Introduction
With improvements in living standards, increasing
attention is focused on intestinal health. Intestinal health is
defined as a state, in which there are no signs of gut disease, no
risk of intestinal disease and no intestinal diseases (1). The adhesion of intestinal flora to
the intestinal wall can have a beneficial or deleterious role in
hosts, and determines the health status of the gut (2). Therefore, investigations of
intestinal flora are essential for intestinal health. A previous
study showed that the diversity of the colonic mucosa was closely
associated with bacterial microflora in patients with active
inflammatory bowel disease (3).
Disturbances of microbiota may also induce immune dysregulation and
can develop into inflammatory bowel disease (4). In addition, gut bacteria can degrade
various xenobiotics, including halogenated aromatic compounds,
which may be beneficial for human health (5).
Various genes associated with intestinal flora have
been identified for investigating intestinal health. In the
microbial community, the diversity of genes can provide several
types of enzymes and pathways associated with intestinal health
(6). Jia et al (7) suggested that cytochrome P450, family
4, subfamily F, polypeptide 14 and tachykinin, precursor 1 interact
with each other and are involved in pathways, including the
neuropeptide signaling pathway, oxidation reduction and metabolism
by regulating intestinal microbiota depletion. In addition, FBJ
murine osteosarcoma viral oncogene homolog has been confirmed to
enhance gut health by altering the ecology of the gut microbiota
and improving the proteolysis of feces (8). Another important gene, interleukin 6
has been confirmed as a key molecule in gut barrier dysfunction
(9). Although several genes
associated with intestinal health have been investigated, further
information is required. Therefore, molecular mechanisms, including
critical genes, pathways and their interactions, require
investigation.
In the present study, microarray analysis was
performed for the screening of differentially expressed genes
(DEGs) between microbiota-depleted mice and control mice.
Subsequently, functional and pathway enrichment analyses of the
DEGs were processed. Finally, literature associated with the DEGs
was mined and their associations were analyzed. The results of the
present study may identify potential important genes for further
investigations on intestinal flora and human health.
Materials and methods
Data acquisition
The gene expression profiles of GSE22648 were
downloaded from the Gene Expression Omnibus database (www.ncbi.nlm.nih.gov/geo) with the platform
GPL6887 (Illumina MouseWG-6 v2.0 expression beadchip) (10). A total of 11 colon intestinal
epithelial cell samples were obtained from mice, including five
microbiota-depleted mice and six control mice, and analyzed.
Data preprocessing and DEG
screening
The Series Matrix File (ftp.ncbi.nlm.nih.gov/geo/series/GSE22nnn/GSE22648/matrix)
was downloaded to transfer probe names into gene symbols based on
the platform annotation information. Using the aggregate function
of the R statistical package, version 3.4.0 (stat.ethz.ch/R-manual/R-devel/library/stats/html/aggregate.html),
the mean value was considered to be the expression value of a gene
when multiple probes mapped to a single gene. For probes with
missing values, nearest neighbor averaging (11) in the impure package (12) of R was used for offset of the
margin value (k value was defaulted to 10). Quantile normalization
was applied for standardization based on the preprocessCore package
of R. The standard matrix was then obtained.
The DEGs between two groups were screened using the
Limma package (13) (bioconductor.org/packages/release/bioc/html/limma.html;
version 3.5) and the differences of mean expression values were
assessed using the unpaired t-test method. In addition, the
Benjamini-Hochberg algorithm (14)
was used for p-value correction. Finally, the thresholds of the
DEGs were log2 fold change |log2FC| >0.585
and adjusted p-value of P<0.05.
Functional and pathway enrichment of
DEGs
The online tool of the Database of Annotation,
Visualization and Integrated Discover (DAVID) (15) was used for Gene Ontology (GO;
www.geneontology.org) and Kyoto
Encyclopedia of Genes and Genomes (KEGG; www.genome.jp/kegg) pathway enrichment analysis of
DEGs. The cut-off criterion was P<0.05.
Mining of DEG-associated
literature
The DEG-associated literature was mined using the
GenCLip 2.0 online tool (ci.smu.edu.cn)
(16). The Gene Cluster with
Literature Profiles module generated statistically overrepresented
key words grouped by a fuzzy cluster algorithm to annotate the
input genes. The key words were generated based on the occurrence
frequencies of free terms in the gene-associated literature or were
provided by the user. The associations among the genes and keywords
were linked to the relevant MEDLINE abstracts in which the
co-occurrence of genes and keywords are highlighted. The Literature
Mining Gene Networks module was used to construct a gene-network of
the input genes and generate sub-networks based on the user defined
query terms. It also calculated the probability of the random
occurrence of the networks through random simulation, and provided
GO and pathway enrichment analyses of genes. Co-citation networks
of DEGs were constructed for the mining of ‘hot genes’ which were
observed to be repeatedly associated with the other genes. At the
same time, the DEGs were clustered to mine the functional
information of key genes in previous studies with thresholds of
P<0.0001 and hit ≥4.
Results
Data preprocessing and DEG
screening
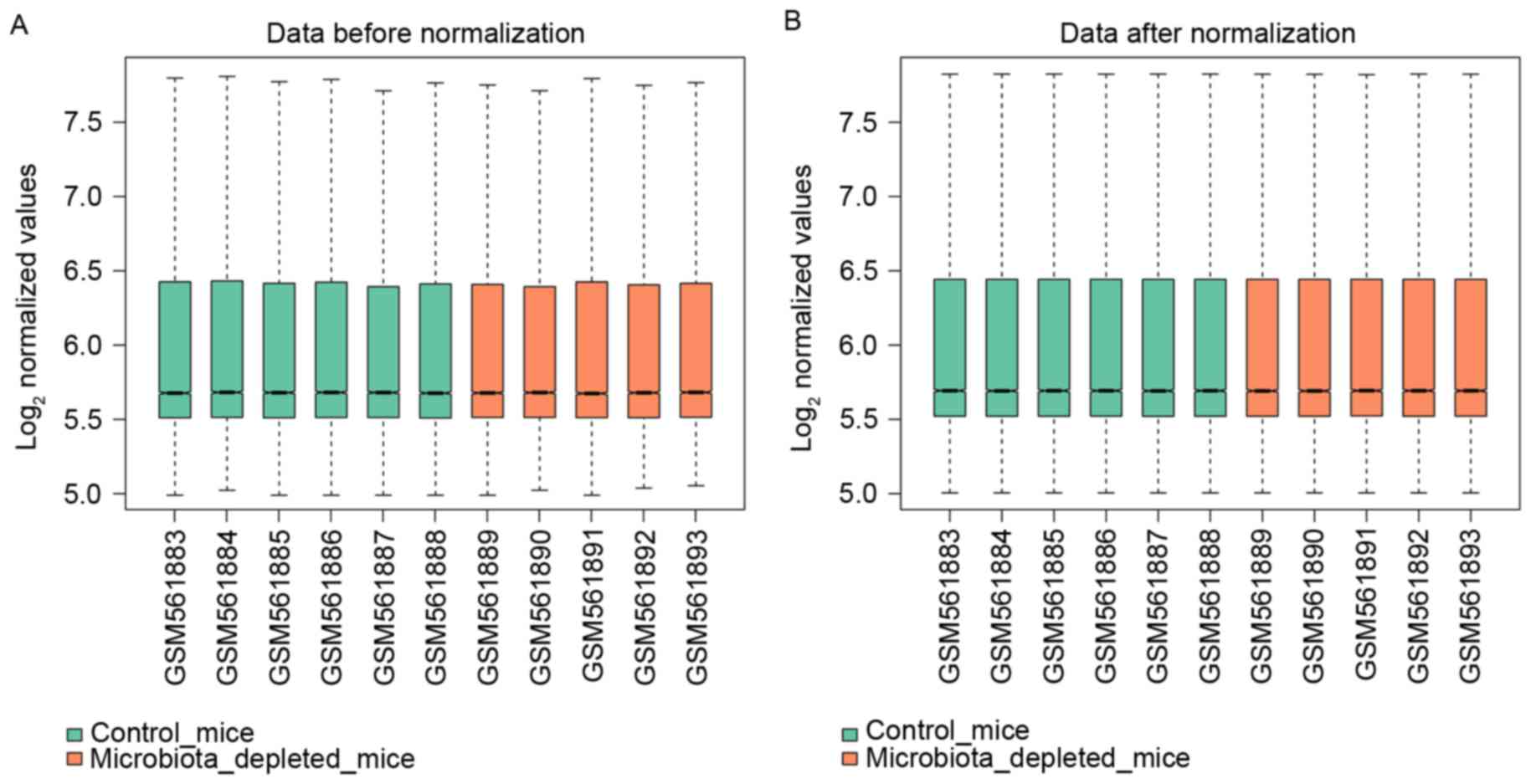
Following data preprocessing, a total of 30,865
expression data of transcripts were obtained from 11 samples. As
shown in Fig. 1, the median of
each sample was at the same level.
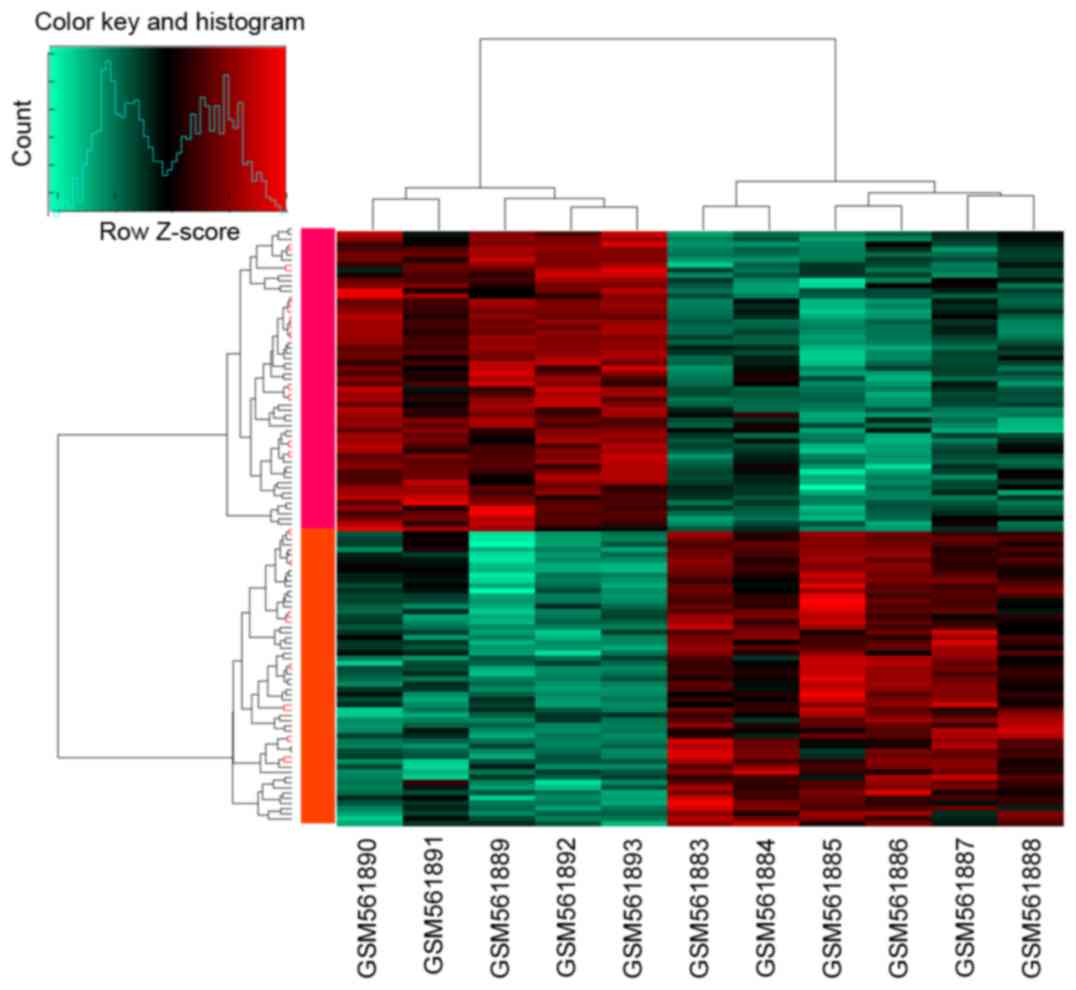
A total of 115 DEGs were screened with the
thresholds of |log2FC| >0.585 and adjusted p-value of
P<0.05, which included 58 upregulated genes and 57 downregulated
genes. The cluster heatmap of the DEGs is shown in Fig. 2.
GO functional and KEGG pathway
enrichment analysis
As shown in Table
I, the screened upregulated genes were significantly enriched
into various GO terms, including microsome, oxidation reduction and
heme binding. They were also enriched in the pathway of drug
metabolism. The 57 downregulated DEGs were enriched in different
functions, including DNA packaging, nucleosome assembly and
nucleosome. These downregulated DEGs were also enriched in
pathways, including arachidonic acid metabolism, systemic lupus
erythematosus and linoleic acid metabolism.
 | Table I.Top five GO and pathway enrichment
terms of differentially expressed genes. |
Table I.
Top five GO and pathway enrichment
terms of differentially expressed genes.
| Ontology | Term | Count | P-value |
|---|
| Upregulated |
|
|
|
| GO-CC |
GO:0005792~microsome | 6 | 5.59E-05 |
| GO-CC |
GO:0042598~vesicular fraction | 6 | 6.56E-05 |
| GO-CC | GO:0005624~membrane
fraction | 8 | 1.62E-04 |
| GO-CC |
GO:0005626~insoluble fraction | 8 | 2.01E-04 |
| GO-CC | GO:0000267~cell
fraction | 8 | 4.21E-04 |
| GO-BP |
GO:0055114~oxidation reduction | 8 | 7.98E-04 |
| GO-MF | GO:0020037~heme
binding | 7 | 8.59E-07 |
| GO-MF |
GO:0046906~tetrapyrrole binding | 7 | 1.14E-06 |
| GO-MF | GO:0009055~electron
carrier activity | 7 | 6.14E-06 |
| GO-MF |
GO:0070330~aromatase activity | 4 | 6.55E-05 |
| GO-MF | GO:0005506~iron ion
binding | 7 | 1.21E-04 |
| KEGG_PATHWAY | mmu00982:Drug
metabolism | 4 | 0.00128 |
| Downregulated |
|
|
|
| GO-BP | GO:0006323~DNA
packaging | 6 | 7.41E-06 |
| GO-BP |
GO:0006334~nucleosome assembly | 5 | 4.43E-05 |
| GO-BP |
GO:0031497~chromatin assembly | 5 | 4.93E-05 |
| GO-BP |
GO:0034728~nucleosome organization | 5 | 5.19E-05 |
| GO-BP |
GO:0065004~protein-DNA complex
assembly | 5 | 5.19E-05 |
| GO-CC |
GO:0000786~nucleosome | 5 | 3.96E-05 |
| GO-CC |
GO:0044427~chromosomal part | 8 | 4.92E-05 |
| GO-CC |
GO:0032993~protein-DNA complex | 5 | 8.38E-05 |
| GO-CC |
GO:0005694~chromosome | 8 | 1.46E-04 |
| GO-CC |
GO:0000785~chromatin | 6 | 1.51E-04 |
| KEGG_PATHWAY |
mmu00590:Arachidonic acid metabolism | 4 | 0.00171 |
| KEGG_PATHWAY | mmu05322:Systemic
lupus erythematosus | 4 | 0.00318 |
| KEGG_PATHWAY | mmu00591:Linoleic
acid metabolism | 3 | 0.00792 |
DEG-associated literature mining
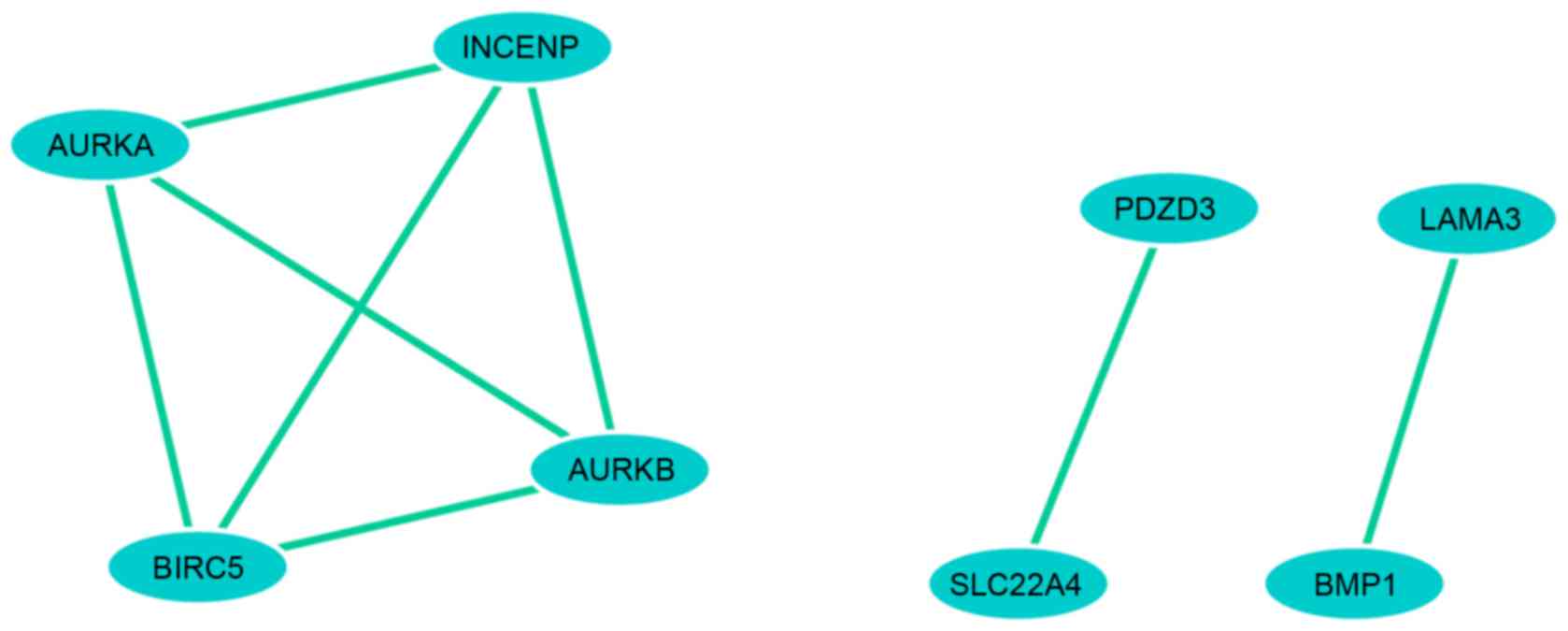
By mining the citation associations between the DEGs
in PubMed, at least eight genes were found to have pairwise
co-citation associations. In addition, four upregulated DEGs and
four downregulated DEGs were identified with high co-citation
levels (Fig. 3 and Table II). Therefore, four downregulated
DEGs, including baculoviral IAP repeat containing 5 (BIRC5),
aurora kinase A (AURKA), aurora kinase B (AURKB) and
inner centromere protein antigens 135/155 kDa (INCENP) were
co-cited in several studies. The four upregulated genes were
laminin α3, bone morphogenetic protein 1, solute carrier family 22
(organic cation/zwitterion transporter), member 4 (SLC22A4)
and PDZ domain containing 3.
| Table II.Results of literature-associated
mining of differentially expressed genes. |
Table II.
Results of literature-associated
mining of differentially expressed genes.
| Gene | Co-genes (n) | Co-cites (n) | Total (n) | log2
fold change |
|---|
| BIRC5 | 3 | 143 | 5,645 | −0.971 |
| AURKA | 3 | 114 |
990 | −1.054 |
| AURKB | 3 | 169 |
537 | −0.700 |
| INCENP | 3 | 127 |
247 | −0.710 |
| LAMA3 | 1 |
1 |
301 | 0.896 |
| BMP1 | 1 |
1 |
445 | 0.663 |
| SLC22A4 | 1 |
2 |
271 | 0.790 |
| PDZD3 | 1 |
2 |
27 | 0.715 |
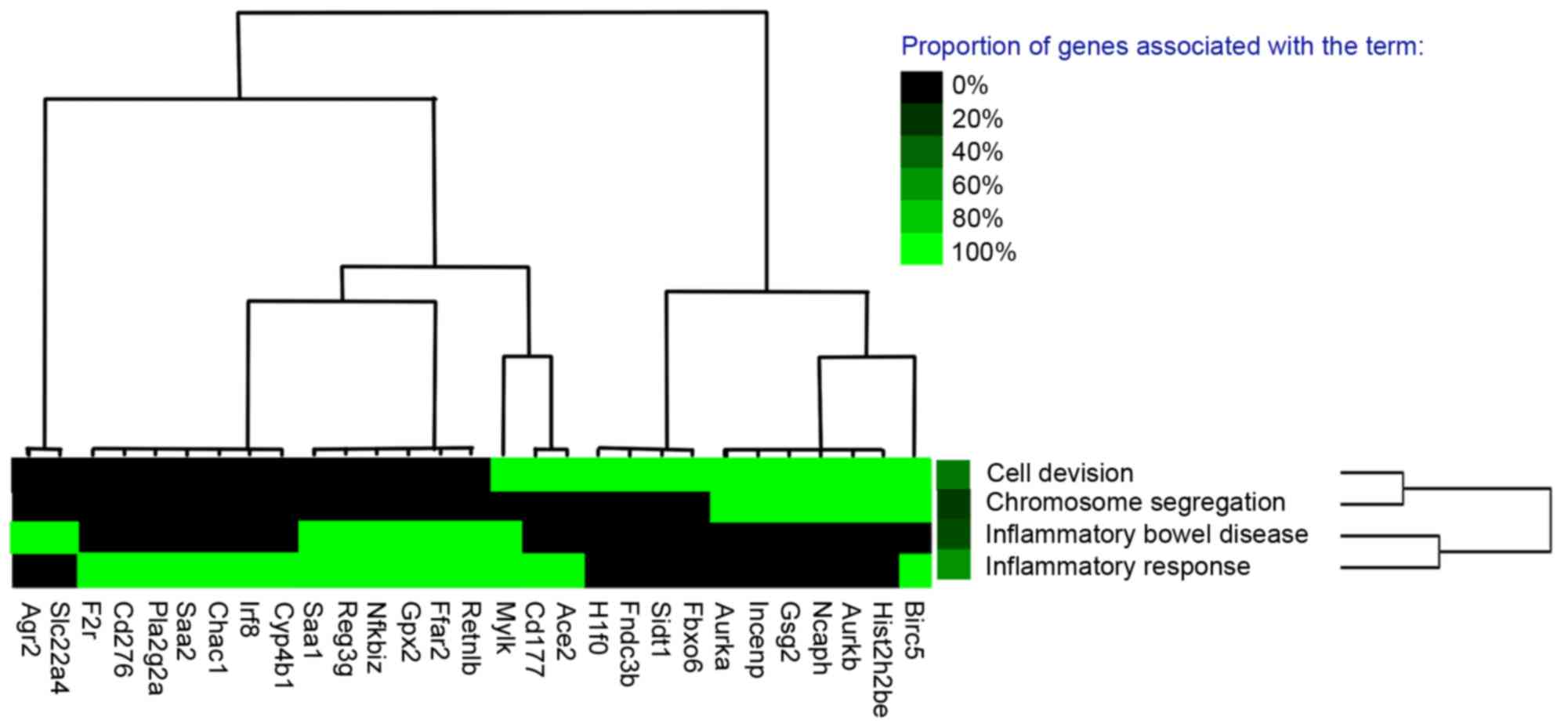
Gene cluster analysis with the literature profiles
of all DEGs was processed (Fig. 4
and Table III). The results
showed that a total of 19 genes, including BIRC5,
AURKB, angiotensin I converting enzyme 2 and free fatty acid
receptor 2, were enriched in four modules, including cell division,
chromosome segregation, inflammatory bowel disease and inflammatory
response. In addition, these modules were associated with various
functional terms, including inflammatory response, chromosome
condensation, chromosomal passenger complex and phospholipase
A2.
| Table III.Modules of literature-associated
differentially expressed genes. |
Table III.
Modules of literature-associated
differentially expressed genes.
| Key word | Hit | Q-value | Genes |
|---|
| #single1 Enrichment
Score: 6.21 |
|
|
|
| INFLAMMATORY
RESPONSE | 17 | 0.000223 | ACE2;
BIRC5; CD177; CD276; CHAC1;
CYP4B1; F2R; FFAR2; GPX2; IRF8;
MYLK; NFKBIZ; PLA2G2A; REG3G;
RETNLB; SAA1; SAA2 |
| #cluster1
Enrichment Score: 6.17 |
|
|
|
| CHROMOSOME
CONDENSATION | 6 | 0.000023 | AURKB;
BIRC5; H1F0; HIST2H2BE; INCENP;
NCAPH |
| CHROMOSOME
SEGREGATION | 7 | 0.000622 | AURKA;
AURKB; BIRC5; GSG2; HIST2H2BE;
INCENP; NCAPH |
| CELL DIVISION | 14 | 0.000568 | ACE2;
AURKA; AURKB; BIRC5; CD177;
FBXO6; FNDC3B; GSG2; H1F0;
HIST2H2BE; INCENP; MYLK; NCAPH;
SIDT1 |
| #cluster2
Enrichment Score: 5.66 |
|
|
|
| CHROMOSOMAL
PASSENGER COMPLEX | 5 | 0.000016 | AURKB;
BIRC5; GSG2; INCENP; NBL1 |
| SPINDLE ASSEMBLY
CHECKPOINT | 5 | 0.002422 | AURKA;
AURKB; BIRC5; GSG2; INCENP |
| SPINDLE
CHECKPOINT | 5 | 0.003230 | AURKA;
AURKB; BIRC5; GSG2; INCENP |
| #single2 Enrichment
Score: 5.42 |
|
|
|
| INFLAMMATORY BOWEL
DISEASE | 9 | 0.000590 | AGR2;
FFAR2; GPX2; MYLK; NFKBIZ;
REG3G; RETNLB; SAA1; SLC22A4 |
| #single3 Enrichment
Score: 4.73 |
|
|
|
| PHOSPHOLIPASE
A2 | 6 | 0.002282 | CD177;
MYLK; PLA2G2A; PLA2G3; SAA1;
SMPDL3B |
Discussion
Gut bacteria and their reactions are important in
gut health and general human health (17). Understanding the process and
potential mechanism of the effect of microbial intestinal
metabolism on human health is important for the treatment of
intestinal diseases (18). In the
present study, four upregulated DEGs and four downregulated DEGs
were identified with high co-citation levels. Among these genes,
the AURKA, INCENP, BIRC5 and AURKB
downregulated genes and SLC22A4 upregulated gene were
important DEGs, which were enriched in four modules, including cell
division, chromosome segregation, inflammatory bowel disease and
inflammatory response.
A previous study indicated that intestinal cell
proliferation is associated with epithelial homeostasis,
regeneration (19), and various
diseases, including obstructive jaundice (20) and cancer (4). In the present study, AURKA was
identified with a high number of citations on literature mining.
This gene encodes a protein, which is a cell cycle regulated
kinase, and is also closely associated with chromosome segregation
(21). Ratushny et al
(22) suggested that by combining
with AURKA, SRC kinase was activated and further affected
various biological processes, including tumor invasion. In
addition, AURKA locates at the spindle poles and regulates
cytokinesis by regulating centrosome maturation (23). In the colon of HNF4α mutant mice,
the expression of AURKA was significantly upregulated, and
HNF4α was a key regulator for ion transport in the gut mucosa.
Therefore, the findings of the present study suggested that
AURKA may be involved in the functional modules of
chromosome segregation and cell division, and may be important in
affecting intestinal microbiome homeostasis in intestinal
health.
AURKA is concentrated primarily at the spindle poles
in the period of meiosis (MI) and meiosis II, whereas AURKB
locates at kinetochores in the period of MI (24). As with AURKA, AURKB
was identified in the present study as being enriched in functions
of chromosome segregation and cell division. Fenton et al
(25) processed a microarray
analysis, and showed that leptin may upregulate the expression of
AURKB and further induce colon epithelial cell
proliferation. In addition, the expression of AURKB can be
regulated by microRNA (miR)-24 by recognizing complementary
sequences (26). In addition, in
irritable bowel syndrome, miR-24 has been confirmed to increase gut
permeability (27). Therefore,
AURKB may also be a target in the treatment of intestinal
disease.
INCENP is a complex partner of AURKB for kinase
activation, and the regulation of chromosome segregation and
cytokinesis (28). In the present
study, INCENP was screened to have a high number of
citations, and was enriched in cell division and chromosome
segregation. INCENP has also been confirmed to be an important
member of the chromosomal passenger complex, which is an essential
regulator for the completion of cell division (29). In addition, INCENP/Sli15
phosphorylation can activate Aurora B/IpI1, which is important for
the activity of the chromosomal passenger complex (30). The expression of INCENP has
also been shown to be increased in several types of cancer,
including colorectal cancer (31).
Therefore, the present study hypothesized that INCENP may
affect intestinal health through interacting with AURKB.
In the present study, BIRC5 was enriched in
three modules, including cell division, chromosome segregation and
inflammatory response. Neufert et al (32) confirmed that BIRC5 can limit
bacterial growth and is involved in mucosal wound healing, which
suggests a promising therapeutic approach for disturbed intestinal
homeostasis. In addition, activated signal transducer and activator
of transcription 3 can induce the transcription of BIRC5 and
regulate tissue defense during the process of gastrointestinal
infection. Previous studies have also shown that microbiota and
commensal bacteria can induce inflammatory response in
immunocompromised hosts (33), and
when the normal microbial community is disrupted, the risk of
inflammation is increased (34),
This offers novel insights into the pathogenesis of disease for
treating intestinal disorders. The gut microbiota can also affect
host metabolism and produce various pro-inflammatory molecules,
including lipopolysaccharide, assisting in the development of
treatments for metabolic diseases (35). Therefore, BIRC5 may be a
critical gene in the effects of microbiota on intestinal
health.
Of the upregulated DEGs identified in the present
study, SLC22A4 was found to have a high number of citations
and was enriched in the module of inflammatory bowel disease. A
mutation of SLC22A4, which encodes organic carnitine
transporter 1, has been confirmed to have a clinical role in gut
health (36). In addition,
Tokuhiro et al (37) showed
that the organic cation transporter SLC22A4 functioned as a
transporter in the inflammatory milieu. A previous study also
confirmed that polymorphisms of SLC22A4 were closely
associated with early inflammatory bowel disease (38). The development of the gut
microbiota is essential for promoting the gut immune system and
maintaining system balance (39).
Therefore, SLC22A4 may be an essential gene for intestinal
health via modulating the immune system.
In conclusion, the present study performed
bioinformatics analysis of the effects of gut flora on molecular
changes in intestinal epithelial cells. AURKA,
INCENP, BIRC5, AURKB and SLC22A4 were
identified as potential target genes for intestinal disease
treatment through their involvement in various functions, including
cell division, chromosome segregation, inflammatory bowel disease
and inflammatory response. These results indicate novel therapeutic
targets for diseases caused by disturbed intestinal homeostasis.
However, there were limitations to the present study; gut
microbiota are associated with the delivery mode and the type of
feeding in infants, and changes throughout life between infancy and
elderly. Therefore, further experiments are required to confirm the
results obtained in the present study.
References
|
1
|
Abreu MT, Fukata M and Arditi M: TLR
signaling in the gut in health and disease. J Immunol.
174:4453–4460. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jeurissen SH, Lewis F, van der Klis JD,
Mroz Z, Rebel JM and Ter Huurne AA: Parameters and techniques to
determine intestinal health of poultry as constituted by immunity,
integrity and functionality. Curr Issues Intest Microbiol. 3:1–14.
2002.PubMed/NCBI
|
|
3
|
Ott SJ, Musfeldt M, Wenderoth DF, Hampe J,
Brant O, Fölsch UR, Timmis KN and Schreiber S: Reduction in
diversity of the colonic mucosa associated bacterial microflora in
patients with active inflammatory bowel disease. Gut. 53:685–693.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Round JL and Mazmanian SK: The gut
microbiota shapes intestinal immune responses during health and
disease. Nat Rev Immunol. 9:313–323. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Qin J, Li R, Raes J, Arumugam M, Burgdorf
KS, Manichanh C, Nielsen T, Pons N, Levenez F, Yamada T, et al: A
human gut microbial gene catalogue established by metagenomic
sequencing. Nature. 464:59–65. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Guarner F and Malagelada JR: Gut flora in
health and disease. Lancet. 361:512–519. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jia ZY, Xia Y, Tong D, Yao J, Chen HQ and
Yang J: Module-based functional pathway enrichment analysis of a
protein-protein interaction network to study the effects of
intestinal microbiota depletion in mice. Mol Med Rep. 9:2205–2212.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Smith AG, O'Doherty JV, Reilly P, Ryan MT,
Bahar B and Sweeney T: The effects of laminarin derived from
Laminaria digitata on measurements of gut health: Selected
bacterial populations, intestinal fermentation, mucin gene
expression and cytokine gene expression in the pig. Br J Nutr.
105:669–677. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Song M and Kellum JA: Interleukin-6. Crit
Care Med. 33 12 Suppl:S463–S465. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Reikvam DH, Erofeev A, Sandvik A, Grcic V,
Jahnsen FL, Gaustad P, McCoy KD, Macpherson AJ, Meza-Zepeda LA and
Johansen FE: Depletion of murine intestinal microbiota: Effects on
gut mucosa and epithelial gene expression. PLoS One. 6:e179962011.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Altman NS: An introduction to kernel and
nearest-neighbor nonparametric regression. The American
Statistician. 46:175–185. 1992. View Article : Google Scholar
|
|
12
|
Hastie T, Tibshirani R, Narasimhan B and
Gilbert C: Impute: Imputation for microarray data. Bioinformatics.
17:520–525. 2001.PubMed/NCBI
|
|
13
|
Shabalin AA, Tjelmeland H, Fan C, Perou CM
and Nobel AB: Merging two gene-expression studies via
cross-platform normalization. Bioinformatics. 24:1154–1160. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Rudy J and Valafar F: Empirical comparison
of cross-platform normalization methods for gene expression data.
BMC Bioinformatics. 12:4672011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Leek JT, Johnson WE, Parker HS, Fertig EJ,
Jaffe AE, Zhang Y, Torres LC and Storey JD: sva: Surrogate Variable
Analysis. R package version 3. 2013.
|
|
16
|
Wang JH, Zhao LF, Lin P, Su XR, Chen SJ,
Huang LQ, Wang HF, Zhang H, Hu ZF, Yao KT and Huang ZX: GenCLiP
2.0: A web server for functional clustering of genes and
construction of molecular networks based on free terms.
Bioinformatics. 30:2534–2536. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Puupponen-Pimiä R, Aura AM,
Oksman-Caldentey KM, Myllärinen P, Saarela M, Mattila-Sandholm T
and Poutanen K: Development of functional ingredients for gut
health. Trends Food Sci Technol. 13:3–11. 2002. View Article : Google Scholar
|
|
18
|
Pérez-Cobas AE, Gosalbes MJ, Friedrichs A,
Knecht H, Artacho A, Eismann K, Otto W, Rojo D, Bargiela R, von
Bergen M, et al: Gut microbiota disturbance during antibiotic
therapy: A multi-omic approach. Gut. 62:1591–1601. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Barker N: Adult intestinal stem cells:
Critical drivers of epithelial homeostasis and regeneration. Nat
Rev Mol Cell Biol. 15:19–33. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Assimakopoulos SF, Tsamandas AC, Louvros
E, Vagianos CE, Nikolopoulou VN, Thomopoulos KC, Charonis A and
Scopa CD: Intestinal epithelial cell proliferation, apoptosis and
expression of tight junction proteins in patients with obstructive
jaundice. Eur J Clin Invest. 41:117–125. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ding J, Swain JE and Smith GD: Aurora
kinase-A regulates microtubule organizing center (MTOC)
localization, chromosome dynamics, and histone-H3 phosphorylation
in mouse oocytes. Mol Reprod Dev. 78:80–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ratushny V, Pathak HB, Beeharry N,
Tikhmyanova N, Xiao F, Li T, Litwin S, Connolly DC, Yen TJ, Weiner
LM, et al: Dual inhibition of SRC and Aurora kinases induces
postmitotic attachment defects and cell death. Oncogene.
31:1217–1227. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Nikonova AS, Astsaturov I, Serebriiskii
IG, Dunbrack Jr RL and Golemis EA: Aurora A kinase (AURKA) in
normal and pathological cell division. Cell Mol Life Sci.
70:661–687. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shuda K, Schindler K, Ma J, Schultz RM and
Donovan PJ: Aurora kinase B modulates chromosome alignment in mouse
oocytes. Mol Reprod Dev. 76:1094–1105. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Fenton JI, Lavigne JA, Perkins SN, Liu H,
Chandramouli GV, Shih JH, Hord NG and Hursting SD: Microarray
analysis reveals that leptin induces autocrine/paracrine cascades
to promote survival and proliferation of colon epithelial cells in
an Apc genotype-dependent fashion. Mol carcinog. 47:9–21. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Felekkis K, Touvana E, Stefanou C and
Deltas C: microRNAs: A newly described class of encoded molecules
that play a role in health and disease. Hippokratia. 14:236–240.
2010.PubMed/NCBI
|
|
27
|
Vicario M, Martínez C and Santos J: Role
of microRNA in IBS with increased gut permeability. Gut.
59:710–712. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Honda R, Körner R and Nigg EA: Exploring
the functional interactions between Aurora B, INCENP, and survivin
in mitosis. Mol Biol Cell. 14:3325–3341. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Guta FF, O'Tooleb P and Walsha MA: MS04
P01 Structural and functional studies of the probiotic organism
Lactobacillus salivarius Mario Bumanna, Heinz. Acta Cryst.
63:s1272007. View Article : Google Scholar
|
|
30
|
Yang J, Zappacosta F, Annan RS, Nurse K,
Tummino PJ, Copeland RA and Lai Z: The catalytic role of INCENP in
Aurora B activation and the kinetic mechanism of Aurora B/INCENP.
Biochem. J. 417:355–360. 2009.
|
|
31
|
Chang JL, Chen TH, Wang CF, Chiang YH,
Huang YL, Wong FH, Chou CK and Chen CM: Borealin/Dasra B is a cell
cycle-regulated chromosomal passenger protein and its nuclear
accumulation is linked to poor prognosis for human gastric cancer.
Exp Cell Res. 312:962–973. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Neufert C, Pickert G, Zheng Y, Wittkopf N,
Warntjen M, Nikolaev A, Ouyang W, Neurath MF and Becker C:
Activation of epithelial STAT3 regulates intestinal homeostasis.
Cell Cycle. 9:652–655. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hooper LV, Littman DR and Macpherson AJ:
Interactions between the microbiota and the immune system. Science.
336:1268–1273. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kamada N, Seo SU, Chen GY and Núñez G:
Role of the gut microbiota in immunity and inflammatory disease.
Nat Rev Immunol. 13:321–335. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tremaroli V and Bäckhed F: Functional
interactions between the gut microbiota and host metabolism.
Nature. 489:242–249. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Shekhawat PS, Sonne S, Carter AL, Matern D
and Ganapathy V: Enzymes involved in L-carnitine biosynthesis are
expressed by small intestinal enterocytes in mice: Implications for
gut health. J Crohns Colitis. 7:e197–e205. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tokuhiro S, Yamada R, Chang X, Suzuki A,
Kochi Y, Sawada T, Suzuki M, Nagasaki M, Ohtsuki M, Ono M, et al:
An intronic SNP in a RUNX1 binding site of SLC22A4, encoding an
organic cation transporter, is associated with rheumatoid
arthritis. Nat Genet. 35:341–348. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ferraris A, Torres B, Knafelz D, Barabino
A, Lionetti P, de Angelis GL, Iacono G, Papadatou B, D'Amato G, Di
Ciommo V, et al: Relationship between CARD15, SLC22A4/5 and DLG5
polymorphisms and early-onset inflammatory bowel diseases: An
Italian multicentric study. Inflamm Bowel Dis. 12:355–361. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kelly D, King T and Aminov R: Importance
of microbial colonization of the gut in early life to the
development of immunity. Mutat Res. 622:58–69. 2007. View Article : Google Scholar : PubMed/NCBI
|