Introduction
According to the Report on the Status of Nutrition
and Chronic Diseases of Chinese residents (2015) (1), the prevalence of diabetes mellitus
(DM) reached up to 9.7% in 2012, which resulted in a heavy social
and economic burden on individuals, families and the country
(2). Therefore, research regarding
the pathogenesis, prevention and therapy of DM is of primary
concern. Substantial evidence implicates that obesity is a key risk
factor for type 2 DM (T2DM) (3).
The root etiology of obesity is chronic energy imbalance, which
produces adipocyte hypertrophy and hyperplasia, and these processes
lead to adipocyte dysfunction, induce insulin resistance and result
in T2DM (4,5). Accordingly, exploring the
characteristics of adipocytes may help the treatment of obesity and
T2DM.
The Hippo-yes associated protein (YAP) signaling
pathway has been identified to serve crucial roles in the
regulation of cell proliferation, cell apoptosis and cell
differentiation, and may modulate development, organ size, tissue
homeostasis and tumorigenesis (6,7). The
core of the mammalian Hippo-YAP pathway has been well established.
In general, upon activation, the extracellular signals are
transduced to the mammalian ste20 kinase 1/2 (MST1/2), then, the
complex is assembled by MST1/2 and phosphorylates salvador homolog
1 (Sav1), which activates S/T protein kinase large tumor suppressor
1/2 (LATS1/2). Activated LATS1/2 in complex with Mob phosphorylates
and inhibits two homologous transcription coactivators, namely, YAP
and its paralog transcriptional co-activator with PDZ-binding motif
(TAZ). Finally, YAP/TAZ regulates the expression of a large number
of genes important for cell proliferation (8). Ankyrin repeat domain 1 (ANKRD1),
connective tissue growth factor (CTGF) and cysteine rich angiogenic
inducer 61 (Cyr61) are well-characterized YAP target genes that
regulate cell proliferation (9).
Several studies have indicated that the Hippo-YAP signaling pathway
is involved in adipogenesis (10–12).
At the early phase of adipogenic differentiation, adipogenesis is
governed by the activity of a series of key transcription factors,
especially peroxisome proliferator-activated receptor γ (PPARγ) and
CCAAT/enhancer binding protein α (C/EBPα), which coordinates the
expression of adipogenic genes characteristic of terminally
differentiated adipocytes, including adipocyte protein 2 (aP2)
(13).
Liraglutide is a glucagon-like peptide-1 (GLP-1)
analogue widely used for the therapy of T2DM, and it is also
recommended for the treatment of obesity (14). As an insulinotropic hormone, GLP-1
is secreted by intestinal L cells in response to nutrient
ingestion, and acts on the GLP-1 receptor to facilitate
glucose-dependent insulin secretion, decreases glucagon levels and
improves β-cell neogenesis (14).
GLP-1 and liraglutide have previously been reported to regulate
adipocyte formation via extracellular signal-regulated kinase,
protein kinase C, protein kinase B and Wnt signaling pathways
(15–17). However, it is not clear whether
other signaling pathways are involved in the regulation of
adipogenic differentiation by liraglutide.
The present study investigated the effects and
possible underlying mechanisms of liraglutide on preadipocyte
3T3-L1 cell adipogenesis. These findings may contribute to the
development of novel drugs to aid weight loss and increase insulin
sensitivity.
Materials and methods
Reagents
Liraglutide was purchased from Novo Nordisk (West
Sussex, UK). Anti-β-actin (cat. no. A1978-100UL) for the western
blotting was purchased from Sigma-Aldrich; Merck KGaA (Darmstadt,
Germany). Anti-PPARγ (cat. no. 2435), anti-C/EBPα (cat. no. 2295),
anti-aP2 (cat. no. 3544), anti-YAP (cat. no. 14074),
anti-phosphorylated (p)-YAP (cat. no. 46931), anti-LATS1 (cat. no.
9153) and anti-MST1 (cat. no. 3682) for western blotting were all
purchased from Cell Signaling Technology, Inc. (Danvers, MA, USA).
Anti-Ki67 (cat. no. 27309-1-AP) for the immunofluorescence assay
was purchased from ProteinTech Group, Inc. (Chicago, IL, USA).
Cell lines and cell culture
Preadipocyte 3T3-L1 cells were cultured in
Dulbecco's modified Eagle's medium (HyClone; GE Healthcare Life
Sciences, Logan, UT, USA) supplemented with 10% fetal bovine serum
(FBS), 200 U/ml penicillin and 200 U/ml streptomycin, and were
maintained in a humidified incubator containing 5% CO2
at 37°C.
Cell differentiation and
treatment
For adipocyte differentiation, 3T3-L1 cells that
reached ~90% confluence were cultured in an adipocyte-inducing
medium (AIM; α-MEM containing 10% FBS, 0.5 µM dexamethasone, 0.25
mM methylisobutylxanthine, 5 µg/ml of insulin and 50 µM
indomethacin) in a humidified atmosphere at 37°C with 5%
CO2 for 3 days. For liraglutide treatment, cells were
cultured in AIM with 0 nM (vehicle), 10, 100, or 1,000 nM
liraglutide.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc., Waltham, MA, USA). A total of 0.5 µg RNA of each sample was
reverse-transcribed into cDNA with a high-capacity cDNA reverse
transcription kit (Thermo Fisher Scientific, Inc.) with the
following thermocycling parameters: 25°C for 10 min, 37°C for 2 h
and 85°C for 5 min. qPCR was performed using SGExcel Fast SYBR
Mixture kits (Sangon Biotech Co., Ltd., Shanghai, China) on the
Light Cycler® 96 Real-Time PCR system (Roche Diagnostics
GmbH, Mannheim, Germany), and the thermocycling parameters were
followed as previously described (18), which consisted of 40 cycles (95°C
for 10 sec, 60°C for 10 sec and 72°C for 10 sec) following an
initial denaturation step (95°C for 2 min). The mRNA expression
level of non-POU-domain-containing, octamer binding protein (NONO)
was used as an internal control as previously reported (19). Relative expression was calculated
using the comparative threshold cycle method (2−ΔΔCq)
(20). The primer sequences used
in the study were as follows: C/EBPα forward,
5′-CTGATTCTTGCCAAACTGAG-3′ and reverse,
5′-GAGGAAGCTAAGACCCACTAC-3′; PPARγ forward,
5′-CTTGACAGGAAAGACAACGG-3′ and reverse, 5′-GCTTCTACGGATCGAAACTG-3′;
aP2 forward, 5′-AAATCACCGCAGACGACAGG-3′ and reverse,
5′-GGCTCATGCCCTTTCATAAAC-3′; ANKRD1 forward,
5′-CTTGATGACCTTCGGTGCG-3′ and reverse, 5′-GCTCTTCTGTTGGGAAATGCT-3′;
CTGF forward, 5′-TCTCCACCCGAGTTACCAATG-3′ and reverse,
5′-TGCAGCCAGAAAGCTCAAAC-3′; Cry61 forward,
5′-GGAAAAGGCAGCTCACTGAAG-3′ and reverse,
5′-GCACTCTGGGTTGTCATTGGTA-3′; MST1 forward,
5′-TCCCTCAGGATGGAGACTATGA-3′ and reverse, 5′-AAGGCTGGGCTGGTGTTG-3′;
NONO forward, 5′-TGCTCCTGTGCCACCTGGT-3′ and reverse,
5′-CCGGAGCTGGACGGTTGAAT-3′.
Western blotting
The cells were incubated in AIM or treated with
different concentrations of liraglutide (0, 10, 100 or 1,000 nM)
for 3 days. Following this, treated cells were lysed with
radioimmunoprecipitation assay lysis buffer (Beyotime Institute of
Biotechnology, Haimen, China) and protein concentration was
determined with bicinchoninic acid protein assay. Immunoblotting
was performed as previously described (21). Briefly, protein (20 µg/lane) was
separated on a 12% SDS denatured polyacrylamide gel and then
transferred onto a polyvinylidene difluoride membrane. The
membranes were blocked with 5% skim milk at room temperature for 1
h, and were subsequently incubated with anti-β-actin, anti-PPARγ,
anti-C/EBPα, anti-aP2, anti-YAP, anti-p-YAP, anti-LATS1 and
anti-MST at 4°C overnight (all at 1:1,000). Membranes were washed
in PBS with 0.1% Tween-20 and incubated with horseradish peroxidase
(HRP)-conjugated goat anti-mouse IgG and goat anti-rabbit IgG
secondary antibodies (1:3,000, cat. no. ZDR-5307 and ZDR-5306;
OriGene Technologies, Inc., Beijing, China) according to the
manufacturer's instructions. Finally, the protein of interest was
visualized using Immobilon Western Chemiluminescent HRP substrate
(EMD Millipore, Billerica, MA, USA). Intensity analysis of images
was performed using ImageJ software (version 1.48v; National
Institutes of Health, Bethesda, MD, USA).
Oil red O staining
3T3-L1 cells that reached ~90% confluence were
cultured in AIM or treated with different concentrations of
liraglutide (0, 10, 100 or 1,000 nM) for 3 days. Following this,
the treated adipocyte-induced 3T3-L1 cells were gently washed twice
with cold PBS, and then fixed in 4% paraformaldehyde for 10 min at
room temperature. Following this, the cells were washed twice with
deionized water, and stained with 60% saturated Oil Red O for 5 min
at room temperature. For Oil Red O quantification, 4% IGEPAL CA 630
(Sigma-Aldrich; Merck KGaA) in isopropanol was added to each well,
and the plate was then rocked on a shaker for 15 min. Light
absorbance by the extracted dye was measured at a wavelength of 520
nm.
Cell immunofluorescence
Adipocyte-induced 3T3-L1 cells were treated with
different concentrations of liraglutide (0, 10, 100 or 1,000 nM) in
a humidified atmosphere at 37°C with 5% CO2 for 2 days
using the aforementioned procedure. Cells were fixed in 4%
paraformaldehyde for 15 min and permeabilized with 1% Triton X-100
at room temperature for 15 min. Following blocking with 1% bovine
serum albumin at room temperature for 2 h, cells were incubated
with rabbit anti-Ki67 (1:200) at 4°C for 12 h and incubated with
goat anti-rabbit Alexa Fluor 488 at room temperature for 1 h
(1:3,000, cat. no. A-31565; Invitrogen; Thermo Fisher Scientific,
Inc.). The images were acquired using the AV300-ASW confocal
microscope (Olympus Corporation, Tokyo, Japan). Image
magnification, ×100.
Cell proliferation assay
A Cell Counting Kit (CCK)-8 kit (Beyotime Institute
of Biotechnology) was used for the evaluation of cell
proliferation. A total of 3×103 cells/well were seeded
into 96-well plates, which were cultured in AIM medium and treated
with different concentrations of liraglutide (0, 0.1, 1, 10, 100 nM
or 1, 10, 100 µM) in a humidified atmosphere at 37°C with 5%
CO2 for 48 h. CCK-8 reagent was then added to each well
and incubated for 2 h at 37°C. The absorbance was measured using a
pan-wavelength microplate reader at a wavelength of 450 nm
(SynergyMx; BioTek Instruments, Inc., Winooski, VT, USA).
Cell apoptosis assay by flow cytometry
(FCM)
Adipocyte-inducing 3T3-L1 cells at ~90% confluence
were treated with different concentrations of liraglutide (0, 10,
100 or 1,000 nM) in a humidified atmosphere at 37°C with 5%
CO2 for 2 days. Subsequently, the cells were used for
apoptosis analysis. For FCM analysis, an Annexin V-APC Apoptosis
Detection kit from Tianjin Biotech Co., Ltd. (Beijing, China) was
used. According to the manufacturer's instructions, the cells were
washed twice with cold PBS and resuspended in 1X binding buffer at
a concentration of 1×106 cells/ml. This solution (100
µl) was transferred to a 5 ml culture tube and 5 µl FITC-Annexin V
and 5 µl PI was added. Cells were gently vortexed and incubated for
15 min at room temperature in the dark. Subsequently, 400 µl of 1X
binding buffer was added to each tube and was analyzed by FCM
(FACSAria I; BD Biosciences, Franklin Lakes, NJ, USA).
Cell transfection
According to the manufacturer's instructions,
transfection was performed using Lipofectamine RNAiMAX in
Opti-MEM® I Medium (both from Invitrogen; Thermo Fisher
Scientific, Inc.) at 37°C for 12 h. Briefly, cells were counted and
seeded in 12 well plates at 1.5×105 cells/well the day
prior to transfection to ensure ~50% confluency on the day of
transfection. For 3T3-L1 cell transfection, 50 nM MST1 siRNA
(siMST1) or its negative control (siNC; Shanghai GenePharma Co.,
Ltd., Shanghai, China) were used. The siRNA sequences were as
follows: siMST1-1, 5′-GGGACUAGAAUACCUUCAU-3′; siMST1-2,
5′-GGGAAUAACUGCCAUAGAA-3′; siMST1-3, 5′-GGAGAACUCAGAGGAGGAU-3′;
siNC, 5′-UUCUCCGAACGUGUCACGU-3′.
Statistical analysis
The results are expressed as the mean ± standard
deviation. A one-way analysis of variance was used followed by
Tukey's or Dunnett's post hoc test to analyze the differences
between multiple groups. Statistical analysis was performed using
SPSS software, version 17 (SPSS, Inc., Chicago, IL, USA). P<0.05
was considered to indicate a statistically significant
difference.
Results
Liraglutide promotes adipogenic
differentiation of 3T3-L1 cells
The authors of the present study previously
demonstrated that adipogenic induced 3T3-L1 cells may produce a
significant increase in lipid droplet numbers when treated with
liraglutide (18). In the current
study, the adipocyte formation effect of liraglutide was
investigated at the early phase of adipogenic differentiation.
Preadipocyte 3T3-L1 cells were cultured in AIM, and were treated
with different concentrations of liraglutide (0, 10, 100 or 1,000
nM). Following 5 days treatment, Oil Red O staining was performed
to examine the lipid droplet production. In a dose-dependent
manner, 1,000 nM Liraglutide treatment led to a maximal increase of
lipid droplet numbers (34% increase in Oil Red O staining compared
with the vehicle group; P<0.05; Fig. 1A and B). Liraglutide also
significantly increased the mRNA expression levels of adipocyte
marker genes, including C/EBPα, PPARγ and aP2 compared with the
levels in the vehicle group (Fig.
1C-E). When cells were treated with 10, 100 and 1,000 nM
liraglutide, the mRNA expression levels of C/EBPα increased by
1.23-fold (P<0.05), 1.58-fold (P<0.01) and 1.63-fold
(P<0.01), respectively (Fig.
1C), the mRNA expression levels of PPARγ increased by 1.18-fold
(P<0.05), 1.54-fold (P<0.001) and 1.59-fold (P<0.001),
respectively (Fig. 1D), and the
mRNA expression levels of aP2 increased by 1.38-fold (P<0.01),
1.75-fold (P<0.001) and 1.96-fold (P<0.001), respectively
(Fig. 1E). Enhanced expression of
C/EBPα, PPARγ and aP2 was further verified by western blot analysis
(Fig. 1F). These data suggested
that liraglutide accelerates adipogenesis at an early stage of
differentiation.
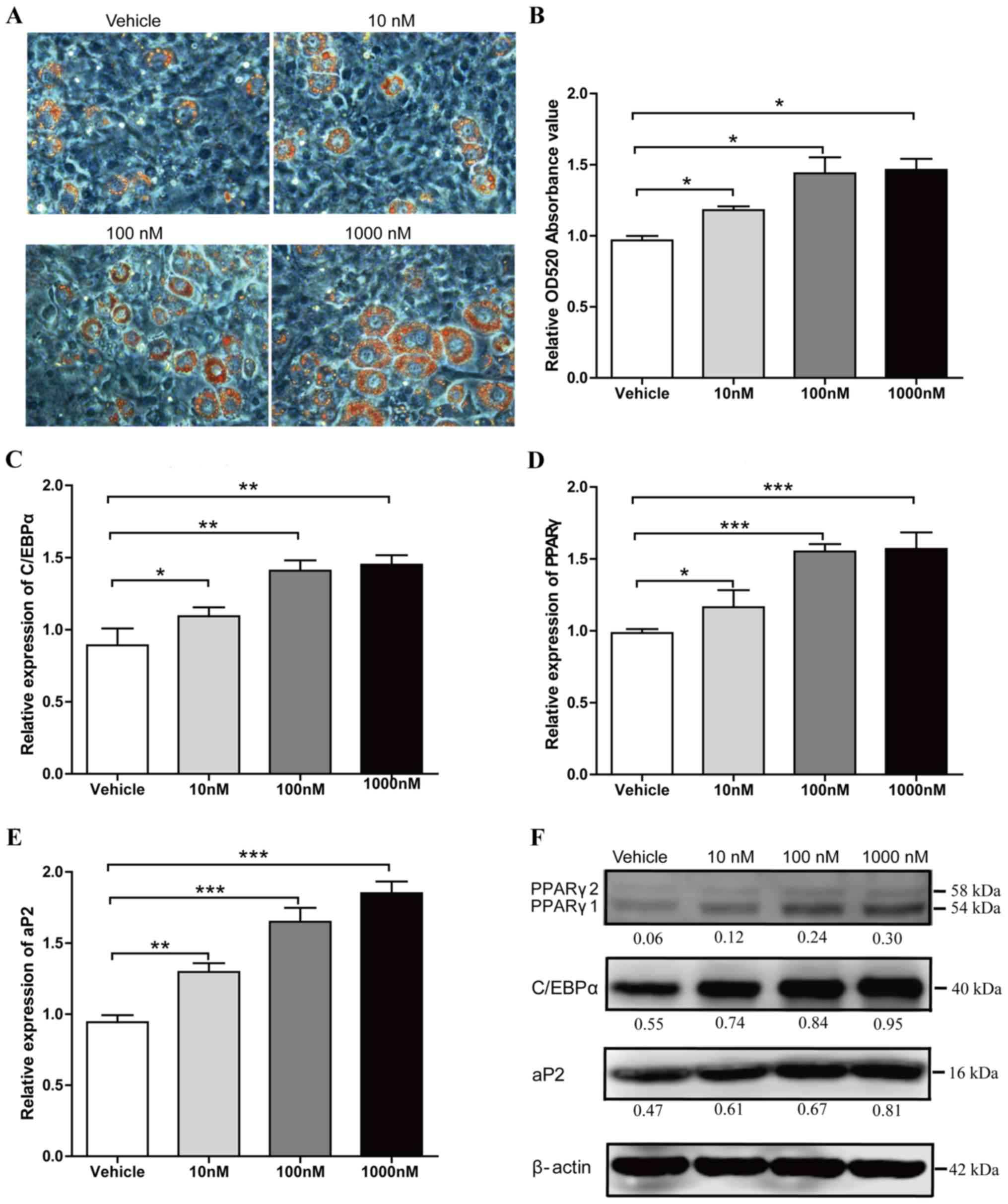 | Figure 1.Liraglutide promotes adipogenic
differentiation of 3T3-L1 cells. Preadipocyte 3T3-L1 cells were
cultured in adipocyte-inducing medium and treated with different
concentrations of liraglutide. (A) Representative images of
differentiated and treated cells were labeled with Oil Red O at day
5. (B) Oil Red O extracted with isopropanol was measured at OD520,
the values represent the mean ± standard deviation. The mRNA
expression levels of key transcription factors, (C) C/EBPα and (D)
PPARγ, as well as (E) the adipocyte-specific gene aP2, were
analyzed by reverse transcription-quantitative polymerase chain
reaction at day 2. Results represent the mean ± standard deviation.
(F) The protein expression levels of C/EBPα, PPARγ and aP2 were
assessed by western blot analysis at day 3 of differentiation. The
value under each lane indicates the relative expression level of
the genes, which is represented by the intensity ratio between
PPARγ, C/EBPα or aP2 and β-actin bands in each lane. β-actin was
used as an internal control. Image magnification, ×200, and all
data represent three separate experiments. One-way analysis of
variance was used followed by Tukey's post hoc test to analyze the
differences between the other groups with the vehicle group in B-E.
*P<0.05, **P<0.01 and ***P<0.001 vs. vehicle group. OD,
optical density; C/EBPα, CCAAT/enhancer binding protein α; PPARγ,
peroxisome proliferator-activated receptor γ; ap2, adipocyte
protein 2. |
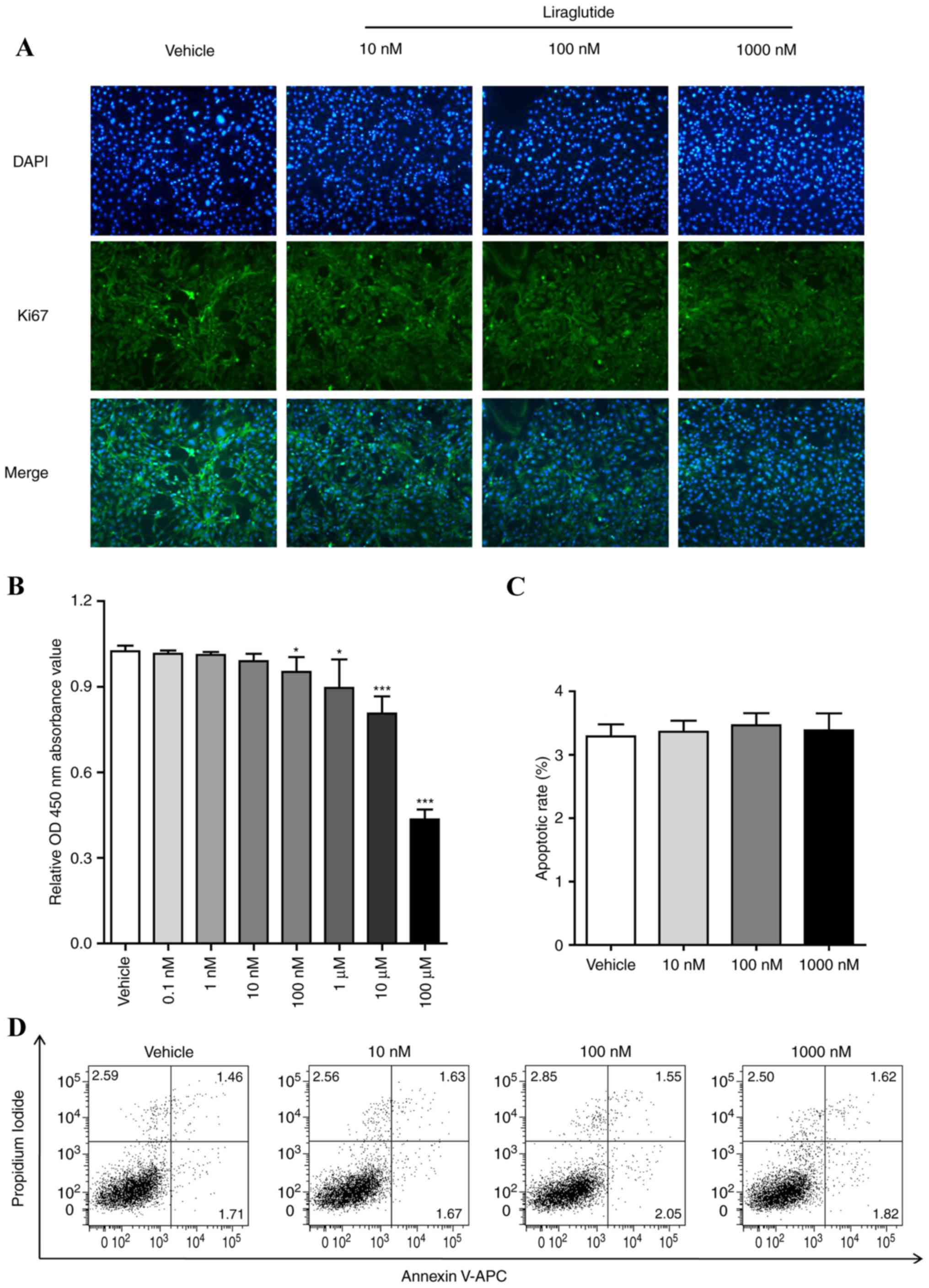
Liraglutide suppresses the
proliferation of 3T3-L1 cells and does not affect apoptosis
Considering that there is a stage of mitotic clonal
expansion prior to the terminal differentiation stage of
adipogenesis, the effect of liraglutide on cell proliferation of
3T3-L1 cells was examined. 3T3-L1 cells were treated with different
concentrations of liraglutide (0, 10, 100 or 1,000 nM) for 48 h,
the expression of Ki67, which is a marker for determining the
growth fraction of a given cell population, was determined by
immunostaining. As presented in Fig.
2A, the green fluorescence in the cell nucleus treated with
different concentrations of liraglutide (0, 10, 100 or 1,000 nM)
was decreased in a dose-dependent manner. 3T3-L1 cells were treated
using the aforementioned procedure and CCK-8 analysis was performed
to measure the cell proliferation. The results demonstrated that
liraglutide reduced the cell growth in a dose-dependent manner
(Fig. 2B). The effect of
liraglutide on the cell apoptosis of preadipocyte 3T3-L1 was also
examined. 3T3-L1 cells were treated with different concentrations
of GLP-1 (0, 10, 100 or 1,000 nM) for 48 h, and cell apoptosis was
assessed by FCM assay. As presented in Fig. 2C and D, liraglutide did not affect
apoptosis of 3T3-L1 cells compared with the vehicle group. It was
also observed that apoptosis of 3T3-L1 cells were not influenced
when the cells were induced with AIM and liraglutide following 5
days (data not shown).
Liraglutide activates the Hippo-YAP
signaling pathway during adipogenesis
Previous investigations have demonstrated that the
Hippo-YAP signaling pathway is involved in cell proliferation and
adipogenesis. To clarify whether liraglutide affects this signaling
pathway during adipogenesis, the expression of MST1, LATS1, YAP and
Ser127 phosphorylated-YAP [p-YAP (S127)] were measured at the early
phase of differentiation. 3T3-L1 cells were incubated in AIM
containing 0, 10, 100 and 1,000 nM liraglutide for 48 h, then the
cells were harvested and the expression levels of these proteins
was measured by western blotting. As presented in Fig. 3A, no significant alteration in the
total protein expression level of YAP was observed, and the levels
of MST1, LATS1 and p-YAP (S127) in liraglutide-treated cells were
markedly increased compared with vehicle control cells. ANKRD1,
CTGF and Cyr61 are well-characterized YAP target genes, and
therefore, it was next examined whether liraglutide altered the
levels of their expression. Compared with the mRNA expression
levels in the vehicle group, when cells were treated with 10, 100
and 1,000 nM liraglutide, the levels of ANKRD1 decreased by
0.56-fold (P<0.01), 0.56-fold (P<0.01) and 0.50-fold
(P<0.01), respectively (Fig.
3B), the levels of CTGF decreased by 0.61-fold (P<0.01),
0.58-fold (P<0.01) and 0.73-fold (P<0.05), respectively
(Fig. 3C), and the mRNA levels of
Cyr61 decreased by 0.54-fold (P<0.01), 0.60-fold (P<0.01) and
0.60-fold (P<0.01), respectively (Fig. 3D). These data also suggested that
liraglutide may reduce 3T3-L1 cell proliferation.
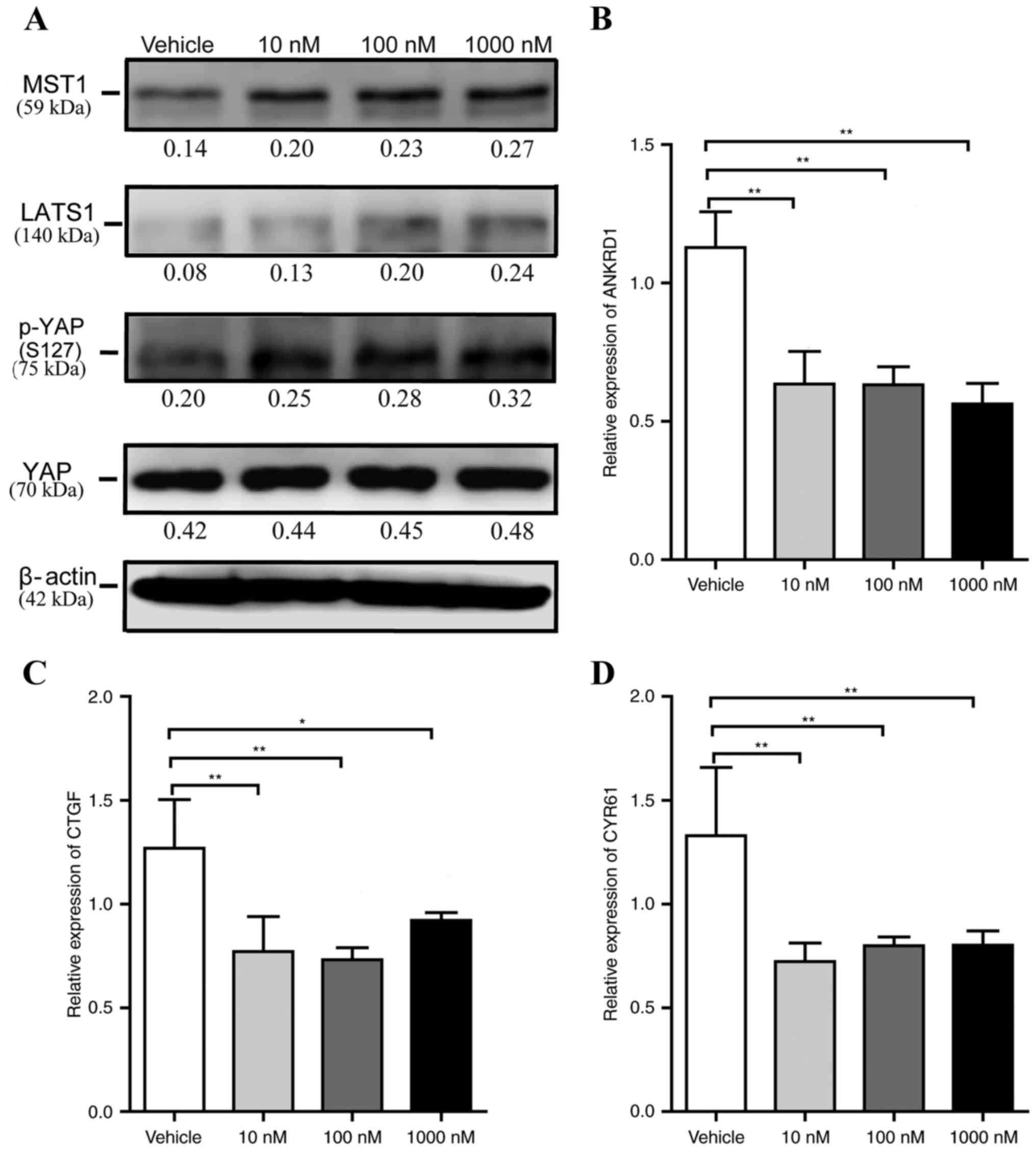 | Figure 3.Liraglutide activates the Hippo-YAP
signaling pathway during adipogenesis. 3T3-L1 cells were cultured
in adipocyte-inducing medium and treated with different
concentrations of liraglutide, and then the cells were harvested at
day 2. (A) Western blot analysis was used to assess the protein
expression levels of the core components of the Hippo-YAP signaling
pathway, including MST1, LATS1, p-YAP (S127) and total YAP. The
value under each lane indicates the relative expression level of
the gene, which is represented by the intensity ratio between MST1,
LATS1, p-YAP (S127) or total YAP and β-actin bands in each lane.
β-actin was used as an internal control. Reverse
transcription-quantitative polymerase chain reaction was used to
determine the mRNA expression levels of YAP specific target genes,
including (B) ANKRD1, (C) CTGF and (D) Cyr61. The values
represented the mean ± standard deviation. All data are
representative of at least three independent experiments. One-way
analysis of variance was applied followed by Dunnett's test to
analyze the differences between the vehicle, 10, 100 and 1,000 nM
groups in B-D. *P<0.05 and **P<0.01 vs. vehicle group. YAP,
Yes-associated protein; p-YAP, phosphorylated yes-associated
protein; MST1, mammalian ste20 kinase 1/2; LATS1/2, large tumor
suppressor 1/2; ANKRD1, ankyrin repeat domain 1; CTGF, connective
tissue growth factor; Cyr61, cysteine rich angiogenic inducer
61. |
Silencing of MST1 attenuates
liraglutide stimulation of adipogenic differentiation
To further clarify whether the Hippo-YAP signaling
pathway mediates liraglutide promotion of adipogenesis, liraglutide
gain-of-function studies were performed on the background of MST1
silencing. 3T3-L1 cells were transfected with siMST1s or siNC for
48 h, and as presented in Fig. 4A and
B, the mRNA and protein expression levels of MST1 were
significantly reduced by all three siMST1s, and the expression of
p-YAP (S127) was also decreased by MST1 siRNAs. Subsequently, one
of three siMST1s was selected to perform further experiments.
3T3-L1 cells were transfected with siMST1 or siNC for 24 h, and
then, the cells were incubated in AIM containing 0 or 100 nM
liraglutide. As shown in Fig. 4C and
D, knockdown of MST1 partially abrogated liraglutide-enhanced
adipogenesis. Consistently, the protein expression levels of
adipogenic differentiation marker genes including C/EBPα, PPARγ and
aP2 were decreased in the cells transfected with siMST1 and
incubated with 100 nM liraglutide, as compared with the cells
transfected with siNC and incubated with 100 nM liraglutide
(Fig. 4E). These results suggested
that the Hippo-YAP signaling pathway may be involved in the process
of liraglutide enforced adipogenesis during the early phase of
differentiation.
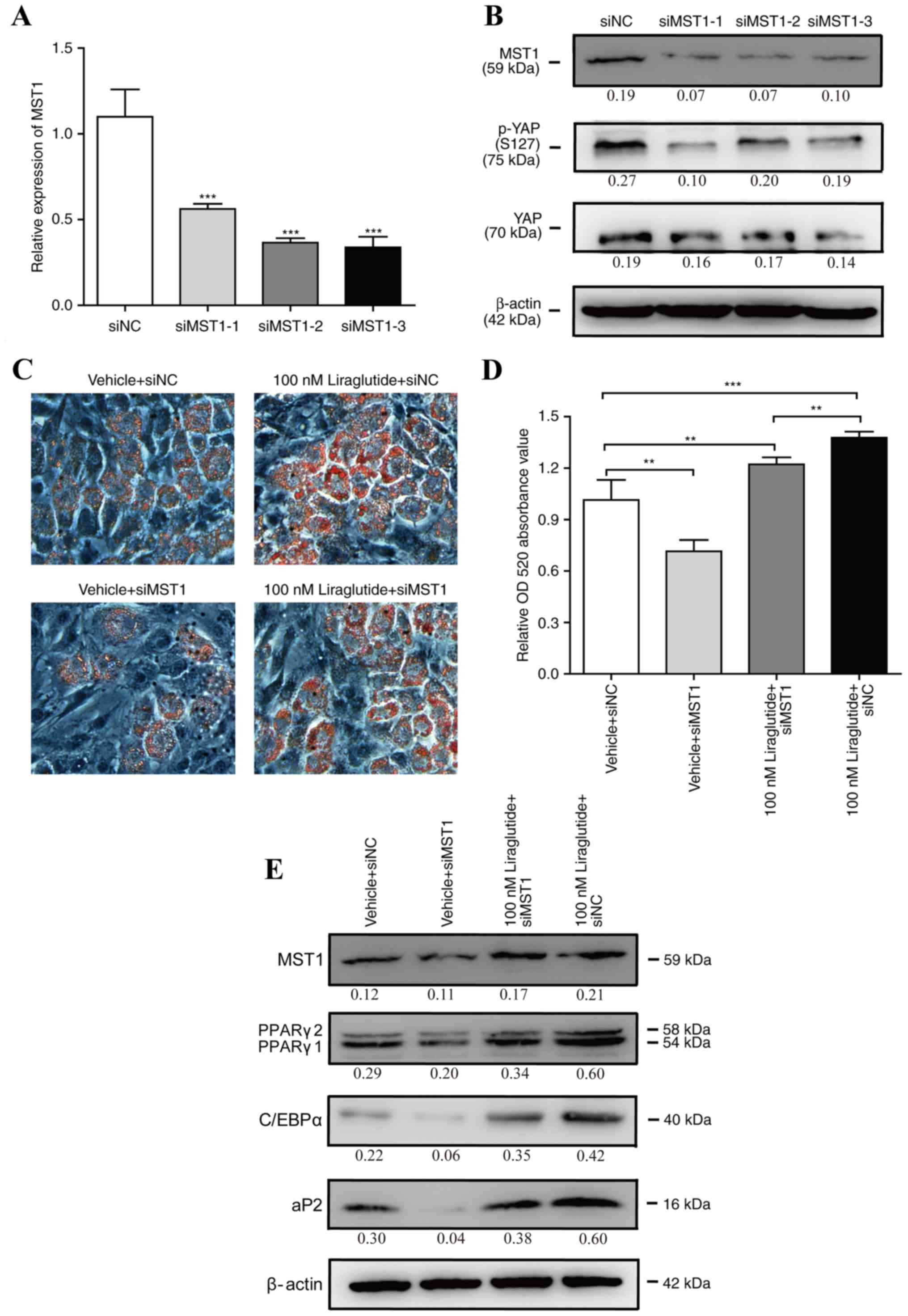 | Figure 4.Silencing of MST1 attenuates
liraglutide stimulation of adipogenic differentiation. 3T3-L1 cells
were transfected with siMST1s or siNC. (A) Following 2 days,
reverse transcription-quantitative polymerase chain reaction was
used to monitor the mRNA expression levels of MST1, and NONO served
as the internal control. The values represent the mean ± standard
deviation. ***P<0.001 vs. siNC group (B); following 3 days,
western blotting was used to measure the protein expression levels
of MST1, p-YAP (S127) and total YAP. The value under each lane
indicates the relative expression level of the gene, which is
represented by the intensity ratio between MST1, p-YAP (S127) or
total YAP and β-actin bands in each lane. β-actin was used as the
internal control. 3T3-L1 cells were transfected with siMST1 or siNC
for 24 h, and then, the cells were incubated in adipocyte-inducing
medium containing 0 or 100 nM liraglutide. (C) Representative
images of differentiated and treated cells were labeled with Oil
Red O at day 5. (D) Oil Red O extracted with isopropanol was
measured at OD520, and the values represented the mean ± standard
deviation. **P<0.01 and ***P<0.001 vs. vehicle + siNC group.
(E) The protein expression levels of MST1, C/EBPα, PPARγ and aP2
were assessed by western blot analysis at day 3 of differentiation.
The value under each lane indicates the relative expression level
of the gene, which is represented by the intensity ratio between
MST1, PPARγ, C/EBPα or aP2 and β-actin bands in each lane. β-actin
was used as an internal control. Image magnification, ×200, and all
data represented three separate experiments. One-way analysis of
variance followed by Tukey's post hoc test was used to analyze the
differences between groups with the siNC group in A or the vehicle
+ siNC group in D. MST1, mammalian ste20 kinase 1; siMST1s,
mammalian ste20 kinase 1 short interfering RNA; siNC, negative
control short interfering RNA; p-YAP, phosphorylated-yes associated
protein; YAP, yes associated protein; OD, optical density; C/EBPα,
CCAAT/enhancer binding protein α; PPARγ, peroxisome
proliferator-activated receptor γ; ap2, adipocyte protein 2. |
Discussion
Obesity results in insulin resistance that results
in individuals having an increased risk of developing T2DM, and
insulin resistance may be partly induced by adipocyte hypertrophy,
hyperplasia and dysfunction (5).
It has been suggested by clinical studies that treatment with
liraglutide controls body weight gain, improves insulin resistance
and reduces adipocyte hyperplasia (22,23).
A previous study revealed that liraglutide treatment may lead to
adipogenic differentiated 3T3-L1 cells to produce a significant
increase in lipid droplet numbers at the stage of terminal
differentiation (18). The present
study focused on the function and underlying mechanism of
liraglutide in the regulation of adipogenesis at the early
phase.
Adipocytes originate from multipotent mesenchymal
stem cells (MSC). During adipogenesis, fibroblast-like pluripotent
MSCs differentiate into mature lipid-laden and insulin-responsive
adipocytes and the process of adipogenesis involves several stages,
which includes mesenchymal precursor and committed preadipocyte
proliferation, as well as their differentiation into terminally
mature adipocytes (24). This
differentiation is tightly regulated by an intricate network of
transcriptional factors, which is governed to a large extent by an
adipocyte-enriched nuclear receptor, PPARγ. PPARγ is both necessary
and sufficient for adipogenesis, and PPARγ cooperates with C/EBPs
to induce the expression of numerous downstream target genes
important for terminal differentiation including aP2 (13). In the current study, it was
observed that liraglutide increased the lipid droplet production of
3T3-L1 cells. It was also demonstrated that liraglutide promoted
the expression of the master transcriptional factors C/EBPα and
PPARγ, and enhanced the expression of the downstream
adipocyte-specific gene aP2 in a dose-dependent manner. These
results suggested that liraglutide may accelerate adipogenesis
through a process that upregulates the expression of C/EBPα and
PPARγ at the early phase of adipogenic differentiation, then
promoted the expression of lipogenesis associated genes including
aP2, and enhanced the accumulated of lipids.
Next, the present study demonstrated that
liraglutide reduced 3T3-L1 cell growth in a dose-dependent manner
and did not affect apoptosis at the early phase of adipogenic
differentiation. Considering that there is a stage of committed
preadipocyte proliferation prior to the terminal differentiation
phase of adipogenesis, it was hypothesized that liraglutide may
force preadipocyte 3T3-L1 cells to achieve transformation into
mature adipocytes earlier, and enhance adipogenesis. Previously,
the Hippo-YAP signaling pathway has been demonstrated to have an
important role in the regulation of cell proliferation and
differentiation. This pathway was initially defined through genetic
studies in Drosophila for tumor suppressor genes (6). In mammalian systems, the core
components of the Hippo-YAP signaling pathway initiate a kinase
cascade, which acts on a transcriptional complex to regulate the
expression of target downstream genes that control cell
proliferation (25). Briefly, as
STE20 family protein kinases, MST1/2 is associated with Sav1/WW45
to phosphorylate Mob1 and LATS1/2. Phosphorylated Mob1 binds to the
autoinhibitory motif in LATS1/2, which activates their
phosphorylation loop and kinase activity. Next, the active complex
(combined LATS1/2 with Mob1) phosphorylates downstream effectors
YAP/TAZ, and this leads to their cytoplasmic retention and
inhibition. Dephosphorylated YAP/TAZ accumulates in the nucleus and
binds to DNA-binding transcription factors to initiate the
expression of growth-promoting and apoptosis-inhibiting genes
(7,25). Therefore, the active Hippo-YAP
signaling pathway induces the cytoplasmic accumulation of
phosphorylated YAP and the inhibition of growth promoting genes. In
the present study, it was demonstrated that liraglutide increased
the levels of the core components of the Hippo-YAP signaling
pathway, including MST1, LATS1 and p-YAP (S127) at the early phase
of adipogenesis. Consistently, YAP specific target genes were
downregulated in liraglutide-treated 3T3-L1 cells, including
ANKRD1, CTGF and Cyr61. Silencing of MST1 reduced adipogenic
differentiation of 3T3-L1 cells, and silencing of MST1 counteracted
the effect of increasing adipogenesis by liraglutide. Previous
studies have reported that MST2 interacts with Sav1 to activate
PPARγ and augments PPARγ-induced adipocyte differentiation
(26). LATS2 phosphorylated YAP
and TAZ and retained them in the cytoplasm, leading to the
reduction of cell proliferation and the promotion of cell
adipogenic differentiation (10).
The results of the present study and previous studies suggest that
the activation of the Hippo-YAP signaling pathway may be involved
in the process of liraglutide enhanced adipogenic
differentiation.
In conclusion, the present study demonstrated that
liraglutide promoted adipogenic differentiation of preadipocyte
3T3-L1 cells. In addition, liraglutide may activate the Hippo-YAP
signaling pathway leading to proliferation inhibition of committed
preadipocyte, and accordingly, 3T3-L1 cells achieve transformation
into mature adipocytes sooner. The results may help to expand the
knowledge regarding the underlying mechanism of liraglutide
facilitating adipogenesis, and may provide a theoretical support
for liraglutide in T2DM and obesity treatment.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81501846 and
81270927), the Scientific Foundation of Tianjin Medical University
(grant no. 2014KYM16), the Scientific Foundation of Tianjin
Metabolic Diseases Hospital and Tianjin Institute of Endocrinology,
Tianjin Medical University (grant no. 2014RC01) and the Tianjin
Municipal Natural Science Foundation of China (grant no.
16JCYBJC26800).
References
|
1
|
Yu D, He Y, Guo Q, Fang H, Xu X, Fang Y,
Li J and Zhao L: Trends of energy and nutrients intake among
Chinese population in 2002–2012. J Hygiene Res. 45:527–533.
2016.
|
|
2
|
Zhang N, Du SM and Ma GS: Current
lifestyle factors that increase risk of T2DM in China. Eur J Clin
Nutr. 71:832–838. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Flier JS: Obesity wars: Molecular progress
confronts an expanding epidemic. Cell. 116:337–350. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Guilherme A, Virbasius JV, Puri V and
Czech MP: Adipocyte dysfunctions linking obesity to insulin
resistance and type 2 diabetes. Nat Rev Mol Cell Biol. 9:367–377.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
de Ferranti S and Mozaffarian D: The
perfect storm: Obesity, adipocyte dysfunction, and metabolic
consequences. Clin Chem. 54:945–955. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhao B, Li L, Lei Q and Guan KL: The
Hippo-YAP pathway in organ size control and tumorigenesis: An
updated version. Genes Dev. 24:862–874. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yu FX and Guan KL: The Hippo pathway:
Regulators and regulations. Genes Dev. 27:355–371. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang Y, Park HW, Jewell JL, Chen Q, Deng
Y, Pan D, Taylor SS, Lai ZC and Guan KL: Protein kinase A activates
the Hippo pathway to modulate cell proliferation and
differentiation. Genes Dev. 27:1223–1232. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhao B, Panupinthu N, Jewell JL, Lian I,
Wang LH, Zhao J, Yuan H, Tumaneng K, Li H, Fu XD, et al: Regulation
of the Hippo-YAP pathway by G-protein-coupled receptor signaling.
Cell. 150:780–791. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
An Y, Kang Q, Zhao Y, Hu X and Li N: Lats2
modulates adipocyte proliferation and differentiation via hippo
signaling. PLoS One. 8:e720422013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Seo E, Basu-Roy U, Gunaratne PH, Coarfa C,
Lim DS, Basilico C and Mansukhani A: SOX2 regulates YAP1 to
maintain stemness and determine cell fate in the osteo-adipo
lineage. Cell Rep. 3:2075–2087. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen SN, Gurha P, Lombardi R, Ruggiero A,
Willerson JT and Marian AJ: The hippo pathway is activated and is a
causal mechanism for adipogenesis in arrhythmogenic cardiomyopathy.
Circ Res. 114:454–468. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ali AT, Hochfeld WE, Myburgh R and Pepper
MS: Adipocyte and adipogenesis. Eur J Cell Biol. 92:229–236. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Russell-Jones D: Molecular,
pharmacological and clinical aspects of liraglutide, a once-daily
human GLP-1 analogue. Mol Cell Endocrinol. 297:137–140. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Challa TD, Beaton N, Arnold M, Rudofsky G,
Langhans W and Wolfrum C: Regulation of adipocyte formation by
GLP-1/GLP-1R signaling. J Biol Chem. 287:6421–6430. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yang J, Ren J, Song J, Liu F, Wu C, Wang
X, Gong L, Li W, Xiao F, Yan F, et al: Glucagon-like peptide 1
regulates adipogenesis in 3T3-L1 preadipocytes. Int J Mol Med.
31:1429–1435. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu R, Li N, Lin Y, Wang M, Peng Y, Lewi K
and Wang Q: Glucagon like peptide-1 promotes adipocyte
differentiation via the Wnt4 mediated sequestering of beta-catenin.
PLoS One. 11:e01602122016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhu E, Yang Y, Zhang J, Li Y, Li C, Chen L
and Sun B: Liraglutide suppresses obesity and induces brown
fat-like phenotype via Soluble Guanylyl Cyclase mediated pathway in
vivo and in vitro. Oncotarget. 7:81077–81089. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Arsenijevic T, Grégoire F, Delforge V,
Delporte C and Perret J: Murine 3T3-L1 adipocyte cell
differentiation model: Validated reference genes for qPCR gene
expression analysis. PLoS One. 7:e375172012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhu E, Wang X, Zheng B, Wang Q, Hao J,
Chen S, Zhao Q, Zhao L, Wu Z and Yin Z: miR-20b suppresses Th17
differentiation and the pathogenesis of experimental autoimmune
encephalomyelitis by targeting RORγt and STAT3. J Immunol.
192:5599–5609. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Astrup A, Rössner S, Van Gaal L, Rissanen
A, Niskanen L, Al Hakim M, Madsen J, Rasmussen MF and Lean ME;
NN8022-1807 Study Group, : Effects of liraglutide in the treatment
of obesity: A randomised, double-blind, placebo-controlled study.
Lancet. 374:1606–1616. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ng SY and Wilding JP: Liraglutide in the
treatment of obesity. Expert Opin Biol Ther. 14:1215–1224. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lefterova MI and Lazar MA: New
developments in adipogenesis. Trends Endocrinol Metab. 20:107–114.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gumbiner BM and Kim NG: The Hippo-YAP
signaling pathway and contact inhibition of growth. J Cell Sci.
127:709–717. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Park BH, Kim DS, Won GW, Jeon HJ, Oh BC,
Lee Y, Kim EG and Lee YH: Mammalian ste20-like kinase and SAV1
promote 3T3-L1 adipocyte differentiation by activation of PPARγ.
PLoS One. 7:e309832012. View Article : Google Scholar : PubMed/NCBI
|