Introduction
Gastric cancer is the fourth most common cancer and
the second most frequent cause of cancer-associated mortality
worldwide (1–3). Although several aggressive treatment
strategies, including surgery, radiotherapy and chemotherapy, are
used to treat gastric cancer, the treatment outcomes for advanced
gastric cancer remain unsatisfactory (4). The majority of patient mortalities
are due to tumor recurrence and metastasis. At present, there are
no effective therapeutic methods to prevent tumor recurrence and
metastasis (5). Therefore, the
development of effective and low toxicity drugs for the inhibition
of tumor recurrence and metastasis is required.
Natural, biologically active products are widely
used in clinical and basic research due to their low toxicity and
often potent effects (6). At
present, plant-derived anticancer drugs used clinically include
vinblastine, vincristine, paclitaxel, and camptothecin (7,8).
Salidroside is a phenyl propanoid glycoside extracted from the
flowering plant Rhodiola rosea. Salidroside has been
reported to have various pharmacological actions, including
anti-inflammatory (9,10), anti-tumor (11), and neuroprotective effects
(12). Growing evidence has
demonstrated that salidroside significantly affects the
proliferation, migration and apoptosis of various tumor types,
including fibrosarcoma, breast, colon and bladder cancer (6,7,11–13).
However, the effects of salidroside on gastric cancer cell
proliferation, migration and invasion remain to be elucidated.
Reactive oxygen species (ROS) within cells have been
identified as important second messengers in intracellular
signaling cascades, which induce and maintain the oncogenic
phenotype of cancer cells (14,15).
ROS are tumorigenic by virtue of their ability to increase cell
proliferation, survival and cellular migration (15). Therefore, inhibiting the production
of ROS may be exhibits anticancer effects in the treatment of
tumour (7,16,17).
Stress-inducible heat shock protein 70 (HSP70) is
expressed at extremely low levels in cells under normal conditions
and is not essential for life (18,19).
However, HSP70 is highly expressed in various tumor tissues,
including breast cancer and melanoma. In addition, HSP70 expression
is associated with cancer cell proliferation, apoptosis,
metastasis, prognosis and differentiation (19). Therefore, inhibition of HSP70
expression may serve as an effective strategy in cancer
therapy.
In the present study, the effects of salidroside on
the proliferation, migration and invasion of BGC-823 cells and the
potential molecular mechanisms were investigated. The results
revealed that salidroside inhibited BGC-823 cell proliferation,
migration and invasion in a dose-dependent manner, through the
suppression of HSP70 expression, as well as ROS-mediated and
proto-oncogene tyrosine-protein kinase Src (Src)-associated
signaling pathways. The present study may provide a novel
perspective for the application of salidroside in tumor
therapy.
Materials and methods
Antibodies and reagents
Salidroside (purity, >98%) was purchased from
Sigma-Aldrich (Merck KGaA, Darmstadt, Germany). The antibodies
against phosphorylated (p)-Src (Tyr416; cat. no. 6943s; 1:1,000),
Src (cat. no. 2109s; 1:1,000), p-protein kinase B (Akt; Ser473;
cat. no. 4060s; 1:500), Akt (cat. no. 9272s; 1:1,000), p-p44/42
mitogen-activated protein kinase 1 (p-ERK; Thr202/Tyr204; cat. no.
4376s; 1:1,000), ERK (cat. no. 9102s; 1:1,000), p-focal adhesion
kinase 1 (FAK; Tyr576/577; cat. no. 3281s; 1:1,000), FAK (cat. no.
3285s; 1:1,000), HSP70 (cat. no. 4873s; 1:1,000), E-cadherin (cat.
no. 3195s; 1:1,000), N-cadherin (cat. no. 4061s; 1:1,000), β-actin
(cat. no. 4970s; 1:1,000) and GAPDH (cat. no. 5174s; 1:1,000) were
obtained from Cell Signaling Technology, Inc., (Danvers, MA, USA).
A Cell Counting Kit (CCK)-8 assay was purchased from Nanjing KeyGen
Biotech Co., Ltd. (Nanjing, China). Matrix metalloproteinase
(MMP)-2 and −9 enzyme-linked immunosorbent assay (ELISA) kits were
obtained from ABclonal Biotech Co., Ltd., (Woburn, MA, USA).
Cell culture
The human gastric cancer cell line, BGC-823, was
purchased from Guangzhou Jennio Biotech Co., Ltd., (Guangzhou,
China). Cells were cultured in Dulbecco's modified Eagle's medium
(DMEM; HyClone; GE Healthcare Life Sciences, Logan, UT, USA)
supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA), 100 µg/ml streptomycin and 100
U/ml penicillin at 37°C in an environment containing 5%
CO2.
Cell viability assay
Cell viability was detected using a CCK-8 kit. In
brief, BGC-823 cells were seeded onto a 96-well plate at a density
of 1×104 cells/well. Following 12 h of culture at 37°C,
the cells were treated with different doses of salidroside (25, 50,
100, 200, 400 and 600 µg/ml) for 24 h at 37°C. CCK-8 reagent (10
µl) was subsequently added to each well and the cells were
incubated for a further 2 h at 37°C with 5% CO2. The
absorbance was detected using a Multiskan GO microplate
spectrophotometer (Thermo Fisher Scientific, Inc.) at a wavelength
of 450 nm. Each treatment was repeated three times.
Colony formation assay
BGC-823 cells were incubated in a 6-well culture
plate at 2,000 cells/well. After treatment with (200, 400 and 600
µg/ml) or without salidroside for 6 days at 37°C, the cell culture
medium was discarded. Cells were subsequently fixed with 4%
paraformaldehyde for 20 min at room temperature and stained with
0.1% crystal violet for 30 min at room temperature. Following this,
cells were washed 3 times in PBS and colonies containing >15
cells were counted under a fluorescence inverted microscope
(magnification, ×100), 5 fields of view were assessed, (Olympus
Corporation, Tokyo, Japan) and images were captured.
Scratch wound-healing assay
BGC-823 cells were seeded in a 12-well cell culture
plate and cultured at 37°C to full confluence. The confluent cell
monolayers were wounded by scratching with a pipette tip. Damaged
cells were removed by washing with PBS and remaining cells in the
plate were treated with salidroside (600 µg/ml) and incubated with
DMEM with 10% FBS for 24 h at 37°C. Images were captured of cell
migration over the injured area at 0 and 24 h using fluorescence
inverted microscope (magnification, ×40; Olympus Corporation).
Cell migration and invasion assay
Cell migration was analyzed in 24-well cell culture
plates with a Transwell membrane and 8-µm pore filter inserts (EMD
Millipore, Billerica, MA, USA). To examine invasion, wells were
coated with Matrigel (BD Biosciences, Franklin Lakes, NJ, USA).
Briefly, BGC-823 cells were harvested following treatment with 200,
400 and 600 µg/ml salidroside for 24 h at 37°C and suspended in
serum-free DMEM. Cells (2×104 cells/well) were added to
the upper chamber and 0.6 ml DMEM containing 20% FBS was added to
the lower chamber. After incubation for 24 h in the 5%
CO2 incubator at 37°C, cells that had migrated to the
lower surface of the membrane were fixed with 4% paraformaldehyde
for 20 min at room temperature. The membrane was washed with PBS
three times and cells were subsequently stained with 0.1% crystal
violet for 30 min at room temperature. The cells remaining on the
upper surface were wiped away gently with a cotton swab. Images of
the migrated cells were captured by fluorescence inverted
microscope (magnification, ×100; Olympus Corporation).
Measurement of intracellular ROS
Intracellular ROS levels were determined with a ROS
assay kit (Beyotime Institute of Biotechnology, Haimen, China).
Briefly, BGC-823 cells were treated with 600 µg/ml salidroside for
24 h at 37°C and subsequently incubated with 10
µmol/ldichloro-dihydro-fluorescein diacetate (DCFH-DA) for 30 min
at 37°C in the dark. Cells were washed with PBS prior to
examination by fluorescence microscopy (magnification, ×100;
Olympus Corporation).
Western blotting
Cells were lysed in radioimmunoprecipitation assay
buffer containing proteinase inhibitors (Beyotime Institute of
Biotechnology) for 30 min on ice. The protein was quantified by
BCA/Bradford assay. An equal amount of protein (50 µg) was loaded
and separated with 12% SDS-PAGE, and then the proteins were
transferred onto a nitrocellulose membrane, which was blocked with
5% skim milk powder for 1 h at room temperature. The membrane was
washed with PBS three times and incubated with the indicated
primary antibodies at 4°C overnight, followed by washing the
membrane again with PBS three times. The membrane was subsequently
incubated with IRdye 800CW-conjugated IgG secondary antibody (cat.
no. 926-32210; 1:5,000; LI-COR Biosciences, Lincoln, NE, USA) for 1
h at room temperature in the dark. The proteins were visualized
using an Odyssey infrared imaging system (LI-COR Biosciences).
Cell transfection
Green fluorescent protein (GFP)-labeled HSP70
overexpression plasmids and negative plasmids (supplier:
CMV-MCS-EGFP-SV40-Neomycin) were purchased from Shanghai GeneChem
Co., Ltd. (Shanghai, China). BGC-823 cells (2×105) were
transfected with 2 µg HSP70 overexpression and negative plasmids
for 24 h with Lipofectamine® 3000 reagent (Invitrogen;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
protocol.
ELISA assay
MMP-2 (cat. no. RK00309) and MMP-9 (cat. no.
RK00217) protein levels were detected in BGC-823 cells
(1×106) with ELISA kits (ABclonal Biotech Co., Ltd.)
according to the manufacturer's protocol. Analysis of each group
was repeated three times.
Statistical analysis
Each experiment was repeated three times. Data were
presented as the mean ± standard deviation. SPSS software, version
17.0 (SPSS, Inc., Chicago, IL, USA) was used to perform statistical
analysis. The results were compared using one-way analysis of
variance followed by a post hoc Tukey test for multiple
comparisons. P<0.05 was considered to indicate a statistically
significant difference.
Results
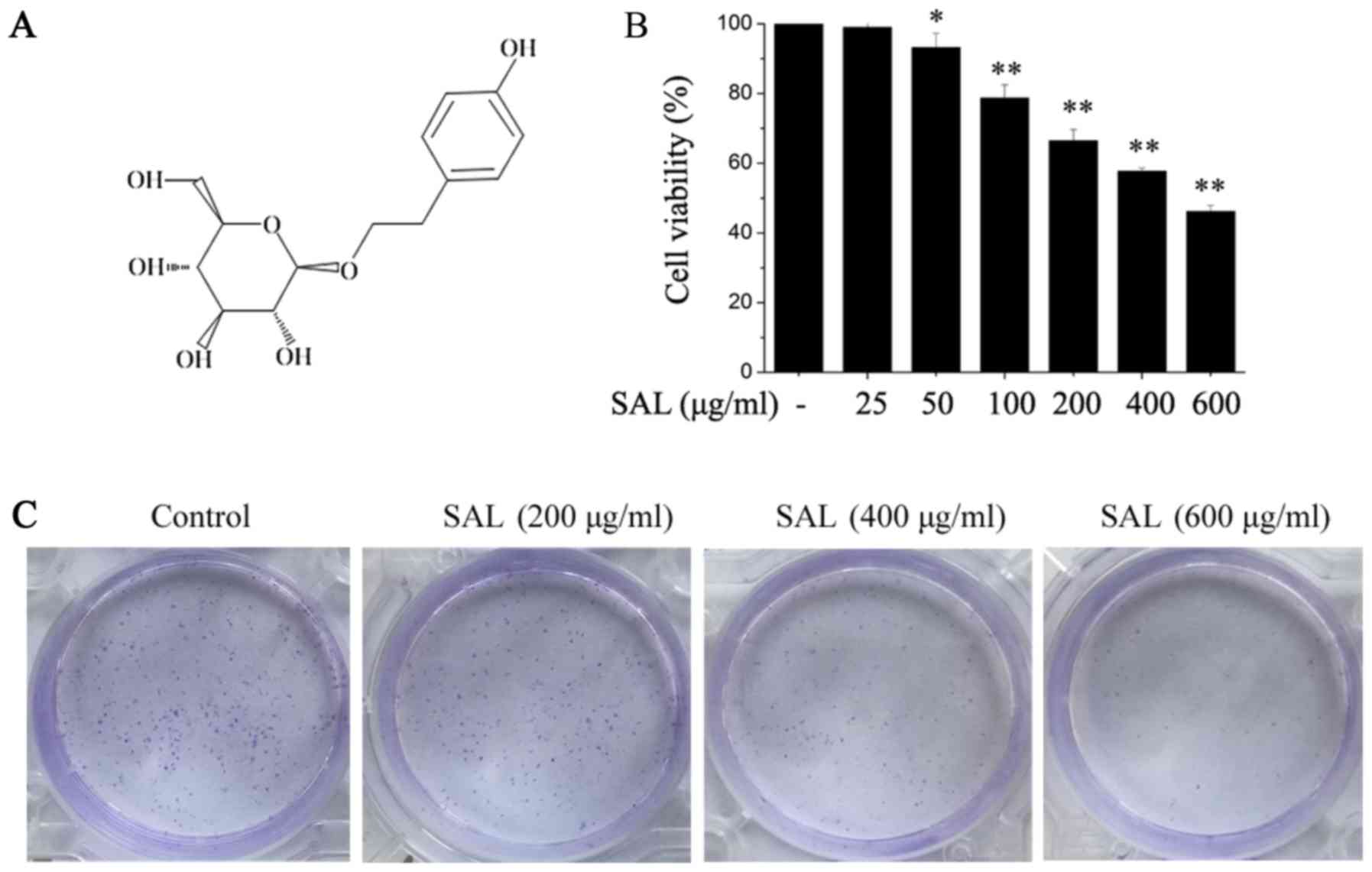
Salidroside inhibits the proliferation
and colony formation of BGC-823 cells
BGC-823 cells were treated with different doses of
salidroside (Fig. 1A) for 24 h,
followed by the addition of CCK-8 at 10 µl/well. Following
incubation for another 2 h, cell viability was detected.
Salidroside significantly suppressed BGC-823 cell viability and the
inhibitory effect was dose-dependent (Fig. 1B). According to these results,
salidroside concentrations of 200, 400 and 600 µg/ml were selected
for the subsequent experiments. The effects of salidroside on the
colony formation of BGC-823 cells were also determined. It was
demonstrated that salidroside clearly inhibited BGC-823 cell colony
formation in a dose-dependent manner (Fig. 1C).
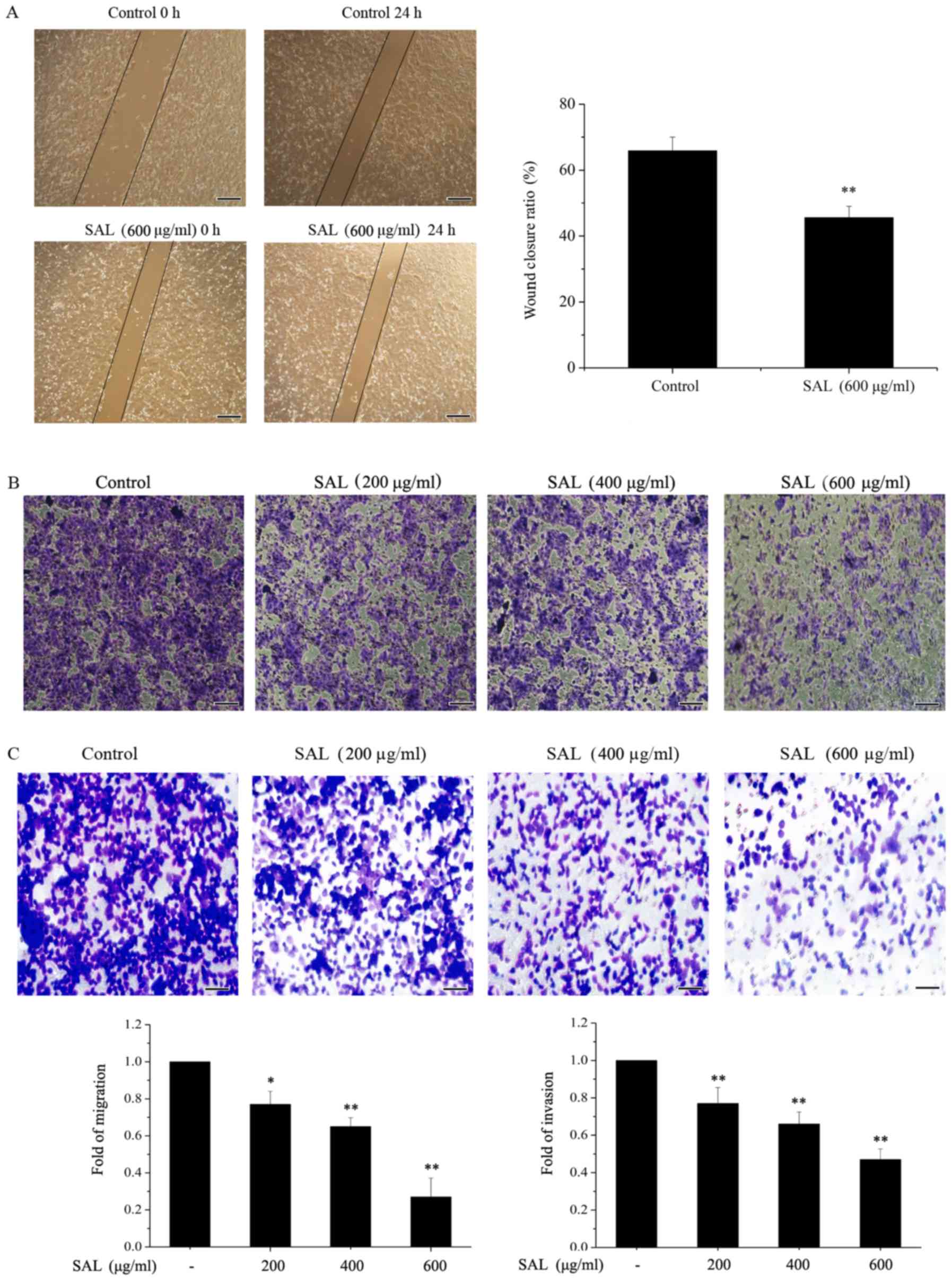
Salidroside suppresses the migration
and invasion of BGC-823 cells
A scratch wound healing assay was used to detect the
migration of BGC-823 cells (Fig.
2A). It was demonstrated that wound closure in cells treated
with salidroside (600 µg/ml) was significantly reduced compared
with the control cells. A Transwell assay was used to determine the
migration and invasion of BGC-823 cells. The migration (Fig. 2B) and invasion (Fig. 2C) of BGC-823 cells treated with
salidroside (200, 400 and 600 µg/ml) for 24 h was markedly
inhibited compared with the control cells, in a dose-dependent
manner.
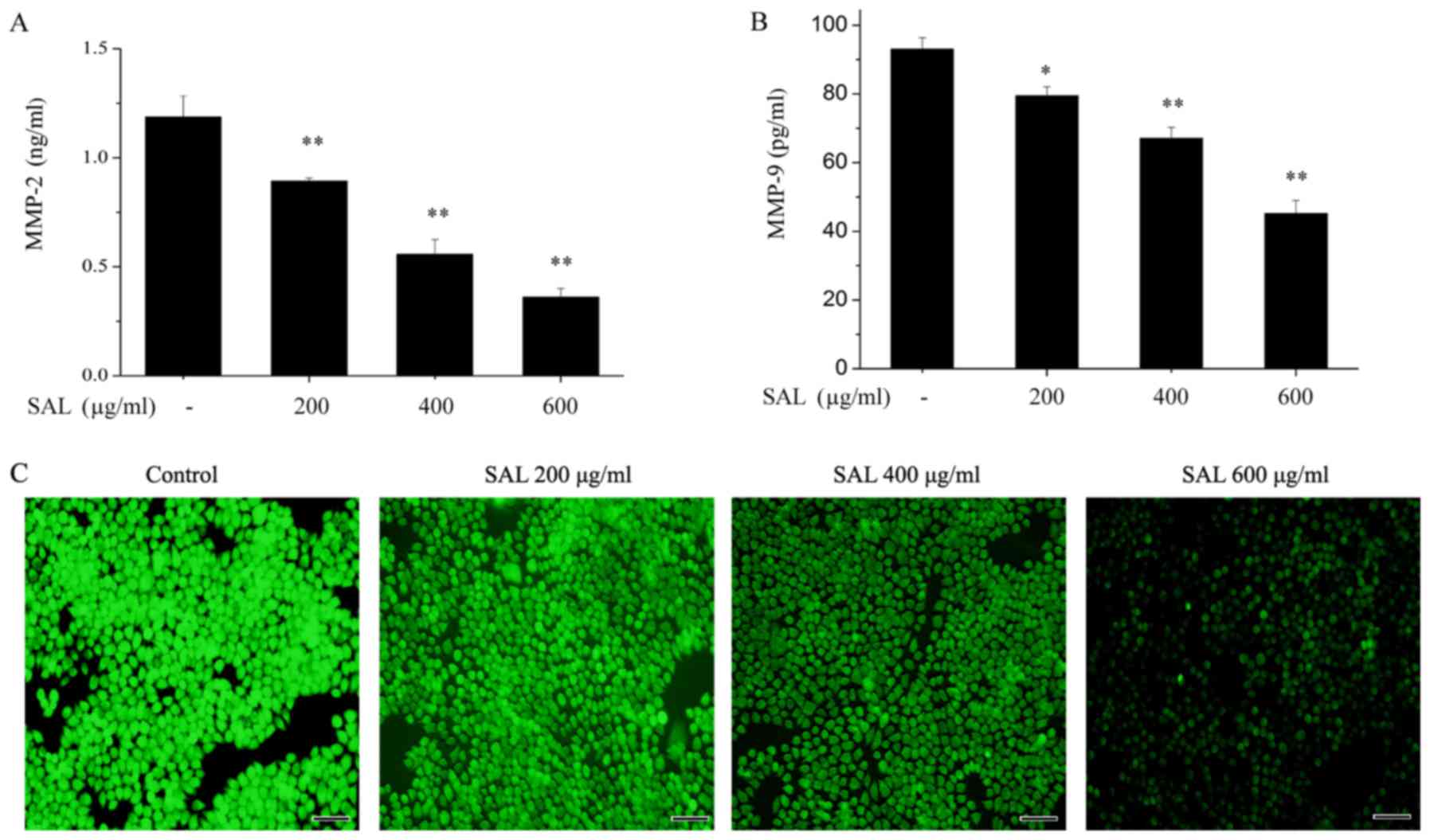
Salidroside reduces the levels of ROS,
MMP-2 and MMP-9 in BGC-823 cells conditioned medium
As MMPs have an important role in tumor cell
invasion (20), the effects of
salidroside on the levels of MMP-2 and MMP-9 levels were detected
by ELISA. Salidroside significantly reduced the levels of MMP-2
(Fig. 3A) and MMP-9 (Fig. 3B) in the treated medium of BGC-823
cells.
As oxidative stress may induce tumor cell migration
and invasion through MMP upregulation (20), the effects of salidroside on ROS
levels in BGC-823 cells were investigated. Intracellular ROS levels
were detected with DCFH-DA. As presented in Fig. 3C, salidroside treatment clearly
reduced the intracellular ROS levels in a dose-dependent
manner.
Salidroside inhibits the expression of
epithelial-mesenchymal transition (EMT) markers and the
phosphorylation of Src-associated signaling pathway proteins in
BGC-823 cells
A previous study reported that Src activation
governs a variety of signaling pathways, including proliferation,
migration and invasion, through various proteins, including Akt,
signal transducer and activator of transcription (STAT)3, ERK and
FAK (21,22). Therefore, the effects of
salidroside on Src-associated signaling pathways were investigated.
As presented in Fig. 4A,
salidroside treatment inhibited the phosphorylation of Src, Akt,
STAT3, ERK and FAK in a dose-dependent manner. EMT is thought to be
involved in the migration and invasion of tumor cells (23). Therefore, the effects of
salidroside on the expression of EMT markers were examined. The
protein expression levels of the epithelial marker E-cadherin and
the mesenchymal marker N-cadherin were detected in BGC-823 cells
treated with different concentrations of salidroside for 24 h.
Salidroside treatment significantly reduced the level of N-cadherin
and enhanced the expression of E-cadherin (Fig. 4B). The aforementioned results
suggested that salidroside may have inhibited the proliferation,
migration and invasion of BGC-823 cells through its effects on EMT
via Src-associated signaling pathways.
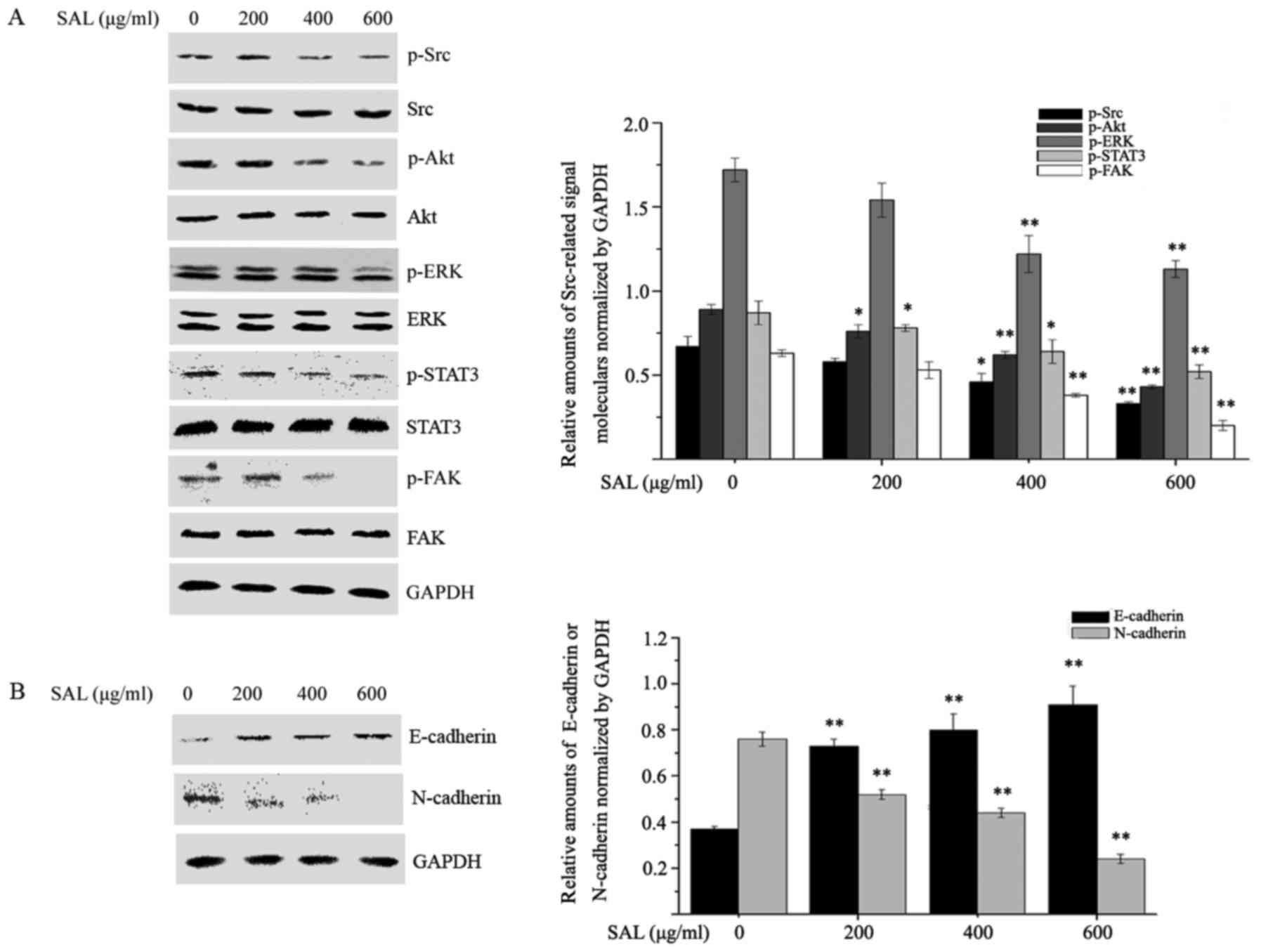 | Figure 4.Salidroside affects the
phosphorylation of Src-associated signaling pathway proteins and
epithelial-mesenchymal transition marker expression. BGC-823 cells
were treated with different concentrations of salidroside for 24 h
and (A) the phosphorylation of Src, Akt, STAT3, ERK and FAK, in
addition to (B) the levels of E-cadherin and N-cadherin were
detected by western blot analysis. *P<0.05, **P<0.01 vs.
control. p, phosphorylated; Akt, protein kinase B; Src,
proto-oncogene tyrosine-protein kinase Src; STAT3, signal
transducer and activator of transcription 3; ERK, mitogen-activated
protein kinase 1; FAK, focal adhesion kinase 1; SAL,
salidroside. |
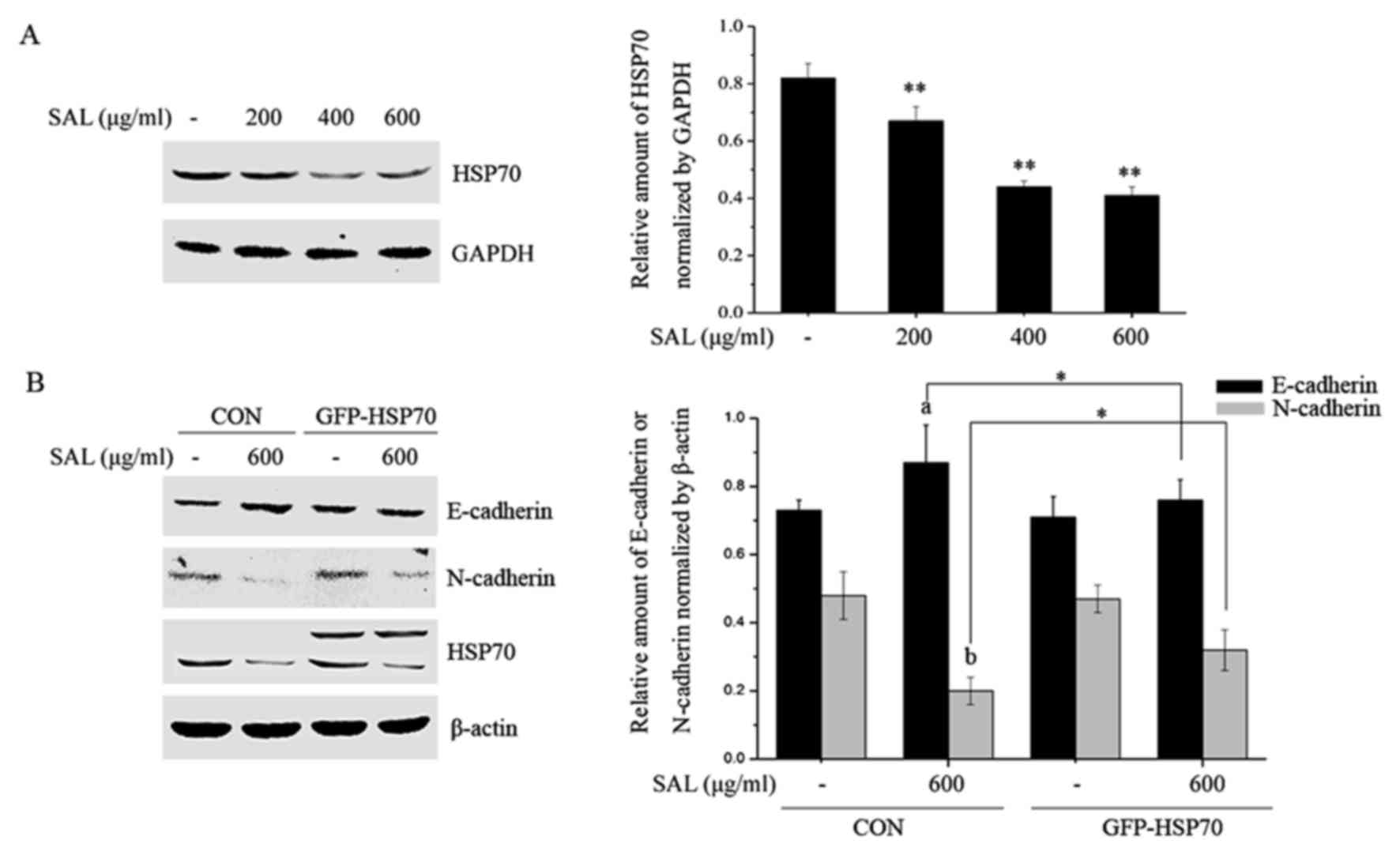
Salidroside attenuates HSP70
expression and HSP70 overexpression rescues cells from the effects
of salidroside on EMT markers
HSP70 is highly expressed in various tumor tissues
and its expression is positively correlated with cell proliferation
and metastasis (19); therefore,
the effects of salidroside on HSP70 expression were investigated.
BGC-823 cells were treated with different doses of salidroside for
24 h, total proteins were collected and the HSP70 levels were
determined by western blotting. Salidroside significantly
attenuated the expression of HSP70 (Fig. 5A). Subsequently, BGC-823 cells were
transfected with negative and HSP70 overexpression plasmids and the
role of HSP70 in salidroside-induced EMT marker expression was
investigated. As presented in Fig.
5B, salidroside treatment significantly enhanced levels of
E-cadherin and reduced the N-cadherin levels in negative plasmid
transfected BGC-823 cells. The alterations in E-cadherin and
N-cadherin expression induced by salidroside were partially
prevented by HSP70 overexpression.
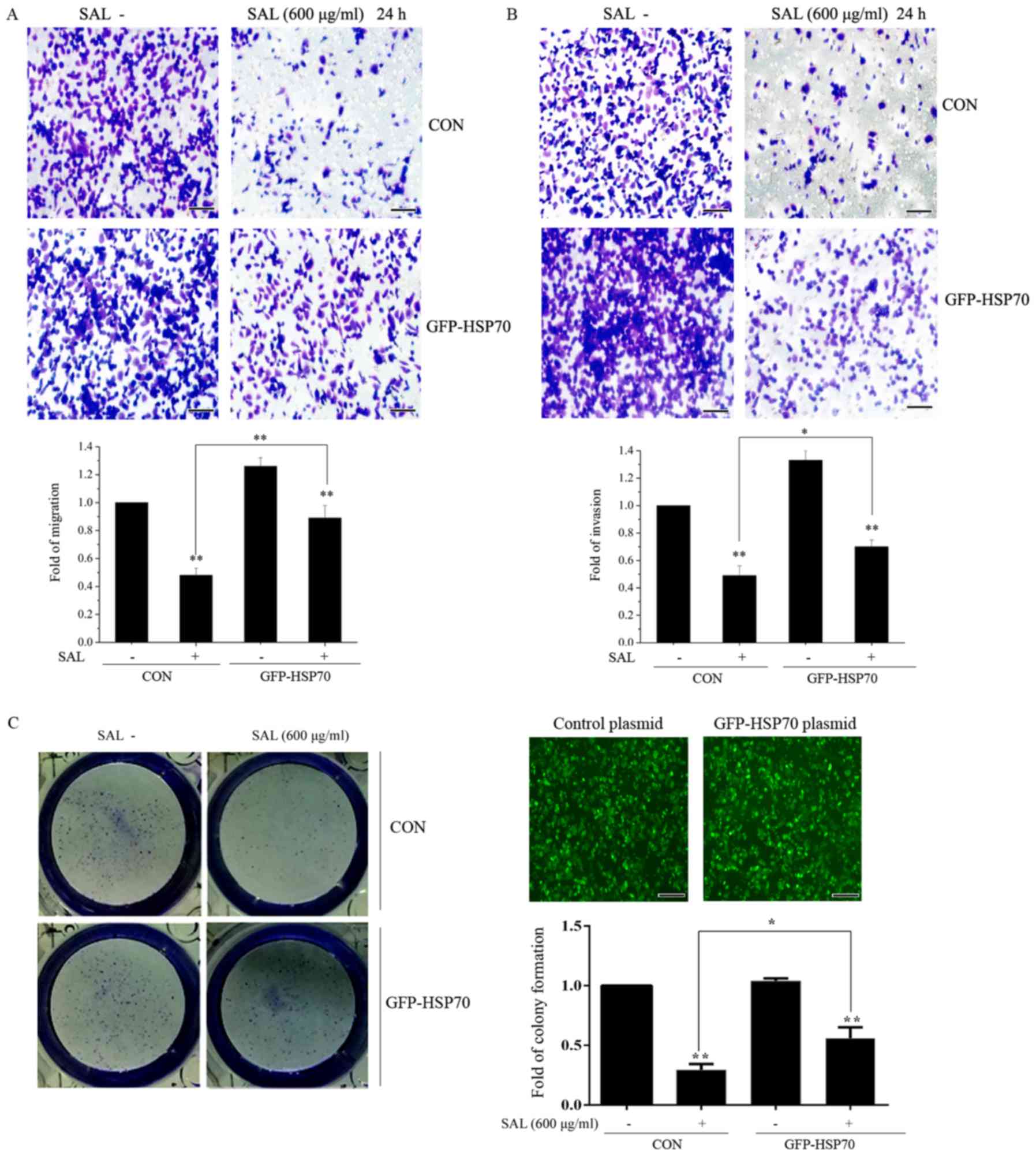
HSP70 suppresses the effects of
salidroside on colony formation, migration and invasion in BGC-823
cells
The effect of HSP70 on colony formation, migration
and invasion was investigated. BGC-823 cells were transfected with
control or HSP70 overexpression plasmids for 24 h. Following
transfection, cells were treated with salidroside for a further 24
h and colony formation, migration and invasion were subsequently
examined. As presented in Fig. 6A and
B, it was demonstrated that salidroside significantly
suppressed the migration and invasion of BGC-823 cells in control
plasmid transfected BGC-823 cells. However, the inhibitory effects
induced by salidroside were prevented in BGC-823 cells transfected
with HSP70 overexpression plasmid. Salidroside significantly
attenuated colony formation in control plasmid transfected BGC-823
cells (Fig. 6C). This inhibitory
effect was reversed by HSP70 overexpression. Taken together, these
results suggested that HSP70 may have antagonized the effects of
salidroside on BGC-823 cell proliferation and migration. Taken
together, the results demonstrated that the inhibition of HSP70
expression may be a molecular mechanism by which salidroside
suppresses BGC-823 cell proliferation, migration and invasion.
Discussion
Salidroside has been reported to have anti-tumor
effects in vitro and in vivo (11,20).
In the present study, it was demonstrated that salidroside
inhibited the proliferation, colony formation, migration and
invasion of BGC-823 cells. The potential mechanisms may be
associated with the inhibitory effects of salidroside on
ROS-mediated and Src-associated signaling pathways, as well as
HSP70 expression.
Inhibition of tumor growth is an important aim in
all strategies used to prevent tumor progression. Dysregulated cell
proliferation is a hallmark of cancer development (24). In the present study, it was
confirmed that salidroside, a bioactive component extracted from
Rhodiola rosea, had an inhibitory effect on the cell
viability of BGC-823 cells through a CCK-8 assay, and this
inhibitory effect was concentration-dependent. In addition,
salidroside was demonstrated to inhibit BGC-823 cell colony
formation, particularly at a dose of 600 µg/ml.
Increasing evidence has revealed that salidroside
inhibits the migration and invasion of various tumors (7,20).
Consistent with these findings, salidroside treatment significantly
suppressed BGC-823 cell migration and invasion in the present
study, in a dose-dependent manner. MMPs have a key role in tumor
cell invasion, migration and tumor angiogenesis (25). MMPs are a family of proteolytic
enzymes that facilitate tumor cell migration by degrading the
basement membrane and other components of the extracellular matrix
(26). MMP-2 and MMP-9 are
important members of the MMP family. Downregulation of MMP-2 and
MMP-9 expression inhibits cancer cell invasion and metastasis
(27,28). Previous reports have demonstrated
that salidroside markedly suppresses MMP-2 and MMP-9 expression
(7,20). Consistent with these results, the
present study demonstrated that salidroside decreased MMP-2 and
MMP-9 expression in BGC-823 cells, suggesting that salidroside may
have reduced the metastatic capabilities of BGC-823 cells via
suppression of MMP-2 and MMP-9 expression. As oxidative stress may
induce tumorcell migration and invasion through the upregulation of
MMP expression (20), the effects
of salidroside on ROS levels were investigated. It was revealed
that salidroside treatment significantly inhibited intracellular
ROS generation in a dose-dependent manner.
EMT is a fundamental biological process in which
epithelial cells undergo a dramatic remodeling of the cytoskeleton,
lose basal-apical polarity and acquire an increased capacity to
metastasize to distant organs (29–31).
Alterations in the expression of EMT-associated markers, including
a decrease in E-cadherin and an increase in N-cadherin, are closely
associated with the invasive and metastatic capacity of cancer
cells (29). The results of the
present study confirmed that salidroside treatment enhanced the
expression of E-cadherin and reduced the expression of
N-cadherin.
A previous study indicated that Src activation
governs several pathways, including those involved in survival,
angiogenesis, proliferation, migration and invasion, through a
variety of proteins, including
phosphatidiylinositol-4,5-biphosphate 3-kinase (PI3K)/Akt, STAT3,
ERK and FAK (32). In addition,
Src activation is associated with an epidermal to mesenchymal-like
transition in models of epithelial cancer (22). Therefore, the effects of
salidroside on the Src-associated signaling pathway proteins were
further investigated in the current study. Salidroside treatment
significantly inhibited the phosphorylation of Src, Akt, ERK, STAT3
and FAK, suggesting that salidroside may have inhibited the
migration and invasion of BGC-823 cells via suppression of
ROS-mediated Src-associated signaling pathway activation.
One of the most effective markers for the detection
of tumor cells is HSP70. HSP70 expression is positively correlated
with cell proliferation and metastasis. Therefore, inhibition of
HSP70 expression may serve as a target for cancer therapy (19). In the present study, whether the
inhibitory effects of salidroside on migration, invasion and
proliferation were partly associated with HSP70 expression was
investigated. The results revealed that salidroside treatment
significantly attenuated HSP70 expression. Overexpression of HSP70
reversed the inhibitory effects of salidroside on EMT marker
expression, as well as the proliferation, migration and invasion of
BGC-823 cells. Taken together, these results suggested that HSP70
expression downregulation may be another possible molecular
mechanism by which salidroside suppressed the proliferation,
migration and invasion of BGC-823 cells. Transforming growth factor
(TGF)-β/mothers against decapentaplegic homologs (Smads) signaling
is thought to be important in the promotion of EMT in human breast
cancer (33). Additionally, a
previous report revealed that HSP70 inhibits high glucose-induced
EMT by modulating Smad expression and activation in peritoneal
mesothelial cells (34).
Furthermore, HSP70 increases the cellular defense capacity through
inhibition of TGF-β/Smad and ERK signaling pathways, thereby
protecting peritoneal mesothelial cells from advanced glycation end
products-induced EMT (35).
Collectively, these reports suggest that the potential molecular
mechanism by which HSP70 regulates EMT may be through Smad
signaling pathways. However, whether HSP70 regulated EMT in BGC-823
cells through Smad is still unclear. In the future, the authors of
the present study will investigate the effect of HSP70 on Smad
signaling pathway activation in gastric cancer.
The present study indicated that HSP70 and
Src-associated signaling pathways may have contributed to the
proliferation and metastasis of BGC-823 cells. However, the
association between HSP70 and these pathways requires further
clarification. Resveratrol has been reported to downregulate HSP70
expression through modulation of Akt and ERK1/2 pathways in chronic
myelogenous leukemia cells (36).
In addition, it has been demonstrated that the PI3K/Akt signaling
pathway is involved in the induction of HSP70 expression and that
blockade of the PI3K/Akt pathway enhances the sensitivity of Raji
cells to chemotherapy through downregulation of HSP70 (37). Banerjee Mustafi et al
(38) reported that heat stress
upregulates the expression of HSP70 through a ROS-mediated p38
mitogen activated protein kinase-Akt signaling pathway.
Furthermore, Src activation governs a variety of pathways,
including PI3K/Akt, STAT3, ERK and FAK (32). Therefore, based on the
aforementioned results, it was hypothesized that a potential
mechanism by which salidroside inhibited the proliferation and
migration of BGC-823 cells in the present study may be through
HSP70 downregulation via suppression of ROS-mediated Src-associated
signaling pathway activation (Fig.
7).
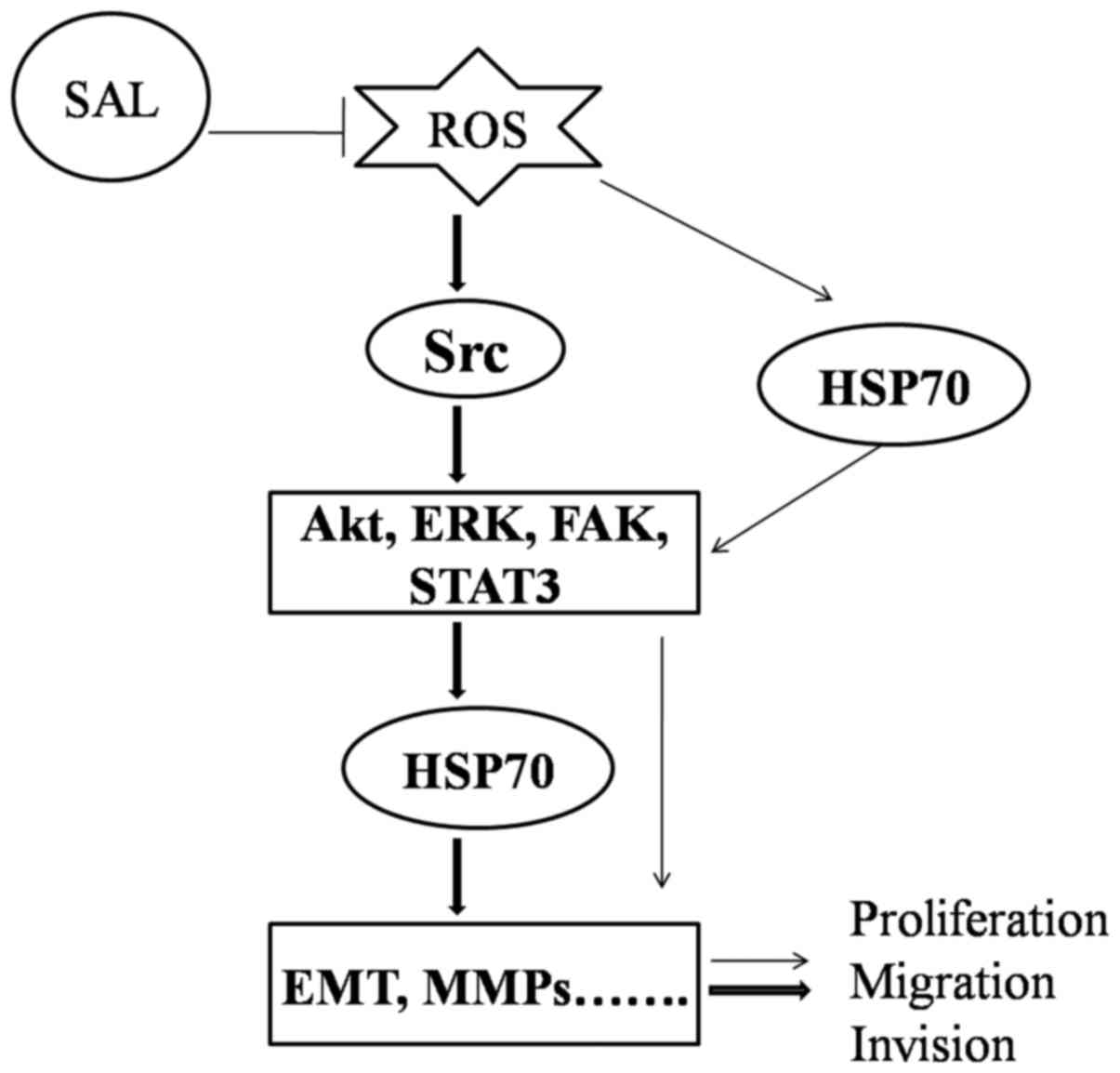 | Figure 7.Schematic diagram illustrating the
signaling pathways involved in the inhibitory effect of salidroside
on biological function, via Src-associated signaling pathways and
HSP70 expression. SAL, salidroside; ROS, reactive oxygen species;
Src, proto-oncogene tyrosine-protein kinase Src; HSP70, heat shock
protein 70; Akt, protein kinase B; STAT3, signal transducer and
activator of transcription 3; ERK, mitogen-activated protein kinase
1; FAK, focal adhesion kinase 1; EMT, epithelial-mesenchymal
transition; MMP, matrix metalloproteinase. |
Budina-Kolomets et al (18) revealed that p-FAK is a client
protein of HSP70, and inhibition of HSP70 may suppress
FAK-dependent invasion in human melanoma cells (18). In addition, Diao et al
(39) reported that exosomal HSP70
expression triggers STAT3 phosphorylation in myeloid-derived
suppressor cells. Based on these findings and the results of the
present study, it was theorized that salidroside may have also
inhibited the proliferation and migration of BGC-823 cells through
the downregulation of HSP70 expression, followed by suppression of
the Src-mediated phosphorylation of FAK and STAT3 (Fig. 7). However, the present study was
unable to obtain clear evidence of the association between HSP70
and Src-associated signaling, which will be investigated in future
research.
In conclusion, the results of the present study
demonstrated that salidroside significantly inhibited BGC-823 cell
proliferation, migration and invasion. Additionally, salidroside
treatment inhibited ROS-mediated Src-associated signaling pathway
protein phosphorylation and HSP70 expression. Taken together, these
data suggested that salidroside suppressed the proliferation,
migration and invasion of BGC-823 cells, at least partially through
ROS-activated Src-associated signaling pathways and HSP70. The
present study provides novel insights into the antitumor effects of
salidroside in gastric cancer.
Acknowledgements
Not applicable.
Funding
This study was supported by the National Nature
Science Foundation of China (grant no. 81601380), Natural Science
Research Project of Anhui Colleges and Universities (grant no.
KJ2016SD59), Outstanding Young Talent Support Program Key Projects
in Anhui Colleges and Universities (grant no. gxyqZD2016173) and
Anhui Province Key Laboratory of Active Biological Macromolecules
(grant no. 1306C083008).
Availability of data and materials
The data and materials during the current study are
available from the corresponding author on reasonable request.
Author's contributions
Conceived and designed the experiments: ZQ and YZ.
Performed the experiments: ZQ, TT, LS and YM. Analyzed the data:
SQ, LL, YL and LY. Wrote the paper: ZQ.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
DMEM
|
Dulbecco's modified Eagle's medium
|
|
HSP70
|
heat shock protein 70
|
|
GFP-HSP70
|
green fluorescent protein-tagged
HSP70
|
|
EMT
|
epithelial-mesenchymal transition
|
|
ROS
|
reactive oxygen species
|
References
|
1
|
Zhang L, Xu Z, Xu X, Zhang B, Wu H, Wang
M, Zhang X, Yang T, Cai J, Yan Y, et al: SALL4, a novel marker for
human gastric carcinogenesis and metastasis. Oncogene.
33:5491–5500. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wang L, Chen X, Du Z, Li G, Chen M, Liang
G and Chen T: Curcumin suppresses gastric tumor cell growth via
ROS-mediated DNA polymerase gamma depletion disrupting cellular
bioenergetics. J Exp Clin Cancer Res. 36:472017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Brenner H, Rothenbacher D and Arndt V:
Epidemiology of stomach cancer. Methods Mol Biol. 472:467–477.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
van Hagen P, Hulshof MC, van Lanschot JJ,
Steyerberg EW, van Berge Henegouwen MI, Wijnhoven BP, Richel DJ,
Nieuwenhuijzen GA, Hospers GA, Bonenkamp JJ, et al: Preoperative
chemoradiotherapy for esophageal or junctional cancer. N Engl J
Med. 366:2074–2084. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Karimi P, Islami F, Anandasabapathy S,
Freedman ND and Kamangar F: Gastric cancer: Descriptive
epidemiology, risk factors, screening, and prevention. Cancer
Epidemiol Biomarkers Prev. 23:700–713. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sun KX, Xia HW and Xia RL: Anticancer
effect of salidroside on colon cancer through inhibiting JAK2_STAT3
signaling pathway. Int J Clin Exp Pathol. 8:615–621.
2015.PubMed/NCBI
|
|
7
|
Zhao G, Shi A, Fan Z and Du Y: Salidroside
inhibits the growth of human breast cancer in vitro and in vivo.
Oncol Rep. 33:2553–2560. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Paterson I and Anderson EA: Chemistry. The
renaissance of natural products as drug candidates. Science.
310:451–453. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Qi Z, Qi S, Ling L, Lv J and Feng Z:
Salidroside attenuates inflammatory response via suppressing
JAK2-STAT3 pathway activation and preventing STAT3 transfer into
nucleus. Int Immunopharmacol. 35:265–271. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Guan S, Feng H, Song B, Guo W, Xiong Y,
Huang G, Zhong W, Huo M, Chen N, Lu J and Deng X: Salidroside
attenuates LPS-induced pro-inflammatory cytokine responses and
improves survival in murine endotoxemia. Int Immunopharmacol.
11:2194–2199. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Fan XJ, Wang Y, Wang L and Zhu M:
Salidroside induces apoptosis and autophagy in human colorectal
cancer cells through inhibition of PI3K/Akt/mTOR pathway. Oncol
Rep. 36:3559–3567. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang L, Ding W, Sun H, Zhou Q, Huang J,
Li X, Xie Y and Chen J: Salidroside protects PC12 cells from
MPP(+)-induced apoptosis via activation of the PI3K/Akt pathway.
Food Chem Toxicol. 50:2591–2597. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu Z, Li X, Simoneau AR, Jafari M and Zi
X: Rhodiola rosea extracts and salidroside decrease the growth of
bladder cancer cell lines via inhibition of the mTOR pathway and
induction of autophagy. Mol Carcinog. 51:257–267. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Forman HJ, Torres M and Fukuto J: Redox
signaling. Mol Cell Biochem. 234–235:49–62. 2002. View Article : Google Scholar
|
|
15
|
Storz P: Reactive oxygen species in tumor
progression. Front Biosci. 10:1881–1896. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang J, Li JZ, Lu AX, Zhang KF and Li BJ:
Anticancer effect of salidroside on A549 lung cancer cells through
inhibition of oxidative stress and phosphor-p38 expression. Oncol
Lett. 7:1159–1164. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Okon IS and Zou MH: Mitochondrial ROS and
cancer drug resistance: Implications for therapy. Pharmacol Res.
100:170–174. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Budina-Kolomets A, Webster MR, Leu JI,
Jennis M, Krepler C, Guerrini A, Kossenkov AV, Xu W, Karakousis G,
Schuchter L, et al: HSP70 inhibition limits FAK-dependent invasion
and enhances the response to melanoma treatment with BRAF
inhibitors. Cancer Res. 76:2720–2730. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ciocca DR and Calderwood SK: Heat shock
proteins in cancer: Diagnostic, prognostic, predictive, and
treatment implications. Cell Stress Chaperones. 10:86–103. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sun C, Wang Z, Zheng Q and Zhang H:
Salidroside inhibits migration and invasion of human fibrosarcoma
HT1080 cells. Phytomedicine. 19:355–363. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sutton P, Borgia JA, Bonomi P and Plate
JM: Lyn, a Src family kinase, regulates activation of epidermal
growth factor receptors in lung adenocarcinoma cells. Mol Cancer.
12:762013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Summy JM and Gallick GE: Treatment for
advanced tumors: SRC reclaims center stage. Clin Cancer Res.
12:1398–1401. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ablin RJ, Owen S and Jiang WG: Prostate
transglutaminase (TGase-4) induces epithelial-to-mesenchymal
transition in prostate cancer cells. Anticancer Res. 37:481–487.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lv C, Huang Y, Liu ZX, Yu D and Bai ZM:
Salidroside reduces renal cell carcinoma proliferation by
inhibiting JAK2/STAT3 signaling. Cancer Biomark. 17:41–47. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yan L, Lin B, Gao L, Gao S, Liu C, Wang C,
Wang Y, Zhang S and Iwamori M: Lewis (y) antigen overexpression
increases the expression of MMP-2 and MMP-9 and invasion of human
ovarian cancer cells. Int J Mol. Sci. 11:4441–4452. 2010.
|
|
26
|
Rajoria S, Suriano R, George A, Shanmugam
A, Schantz SP, Geliebter J and Tiwari RK: Estrogen induced
metastatic modulators MMP-2 and MMP-9 are targets of
3,3′-diindolylmethane in thyroid cancer. PloS One. 6:e158792011.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lai WW, Hsu SC, Chueh FS, Chen YY, Yang
JS, Lin JP, Lien JC, Tsai CH and Chung JG: Quercetin inhibits
migration and invasion of SAS human oral cancer cells through
inhibition of NF-κB and matrix metalloproteinase-2/-9 signaling
pathways. Anticancer Res. 33:1941–1950. 2013.PubMed/NCBI
|
|
28
|
Braicu EI, Gasimli K, Richter R, Nassir M,
Kümmel S, Blohmer JU, Yalcinkaya I, Chekerov R, Ignat I, Ionescu A,
et al: Role of serum VEGFA, TIMP2, MMP2 and MMP9 in monitoring
response to adjuvant radiochemotherapy in patients with primary
cervical cancer-results of a companion protocol of the randomized
NOGGO-AGO phase III clinical trial. Anticancer Res. 34:385–391.
2014.PubMed/NCBI
|
|
29
|
Liu L, Zhang J, Yang X, Fang C, Xu H and
Xi X: SALL4 as an epithelial-mesenchymal transition and drug
resistance inducer through the regulation of c-Myc in endometrial
cancer. PloS One. 10:e01385152015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Weinberg RA: Mechanisms of malignant
progression. Carcinogenesis. 29:1092–1095. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mani SA, Guo W, Liao MJ, Eaton EN, Ayyanan
A, Zhou AY, Brooks M, Reinhard F, Zhang CC, Shipitsin M, et al: The
epithelial-mesenchymal transition generates cells with properties
of stem cells. Cell. 133:704–715. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lin SY, Chang HH, Lai YH, Lin CH, Chen MH,
Chang GC, Tsai MF and Chen JJ: Digoxin suppresses tumor malignancy
through inhibiting multiple Src-related signaling pathways in
non-small cell lung cancer. PloS One. 10:e01233052015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Smith AL, Iwanaga R, Drasin DJ, Micalizzi
DS, Vartuli RL, Tan AC and Ford HL: The miR-106b-25 cluster targets
Smad7, activates TGF-β signaling and induces EMT and tumor
initiating cell characteristics downstream of Six1 in human breast
cancer. Oncogene. 31:5162–5171. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liu J, Bao J, Hao J, Peng Y and Hong F:
HSP70 inhibits high glucose-induced Smad3 activation and attenuates
epithelial-to-mesenchymal transition of peritoneal mesothelial
cells. Mol Med Rep. 10:1089–1095. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yang J, Zhu T, Liu X, Zhang L, Yang Y,
Zhang J and Guο M: Heat shock protein 70 protects rat peritoneal
mesothelial cells from advanced glycation end-products-induced
epithelial-to-mesenchymal transition through mitogen activated
protein kinases/extracellular signal-regulated kinases and
transforming growth factor-β/Smad pathways. Mol Med Rep.
11:4473–4481. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Mustafi Banerjee S, Chakraborty PK and
Raha S: Modulation of Akt and ERK1/2 pathways by resveratrol in
chronic myelogenous leukemia (CML) cells results in the
downregulation of Hsp70. PloS One. 5:e87192010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Fang X, Jiang Y, Feng L, Chen H, Zhen C,
Ding M and Wang X: Blockade of PI3K/AKT pathway enhances
sensitivity of Raji cells to chemotherapy through down-regulation
of HSP70. Cancer Cell Int. 13:482013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Mustafi Banerjee S, Chakraborty PK, Dey RS
and Raha S: Heat stress upregulates chaperone heat shock protein 70
and antioxidant manganese superoxide dismutase through reactive
oxygen species (ROS), p38MAPK, and Akt. Cell Stress Chaperones.
14:579–589. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Diao J, Yang X, Song X, Chen S, He Y, Wang
Q, Chen G, Luo C, Wu X and Zhang Y: Exosomal Hsp70 mediates
immunosuppressive activity of the myeloid-derived suppressor cells
via phosphorylation of Stat3. Med Oncol. 32:4532015. View Article : Google Scholar : PubMed/NCBI
|