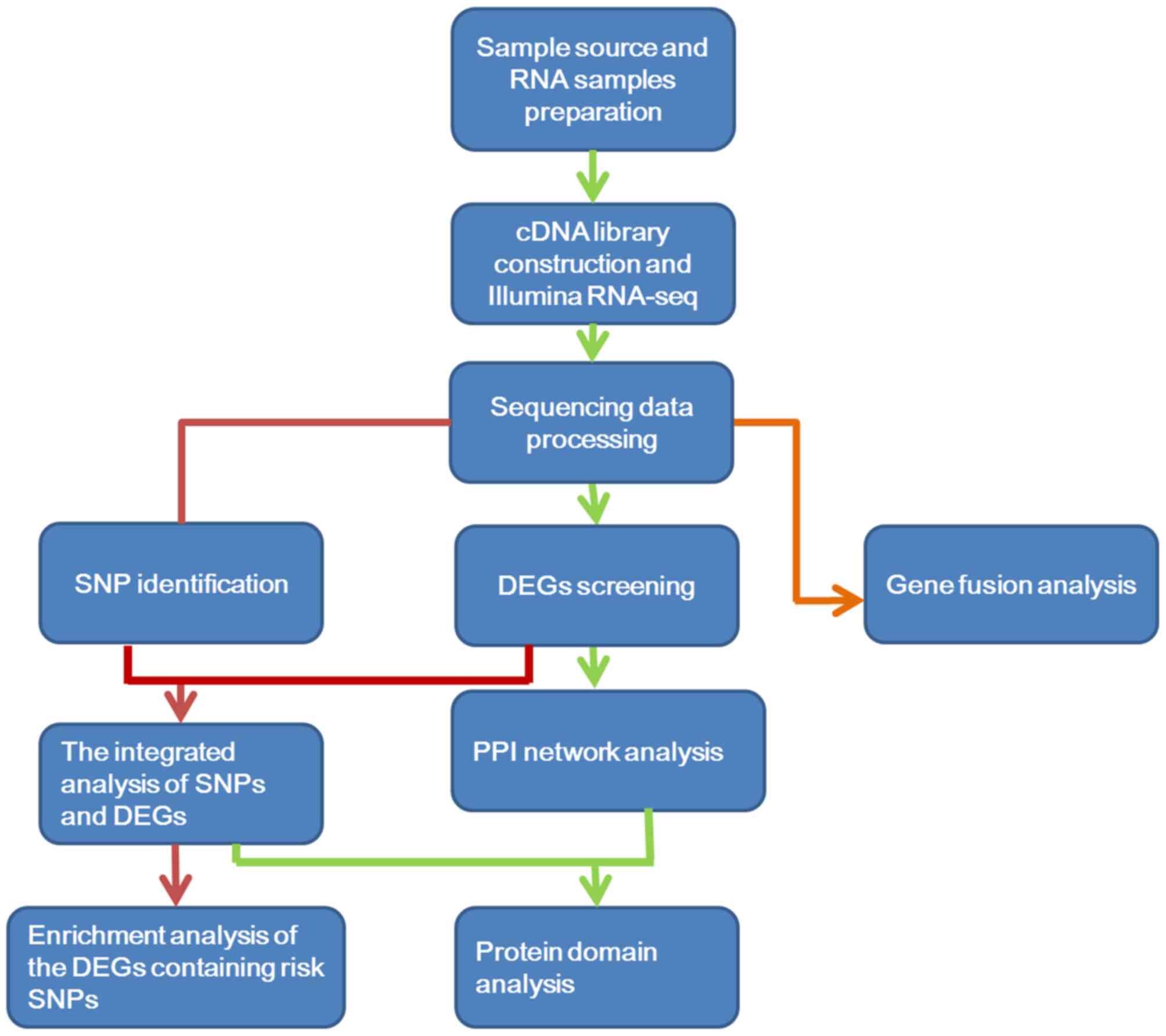
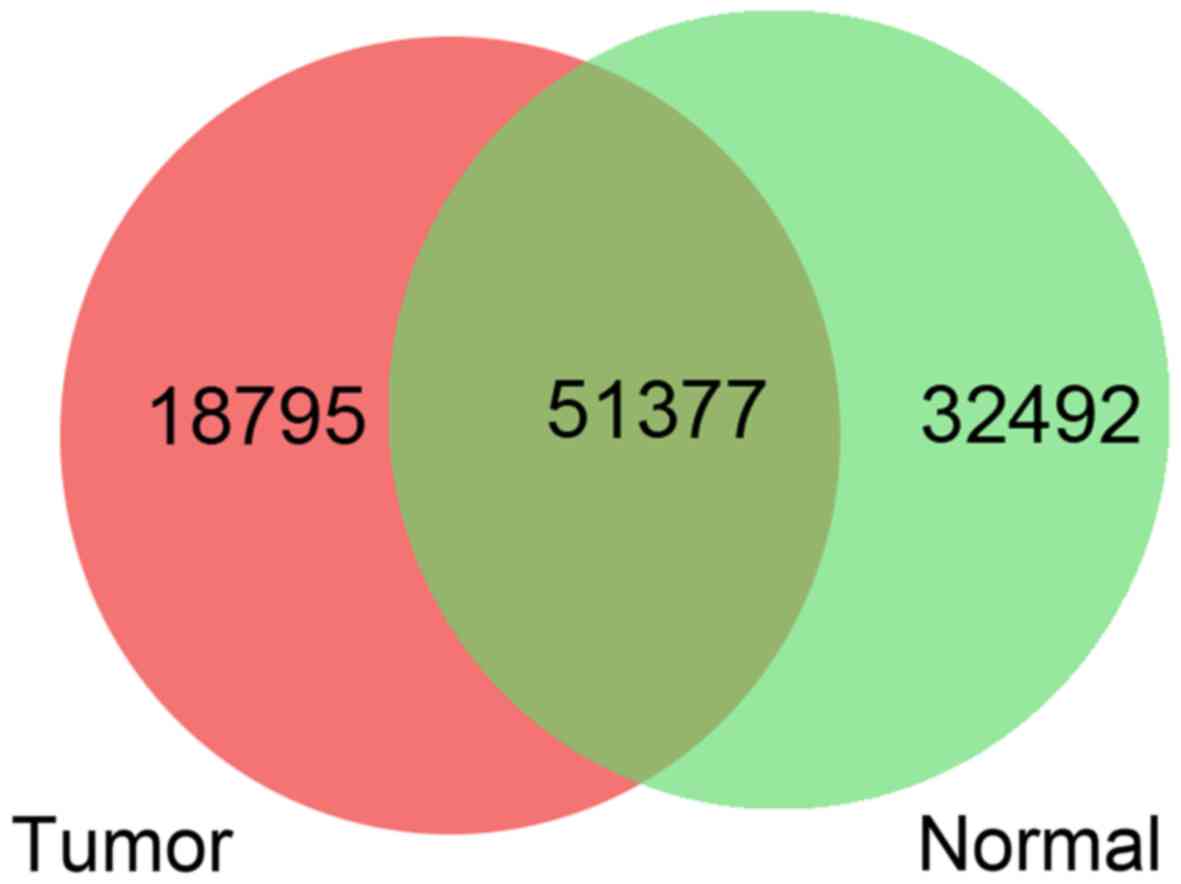
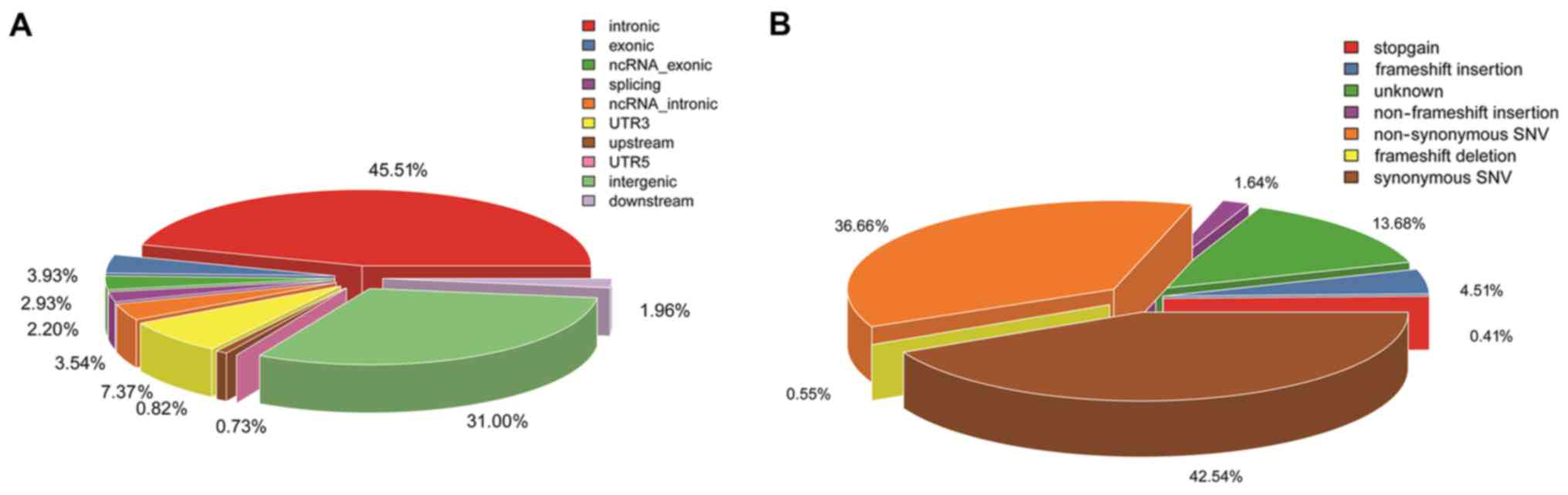
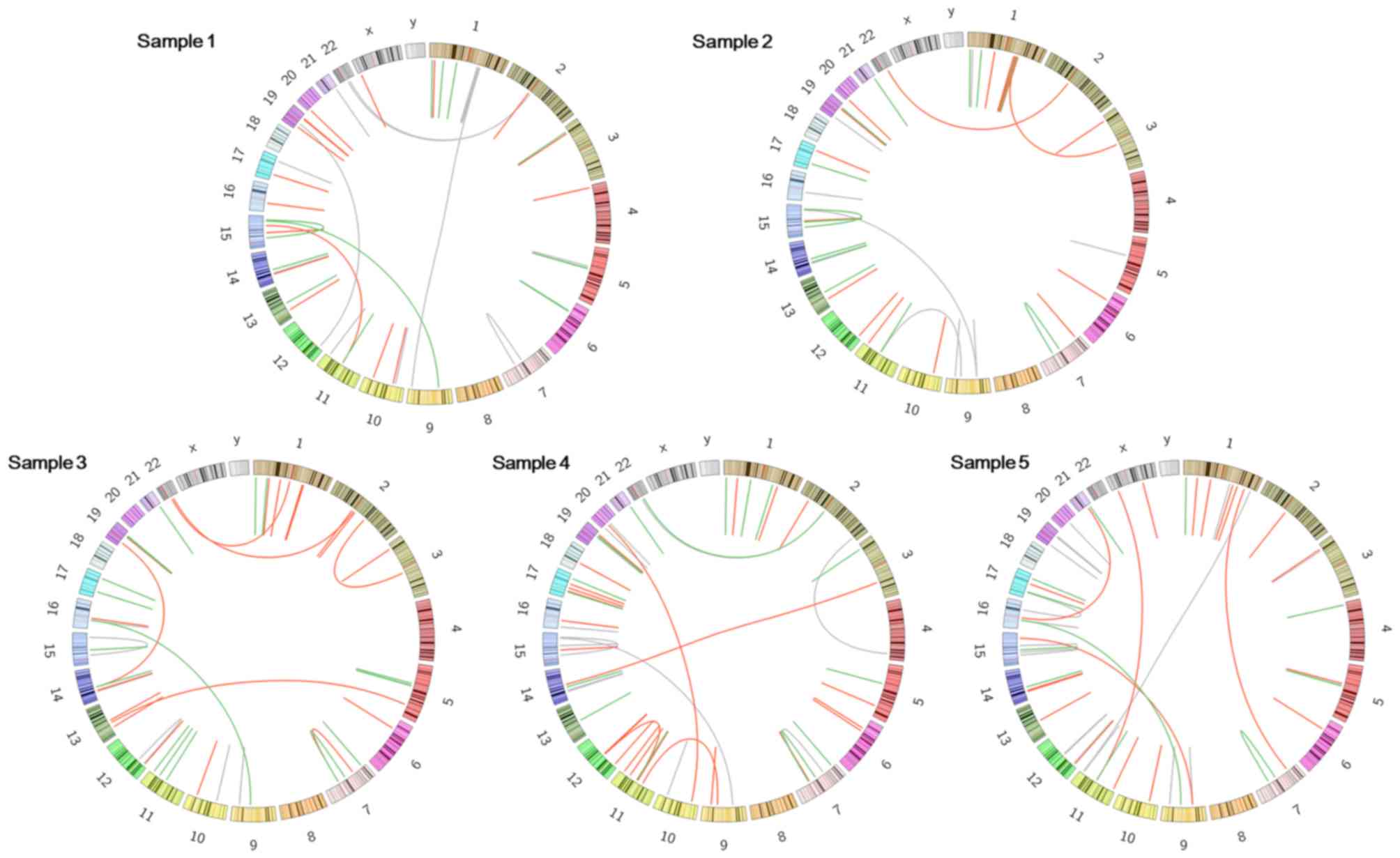
| GO_BP |
GO:0001655~urogenital system
development | 6 | ACE, AR, BMP2,
PGF, WFS1, ZBTB16 | 1.38E-03 |
| GO_BP |
GO:0055114~oxidation reduction | 13 | ALDH6A1, SORD,
FOXRED2, OGDHL, AASS, CYP20A1, SOD3, IYD, ALDH1A1, ERO1LB, DIO2,
BCO2, WWOX | 2.83E-03 |
| GO_BP | GO:0031667~response
to nutrient levels | 7 | BMP2, GATM,
MAP1LC3A, PPARG, LIPG, ADIPOR2, GHR | 3.56E-03 |
| GO_BP | GO:0001822~kidney
development | 5 | ACE, BMP2, PGF,
WFS1, ZBTB16 | 5.75E-03 |
| GO_BP | GO:0043434~response
to peptide hormone stimulus | 6 | AR, EGR2, GATM,
PPARG, FOXO1, GHR | 5.91E-03 |
| GO_CC |
GO:0044421~extracellular region part | 18 | TG, MATN2, BMP2,
PODN, SORD, PGF, MMP15, CCL28, SOD3, TNFSF10, ACE, FBLN2, EMID1,
NUCB2, LIPG, ANGPTL1, MFAP4, GHR | 5.34E-03 |
| GO_CC | GO:0005794~Golgi
apparatus | 16 | TG, SLC39A14,
RAP1GAP, NCS1, ACACB, MAN1A1, SOD3, CSGALNACT1, AP1S3, SYNE1,
EMID1, NUCB2, NDRG2, GRB14, WWOX, SMPD3 | 1.14E-02 |
| GO_CC | GO:0005925~focal
adhesion | 5 | TNS3, PGM5,
PRUNE, LAYN, LPP | 1.35E-02 |
| GO_CC |
GO:0005924~cell-substrate adherens
junction | 5 | TNS3, PGM5,
PRUNE, LAYN, LPP | 1.53E-02 |
| GO_CC | GO:0031985~Golgi
cisterna | 3 | CSGALNACT1,
NUCB2, SMPD3 | 1.65E-02 |
| GO_MF | GO:0042562~hormone
binding | 4 | ALDH1A1, AR,
ADIPOR2, GHR | 7.23E-03 |
| GO_MF | GO:0005096~GTPase
activator activity | 7 | ALDH1A1, AGAP11,
RAP1GAP, TBC1D4, OPHN1, RGS16, STARD13 | 8.86E-03 |
| GO_MF | GO:0008289~lipid
binding | 10 | ALDH1A1,
ALDH6A1, AR, MAP1LC3A, SDPR, OSBPL1A, FAAH, PPARG, CYTH3,
PLEKHA2 | 1.06E-02 |
| GO_MF |
GO:0005543~phospholipid binding | 6 | MAP1LC3A, SDPR,
OSBPL1A, FAAH, CYTH3, PLEKHA2 | 1.44E-02 |
| GO_MF | GO:0050662~coenzyme
binding | 6 | ALDH6A1, ERO1LB,
SORD, OGDHL, FOXRED2, WWOX | 1.57E-02 |
| Pathway | hsa05200: Pathways
in cancer | 8 | AR, BMP2, PGF,
PPARG, FOXO1, ZBTB16, TCF7L1, DAPK1 | 1.28E-02 |
|
| B, Significant GO
and KEGG terms enriched for the downregulated genes containing risk
SNPs |
|
| Category | Term | Gene number | Gene symbol | P-value |
| GO_BP |
GO:0022604~regulation of cell
morphogenesis | 20 | PALM, LST1,
LZTS1, PTPRF, LIMK1, PLXNB2, CDH2, CDH4, TTL, TGFβ 2, NRCAM, FYN,
TIAM1,MYH14, CDC42EP3, ARAP1, RASA1, MYH10, CDC42EP5, FN1 | 4.41E-09 |
| GO_BP | GO:0007155~cell
adhesion | 48 | MPZL3, NRP2,
CLDN4, POSTN, CD151, CDSN, CTNNB1, NRCAM, PCDH1, WISP1,
CD44 | 2.56E-08 |
| GO_BP |
GO:0022610~biological adhesion | 48 | MPZL3, NRP2,
CLDN4, POSTN, CD151, CDSN, CTNNB1, NRCAM, PCDH1, WISP1,
CD44 | 2.63E-08 |
| GO_BP |
GO:0043062~extracellular structure
organization | 20 | MPZL3, MYO6,
ADAMTS14, LGALS3, SPOCK2, TNC, TGFβ R1, POSTN, CDH2, COL5A2,
CTNNB1, ANXA2, TGFβ2, DVL1, NRCAM, AGT, COL1A2, AGRN, COL1A1,
ADAMTS2 | 1.63E-07 |
| GO_BP | GO:0009611~response
to wounding | 36 | DCBLD2, TNC,
CLU, TLR2, CDH3, TNFRSF4, TGFβ2, CCL22, HRH1, NOD1, CD44, MAP3K1,
SERPINE1, ITGB6, CFI, BLNK, RAB27A, FN1, NOX4, CIITA, NFKBIZ, LYN,
OLR1, CFB, IL1RN, ANXA1, CHST2, NFAM1, CD55, BAX, VCAN, CTSB,
PARP4, PROS1, ACVR1, MYH10 | 2.51E-06 |
| GO_MF | GO:0005509~calcium
ion binding | 55 | PXDN, LTBP1,
S100A5, LDLR, ARSJ, EFHD2, PCDH1, GALNTL6, PADI2, ACTN1, NPTXR,
CAPN12 | 6.18E-07 |
| GO_MF | GO:0030695~GTPase
regulator activity | 31 | TBC1D9,
RASGEF1A, IQGAP1, PLEKHG2, RINL, TIAM1, RAPGEF5, RAP1GAP2, DOCK10,
RASA1, RHOBTB3, NGEF, ARHGEF2, ABR, TNIK, PSD3, DOCK9, SIPA1L2,
ARHGAP23, DOCK3, ANXA2, MAP4K4, CDC42BPG, RASGRF1, RGS4, RIN1,
SYTL2, SH3BP1, LRRK2, SYTL1, ARAP1 | 2.83E-06 |
| GO_MF |
GO:0060589~nucleoside-triphosphatase
regulator activity | 31 | TBC1D9,
RASGEF1A, IQGAP1, PLEKHG2, RINL, TIAM1, RAPGEF5, RAP1GAP2, DOCK10,
RASA1, RHOBTB3, NGEF, ARHGEF2, ABR, TNIK, PSD3, DOCK9, SIPA1L2,
ARHGAP23, DOCK3, ANXA2, MAP4K4, CDC42BPG, RASGRF1, RGS4, RIN1,
SYTL2, SH3BP1, LRRK2, SYTL1, ARAP1 | 4.39E-06 |
| GO_MF | GO:0019899~enzyme
binding | 34 | CDH2, IQGAP1,
CTNNB1, MAP3K1, SERPINE1, CDK5RAP2, FAS, DOCK10, INSR, RASA1,
CDC42EP5, RHOBTB3, FMNL2, ARHGEF2, LYN, TGFβR1, DOCK9, DOCK3, FLNA,
DVL1, ANXA2, NRIP1, RFC5, MICALCL, CCND1, MAST2, CCND3, CCND2,
IGF2R, SYTL2, PARP4, CACNA1C, SYTL1, ADAMTS4 | 2.76E-05 |
| GO_MF |
GO:0008092~cytoskeletal protein
binding | 32 | SHROOM4, FXYD5,
SDC4, KLHL2, NRCAM, FRMD3, CORO2A, FRMD5, MACF1, CDK5RAP2, MSN,
CDC42EP3, RAB27A, FMNL2, ARHGEF2, MYO6, HTT, MYO1E, MYO1D, MYO1G,
ACTN1, FLNA, ANXA2, FYN, ARPC5L, SPTBN2, FLII, MYH14, DST, FHOD1,
LCP1, MYH10 | 7.90E-05 |
| GO_CC | GO:0044459~plasma
membrane part | 119 | MICB, SRCIN1,
PTGS2, ATP1B3, GABRB2, TLR2, CTNNB1, NRCAM, ST3GAL5, CD44,
KCNK5 | 2.22E-11 |
| GO_CC | GO:0005886~plasma
membrane | 171 | MICB, ATP1B3,
PTGS2, SRCIN1, GABRB2, TLR2, IQGAP1, CTNNB1, NRCAM, MCOLN3 | 2.42E-10 |
| GO_CC |
GO:0031012~extracellular matrix | 34 | ADAMTS17, LTBP1,
ADAMTS14, SPOCK2, TNC, ADAMTSL5, POSTN, TGFβ2, TIMP1, LAMB3, CD44,
FBN3, AGRN, FBN2, FGF1, LOXL1, FN1, LGALS3, COL13A1, PAPLN, SPARC,
EMILIN2, COL5A2, ANXA2, ADAMTS9, BGN, NAV2, COL1A2, LAMC2, VCAN,
COL1A1, DST, ADAMTS2, ADAMTS4 | 3.48E-09 |
| GO_CC |
GO:0005578~proteinaceous extracellular
matrix | 32 | ADAMTS17, LTBP1,
ADAMTS14, SPOCK2, TNC, ADAMTSL5, POSTN, TIMP1, LAMB3, FBN3, AGRN,
FBN2, FGF1, LOXL1, FN1, LGALS3, COL13A1, PAPLN, SPARC, EMILIN2,
COL5A2, ANXA2, ADAMTS9, BGN, NAV2, COL1A2, LAMC2, VCAN, COL1A1,
DST, ADAMTS2, ADAMTS4 | 7.79E-09 |
| GO_CC | GO:0005887~integral
to plasma membrane | 71 | MICB, LDLR,
CLDN4, SLC20A1, ATP1B3, CORIN, GABRB2, TLR2, GABBR2, CD151,
SDC4 | 2.00E-08 |
| Pathway | hsa04514: Cell
adhesion molecules (CAMs) | 19 | CLDN16, ICAM1,
CLDN4, PTPRF, CDH2, HLA-B, CDH3, SDC4, CDH4, NRCAM, ITGA9, ITGB8,
CD58, CLDN1, HLA-DPA1, VCAN, CNTNAP1, HLA-DOA, HLA-DRA | 1.16E-06 |
| Pathway | hsa04512:
ECM-receptor interaction | 15 | TNC, ITGA11,
ITGA3, SDC4, COL5A2, ITGA9, LAMB3, CD44, ITGB8, ITGB6, COL1A2,
LAMC2, AGRN, COL1A1, FN1 | 1.68E-06 |
| Pathway | hsa04510: Focal
adhesion | 22 | TNC, MET,
ITGA11, ACTN1, ITGA3, COL5A2, FLNA, CTNNB1, ITGA9, LAMB3, CCND1,
CCND3, CCND2, ITGB8, RASGRF1, FYN, ITGB6, COL1A2, LAMC2, COL1A1,
SHC3, FN1 | 1.17E-05 |
| Pathway | hsa05412:
Arrhythmogenic right ventricular cardiomyopathy (ARVC) | 12 | ITGA9, DSG2,
PKP2, ITGB8, ITGB6, ITGA11, CACNB1, ACTN1, ITGA3, CDH2, CACNA1C,
CTNNB1 | 8.78E-05 |
| Pathway | hsa04115: p53
signaling pathway | 11 | CCNE2, CCND1,
TNFRSF10B, CCND3, CCND2, ZMAT3, BAX, SERPINE1, DDB2, FAS,
PERP |