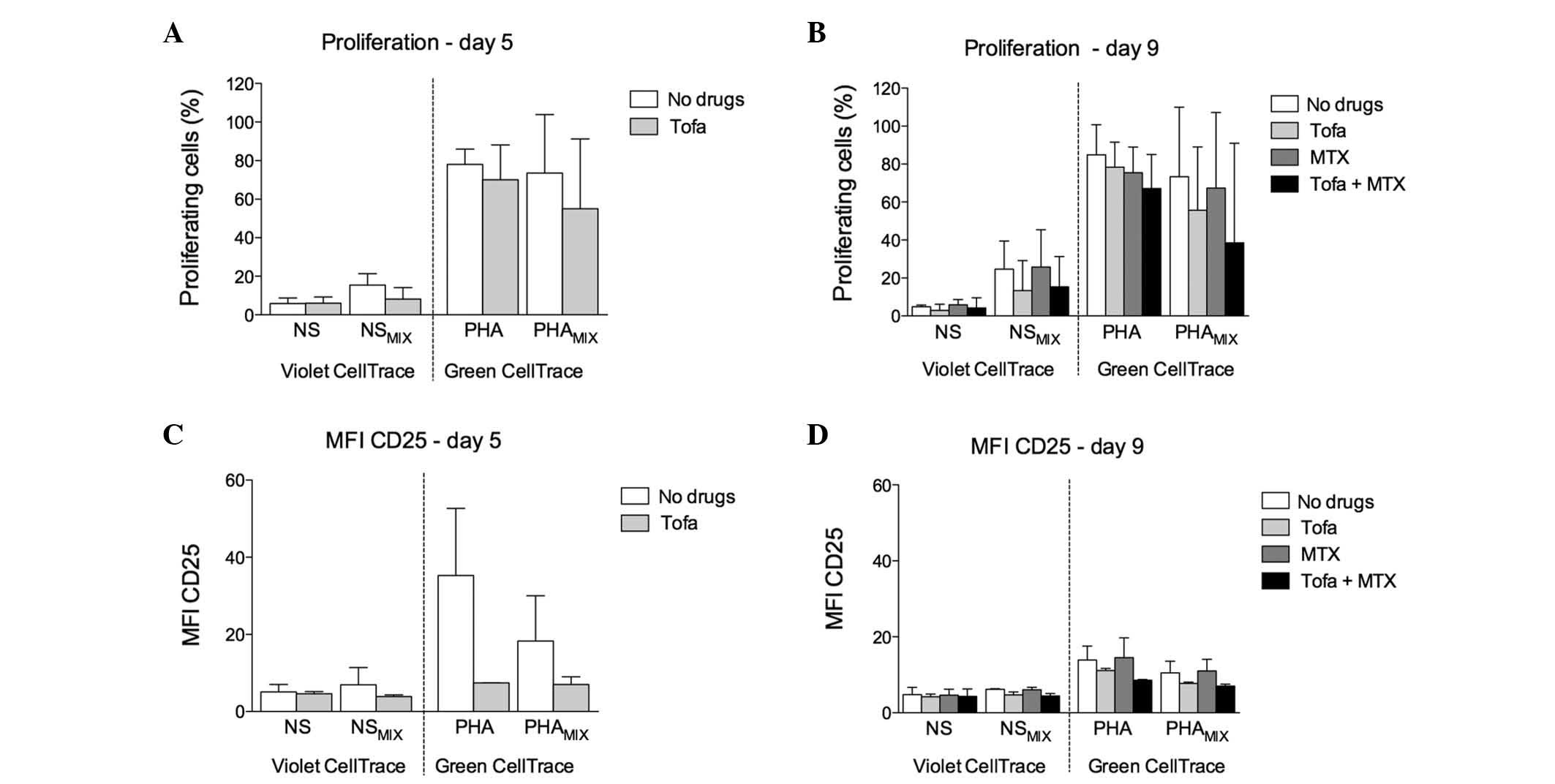 | Figure 4Effect of sequential treatment with
Tofa and MTX on cell proliferation and activation. Percentage of
proliferating cells (A) after the incubation in presence of Tofa
(day 5) and (B) following subsequent incubation with MTX at day 9.
Cell activation, presented as MFI, of CD25 (C) at day 5 and (D) at
day 9. NS, NS-cells cultured alone; PHA, PHA-stimulated cells
cultured alone; NSMIX, NS-cells co-cultured in the
presence of PHA-stimulated cells; PHAMIX, PHA-stimulated
cells co-cultured in the presence of NS-cells. To distinguish
between the two populations, NS-cells were stained with CellTrace™
Violet, while the PHA-stimulated cells were stained with CellTrace™
Green. NS, non-stimulated; PHA, phytohaemagglutinin; MTX,
methotrexate; Tofa, tofacitinib; MFI, median fluorescence
intensity. |
At day 9, sequential treatment with Tofa and MTX
induced an additional reduction of proliferation when compared with
Tofa treatment alone: PHAMIX + Tofa-MTX, 38.41±37.09 vs.
PHAMIX + Tofa, 55.65±23.65%. Furthermore, NS-cells
displayed low proliferation: NSMIX, 24.65±10.45%, which
was marginally influenced by treatment with Tofa: NSMIX
+ Tofa, 13.31±11.20%, and was not affected by the addition of MTX
during the second incubation: NSMIX + MTX, 25.75±13.95%;
NSMIX + Tofa-MTX, 15.33±11.27% (Fig. 4B).
Tofa induced an enduring reduction in
activation, but with reduced effect on NS-cells and MTX exerted
little effect
At day 5, although the differences observed were not
significant, Tofa treatment appeared to decrease the activation of
PHA-stimulated cells in the MIX condition: PHAMIX,
18.30±8.30 vs. PHAMIX + Tofa, 7.04±1.40 (Fig. 4C). Accordingly, NS-cells underwent
marginal activation (NSMIX, 6.95±3.16) that was reduced
by Tofa (NSMIX + Tofa, 3.89±0.32; Fig. 4C).
Conversely, at day 9, the addition of MTX as the
second drug did not appear to have an effect on PHA-stimulated
cells, regardless of the previous Tofa treatment:
PHAMIX, 10.53±2.17; PHAMIX + Tofa, 7.74±0.22;
PHAMIX + MTX, 11.04±2.16; PHAMIX + Tofa-MTX,
7.04±0.36 (Fig. 4D).
Tofa treatment reduced IFN-γ and IL-4
production, but MTX did not exert an effect
At day 5, Tofa treatment reduced cytokine secretion,
particularly secretion of IFN-γ: PHA, 21,857. 07±1,701.98 vs. PHA +
Tofa, 1,978.82±120.12 pg/ml (P<0.05) and of IL-4: PHA, 6.74±0.91
vs. PHA + Tofa, 4.42±0.44 pg/ml (P<0.05) in PHA-stimulated cells
and this trend was maintained in the MIX condition (Fig. 5).
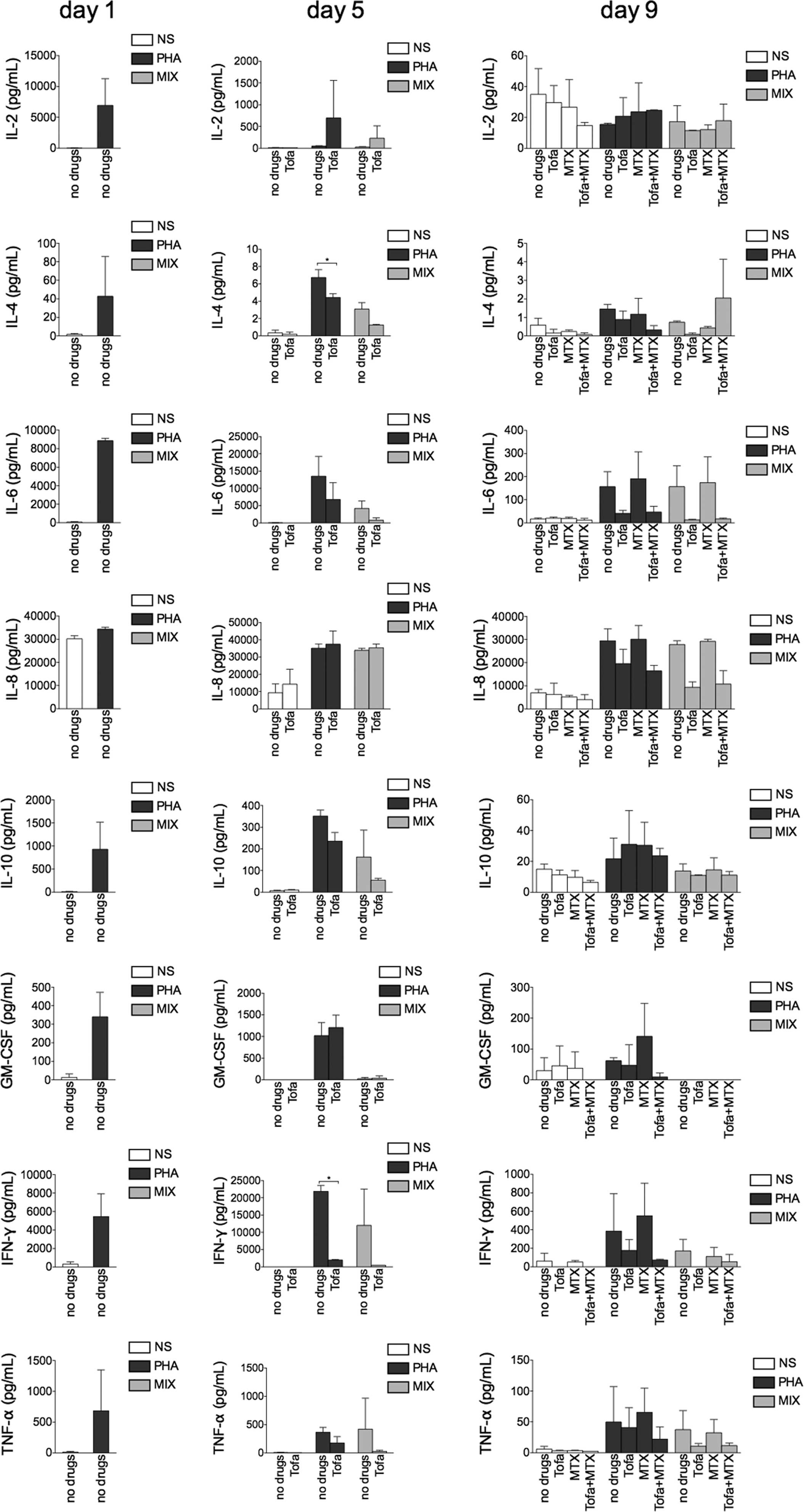 | Figure 5Cytokine evaluation in cell culture
supernatants following treatment with Tofa then MTX. The bar charts
represent the cytokine concentration, expressed as pg/ml, evaluated
with Bio-Plex® technology. NS, supernatants from
unstimulated cells; PHA, supernatants from PHA-stimulated cells;
MIX, supernatants from mixed culture of NS and PHA-stimulated
cells. *P<0.05. NS, non-stimulated; IL, interleukin;
GM-CSF, granulocyte-macrophage colony-stimulating factor; IFN-γ,
interferon-γ; TNF-α, tumour necrosis factor-α; PHA,
phytohaemagglutinin; MTX, methotrexate; Tofa, tofacitinib. |
At day 9, MTX alone did not induce a reduction of
cytokine secretion in the PHA-stimulated cells when compared with
untreated cells, although MTX treatment exerted an additional
effect to the Tofa treatment, in particular for IL-4 and IFN-γ. In
the MIX condition, this effect was less evident.
Drug treatments, in PHA-stimulated cells, induced a
higher production of IL-2. By contrast, Tofa treatment, alone and
in combination with MTX, induced a decreased production of IL-8
(Fig. 5).
Discussion
The therapeutic approach in rheumatic diseases is
based upon the administration of anti-inflammatory glucocorticoids,
DMARDs or biologics. Recently, novel drugs are also being developed
(3), and there is evidence to
suggest that combined or sequential administration of
well-established therapeutic strategies improves the patient
outcome without the high costs of biologics (7,13).
The possible mechanisms of action of combination
therapy have been poorly investigated in the pre-clinical and
laboratory settings. However, there is the potential to obtain
greater advantages in terms of better outcomes for patients, higher
safety and lower costs by exploiting the different characteristics
of novel and existing drugs in combination treatments.
Among the drugs available for rheumatic diseases,
MTX has achieved marked results in past decades and Tofa is among
the most innovative, recently developed molecules.
Although there are clinical studies that compare the
two drugs (14) or that evaluate
their use in combined treatments (15), to the best of our knowledge, data
regarding in vitro models, which examine the mechanisms of
action (10,11,16)
to optimize their pharmacological action when used in combination,
are lacking.
The present study demonstrated that MTX and Tofa
exerted different effects on cell activation and proliferation, and
it was proposed that these differences may be exploited by
sequential therapeutic strategies, to maximize the anti-rheumatic
effect. MTX showed a persistent antiproliferative effect and a
moderate downregulation of activation on PHA-stimulated cells,
which was not influenced by subsequent Tofa treatment. By contrast,
Tofa marginally effected activation, but only slight
antiproliferative action that, however, could be potentiated by
sequential treatment with MTX.
Recent studies have demonstrated that Tofa only
exerts its effect on recently activated cells. Doses higher than
those typically administered block the activation and proliferation
of stimulated lymphocytes, but the effect is transient, with the
levels of activation and proliferation restored following
withdrawal of the drug (10). The
present study proposed that proliferation of the cells following
Tofa treatment-and-withdrawal may be exploited to render cells
increasingly sensitive to sequential treatment with other drugs,
such as MTX. Indeed, MTX demonstrated a strong antiproliferative
effect on recently activated cells, during the first incubation,
although not on continuously proliferating cells during the second
time frame. However, the antiprolifera-tive effect of MTX during
the second time frame was evident when Tofa was administered first,
directly after cell stimulation.
Furthermore, on day 5, the two drugs tended to
reduce the secretion of pro-inflammatory cytokines by cells
recently stimulated with PHA; notably, IFN-γ and IL-4 levels were
significantly reduced in Tofa-treated cells (P<0.05) and this
trend was also observed in MTX-treated cells, although this
difference was not identified to be statistically significant.
By contrast, during the second time frame, the use
of different sequences of drugs caused conflicting effects on
cytokine secretion. When the sequence of MTX + Tofa was used, an
increased secretion of cytokines by cells treated with MTX only
during the first time frame was observed at day 9, as compared with
the other treatments. This effect may indicate that cell activation
became unhindered following withdrawal of MTX, which resulted in a
revived level of activation; conversely, when Tofa was added to the
culture, a reduction of cytokine secretion was observed, when Tofa
was administered alone or following MTX.
When MTX was administered alone during the second
time frame, it did not reduce, but caused an increase in, cytokine
release; however, treatment with Tofa in the first incubation
period resulted in a greater inhibition of cytokine release, which
was more marked following Tofa + MTX treatment. This was likely due
to the inability of the second drug alone to act on the cells that
were activated later, and could suggest that sequential drug
treatments may themselves be influenced by timing and sequence.
Exceptions were observed for IL-8 and -2. IL-8
showed high levels of secretion at all times and did not undergo
any significant variation regardless of the treatment; this may be
due to the fact that this cytokine was predominantly produced by
other cells, rather than T lymphocytes, which did not undergo drug
treatments. Conversely, IL-2 concentration increased following Tofa
treatment, whether Tofa was administered as the first drug or
following MTX treatment. This may be due to positive feedback from
IL-2 signalling, when the signalling pathway was blocked by Tofa.
Notably, in the MIX condition, the trend of the effects on cytokine
secretion, observed in PHA-stimulated cells, was largely reproduced
at day 5. During the second time frame, certain differences between
PHA-stimulated cells and the MIX condition may have been apparent,
as the supernatant was collected from the culture wells and it
represents the cytokines that were produced by the total of the
cells; whereas, in the MIX condition, only a quarter of the cells
were pre-stimulated with PHA. It should be noted, however, that the
present study had certain limitations: Due to the experimental
design, a large amount of cells was required; therefore, buffy
coats from healthy donors were used, which showed a higher
variability in results than peripheral blood cells or cell lines.
Moreover, the standard deviation at day 9 was very large, which was
probably due to the long culture period.
Furthermore, the present study proposed an in
vitro model that was constructed to simulate rheumatic disease,
where only certain lymphocytes are activated (putative pathogenic
lymphocytes) while the majority remain unstimulated (17). In this model, the unstimulated
cells (NS-cells) were stained with CellTrace™ Violet, while
PHA-stimulated cells were stained with CellTrace™ Green and the two
populations were co-cultured. The staining with different tracers
allowed independent tracking of the fate of the directly stimulated
and unstimulated populations. The activation and proliferation of
NS-cells in the MIX condition was notable. These cells tended to
proliferate, which may have been due to bystander stimulation.
These cells were sensitive to the two drugs administered during the
first time frame, while drug treatment in the second time frame,
sequential or not, appeared to exert no effect; which was
consistent with the result of cytokine secretion in the
supernatant.
This particular issue must be considered further.
In vivo, certain lymphocytes in the same milieu of
pathogenic cells may undergo bystander activation, possibly
contributing to the spread of autoimmune activation and disease
progression. Indeed, T cells that are stimulated by a cytokine
cocktail induce the release of proinflammatory cytokines from
monocytes. A previous study identified that signalling pathways in
the responding monocytes were identical, regardless of whether the
monocytes were placed in contact with T cells isolated from
rheumatoid arthritis synovial tissue (18).
A previous study indicated that bystander cells
accumulate pathogenic potential with age (19), therefore, it is of particular
interest to control these cells, particularly in long-term
treatment strategies. Furthermore, therapy tends to modify the
course of rheumatic disease, possibly indicating a role of
selection due to pathogenic cell clones escaping from drug action;
although, to the best of our knowledge, the underlying mechanisms
remain to be fully demonstrated, it is likely that the escaping
clones are the most pathogenic ones and are able to spread the
immune defect (20,21). Thus, these considerations support
the current therapeutic strategies, which are based on different
approaches that combine drugs in consecutive use (such as step-up
and top-down).
In conclusion, the present study does not directly
correlate with the mechanisms of rheumatic disease in vivo;
however, does indicate that it is possible to obtain a simplified
in vitro model to evaluate the effect of sequential
treatment methods with various anti-rheumatic drugs. Furthermore,
this model allows the outcome of the primary activated cells (i.e
pathogenic lymphocytes) to be tracked, but also enables evaluation
of the fate of the cells that are bystander-activated by the
immunological milieu during active disease.
Acknowledgments
The present study was supported by a grant from the
Maternal and Child Institute 'Burlo Garofolo' (grant no. RC
28/2009).
Abbreviations:
|
MTX
|
methotrexate
|
|
Tofa
|
tofacitinib
|
|
DMARDs
|
disease modifying anti-rheumatic
drugs
|
|
PBMCs
|
peripheral blood mononuclear cells
|
References
|
1
|
McGonagle D, Aziz A, Dickie LJ and
McDermott MF: An integrated classification of pediatric
inflammatory diseases, based on the concepts of autoinflammation
and the immunological disease continuum. Pediatr Res. 65:38R–45R.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Uhlig T, Moe RH and Kvien TK: The burden
of disease in rheumatoid arthritis. Pharmacoeconomics. 32:841–851.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kremer JM, Bloom BJ, Breedveld FC, Coombs
JH, Fletcher MP, Gruben D, Krishnaswami S, Burgos-Vargas R,
Wilkinson B, Zerbini CA and Zwillich SH: The safety and efficacy of
a JAK inhibitor in patients with active rheumatoid arthritis:
Results of a double-blind, placebo-controlled phase IIa trial of
three dosage levels of CP-690,550 versus placebo. Arthritis Rheum.
60:1895–1905. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tanaka Y and Yamaoka K: JAK inhibitor
tofacitinib for treating rheumatoid arthritis: From basic to
clinical. Mod Rheumatol. 23:415–424. 2013. View Article : Google Scholar
|
|
5
|
Shi JG, Chen X, Lee F, Emm T, Scherle PA,
Lo Y, Punwani N, Williams WV and Yeleswaram S: The
pharmacokinetics, pharmacodynamics, and safety of baricitinib, an
oral JAK 1/2 inhibitor, in healthy volunteers. J Clin Pharmacol.
54:1354–1361. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Keystone EC, Taylor PC, Drescher E,
Schlichting DE, Beattie SD, Berclaz PY, Lee CH, Fidelus-Gort RK,
Luchi ME, Rooney TP, et al: Safety and efficacy of baricitinib at
24 weeks in patients with rheumatoid arthritis who have had an
inadequate response to methotrexate. Ann Rheum Dis. 74:333–340.
2015. View Article : Google Scholar
|
|
7
|
Wailoo A, Hernández MA, Scott IC, Ibrahim
F and Scott DL: Cost-effectiveness of treatment strategies using
combination disease-modifying anti-rheumatic drugs and
glucocorticoids in early rheumatoid arthritis. Rheumatology
(Oxford). 53:1773–1777. 2014. View Article : Google Scholar
|
|
8
|
Verschueren P, Esselens G and Westhovens
R: Daily practice effectiveness of a step-down treatment in
comparison with a tight step-up for early rheumatoid arthritis.
Rheumatology (Oxford). 47:59–64. 2008. View Article : Google Scholar
|
|
9
|
Lee YH, Bae SC and Song GG: Comparative
efficacy and safety of tofacitinib, with or without methotrexate,
in patients with active rheumatoid arthritis: A Bayesian network
meta-analysis of randomized controlled trials. Rheumatol Int.
35:1965–1974. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Piscianz E, Valencic E, Cuzzoni E, De
Iudicibus S, De Lorenzo E, Decorti G and Tommasini A: Fate of
lymphocytes after withdrawal of tofacitinib treatment. PLoS One.
9:e854632014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Piscianz E, Cuzzoni E, De Iudicibus S,
Valencic E, Decorti G and Tommasini A: Differential action of
3-hydroxyanthranilic acid on viability and activation of stimulated
lymphocytes. Int Immunopharmacol. 11:2242–2245. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lyons AB, Blake SJ and Doherty KV: Flow
cytometric analysis of cell division by dilution of CFSE and
related dyes. Curr Protoc Cytom Chapter. 9:Unit9.11. 2013.
View Article : Google Scholar
|
|
13
|
O'Dell JR, Mikuls TR, Taylor TH, et al:
CSP 551 RACAT Investigators: Therapies for active rheumatoid
arthritis after methotrexate failure. N Engl J Med. 369:307–318.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lee EB, Fleischmann R, Hall S, Wilkinson
B, Bradley JD, Gruben D, Koncz T, Krishnaswami S, Wallenstein GV,
Zang C, et al: Tofacitinib versus methotrexate in rheumatoid
arthritis. N Engl J Med. 370:2377–2386. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Burmester GR, Blanco R, Charles-Schoeman
C, Wollenhaupt J, Zerbini C, Benda B, Gruben D, Wallenstein G,
Krishnaswami S, Zwillich SH, et al: Tofacitinib (CP-690,550) in
combination with methotrexate in patients with active rheumatoid
arthritis with an inadequate response to tumour necrosis factor
inhibitors: A randomised phase 3 trial. Lancet. 381:451–460. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Maeshima K, Yamaoka K, Kubo S, Nakano K,
Iwata S, Saito K, Ohishi M, Miyahara H, Tanaka S, Ishii K, et al:
The JAK inhibitor tofacitinib regulates synovitis through
inhibition of interferon-γ and interleukin-17 production by human
CD4+ T cells. Arthritis Rheum. 64:1790–1798. 2012. View Article : Google Scholar
|
|
17
|
Cope AP, Schulze-Koops H and Aringer M:
The central role of T cells in rheumatoid arthritis. Clin Exp
Rheumatol. 25(5 Suppl 46): S4–S11. 2007.PubMed/NCBI
|
|
18
|
Brennan FM, Smith NM, Owen S, Li C, Amjadi
P, Green P, Andersson A, Palfreeman AC, Hillyer P, Foey A, et al:
Resting CD4+ effector memory T cells are precursors of
bystander-activated effectors: A surrogate model of rheumatoid
arthritis synovial T-cell function. Arthritis Res Ther. 10:R362008.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kobayashi M, Yasui N, Ishimaru N, Arakaki
R and Hayashi Y: Development of autoimmune arthritis with aging via
bystander T cell activation in the mouse model of Sjögren's
syndrome. Arthritis Rheum. 50:3974–3984. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kobari Y, Misaki Y, Setoguchi K, Zhao W,
Komagata Y, Kawahata K, Iwakura Y and Yamamoto K: T cells
accumulating in the inflamed joints of a spontaneous murine model
of rheumatoid arthritis become restricted to common clonotypes
during disease progression. Int Immunol. 16:131–138. 2004.
View Article : Google Scholar
|
|
21
|
VanderBorght A, Geusens P, Raus J and
Stinissen P: The autoimmune pathogenesis of rheumatoid arthritis:
Role of autoreactive T cells and new immunotherapies. Semin
Arthritis Rheum. 31:160–175. 2001. View Article : Google Scholar : PubMed/NCBI
|