Introduction
The tendon is a vital component of the
musculoskeletal system. Its strong mechanical properties enable it
to transmit muscle-contraction force to the skeleton in order to
maintain posture or produce motion (1). This property is a consequence of its
highly organized structure, consisting of hypocellular connective
tissue arranged in a specific spatial organization of type I
collagen (Col1) fibers (2). Tendon
injuries are common, and account for nearly one third of all
musculoskeletal conditions (3). In
addition, the repair process of a tendon injury is slow, and
commonly results in scar tissue and incomplete recovery (4). Therefore, a full understanding of the
pathogenesis and healing mechanism of tendon repair is urgently
required. Degeneration is considered the intrinsic pathological
mechanism of chronic tendon injury or tendinopathies (5). Previous studies have highlighted the
importance of inflammatory cell infiltration and inflammatory
cytokine gene expression in animal and human tendon disease
(6,7), indicating that inflammation may have
an important role in the tendon healing process (8).
It has been demonstrated that interleukin (IL)-10
gene expression is significantly upregulated 2 weeks following the
occurrence of Achilles tendon rupture in humans, between the
inflammatory and the proliferative phases during the healing
process (9,10). Notably, in vivo
overexpression of IL-10 has been demonstrated to significantly
increase the maximum stress in a tendon-healing model (11). In other tissues and cells, IL-10
has been demonstrated to: i) provide pro-survival cues to
melanocytes by exerting anti-apoptotic effects (12); ii) inhibit bone marrow fibroblast
progenitor cells from homing and transdifferentiating into
myofibroblasts, thereby modulating cardiac fibrosis (13); and iii) reduce type I collagen in
cultured human skin fibroblasts (14). However, the exact impact of
upregulated IL-10 gene expression on injured tendons has not been
fully elucidated.
Recently, tendon-derived stem cells (TDSCs) have
been identified in various species including humans, rabbits, rats
and mice (15–17). The characteristic properties of
stem cells, including proliferation, cloning and multipotency,
allow them to differentiate into tendon-like tissues in
vitro and/or in vivo (15). A previous study indicated that
TDSCs form tendon-like tissues in nude mouse or nude rat models
(17), which suggests that TDSCs
may contribute to tendon repair. To understand how inflammatory
cytokines impact the regenerative and degenerative potentials of
TDSCs, the present study focused on IL-10, a cytokine that is
upregulated in injured tendons, and examined the effects of IL-10
on the function of TDSCs.
Materials and methods
Animals
All aspects of the research were approved by the
Institutional Animal Care and Use Committee of Nanfang Hospital,
Southern Medical University (Guangzhou, China). Female
Sprague-Dawley rats (n=2; 6-weeks-old; 170–200 g) were purchased
from the Laboratory Animal Center of Southern Medical University
(Guangzhou, China).
Isolation of TDSCs
TDSCs were isolated from the Achilles tendons of
Sprague-Dawley rats as previously reported (18,19).
Briefly, rats were anesthetized via an intramuscular injection of
pentobarbital (30 mg/kg) and were subsequently sacrificed.
Following this, the Achilles tendons were dissected and incubated
in 600 U/ml (3 mg/ml) type I collagenase (cat. no. C0130;
Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) and PBS for 2 h at
37°C with gentle shaking. The dissociated cells were plated at a
density of 140 cells/cm2 in 100 mm dishes and cultured
in Dulbecco's modified Eagle's medium (DMEM) containing 20% fetal
bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc., Waltham,
MA, USA), 100 U/ml penicillin and 100 µg/ml streptomycin (Gibco;
Thermo Fisher Scientific, Inc.) for 8–10 days at 5% CO2
and 37°C. TDSCs at passage 3 or 4 were used in the subsequent
experiments. The stem cell characteristics of TDSCs, including the
proliferation, clonogenicity and multi-lineage differentiation
potential, were confirmed prior to use in subsequent experiments
using standard assays, including colony-forming unit fibroblast
assays, Oil red O or Alizarin red staining and Alcian blue
staining, as described previously (19).
Cell proliferation assay
To perform the cell proliferation assay, TDSCs were
plated at a density of 103 cells/well in a 96-well plate
cultured in DMEM containing 10% FBS and allowed to adhere overnight
at 5% CO2 and 37°C. Following this, DMEM containing 10%
FBS with 0.1, 1, 10 and 100 ng/ml rat IL-10 (cat. no. 400-19;
PeproTech, Inc., Rocky Hill, NJ, USA) was added to TDSCs, which
were then cultured for 1, 3 or 5 days at 37°C. Untreated cells
cultured for 1, 3 or 5 days at 37°C were treated as the control.
TDSC proliferation was subsequently determined using a Cell
Counting Kit-8 assay (CCK-8; cat. no. KL640; Dojindo Molecular
Technologies, Inc., Kumamoto, Japan) according to previously
published protocol (20,21).
Cell cycle analysis
Based on the results of aforementioned cell
proliferation assays, TDSCs that were either untreated or treated
with IL-10 (10 ng/ml) cultured in DMEM containing 10% FBS for 3
days were washed once in PBS and fixed with 500 µl cold 70% ethanol
in PBS for 2 h at 4°C. TDSCs were centrifuged at 800 × g at 4°C for
5 min and washed again in PBS, then resuspended in 100 µl RNase A
(Nanjing KeyGen Biotech Co., Ltd., Nanjing, China) and incubated at
37°C for 30 min. Following this, TDSCs were incubated with 400 µl
propidium iodide (PI; Nanjing KeyGen Biotech Co., Ltd.) at 4°C for
30 min, and analyzed with a flow cytometer (FACScan; BD
Biosciences, San Jose, CA, USA) and FlowJo 7.6.1 software (FlowJo
LLC, Ashland, OR, USA).
Wound healing assay
Cell migration was determined with a wound-healing
assay, as previously described (22). TDSCs were grown to 100% confluence
in 6-well plates and treated with IL-10 (10 ng/ml) or DMEM
containing 2% FBS at 37°C. A sterile P200 pipette tip was used to
create a scratch across the cell monolayer. Cultures were
subsequently washed once with 1 ml growth medium to remove the
damaged and detached cells. Following medium replacement, TDSCs
were cultured for 24 h at 37°C. Cell cultures were examined with a
phase-contrast microscope (Olympus Corporation, Tokyo, Japan), and
images of the entire strip, including three from the same scratch
fields, were acquired at 0, 12 and 24 h after cells were scratched.
The recovered gap area of the entire strip was measured at
different time points using ImageJ 1.8.0 software (National
Institutes of Health, Bethesda, MD, USA). The migration rate was
based on the calculated recovered gap area and was expressed as a
percentage.
Signal transducer and activator of
transcription 3 (Stat3) inhibitor application
To demonstrate that IL-10 exerts its functions
through the janus kinase (JAK)/Stat3 signaling pathway, TDSCs were
plated at a density of 4×105/well on a 60 mm dish,
cultured in DMEM containing 10% FBS and allowed to adhere overnight
at 5% CO2 and 37°C. TDSCs were subsequently cultured in
DMEM containing 10% FBS, supplemented with or without 10 ng/ml
IL-10, and with or without 5 µM Stat3 inhibitor (WP1066; cat. no.
S2796; Selleck Chemicals, Shanghai, China) for 3 days at 37°C.
Following the above treatments, cells were collected and used in
further experiments.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
To investigate the effect of IL-10 on spontaneous
tenogenic differentiation, spontaneously differentiated tenocytes
were used as the control group, in which TDSCs were treated with
basic culture medium without IL-10 for 3 days. Following the
treatment of cells with 0, 0.1, 1, 10 or 100 ng/ml IL-10 for 3
days, total RNA was isolated from TDSCs with TRIzol reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocols. Following this, total RNA was
reverse-transcribed into cDNA using the PrimeScript™ RT
Master Mix kit (cat. no. RR036A; Takara Biotechnology Co., Ltd.,
Dalian, China). Reactions were incubated in a
LightCycler® 480 system (Roche Diagnostics, Basel,
Switzerland) for 15 min at 37°C, followed by 5 sec at 85°C and then
cooling to 4°C. The resulting cDNA was subjected to qPCR in a
LightCycler® 480 system with SYBR green reagent (cat.
no. AK8307; Takara Biotechnology Co., Ltd.). The thermocycling
conditions used were as follows: Pre-denaturation at 95°C for 30
sec; followed by 40 cycles of denaturation at 95°C for 5 sec, and
annealing and extension at 60°C for 30 sec. The mean Cq value was
calculated from triplicate reactions. The relative expression
levels of scleraxis (Scx), Col1, tenomodulin (Tnmd), collagen type
3 (Col3), mohawk (Mkx), early growth response gene 1 (Egr1),
fibromodulin (Fmod), lumican (Lum), decorin (Dcn) and biglycan
(Bgn) were calculated based on a standard curve and normalized to
GAPDH (23). The primer
sequences used are listed in Table
I.
 | Table I.Primers used for reverse
transcription-quantitative polymerase chain reaction. |
Table I.
Primers used for reverse
transcription-quantitative polymerase chain reaction.
| Gene | Forward
primers | Reverse
primers | NCBI Accession
no.a |
|---|
| GAPDH |
5′-AAGCTCATTTCCTGGTATGACA-3′ |
5′-TCTTACTCCTTGGAGGCCATGT-3′ | NM_017008.4 |
| Scx |
5′-AGAACACCCAGCCCAAACA-3′ |
5′-CGGTCTTTGCTCAACTTTCT-3′ | NM_001130508 |
| Col1 |
5′-GTGCTAAGGGTGAAGCTGGT-3′ |
5′-CATCAGCACCAGGGTTTCCAG-3′ | NM_053304 |
| Tnmd |
5′-GTCACATTCTAAATGCAGAAG-3′ |
5′-CTCCCCCAAAACAGGACAAT-3′ | NM_022290 |
| Col3 |
5′-CTGGAGATAAGGGTGAAGGT-3′ |
5′-GAGGGCCTCCTTCACCTTTCT-3′ | NM_032085 |
| Mkx |
5′-CTATCGCACAGGTAAGCCCA-3′ |
5′-CCCACGTATCAGTTTCTCCCA-3′ | XM_017600733 |
| Egr1 |
5′-AACAACCCTACGAGCACCTG-3′ |
5′-ACCAGCGCCTTCTCGTTATT-3′ | NM_012551 |
| Fmod |
5′-CCCGTGATTGTCCCCAAGAA-3′ |
5′-CAGGTACTTGAGGTTGCGGT-3′ | NM_080698 |
| Lum |
5′-GCTTCACCGGGCTTCAATAC-3′ |
5′-AAATGAGTTTCCAGGCACGC-3′ | NM_031050 |
| Dcn |
5′-CCTAAAGGAGCTGCCCGAAA-3′ |
5′-GCCGCCCAGTTCTATGACAA-3′ | NM_024129 |
| Bgn |
5′-CTGCATTGAGATGGGTGGGA-3′ |
5′-GGTAGTTGAGCTTCAGGCCA-3′ | NM_017087 |
Immunofluorescence staining
Following the aforementioned treatments, Col1
immunofluorescence staining was performed on the cell monolayer in
a two-step procedure. Specifically, Cells were cultured until a
40–60% confluence was reached prior to immunofluorescence staining.
Following this, cells were fixed in 4% paraformaldehyde at room
temperature for 10 min, lysed using 0.5% Triton X-100 and then
blocked with 5% bovine serum albumin (cat. no. 36101ES25; Shanghai
Yusheng Biotechnology Co., Ltd., Shanghai, China) at room
temperature for 30 min. Cells were then incubated overnight at 4°C
with primary rabbit anti-Col1 antibodies (1:100; cat. no.
14695-1-AP; ProteinTech Group, Inc., Chicago, IL, USA) and
subsequently incubated with tetramethylrhodamine-tagged goat
anti-rabbit IgG secondary antibodies (1:100; cat. no. HA1016;
Hangzhou HuaAn Biotechnology Co., Ltd., Hangzhou, China) for 2 h at
room temperature. Following this, cells were incubated with
4′,6-diamidino-2-phenylindole at room temperature for 5 min prior
to being photographed using a fluorescence microscope (Olympus
BX51; magnification, ×100; Olympus Corporation, Tokyo, Japan).
Western blot analysis
TDSCs were collected and radioimmunoprecipitation
assay lysis buffer was used to extract total cell protein. Protein
concentration in the samples was measured using a bicinchoninic
acid protein assay kit (Nanjing KeyGen Biotech Co., Ltd.). Protein
samples (25 µg) were separated via 10% SDS-PAGE, electrotransferred
onto polyvinylidene fluoride membranes and then blocked using 5%
bovine serum albumin (cat. no. 36101ES25; Shanghai Yusheng
Biotechnology Co., Ltd.) at room temperature 1 h. Following this,
membranes were incubated with primary antibodies [rabbit polyclonal
antibodies against Scx (1:500; cat. no. ab58655; Abcam, Cambridge,
MA, USA), Tnmd (1:500; cat. no. ab203676; Abcam), Col1 (1:1,000;
cat. no. 14695-1-AP; ProteinTech Group, Inc., Chicago, IL, USA),
Col3 (1:1,000; cat. no. 13548-1-AP; ProteinTech Group, Inc.), Stat3
(1:1,000; 10253-2-AP; ProteinTech Group, Inc.), phosphorylated
(p)-Stat3 (1:1,000; Tyr705; cat. no. AF3295; Affinity Biosciences,
Cincinnati, OH, USA), protein kinase B (Akt; 1:500; cat. no.
10176-2-AP; ProteinTech Group, Inc.), p-Akt (1:500; cat. no.
66444-1-Ig; ProteinTech Group, Inc.) and GAPDH [1:5,000; cat. no.
10494-1-AP; ProteinTech Group, Inc.)] overnight at 4°C. Membranes
were subsequently incubated with horseradish peroxidase-conjugated
Affinipure Goat Anti-Rabbit IgG (H+L) secondary antibodies
(1:5,000; cat. no. SA00001-2; ProteinTech Group, Inc.) for 2 h at
room temperature. Proteins were visualized with the
electrochemiluminescence kit (cat. no. P0018; Beyotime Institute of
Biotechnology, Shanghai, China) and images were captured using the
Tanon imaging system (Tanon-5200; Tanon Science and Technology Co.,
Ltd., Shanghai, China).
Statistical analysis
Results were analyzed using the SPSS version 20 (IBM
Corp., Armonk, NY, USA) and expressed as the mean ± standard error
of the mean. Data represents at least three replicates for each
experimental condition. A Student's t-test, or one-way analysis of
variance followed by Dunnett's post-hoc test was used to identify
statistical differences. P<0.05 was considered to indicate a
statistically significant difference.
Results
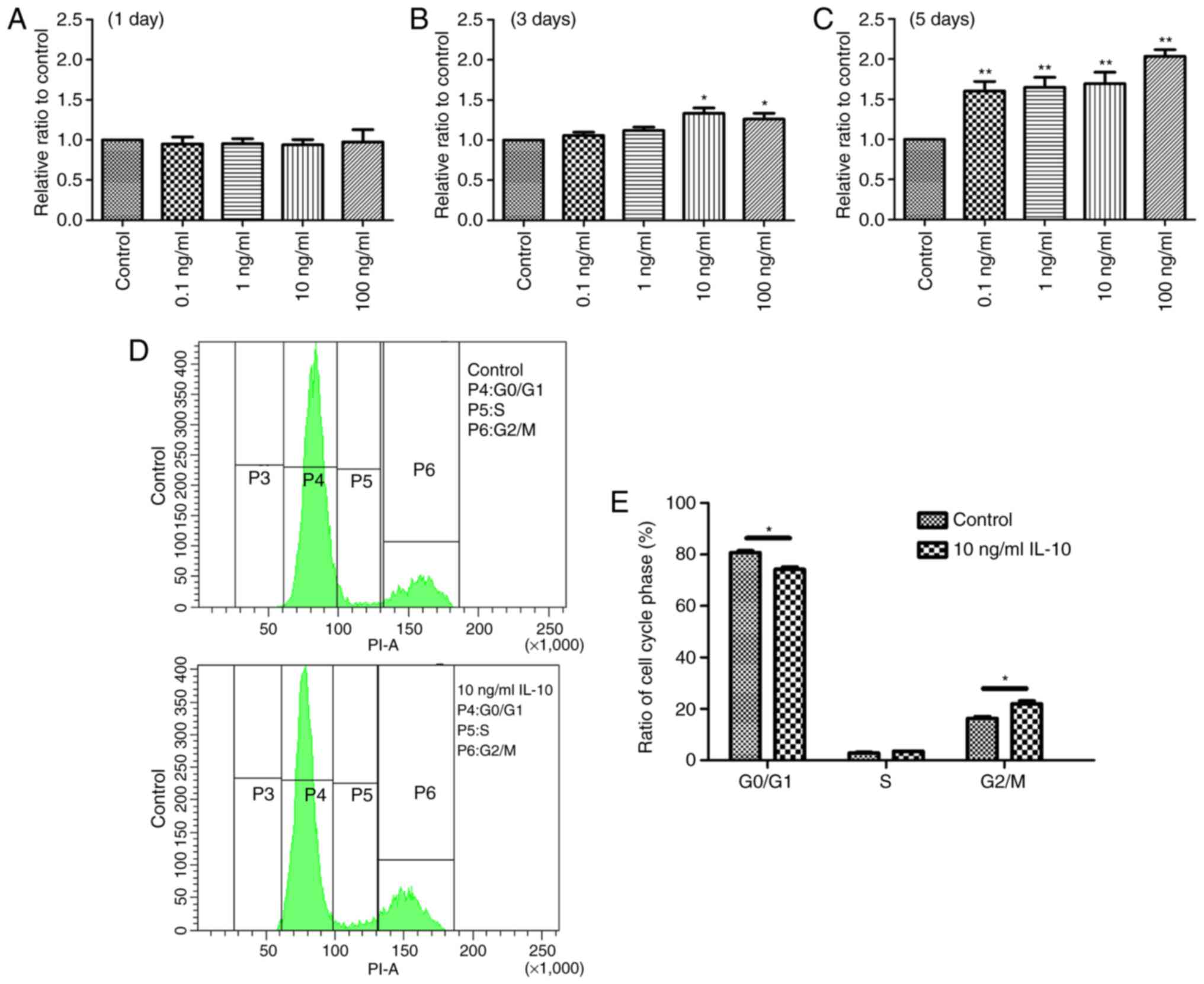
IL-10 stimulates cell proliferation of
TDSCs
The effect of IL-10 on cell proliferation was
initially examined. TDSCs were treated with 0, 0.1, 1, 10 or 100
ng/ml IL-10 for 1, 3 or 5 days, and cell proliferation was
determined with a CCK-8 assay. The results revealed a statistically
significant increase in cell proliferation in the IL-10-treated
cells compared with the controls. At day 1, there was no
significant difference between any groups (Fig. 1A). However, the optical density
values of TDSCs treated with 10 and 100 ng/ml IL-10 were
significantly higher after 3 days compared with the control
(P<0.05; Fig. 1B). At day 5,
all experimental groups exhibited a significant increase in cell
proliferation compared with the control group (Fig. 1C). Based on these results, it was
concluded that a 3 day incubation with 10 ng/ml IL-10 would be the
condition used to induce the desired effects in the subsequent
experiments.
IL-10 leads to cell cycle progression
in TDSCs
IL-10 altered the cell cycle of TDSCs. Following
incubation of TDSCs with 10 ng/ml IL-10 for 3 days, cell cycle
progression was analyzed by flow cytometry. The results
demonstrated that the G1 phase was activated and an increased
number of cells were in the G2/M phase in IL-10-treated cells,
compared with the control (Fig.
1D). In the control and 10 ng/ml IL-10-treated TDSCs, the
percentages of cells in G1 phase was 80.7±0.78 and 74.2±0.95%,
respectively (Fig. 1E), indicating
that IL-10 accelerated G1 phase cell cycle division, compared with
the controls (P<0.05). By contrast, the percentage of control
and 10 ng/ml IL-10-treated TDSCs in the G2/M phase was 16.3±0.71
and 21.9±1.12%, respectively (P<0.05; Fig. 1E). However, the percentage of TDSCs
in the S phase with and without IL-10 treatment was almost
identical.
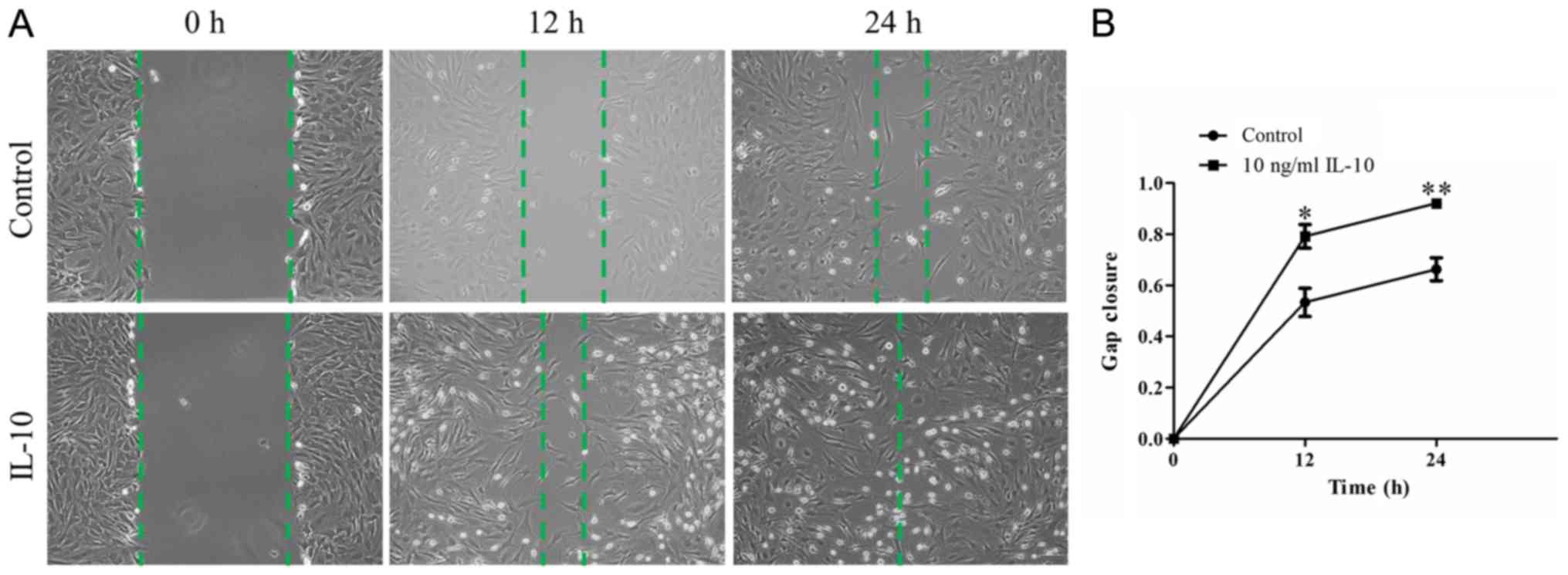
IL-10 promotes TDSC migration
To examine whether IL-10 promotes the repair
capacity of TDSCs, an in vitro wound-healing assay was
performed. A confluent monolayer of TDSCs was wounded and treated
with or without 10 ng/ml IL-10 for 24 h. Microscopy indicated that
wound closure in IL-10-treated TDSCs appeared to be significantly
greater at 12 and 24 h compared with the control cells (Fig. 2A). Quantitative analyses indicated
the cells in the IL-10-treated group had a quicker migration
velocity, compared with that in the control group at the two
different time points. The mean relative gap closure in cells
treated with 0 and 10 ng/ml IL-10 was 53.35±5.54 and 79.18±4.65%
after 12 h, and 66.31±4.50 and 92.10±1.04% after 24 h, respectively
(Fig. 2B; P<0.01).
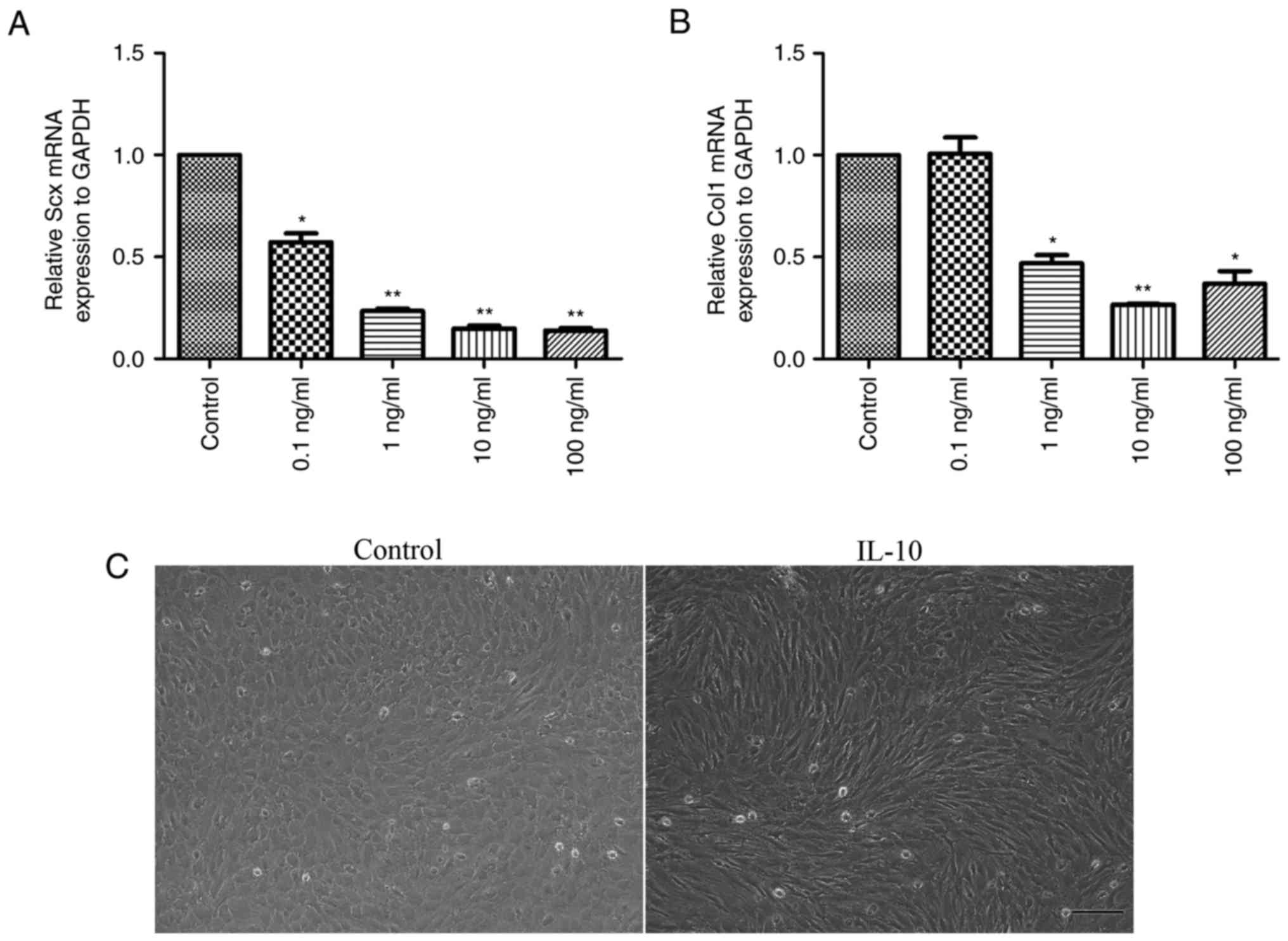
IL-10 suppresses spontaneous tenogenic
differentiation
Following the treatment of cells with 0, 0.1, 1, 10
or 100 ng/ml IL-10 for 3 days, the gene expression levels of tendon
cell markers, Scx and Col1, were analyzed. The results demonstrated
that a significant dose-dependent reduction in the mRNA expression
levels of Scx (Fig. 3A) and Col1
(Fig. 3B) occurred. Notably, cells
treated with 10 or 100 ng/ml IL-10 were suppressed to a larger
extent when compared with the control cells (P<0.01).
Furthermore, it was revealed that cells in the IL-10-treated group
exhibited long spindle-like shapes and orderly arrangement, while
those in control group were ovoid in shape (Fig. 3C).
RT-qPCR analysis revealed that IL-10 significantly
downregulated the gene expression levels of Tnmd, Col3 and Egr1,
Fmod and Lum; however MKx expression was significantly upregulated
compared with the control (Fig.
4A). Additionally, no distinctions were observed in the gene
expression of Dcn and Bgn between the control and IL-10-treated
cells (Fig. 4A).
Immunofluorescence analysis demonstrated that the cytoplasm of the
control group contained more Col1 than that of the IL-10-treated
group (Fig. 4B). At the protein
level, western blot analysis consistently revealed significant
reductions in the expression levels of Col1 (P<0.01), Scx
(P<0.05), Tnmd (P<0.01) and Col3 (P<0.01) in TDSCs
incubated with 10 ng/ml IL-10, compared with the control (Fig. 4C and D).
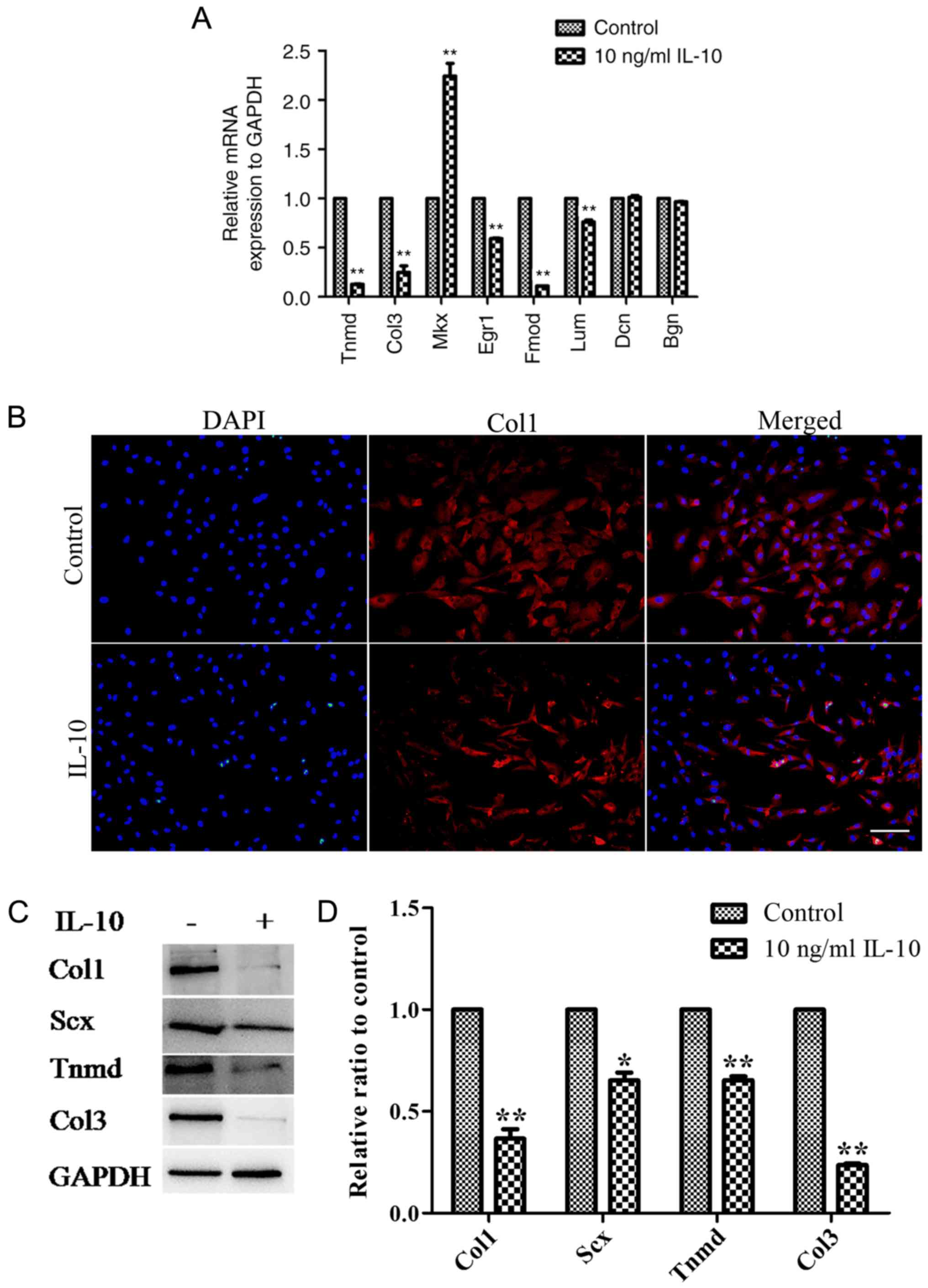 | Figure 4.IL-10 alters the mRNA and protein
expression levels tendon-associated molecules in TDSCs. (A) The
mRNA expression Tnmd, Col3, Mkx, Egr1, Fmod and Lum was altered by
IL-10 treatment. (B) Fluorescent staining of Col1 expression
following treatment with IL-10 for 3 days. (C) Rat TDSCs were
treated with the indicated concentrations of IL-10 for 3 days and
subjected to western blot analysis for Scx, Col1, Tnmd and Col3
protein expression. (D) Densitometric analysis of the western
blotting results. Data are expressed as the mean ± standard error
of the mean. *P<0.05, **P<0.01 vs. control cells. IL-10,
interleukin-10; TDSCs, tendon-derived stem cells; Tnmd,
tenomodulin; Col3, collagen type 3; Mkx, mohawk; Egr1, early growth
response gene 1; Fmod, fibromodulin; Lum, lumican; Dcn, decorin;
Bgn, biglycan. |
IL-10 inhibits spontaneous tenogenic
differentiation of TDSCs by activating the JAK/Stat3 signaling
pathway
In order to investigate the molecular pathways
responsible for the effects of IL-10, the response of TDSCs to
IL-10 (10 ng/ml) was examined. It was demonstrated that Stat3 was
phosphorylated to a significantly higher degree following IL-10
treatment, compared with the control group, although the PI3K/Akt
signaling pathway was not activated (Fig. 5A). These results demonstrated that
TDSCs responded directly to IL-10 stimulation. The effect of IL-10
on the gene expression of tendon cell markers, Col1, Col3 and Tnmd,
was significantly suppressed by treatment of TDSCs with the Stat3
inhibitor, WP1066, which confirmed that IL-10 inhibited spontaneous
tenogenic differentiation of TDSCs by activating the JAK/Stat3
pathway (Fig. 5B).
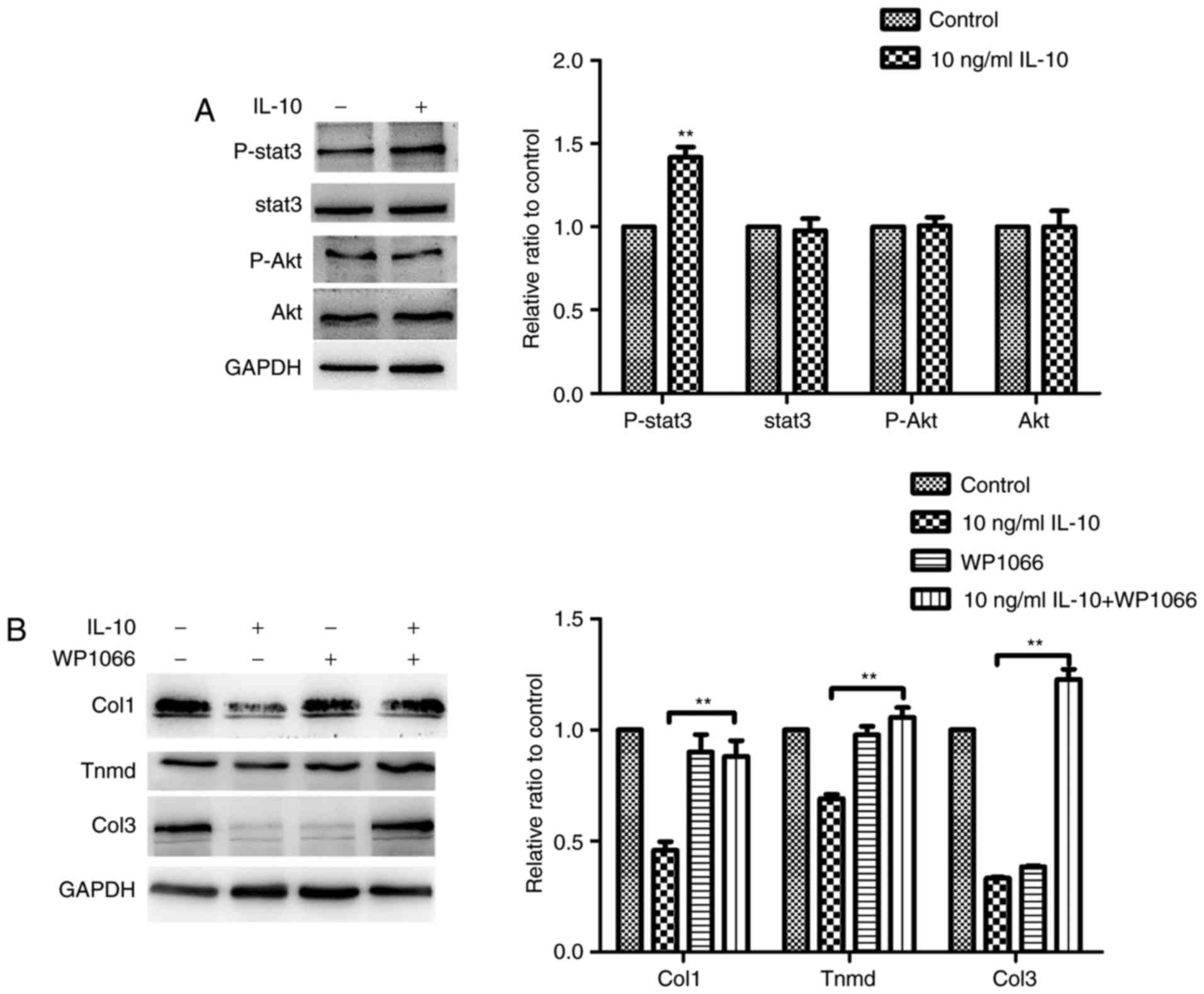 | Figure 5.IL-10 activates Stat3 but has no
effect on Akt expression in TDSCs. (A) Rat TDSCs were treated with
the indicated concentrations of IL-10 and were subjected to western
blot analysis to detect p-Stat3, Stat3, p-Akt and Akt protein
expression. (B) TDSCs were treated with the indicated
concentrations of IL-10 with or without the Stat3 inhibitor WP1066
and subjected to western blot analysis for Col1, Tnmd and Col3
protein expression. Data are expressed as the mean ± standard error
of the mean. *P<0.05, **P<0.01 vs. 10 ng/ml IL-10. IL-10,
interleukin-10; TDSCs, tendon-derived stem cells; p-,
phosphorylated; Stat3, Signal transducer and activator of
transcription 3; Akt, protein kinase B; Tnmd, tenomodulin; Col3,
collagen type 3. |
Discussion
The tendon is a unique connective tissue with few
cells, blood vessels or nerves. Its healing follows a typical wound
healing course, which is divided into three overlapping phases: The
early inflammatory, proliferative and remodeling phases (24). A commonly injured tendon never
regains complete recovery and may result in heterotopic
ossification (25,26). The inferior scar tissue formation
greatly increases the risk of reinjury (27). Recruitment of a sufficient number
of TDSCs is the premise of effective healing, which occurs at the
late inflammatory phase and proliferative phase (24). Furthermore, correct tenogenic
differentiation of TDSCs is key to exerting beneficial effects,
including collagen synthesis, regulation of the tendon
microenvironment and regaining normal tendon tissue (17,28).
It has been reported that IL-10 is upregulated at the
post-inflammatory phase in early human tendon repair (9). IL-10 inhibits the release of
pro-inflammatory mediators (29),
including IL-1β, which was demonstrated to irreversibly inhibit
tenogenic differentiation of injured tendon-derived progenitor
cells in our previous study (30).
Moroguchi et al (31)
demonstrated that IL-10 suppresses the proliferation and expression
of type I collagen in human skin fibroblasts. In addition, IL-10
exerts an anti-apoptotic effect, as well as homing and
differentiation effects in cells (12,13,32).
However, the exact impact of increased IL-10 on injured tendons has
not been fully elucidated.
The results of the present study revealed that the
proliferative capacity of TDSCs was significantly increased when
the cells were treated with IL-10. This indicated that the
upregulation of IL-10 at the later inflammatory phase may promote
tendon healing by stimulating the proliferation of TDSCs. Notably,
it was demonstrated that IL-10 induced cell cycle activation and
transition into the G2/M phase from the G1 phase. Additionally,
IL-10 significantly promoted cell migration when compared with the
control cells, which reflected the repair capacity of TDSCs. In the
regenerative phase, healing cells migrate into the injury/repair
site, actively proliferate and deposit abundant extracellular
matrix components in the tissue (33). Therefore, the proliferation and
migration of TDSCs serves a pivotal role in tendon healing. The
findings suggest that IL-10, which promotes the proliferation and
migration of TDSCs, has a positive impact on tendon healing.
Scx and Tnmd represent tenogenic differentiation
markers (2). Under
self-differentiated culture conditions, the present study revealed
that IL-10 reduced the expression levels of Scx and Tnmd, as well
as Egr-1, a transcriptional factor that functions in tendon
development (34), indicating that
IL-10 may have suppressed TDSC tenogenic differentiation. In
addition, IL-10 reduced the expression levels of the main
tendon-associated collagens, Col1 and Col3, which was consistent
with the results obtained by Abbah et al (35) in human tenocytes. Similarly, gene
expression levels of Fmod and Lum, two leucine-rich repeat
proteins influencing collagen fibrillogenesis (36), were also inhibited in TDSCs by
IL-10. Notably, as Fmod is a critical component of the TDSC niche,
reduction of Fmod affects TSDC differentiation (17). It was also revealed that
overexpression of Fmod enhanced tendon healing in vivo and
in vitro (37).
Furthermore, Fmod-deficient mice developed abnormal tendon and
ectopic ossification (38). It has
been demonstrated Lum has a combined effect together with Fmod at
different developmental stages (36). Taken together, these effects of
IL-10 may have altered the microenvironment of the TDSCs niche to
determine cell fate, consequentially impairing the recovery of
biomechanical properties of regenerating tendons. Furthermore,
IL-10 is predominantly regulated by JAK and Stat3 transcription
factors, and considering that Stat3 is situated downstream of JAK.
Stat3 is phosphorylated by JAK (32,39,40).
The results of the present study determined that IL-10 inhibited
the gene expression of tenogenic differentiation markers in TDSCs
by activating the JAK/Stat3 pathway, which was similar to results
obtained in previous report in ventral mesencephalic neurons
(41).
Upregulated expression of Mkx, a homeobox gene
involved in tendon development, was detected in the present study,
which was similar to our previous results regarding IL-1β (30). Ito et al (42) reported that Mkx mutant mouse
tendons were hypoplastic throughout the body and collagen fibril
diameters were smaller, indicating that Mkx has a critical role in
tendon development by regulating type I collagen production
(42). Although the role of
Mkx in tendon repair has not been fully elucidated,
upregulation of Mkx by IL-10 may be involved in collagen fibril
formation during tendon healing. However, the specific significance
of Mkx upregulation by IL-10 and other cytokines requires further
investigation.
The present study demonstrated that IL-10 exerted
dual effects on TDSCs in vitro. On the one hand, IL-10
enhanced the proliferation and migration of TDSCs. Conversely,
IL-10 also inhibited tenogenic differentiation of TDSCs. However,
in a murine patellar tendon model, Ricchetti et al (11) revealed that overexpression of IL-10
markedly increased its maximum stress, indicating that complex
mechanisms regulate the in vivo effect of IL-10. Although it
remains unclear whether the same pathway regulates of the promotion
of the proliferation of TDSCs and the inhibition of tenogenic
differentiation, it was concluded that timely control of the
negative effect of IL-10 and interventional promotion of its
positive effect may benefit the stimulation of tendon regeneration.
Therefore, effective regulation of IL-10 expression may represent a
therapeutic strategy for the clinical treatment of tendon
diseases.
In summary, the present study revealed that IL-10
exerted dual effects on TDSCs in vitro via strongly
enhancing cell proliferation and migration, as well as inhibiting
TDSC tenogenic differentiation through activation of the JAK/Stat3
pathway.
Acknowledgements
The authors thank Professor Liang Ping for his
revision of this manuscript.
Funding
This study was supported by the National Natural
Science Foundation of China (grant no. 81601900) and the Science
and Technology Project of Guangdong Province (grant no.
2016A020214010).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding authors on reasonable
request.
Authors' contributions
The experiments were designed by GD and KZ.
Experiments were performed by GD, KL, SC, PC and HZ. Data were
analyzed by GD, KZ and BY. The paper was written by GD, KZ and BY.
All authors read and approved the final manuscript, were involved
in revising the manuscript critically for important intellectual
content, study conception and design, as well as data
interpretation and analysis.
Ethical approval and consent to
participate
All aspects of the research were approved by the
Institutional Animal Care and Use Committee of Nanfang Hospital,
Southern Medical University (Guangzhou, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Nourissat G, Berenbaum F and Duprez D:
Tendon injury: From biology to tendon repair. Nat Rev Rheumatol.
11:223–233. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gaut L and Duprez D: Tendon development
and diseases. Wiley Interdiscip Rev Dev Biol. 5:5–23. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kaux JF, Forthomme B, Goff CL, Crielaard
JM and Croisier JL: Current opinions on tendinopathy. J Sports Sci
Med. 10:238–253. 2011.PubMed/NCBI
|
|
4
|
Leadbetter WB: Cell-matrix response in
tendon injury. Clin Sports Med. 11:533–578. 1992.PubMed/NCBI
|
|
5
|
Xu Y and Murrell GA: The basic science of
tendinopathy. Clin Orthop Relat Res. 466:1528–1538. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Battery L and Maffulli N: Inflammation in
overuse tendon injuries. Sports Med Arthrosc Rev. 19:213–217. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Millar NL, Dean BJ and Dakin SG:
Inflammation and the continuum model: Time to acknowledge the
molecular era of tendinopathy. Br J Sports Med. 50:14862016.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Abate M, Silbernagel KG, Siljeholm C, Di
Iorio A, De Amicis D, Salini V, Werner S and Paganelli R:
Pathogenesis of tendinopathies: Inflammation or degeneration?
Arthritis Res Ther. 11:2352009. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ackermann PW, Domeij-Arverud E, Leclerc P,
Amoudrouz P and Nader GA: Anti-inflammatory cytokine profile in
early human tendon repair. Knee Surg Sports Traumatol Arthrosc.
21:1801–1806. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tarafder S, Chen E, Jun Y, Kao K, Sim KH,
Back J, Lee FY and Lee CH: Tendon stem/progenitor cells regulate
inflammation in tendon healing via JNK and STAT3 signaling. FASEB
J. 31:3991–3998. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ricchetti ET, Reddy SC, Ansorge HL, Zgonis
MH, Van Kleunen JP, Liechty KW, Soslowsky LJ and Beredjiklian PK:
Effect of interleukin-10 overexpression on the properties of
healing tendon in a murine patellar tendon model. J Hand Surg Am.
33:1843–1852. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhou J, Ling J, Song J, Wang Y, Feng B and
Ping F: Interleukin 10 protects primary melanocyte by activation of
Stat-3 and PI3K/Akt/NF-κB signaling pathways. Cytokine. 83:275–281.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Verma SK, Garikipati VNS, Krishnamurthy P,
Schumacher SM, Grisanti LA, Cimini M, Cheng Z, Khan M, Yue Y,
Benedict C, et al: Interleukin-10 inhibits bone marrow fibroblast
progenitor cell-mediated cardiac fibrosis in pressure-overloaded
myocardium. Circulation. 136:940–953. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Reitamo S, Remitz A, Tamai K and Uitto J:
Interleukin-10 modulates type I collagen and matrix metalloprotease
gene expression in cultured human skin fibroblasts. J Clin Invest.
94:2489–2492. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang J and Wang JH: Characterization of
differential properties of rabbit tendon stem cells and tenocytes.
BMC Musculoskel Dis. 11:102010. View Article : Google Scholar
|
|
16
|
Rui YF, Lui PP, Li G, Fu SC, Lee YW and
Chan KM: Isolation and characterization of multipotent rat
tendon-derived stem cells. Tissue Eng Part A. 16:1549–1558. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bi Y, Ehirchiou D, Kilts TM, Inkson CA,
Embree MC, Sonoyama W, Li L, Leet AI, Seo BM, Zhang L, et al:
Identification of tendon stem/progenitor cells and the role of the
extracellular matrix in their niche. Nat Med. 13:1219–1227. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang K, Zhang S, Li Q, Yang J, Dong W,
Wang S, Cheng Y, Al-Qwbani M, Wang Q and Yu B: Effects of celecoxib
on proliferation and tenocytic differentiation of tendon-derived
stem cells. Biochem Biophys Res Commun. 450:762–766. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Asai S, Otsuru S, Candela ME, Cantley L,
Uchibe K, Hofmann TJ, Zhang K, Wapner KL, Soslowsky LJ, Horwitz EM
and Enomoto-Iwamoto M: Tendon progenitor cells in injured tendons
have strong chondrogenic potential: The CD105-negative
subpopulation induces chondrogenic degeneration. Stem Cells.
32:3266–3277. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wu YH, Liu W, Zhang L, Liu XY, Wang Y, Xue
B, Liu B, Duan R, Zhang B and Ji Y: Effects of microRNA-24
targeting C-myc on apoptosis, proliferation, and cytokine
expressions in chondrocytes of rats with osteoarthritis via MAPK
signaling pathway. J Cell Biochem. 16–Nov;2017.(Epub ahead of
print).
|
|
21
|
Zhang T, Ji D, Wang P, Liang D, Jin L, Shi
H, Liu X, Meng Q, Yu R and Gao S: The atypical protein kinase RIOK3
contributes to glioma cell proliferation/survival,
migration/invasion and the AKT/mTOR signaling pathway. Cancer Lett.
415:151–163. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liang CC, Park AY and Guan JL: In vitro
scratch assay: A convenient and inexpensive method for analysis of
cell migration in vitro. Nat Protoc. 2:329–333. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Thomopoulos S, Parks WC, Rifkin DB and
Derwin KA: Mechanisms of tendon injury and repair. J Orthop Res.
33:832–839. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Oliva F, Via AG and Maffulli N:
Physiopathology of intratendinous calcific deposition. BMC Med.
10:952012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
O'Brien EJ, Frank CB, Shrive NG,
Hallgrímsson B and Hart DA: Heterotopic mineralization
(ossification or calcification) in tendinopathy or following
surgical tendon trauma. Int J Exp Pathol. 93:319–331. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Butler DL, Juncosa N and Dressler MR:
Functional efficacy of tendon repair processes. Annu Rev Biomed
Eng. 6:303–329. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Guo J, Chan KM, Zhang JF and Li G:
Tendon-derived stem cells undergo spontaneous tenogenic
differentiation. Exp Cell Res. 341:1–7. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fiorentino DF, Zlotnik A, Mosmann TR,
Howard M and O'Garra A: IL-10 inhibits cytokine production by
activated macrophages. J Immunol. 147:3815–3822. 1991.PubMed/NCBI
|
|
30
|
Zhang K, Asai S, Yu B and Enomoto-Iwamoto
M: IL-1β irreversibly inhibits tenogenic differentiation and alters
metabolism in injured tendon-derived progenitor cells in vitro.
Biochem Biophys Res Commun. 463:667–672. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Moroguchi A, Ishimura K, Okano K,
Wakabayashi H, Maeba T and Maeta H: Interleukin-10 suppresses
proliferation and remodeling of extracellular matrix of cultured
human skin fibroblasts. Eur Surg Res. 36:39–44. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sabat R, Grütz G, Warszawska K, Kirsch S,
Witte E, Wolk K and Geginat J: Biology of interleukin-10. Cytokine
Growth Factor Rev. 21:331–344. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tsai WC, Yu TY, Lin LP, Cheng ML, Chen CL
and Pang JH: Prevention of simvastatin-induced inhibition of tendon
cell proliferation and cell cycle progression by geranylgeranyl
pyrophosphate. Toxicol Sci. 149:326–334. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Guerquin MJ, Charvet B, Nourissat G, Havis
E, Ronsin O, Bonnin M, Ruggiu M, Olivera-Martinez I, Robert N, Lu
Y, et al: Transcription factor EGR1 directs tendon differentiation
and promotes tendon repair. J Clin Invest. 123:3564–3576. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Abbah SA, Thomas D, Browne S, O'Brien T,
Pandit A and Zeugolis DI: Co-transfection of decorin and
interleukin-10 modulates pro-fibrotic extracellular matrix gene
expression in human tenocyte culture. Sci Rep. 6:209222016.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ezura Y, Chakravarti S, Oldberg A,
Chervoneva I and Birk DE: Differential expression of lumican and
fibromodulin regulate collagen fibrillogenesis in developing mouse
tendons. J Cell Biol. 151:779–788. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Delalande A, Gosselin M, Suwalski A,
Guilmain W, Leduc C, Berchel M, Jaffrès P, Baril P, Midoux P and
Pichon C: Enhanced achilles tendon healing by fibromodulin gene
transfer. Nanomedicine. 11:1735–1744. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ameye L, Aria D, Jepsen K, Oldberg A, Xu T
and Young MF: Abnormal collagen fibrils in tendons of
biglycan/fibromodulin-deficient mice lead to gait impairment,
ectopic ossification, and osteoarthritis. FASEB J. 16:673–680.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ahmed ST and Ivashkiv LB: Inhibition of
IL-6 and IL-10 signaling and Stat activation by inflammatory and
stress pathways. J Immunol. 165:5227–5237. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Tanuma N, Shima H, Nakamura K and Kikuchi
K: Protein tyrosine phosphatase epsilonC selectively inhibits
interleukin-6- and interleukin-10-induced JAK-STAT signaling.
Blood. 98:3030–3034. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhu Y, Liu Z, Peng YP and Qiu YH:
Interleukin-10 inhibits neuroinflammation-mediated apoptosis of
ventral mesencephalic neurons via JAK-STAT3 pathway. Int
Immunopharmacol. 50:353–360. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Ito Y, Toriuchi N, Yoshitaka T, Ueno-Kudoh
H, Sato T, Yokoyama S, Nishida K, Akimoto T, Takahashi M, Miyaki S
and Asahara H: The Mohawk homeobox gene is a critical regulator of
tendon differentiation. Proc Nat Acad Sci. 107:10538–10542. 2010.
View Article : Google Scholar : PubMed/NCBI
|