Introduction
Tendon injuries are frequent and contribute to a
substantial number of cases of morbidity (≤64% in patients with
rheumatoid arthritis) in sports and in the workplace (1). When the Achilles tendon experiences
an injury, the damaged tissue may cause a severe inflammatory
reaction, resulting in a ‘secondary injury’ to the Achilles tendon
(2,3). Furthermore, the repair of flexor
tendon injuries is complex due to the adhesion forming between the
tendons and the surrounding soft tissue post-surgery (4). Another study also provides evidence
that adhesion formation has often resulted in limited functional
recovery and decreased the ability of the tendon to move smoothly
(5). Tendon adhesion is regarded
as one of the main problems involved with tendon repair, for which
no ideal treatment is currently available (6). Similar to the majority of other
tissue repair mechanisms following injury, tendon repair involves a
series of molecular and cellular events affected by the location of
the injury site, age, sex, nutrition and genetics (7). Thermal pretreatment (TP) serves as a
notable factor in the reduction of the inflammatory response and
tendon healing by inducing molecular chaperone expression (8).
Heat shock proteins (HSPs), including molecular
chaperones, are highly preserved proteins present in eukaryotic and
prokaryotic cells and serve a crucial function in various cellular
processes, including protein folding, regulation of signaling
pathway, degradation of misfolded proteins and the modulation of
immune responses (9). HSPs are
capable of maintaining cellular homeostasis whenever the
microenvironment changes, modulating cell differentiation and
proliferation physiologically and pathologically (10). HSP70, a widespread family of
molecular chaperones, modulate the protein quality control in
addition to homeostasis, serving as an effective drug target for
the treatment of neurodegenerative and hyperproliferative disorders
(11). Wounds are a type of stress
condition that may induce cellular and biochemical responses given
that it may promote a significant increase in HSP70 expression in
coelomocytes (12). HSPs exhibit
properties that promote cell survival in response to all forms of
cellular stress (13). Mushtaq
et al (14) revealed that
HSP70 is necessary for the regulation of epithelial growth,
regeneration or adhesion, promoting corneal epithelial wound
healing. Therefore, the present study hypothesized that the
interaction between HSP70 and TP may affect the healing process in
tendon injury. The present study intends to investigate the
function of TP along with its association with the upregulation of
HSP70 in tendon healing and adhesion in rat models simulating
tendon injury.
Materials and methods
Ethical statement
The animal experiments were conducted in accordance
with the Guide for the Care and Use of Laboratory Animals as
promulgated by the National Institutes of Health (15). Ethical approval was obtained from
the ethics committee of The Affiliated Huai'an No.1 People's
Hospital of Nanjing Medical University (Huai'an, China).
Experimental animals and establishment
of tendon injury rat model
A total of 60 male Sprague-Dawley (SD) rats (aged 10
weeks and weighing between 350–450 g) were obtained from the Hubei
Research Center of Laboratory Animals (Hubei, China) and randomly
assigned into the following groups: Normal (healthy rats) group,
model (rats with tendon injury) group, TP (tendon injury rats
treated with TP) group, negative control (NC)-small interfering RNA
(siRNA) group (tendon injury rats with an injection of 20 µl
suspension of NC-siRNA-transfected cells at the site of the
injury), HSP70-siRNA group (tendon injury rats treated with a 20 µl
suspension of HSP70-siRNA-transfected cells at the site of the
injury) and a combination group (tendon injury rats treated with TP
and a 20 µl suspension of HSP70-siRNA-transfected cells at the site
of the injury) groups (n=10). At 4 and 8 weeks following surgery, 5
rats were assigned to each group. The rats were provided with
adaptive feeding at a room temperature of 23–25°C with a relative
humidity between 45–60% and a light-dark cycle of 12–12 h for 1
week. All rats were allowed ad libitum access to food and
water.
The rats were placed under general anesthesia via an
intraperitoneal injection of a 1% pentobarbital sodium (45 mg/kg)
solution and were maintained stationary in a supine position. To
obtain a precise incision on the model rats, their legs were shaved
and sterilized using iodophor. A 2-cm longitudinal plantar incision
was created in the middle of the left paw of the rat. The skin and
subcutaneous tissue were sequentially cut to expose the flexor
tendon of the deep flexor tendon, and the proximal end of the
tendon bifurcation was transversely sliced at ~3 mm. The incision
was sutured with a 5-0 tendon suture using the Kessler method
(16) and wrapped with sterile
gauze. The rats were then administered a subcutaneous injection of
sodium chloride solution (50 ml/kg) on the nape located on their
back immediately following the surgery and returned to their cages.
They were again provided water and food 4 h subsequent to returning
to their cages. The TP group was then placed in an incubator at a
temperature of 45°C and remained in the incubator for 15 min until
the rectal temperature of rats (measured with a rectal thermometer)
reached ~41.5±0.5°C. The normal group was subsequently maintained
in an incubator at 37°C without any form of TP. All rats were
allowed to recover at room temperature for 24 h, and none of the
rats exhibited symptoms of lethargy and mortality.
Construction of HSP70-siRNA expression
vector
The pSUPER-puro expression system (an original short
hairpin RNA design) was used to construct the HSP70-siRNA
expression vector. Two specific target sequences of HSP70 (GenBank
no. NM_005345) were constructed: HSP70-siRNA sense,
5′-GGACGAGUUUGAGCACAAGTT-3′ and antisense,
5′-CUUGUGCUCAAACUCGUCCTT-3′; NC-siRNA sense,
5′-UUCUCCGAACGUGUCACGUTT-3′ and antisense,
5′-ACGUGACACGUUCGGAGAATT-3′. Each of the two nucleotides in the
target sequence contained 9 hairpin structure sequences. The
hairpin structure was targeted at the Bgl II and Sal
I sites of pSUPER-puro plasmid, thereby constructing a novel
recombinant plasmid. H1 RNA polymerase promoter and puromycin were
used to select a clone with stable expression.
Culture and transfection of tendon
cells
Aponeurosis was homogenized once it was detached
from the tendon, digested with mixed collagenase (100 U/µl; cat.
no. 17101015; Gibco; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) and placed in Dulbecco's modified Eagle's medium (DMEM)
containing 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.) at 37°C in a humidified atmosphere of 5%
CO2. for primary culture, and the medium was changed
once every 4–5 days. When the medium covered 90% of the bottom of
the medium, the cells were treated with trypsin (1:3; Gibco; Thermo
Fisher Scientific, Inc.) to subculture. The cells were passaged
until the 3rd generation, which were used for subsequent
experiments. Tendon cells involved during the logarithmic growth
phase were then seeded into a 12-well plate at a density of
5×104 cells/well and finally incubated for a 20 h
interval at 37°C. Once the medium was replaced by DMEM without the
addition of FBS, the cells were transfected with the constructed
plasmids (4 µg) using Lipofectamine® 2000 reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) for 48 h at 37°C
according to the manufacturer's protocol. Following the
aforementioned procedure, the tendon cells used in the in
vitro culture were examined. Next, the fluorescent intensity
was observed and photographed under a fluorescence inverted
microscope (magnification, ×200; CX41-12C02, Olympus Optical Co.,
Ltd., Tokyo, Japan) following 48 h transfection. The transfection
efficiency was calculated according to the following formula:
Transfection efficiency (%) = number of cells with green
fluorescent protein (GFP)/total cell number ×100% (17,18).
A total of 5 high-power fields were randomly selected, and the mean
values were determined.
Observation of tendon adhesion
Tendon healing at the anastomotic site, grading
(6), areas of adhesion in
peripheral tissues, tendon gliding and smoothness of the
aforementioned anastomotic surface were all observed. At week 4
following the surgery, all surgical incisions healed sufficiently
without any inflammatory reactions or infection. No rats succumbed
to mortality as a result of the surgery. The rats maintained for 4
weeks were then sacrificed; the remaining rats were fed regularly
and sacrificed on the 8th week following surgery. Tendon adhesion
and gliding in rats were evaluated using a single-blind method
(Table I) (6).
 | Table I.Scoring standard for tendon
adhesion. |
Table I.
Scoring standard for tendon
adhesion.
| Point | Severity of tendon
adhesion |
|---|
| 0 | Without adhesion:
The tendons glide freely without injury |
| 1–2 | Mild adhesion: Fine
filaments adhering to peripheral tissues |
| 3–4 | Moderate adhesion:
Scattered filaments adhering to peripheral tissues |
| 5–6 | Severe adhesion: A
large number of filaments adhering to peripheral tissues, and the
tendon cannot be separated from peripheral tissues and cannot
glide |
Biomechanical test
Following the single-blind method, the rats were
injected intraperitoneally with a pentobarbital sodium (150 mg/kg)
solution. Following euthanasia, the second toe was disconnected
from the carpometacarpal joint, and the deep toe flexor tendon of
the second toe was obtained (cut created along ~3 cm of the
carpometacarpal joint). The tendon was then placed in position in a
BA-100T mechanical-standard rubber tension test machine (Zhengbang
Testing Equipment Co., Ltd., Dongguan, Guangdong, China; http://www.ybzhan.cn/tp29536), ensuring that the
tendon was clamped tightly, and that the tendon healing site was
located in the middle area between the clamps. The test began with
the loading of 1-N force to maintain the proper tendon tension. The
initial length was measured. The tendon was stretched to 0.5 mm at
a speed of 0.5 mm/s, and the pretreatment was conducted a total of
three times. Finally, the tendon was loaded to break at a speed of
1 cm/min. The maximum tensile force of tendon and the sliding
distance of the tendon were recorded.
Paraffin sectioning
Following the biomechanical test, the tendon tissue
was subsequently removed and treated with 4% paraformaldehyde at
25°C for 4 h. Subsequent to dehydrating with graded ethanol
(75,80,95, and 100%), the tissue was cleared using xylene,
wax-immersed and then embedded. The paraffin block was placed on a
microtome and coarsely repaired to expose the largest section of
the tissue block, which was then finely cut into thin slices ~5 µm
thick. The sections were then placed in a hot water bath (Slidetec
WATER; Labsun GmbH, Karlsruhe, Germany). The creased slices were
leveled and then placed on a glass slide. The paraffin-embedded
specimens were then dried on a 45°C hotplate (Slidetec HEAT; Labsun
GmbH) for a minimum of 5 min.
Hematoxylin and eosin (H&E)
staining
Subsequent to removal from the copying machine, the
slices were oven-dried in a 37°C oven for 30 min and dewaxed using
graded ethanol (100 and 80% respectively) until fully hydrated. The
slices were then stained using Harris hematoxylin (Shanghai Yuanye
Biotechnology Co., Ltd., Shanghai, China) for 10 min at 25°C. Then,
the slices were washed under tap water for 1 min, differentiated in
1% hydrochloric acid-ethanol, and rinsed under running water to
restore the blue color for 15 min. The slices were then stained
with a 1% ethanol solution of eosin (Shanghai Yuanye Biotechnology
Co., Ltd.) for 3 min at 25°C, differentiated in 90% ethanol for 30
sec and washed with a 95% ethanol solution for 1 min in addition to
with dimethyl carbonate (Shanghai Yuanye Biotechnology, Co., Ltd.)
for 1 min. The slices were then washed using dimethylbenzene
(Sinopharm Chemical Reagent Co., Ltd., Shanghai, China) three times
(2 min each time) and sealed using a neutral plasma (cat. no.
GT21316; Beijing Huayueyang Biotechnology Co., Ltd., Beijing,
China; http://huayueyang.biogo.net/). The
morphological structure of the tendons was observed under a light
microscope (magnification, ×200; CX41-12C02, Olympus Optical Co.,
Ltd.).
Immunohistochemistry
Subsequent to oven-drying in a 37°C oven for 30 min,
the slices were dewaxed using dimethylbenzene (Sinopharm Chemical
Reagent Co., Ltd.) followed by a dehydration process in graded
ethanol (100,95,80, and 75% respectively). Following rehydration,
endogenous peroxidase activity was inhibited by incubating with 3%
hydrogen peroxide for 1 h at 37°C. Then, the slices were washed
with phosphate buffer saline (PBS; Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) three times (2 min each time) followed by 10
min of incubation in a sodium citrate buffer (pH 6.0, 0.01 mol/l)
at 96°C for antigen retrieval. Subsequent to washing with PBS, the
slices were then treated with a rabbit anti-rat HSP70 antibody
[1:100; HSP70(W27)(SC-24); Hebei Bio-High Technology Development
Co., Ltd., Shijiazhuang, Hebei, China] and placed in a 4°C
incubator overnight. Subsequent to washing with PBS thrice (5 min
each time), a goat anti-rabbit immunoglobulin G secondary antibody
(1:2,000; D111018; Shanghai Sangon Biological Engineering
Technology & Services Co., Ltd., Shanghai, China) conjugated to
horseradish peroxidase was also added followed by an incubation
period at room temperature for 15 min. The slices were then
developed using diaminobenzidine (100 µl; Wuhan Boster Biological
Technology, Ltd., Wuhan, China) and further incubated at room
temperature for an additional 5–10 min. Furthermore, the slides
were washed with distilled water to terminate the color and further
stained using hematoxylin for 5–10 min at 25°C. Then, the slides
were sealed using neutral plasma. Positive HSP70 expression was
present in small brown yellow granules observed in the nucleus or
cytoplasm. Five high-power fields were randomly selected for each
slice with 1,000 cells per field of view under a light microscope
(magnification, ×200; CX41-12C02, Olympus Optical Co., Ltd.). The
number of positive cells in the field of view were calculated, and
the mean positive expression rate was determined as follows: Mean
positive expression rate=number of positive cells/total cells
(%).
Histomorphological observation
Tendon tissues of the rats were obtained from the
hind limbs of the model rats the 4 and 8th week following surgery,
placed in 10% buffered formalin for 24 h and transferred into an
80% ethanol gradient. Paraffin-embedded tissues were then cut into
5-µm thick slices and stained using H&E staining, as described
above. Tendon development, filament formation, shape, the number of
fibroblasts and collagen fiber arrangements were observed under an
electron microscope (magnification, ×200; CX41-12C02, Olympus
Optical Co., Ltd.). The inflammatory reaction of the specimens from
rats from the 4 and 8th week post-surgery was evaluated at this
juncture of the experiment. The grading criteria of the
inflammatory reaction were established as follows: Grade 1, without
or with few lymphocytes; grade 2, a small amount of lymphocytes;
grade 3, infiltration of a small amount of neutrophils and
lymphocytes in addition to a giant cell reaction; and grade 4,
macrophage and inflammatory reactions with the marked infiltration
of neutrophils.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Newly formed tendons (0.1 g) were obtained from the
rats at the 4 and 8th week following surgery in each group. Total
RNA was then isolated from the tendon tissues of all groups using
the TRIzol reagent kit (Gibco; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. RNA purity and
concentration were assessed using a NanoDrop 2000c
Spectrophotometer (Thermo Fisher Scientific, Inc.). Then, HSP70
mRNA was reversely transcribed using the A3500 RT system (Promega
Corporation, Madison, WI, USA) at 42°C for 1 h and at 95°C for 5
min according to the manufacturer's protocol. RT-qPCR was conducted
with synthesized cDNA using a Fast SYBR Green PCR kit (Applied
Biosystems; Thermo Fisher Scientific, Inc.). Based on the sequences
provided by GenBank (19),
specific RT stem-loop primers and primers for PCR amplification
(Table II) were designed using
Primer Premier 5.0 (Premier Biosoft International, Palo Alto, CA,
USA). RT-qPCR analyses were conducted on an ABI 7500 PCR instrument
(Applied Biosystems; Thermo Fisher Scientific, Inc.). For the PCR
procedure, predenaturation was performed at a temperature of 94°C
for 5 min followed by 40 cycles of denaturation at a temperature of
94°C for 30 sec, annealing at 58°C for 30 sec and extension at 72°C
for 1 min. The mRNA levels of HSP70 and GAPDH genes in the
experimental and control groups were measured and calculated
utilizing the 2−ΔΔCq method (20). The quantification cycle (Cq) at
which fluorescence intensity is accumulated at a certain quantity
was obtained. The 2−ΔΔCq method was used to determine
the ratio of expression of the target genes between the
experimental and control groups. The following formula was
employed: ΔΔCq=ΔCq experimental
group-ΔCq control group, and
ΔCq=Cq HSP70-Cq GAPDH.
| Table II.Primer sequences for reverse
transcription-quantitative polymerase chain reaction. |
Table II.
Primer sequences for reverse
transcription-quantitative polymerase chain reaction.
| Item | Primer
sequence |
|---|
| Heat shock | Forward,
5′-AAGGTGGAGATCATCGCCAA-3′ |
| protein 70 | Reverse,
5′-GCGATCTCCTTCTTCATCTTGGT-3′ |
| GAPDH | Forward,
5′-AATTCAACGGCACAGTCAAGGC-3′ |
|
| Reverse,
5′-GGATGCAGGGATGATGTTCTGG-3′ |
Western blot analysis
Newly formed tendons (0.1 g) were obtained from rats
at the 4th and 8th week following surgery in each group. Total
protein was then harvested from the tendon tissues of all groups
using a TRIzol reagent kit (Gibco; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. The protein concentration
of the tendon cells was then measured using a bicinchoninic acid
protein assay kit (Beyotime Institute of Biotechnology, Beijing,
China). The extracted protein was then mixed with loading buffer,
boiled for 10 min at 100°C and loaded at a density of 400 µg per
well. The proteins were then separated using 10% sodium dodecyl
sulfate-polyacrylamide gel (Wuhan Boster Biological Technology,
Ltd.) electrophoresis. The voltage for transfer ranged from 80–120
V. Following the aforementioned transfer, the proteins were
transferred onto a polyvinylidene fluoride membrane with wet
spinning at a voltage of 100 V for 45–70 min and subsequently
blocked using 10% dried skimmed milk at room temperature for 1 h.
The proteins were then incubated with the following primary
antibodies at 4°C overnight: Rabbit anti-rat HSP70 monoclonal
antibody (1:300; cat no. sc-221731), transforming growth factor-β
(TGF-β; 1:200; cat. no. sc-203295), insulin-like growth factor I
(IGF-1; 1:200; cat. no. sc-1422) and β-actin (1:100; cat. no.
sc-47778). All antibody used above were purchased from Santa Cruz
Biotechnology, Inc. (Dallas, TX, USA). The proteins were washed
using tris-buffered saline with 0.5% Tween 20 (TBST) three times (3
min per wash). Then, the secondary antibody, horseradish
peroxidase-conjugated goat anti-mouse antibody (1:1,000, ab97057,
Abcam, Cambridge, MA, USA) were added and incubated at room
temperature for 1 h. Subsequent to washing with TBST for an
additional three times (each for 5 min), the samples were finally
developed using an enhanced chemiluminescence method (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). Gray values of the protein
bands were analyzed by using SPSS version 21.0 (IBM Corp. Armonk,
NY, USA). The ratio of the gray values targeting proteins were
determined and β-actin was used as an internal reference for
targeted protein levels.
Statistical analysis
SPSS version 21.0 (IBM Corp., Armonk, NY, USA) was
used for statistical analysis. Measurement data were expressed as
the mean ± standard deviation. Comparisons among groups were
analyzed utilizing a one-way analysis of variance, followed by
Tukey's post hoc test. Repeated measures analysis of variance was
used for comparisons among multiple groups at multiple time points.
P<0.05 was considered to indicate a statistically significant
difference.
Results
TP reduces tendon adhesion and
HSP70-siRNA increases tendon adhesion
Tendon adhesion in each group was observed at weeks
4 and 8 subsequent to the operation. A single-blind method was used
for the evaluation of the tendon adhesion in each group. As
presented in Fig. 1, at the 4th
week following surgery, the adhesion grades of tendon adhesion
among the model, TP, HSP70-siRNA, NC-siRNA and combined groups
exhibited significant differences compared with the normal group
(P<0.05). Compared with the model group, no significant
difference was observed between the NC-siRNA group and the combined
group (P>0.05). Compared with the model group, the TP group
presented a significantly lower adhesion score (P<0.05), whereas
the HSP70-siRNA group exhibited a significantly increased adhesion
score (P<0.05). These scores indicated that TP reduces tendon
adhesion, whereas HSP70 gene silencing increases adhesion.
Successful transfection of HSP70-siRNA
in vitro
Five days following transfection, a fluorescence
inverted microscope was used to observe GFP in transfected tendon
cells. The transfection was confirmed through GFP expression
tracked by fluorescence photography. The transfection confirmed an
efficiency between 50 and 70% (Fig.
2A-B), indicating that the transfection of HSP70-siRNA was
successful. RT-qPCR was used to detect HSP70 expression upon
transfection with different siRNAs (Fig. 2C). Compared with the NC-siRNA
group, HSP70 mRNA expression levels in the HSP70 siRNA-1, HSP70
siRNA-2 and HSP70 siRNA-3 groups were significantly decreased
(P<0.05), and the greatest decrease in HSP70 mRNA expression was
observed in the HSP70 siRNA-2 group. Therefore, HSP70 siRNA-2 was
selected for subsequent experiments.
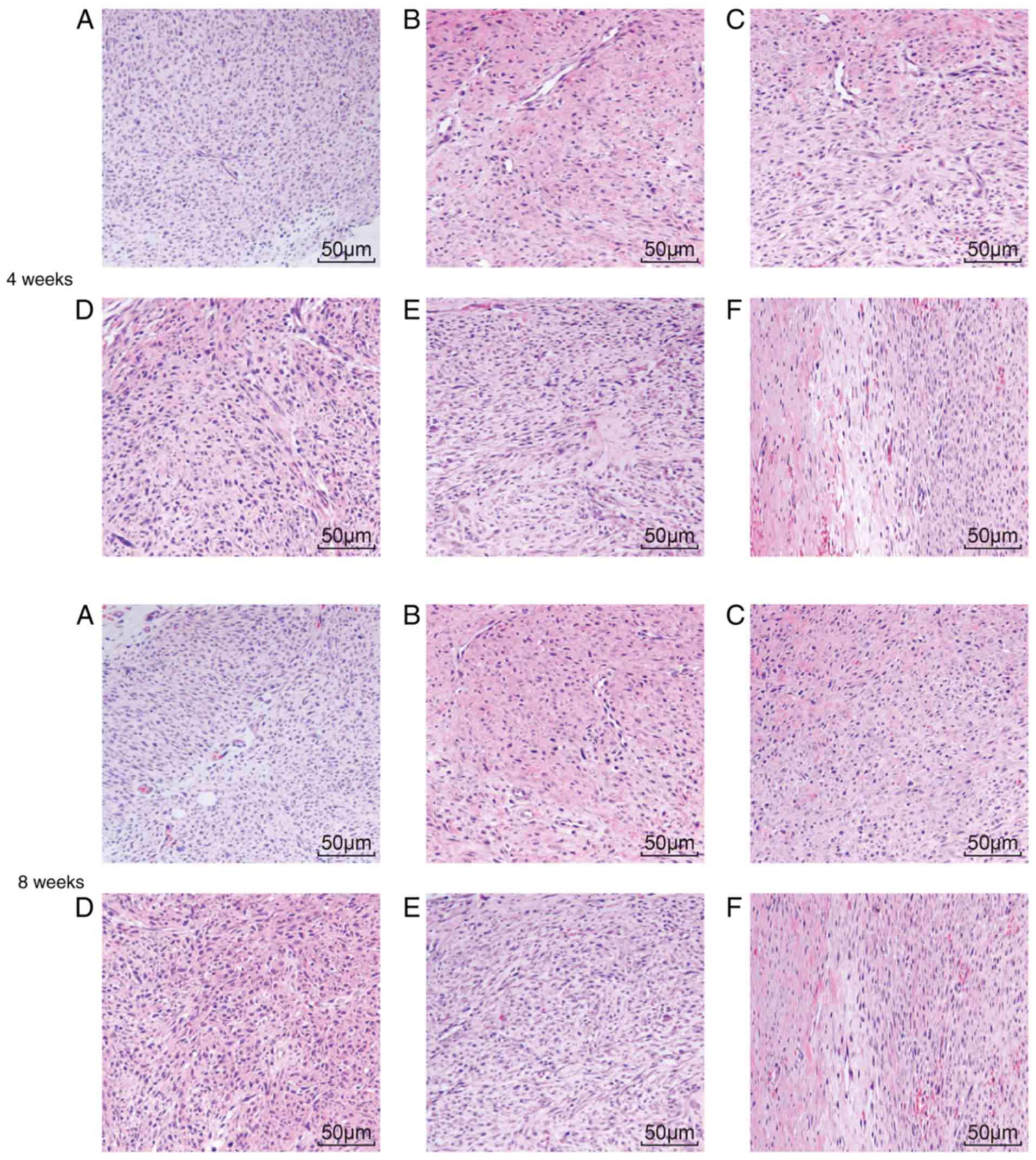
Morphological changes in the tendon
tissues in rats with tendon injuries
A microscope was used to thoroughly examine
morphological changes in blood vessels and collagenous fiber. Four
weeks post-surgery, the model, NC-siRNA and combined groups
exhibited active fibroblast and collagen fiber proliferation, and
the infiltration of inflammatory cells and collagen fibers at the
broken ends in addition to the anastomosis site, including the
production of active phagocytes. Substantial capillary
proliferation was observed in the paratendon at the anastomosis
site, with notable congestive edema and infiltration of
inflammatory cells, which mainly included neutrophils and
monocytes. The inflammation at the anastomosis site was categorized
into 3 grades. Comparatively, fewer inflammatory cells were
observed in the TP group, and the inflammatory response was
characterized as grade 2. The HSP70-siRNA group exhibited a higher
number of inflammatory cells in comparison with the normal group.
Given this increase, the inflammatory response was characterized as
grade 4. At week 8 following surgery, congestive edema and
infiltration of inflammatory cells at the broken ends of the tendon
were reduced in the model, NC-siRNA and combined groups.
Substantial fibroblast proliferation was observed in the tendon
parenchyma close to the broken ends, and newly formed collagen was
additionally observed in the broken ends. The number of novel
capillaries formed was increased, whereas the caliber was broader
compared with that noted one week prior. The TP group presented a
regular arrangement of the collagen fibers that were almost
parallel with the long axis of the tendon. The number of
infiltrated inflammatory cells and capillaries at the anastomosis
site and paratendon were considerably reduced compared with that
noted 4 weeks previously. The inflammatory response was reduced to
grade 1. HSP70-siRNA resulted in an irregular arrangement of
collagen fibers at the anastomosis site. Net migration of the
tendon fibroblasts from the anastomosis site to the outer tendon
was observed. Four weeks following surgery, the TP group exhibited
a notable difference in the optimized infiltration of inflammatory
cells at the early stage and a more regular arrangement of collagen
fibers in the middle and late stages compared with the model,
NC-siRNA, HSP70-siRNA and combined groups (Table III). These results demonstrated
that TP optimized infiltration of inflammatory cells and the
collagenous arrangement.
| Table III.Inflammatory response grading. |
Table III.
Inflammatory response grading.
|
|
| Grade |
|---|
| Group | Sample | 4th week | 8th week |
|---|
| Normal | 10 | 0 | 0 |
| Model | 10 | 3 | 2 |
| Negative
control-siRNA | 10 | 3 | 2 |
| Combined | 10 | 3 | 2 |
| Thermal
pretreatment | 10 | 2 | 1 |
| Heat shock
protein | 10 | 4 | 3 |
| 70-siRNA |
|
|
|
TP increases the maximum tensile force
and tendon gliding distance in rats with tendon injury
Biomechanical tests were performed to detect the
maximum tensile strength along with the tendon sliding distance. As
presented in Table IV, at week 4
post-surgery, significant differences were identified in maximum
tensile strength and tendon gliding distance in all 5 groups
compared with the normal group (P<0.05). Compared with the model
group, the NC-siRNA and combine groups exhibited no differences in
maximum tensile strength and tendon gliding distance (P>0.05).
In contrast, the TP and HSP70-siRNA groups presented a
significantly greater maximum tensile strength and tendon gliding
distance compared with the model group (P<0.05). At week 8
following surgery, no significant differences were noted in the
tendon gliding distance between the normal and TP groups
(P>0.05). These results suggested that TP positively increased
the tensile strength and tendon sliding distance.
| Table IV.Maximum tensile strength and tendon
gliding distance 4 and 8 weeks following surgery. |
Table IV.
Maximum tensile strength and tendon
gliding distance 4 and 8 weeks following surgery.
|
|
| Maximum tensile
strength (N) | Tendon gliding
distance (mm) |
|---|
|
|
|
|
|
|---|
| Group | Sample | 4th week | 8th week | 4th week | 8th week |
|---|
| Normal | 10 | 84.48±4.34 | 88.47±5.41 | 3.30±0.34 | 3.26±0.45 |
| Model | 10 |
61.55±3.41a |
70.67±4.67a |
1.90±0.27a |
1.86±0.22a |
| Negative
control-siRNA | 10 |
59.45±3.56a |
69.58±4.42a |
1.94±0.17a |
1.82±0.32a |
| Combine | 10 |
60.54±4.21a |
69.77±3.56a |
1.91±0.57a |
1.80±0.38a |
| Thermal
pretreatment | 10 |
79.05±4.68a,b |
87.34±4.53b |
2.51±0.41a,b |
3.21±0.24b |
| Heat shock protein
70-siRNA | 10 |
45.78±4.32a,b |
51.67±3.78a,b |
1.34±0.56a,b |
1.21±0.58a,b |
Pathological changes in the tendon
tissues in rats with tendon injuries
H&E staining was used to detect any pathological
changes the tendon tissue in SD male rats. At weeks 4 and 8
following surgery, the normal group exhibited intact collagen
fibers, orderly arranged tendon cells with clear nuclei of uniform
size, evenly distributed chromo-plasm and an intact nuclear
membrane without cell swelling. At week 4 subsequent to surgery,
irregular collagen fibers were observed at the tendon anastomotic
sites in the model, NC-siRNA and combined groups. In addition, few
fibroblasts and collagenocytes of irregular shape and inflammatory
cell infiltration were observed in the peripheral tissue. Collagen
fibers observed in the rat tendons at week 8 following surgery were
arranged in a more orderly manner compared with those observed at
the 4th week following surgery. Collagen fibers at the tendon
anastomotic sites in the TP group at week 8 following surgery were
more orderly arranged compared with those at 4th week post-surgery.
In addition, collagen fibers and fibroblasts exhibited a regular
arrangement. At the 8th week following surgery, considerable scar
tissue with pleomorphic fiber formed at the incision site of the
rat tendons in the model group, whereas the scar tissues of the TP
group were more mature and better aligned, exhibiting lower
inflammatory cell infiltration. At week 4 and 8 post-surgery, the
HSP70-siRNA group exhibited extremely disordered collagen fibers at
the tendon anastomotic sites (Fig.
3A-F). These results indicated that TP may improve inflammatory
cell infiltration and collagenous arrangement.
TP increases HSP70-positive expression
in the tendon tissues
Immunohistochemistry was used to measure the
positive expression of the HSP70 protein. The tendon cells
exhibiting yellow-brown particles in either the cytoplasm or
nucleus were considered to be HSP70 positive. The
immunohistochemical results demonstrated that HSP70 protein
expression was particularly low in the normal group and the highest
in the TP group. At week 8 following surgery, HSP70 protein
expression levels significantly increased in the TP group compared
with the normal group (P<0.05). The normal, model, NC-siRNA and
combined groups exhibited various levels of HSP70 expression, but
these levels were lower compared with the TP group. HSP70
expression levels in the HSP70-siRNA group were also significantly
reduced compared with the normal group (P<0.05; Fig. 4A-B). Thus, TP increased HSP70
protein expression.
TP increases HSP70 mRNA expression in
tendon tissues
RT-qPCR was used to detect HSP70 mRNA levels. As
presented in Fig. 5A-B, compared
with the control (normal) group, significant differences in HSP70
mRNA expression levels were noted among the 5 groups (P<0.05).
Compared with the control group, HSP70 mRNA levels were
significantly increased in the TP group (P<0.05) but reduced in
the HSP70-siRNA group (P<0.05). Compared with the model group,
HSP70 mRNA levels between the NC-siRNA group and the combined group
exhibited no significant differences (P>0.05). HSP70 mRNA levels
were significantly enhanced in the TP group (P<0.05) and reduced
in the HSP70-siRNA group (P<0.05) compared with the model group.
These results indicated that TP may increase HSP70 mRNA levels.
TP increases HSP70, TGF-β and IGF-1
protein levels
Western blot analysis was used to detect HSP70,
TGF-β and IGF-1 protein levels. As presented in Fig. 6A-D, compared with the control
group, significant differences in HSP70, TGF-β and IGF-1 protein
levels were noted in the 5 other groups (P<0.05). Compared with
the control group, HSP70 protein expression levels were
significantly enhanced in the TP group (P<0.05) but reduced in
the HSP70-siRNA group (P<0.05). Compared with the model group,
HSP70, TGF-β and IGF-1 protein levels between the NC-siRNA and
combined groups did not differ (P>0.05), and HSP70, TGF-β and
IGF-1 protein levels were significantly increased in the TP group
(P<0.05) but decreased in the HSP70-siRNA group (P<0.05). The
aforementioned results demonstrated that TP enhances HSP70, TGF-β
and IGF-1 protein levels.
 | Figure 6.Western blot analysis demonstrated
that TP increased HSP70, TGF-β and IGF-1 protein levels. There were
10 rats in each group, with 5 rats observed 4 weeks following
surgery and 5 rats observed 8 weeks following surgery. (A)
Quantification analysis of HSP70, TGF-β and IGF-1 protein levels in
each group at the 4th week following surgery. (B) Protein bands of
HSP70, TGF-β and IGF-1 4 weeks following surgery in each group. (C)
Quantification analysis of HSP70, TGF-β and IGF-1 protein levels in
each group at the 8th week following surgery. (D) Protein bands of
HSP70, TGF-β and IGF-1 8 weeks following surgery in each group.
Data were analyzed by repeat measures analysis of variance.
*P<0.05 vs. the normal group; #P<0.05 vs. the
model group. NC, negative control; TP, thermal pretreatment; HSP,
heat shock protein; siRNA, small interfering RNA; TGF-β, tumor
growth factor β; IGF-1, insulin-like growth factor 1. |
Discussion
Tendon injury often results in the formation of
tendon adhesion, causing the deterioration of the range of motion
of the joint (21). Patellar
tendon adhesion substantially influences patellofemoral and
tibiofemoral biomechanics and exhibits a close association with
tendon shortening, changes of patellofemoral contact and
tibiofemoral contact and knee arthrofibrosis (22). HSP-72 is upregulated in response to
heat and mechanical stress, and this response is mediated by human
fibroblasts and involves the nuclear translocation of HSP-72
(23). In the present study, the
effect of HSP70 expression following the completion of TP on tendon
healing and adhesion was investigated. The results revealed that TP
increased HSP70 expression while reducing traumatic inflammation
and postoperative adhesion in the tendon, thereby improving the
tendon healing process in rats.
One of the major conclusions is that tendon adhesion
is associated with HSP70. When HSP70 expression was reduced, tendon
adhesion increased. HSPs are ubiquitous proteins that serve
critical functions in the response to biotic and abiotic stresses
(24). A previous study by
Peterson et al (25)
demonstrated that HSPs promote epithelial wound healing in various
tissues, whereas reduced HSP70 expression may potentially prolong
the healing process in the pathologic cornea of animals. Certain
HSP70s, including molecular chaperones, serve a pivotal function in
various biological processes, including the regulation of protein
folding, interactions between proteins and translocation and
degradation across membranes (26). A previous study indicated that
HSP70 suppresses heat-induced apoptosis via inactivation of BCL2
associated X, apoptosis regulator; thus, preventing the release of
pro-apoptotic factors from the mitochondrial intermembrane space
(27). HSP also serves a
cytoprotective function in stressed muscle (28). Furthermore, a study consistent with
the results of the present study demonstrated that the HSP70
expression level in chondrocytes exhibited a close association with
the histological severity in osteoarthritis (29). In addition, HSP70 overexpression
promotes the metabolic activity of chondrocytes while protecting
the chondrocytes from multiple stresses (30).
In addition, the present study provided evidence
that TP increased HSP70 expression while reducing traumatic
inflammation and tendon adhesion following surgery. TP alters
tissue stiffness through the regulation of collagen composition,
altering tissue susceptibility to histotripsy (31). As previously reported, TP is
associated with the induction of HSP70 protein synthesis, and this
mechanism may mitigate tissue damage in lung fibrosis (32). Li et al (33) also identified that appropriate TP
may substantially increase HSP70 expression levels, reduce serum
aspartate aminotransferase and alanine aminotransferase levels,
decrease liver injury and promote liver repair following carbon
tetrachloride-induced liver injury. In contrast, HSPs additionally
influence cell protection, inflammation and immune system function
(34). In addition, as an
anti-apoptotic chaperone and through its chaperone activity, HSP70
tightly regulates inflammatory responses via the nuclear factor-κβ
signaling pathway, which is a critical pathway in mediating
inflammatory responses (35). A
previous study additionally demonstrated that TP reduced tendon
adhesion via upregulating HSP72 expression (36). Furthermore, HSPs serve a
substantial role in dealing with environmental stresses, including
heat stress, whereas TP increases the mRNA levels of rhopalosiphum
padi HSPs (37).
To conclude, the results obtained in the present
study also demonstrated that TP promoted tendon healing via its
increase in tensile strength and tendon sliding distance. Heat
stress contributed to satellite cell proliferation and protein
synthesis in the repair of injured skeletal muscle. Thus, the
recovery of injured skeletal muscle may benefit from heating
(38). HSP70, TGF-β and IGF-1
protein levels were found to be increased in the TP group but
decreased in the HSP70-siRNA group in the present study. TGF-β
exerts a positive effect on tendon healing through the knockout of
TGF-β inducible early gene in the absence of the Smad pathway
(39). One study demonstrated that
increased TGF-β expression of bone marrow-derived mesenchymal stem
cells facilitates tendon healing via modulating the TGF-β/mitogen
activated protein kinase signaling pathway (40). Furthermore, TGF-β1 and IGF-1 served
a notable role in tendon cell proliferation and tendon tissue
regeneration, and these processes were beneficial in the tendon
healing process by regulating tenocyte proliferation and collagen
metabolism (41). Another study
also demonstrated that IGF-1, as an exogenous modulator, served a
critical role in the promotion of tendon healing without promoting
tendon adhesion given that it induces cell proliferation and
collagen production and functions as strong angiogenetic factor
(42). Therefore, the present
study hypothesized that the increased expression of TGF-β and IGF-1
induced by TP promotes tendon healing.
In conclusion, increased HSP70 expression following
TP contributes to tendon healing in rats through attenuating
traumatic inflammation and reducing postoperative tendon adhesion.
Therefore, the present study provided an experimental baseline for
the prevention of tendon adhesion in the clinic. However, due to
the lack of a guaranteed prevention/treatment method, further
studies are mandatory to obtain a more comprehensive understanding
of the specific mechanisms involved with increased HSP70 expression
following TP and how it benefits tendon healing.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
XMT and JD wrote the paper, and conceived and
designed the experiments; HLS analyzed the data; XMT and HLS
collected and provided the samples for this study. JD wrote and
revised the paper. All the authors approved the final version of
the paper.
Ethics approval and consent to
participate
The animal experiments were conducted in accordance
with the Guide for the Care and Use of Laboratory Animals as
promulgated by the National Institutes of Health. Ethical approval
was obtained from the ethics committee of The Affiliated Huai'an
No. 1 People's Hospital of Nanjing Medical University (Huai'an,
China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Longo UG, Lamberti A, Maffulli N and
Denaro V: Tissue engineered biological augmentation for tendon
healing: A systematic review. Br Med Bull. 98:31–59. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bauge C, Leclercq S, Conrozier T and
Boumediene K: TOL19-001 reduces inflammation and MMP expression in
monolayer cultures of tendon cells. BMC Complement Altern Med.
15:2172015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wu PT, Jou IM, Kuo LC and Su FC:
Intratendinous injection of hyaluronate induces acute inflammation:
A possible detrimental effect. PLoS One. 11:e01554242016.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Geary MB, Orner CA, Bawany F, Awad HA,
Hammert WC, O'Keefe RJ and Loiselle AE: Systemic ep4 inhibition
increases adhesion formation in a murine model of flexor tendon
repair. PLoS One. 10:e01363512015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Thomopoulos S, Das R, Sakiyama-Elbert S,
Silva MJ, Charlton N and Gelberman RH: bFGF and PDGF-BB for tendon
repair: Controlled release and biologic activity by tendon
fibroblasts in vitro. Ann Biomed Eng. 38:225–234. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Dabak TK, Sertkaya O, Acar N, Donmez BO
and Ustunel I: The effect of phospholipids (surfactant) on adhesion
and biomechanical properties of tendon: A rat achilles tendon
repair model. Biomed Res Int. 2015:6893142015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ackermann PW, Franklin SL, Dean BJ, Carr
AJ, Salo PT and Hart DA: Neuronal pathways in tendon healing and
tendinopathy-update. Front Biosci (Landmark Ed). 19:1251–1278.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mulhall KJ, McLaughlin R, Kay E, Kiely P,
Bouchier-Hayes D and Murray P: Thermal preconditioning prevents
peritendinous adhesions and inflammation. Clin Orthop Relat Res.
405:258–266. 2002. View Article : Google Scholar
|
|
9
|
Zhang L, Fok JH and Davies FE: Heat shock
proteins in multiple myeloma. Oncotarget. 5:1132–1148. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Afzal E, Ebrahimi M, Najafi SM, Daryadel A
and Baharvand H: Potential role of heat shock proteins in neural
differentiation of murine embryonal carcinoma stem cells (P19).
Cell Biol Int. 35:713–720. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Rauch JN and Gestwicki JE: Binding of
human nucleotide exchange factors to heat shock protein 70 (Hsp70)
generates functionally distinct complexes in vitro. J Biol Chem.
289:1402–1414. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Vazzana M, Siragusa T, Arizza V, Buscaino
G and Celi M: Cellular responses and HSP70 expression during wound
healing in Holothuria tubulosa (Gmelin, 1788). Fish Shellfish
Immunol. 42:306–315. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lancaster GI and Febbraio MA:
Exosome-dependent trafficking of HSP70: A novel secretory pathway
for cellular stress proteins. J Biol Chem. 280:23349–23355. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mushtaq S, Naqvi ZA, Siddiqui AA and Ahmed
N: Albumin precursor and Hsp70 modulate corneal wound healing in an
organ culture model. Acta Histochem. 113:36–42. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Guide for the Care and Use of Laboratory
Animals, . Washington (DC): National Academies Press (US); 2011,
vol. 10.17226/12910.
|
|
16
|
Cury DP, Schäfer BT, de Almeida SRY,
Righetti MMDS and Watanabe IS: Application of a purified protein
from natural latex and the influence of suture type on achilles
tendon repair in rats. Am J Sports Med. 47:901–914. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Han Q, Li L, Liang H, Li Y, Xie J and Wang
Z: Downregulation of lncRNA X inactive specific transcript (XIST)
suppresses cell proliferation and enhances radiosensitivity by
upregulating mir-29c in nasopharyngeal carcinoma cells. Med Sci
Monit. 23:4798–4807. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kim KM, Wagle S, Moon YJ, Wang SI, Park
BH, Jang KY and Kim JR: Interferon β protects against avascular
osteonecrosis through interleukin 6 inhibition and silent
information regulator transcript-1 upregulation. Oncotarget.
9:3562–3575. 2017.PubMed/NCBI
|
|
19
|
Dumas P, Morin MD, Boquel S, Moffat CE and
Morin PJ: Expression status of heat shock proteins in response to
cold, heat, or insecticide exposure in the Colorado potato beetle
Leptinotarsa decemlineata. Cell Stress Chaperones 2019. (Epub ahead
of print).
|
|
20
|
Song H, Xu Y, Shi L, Xu T, Fan R, Cao M,
Xu W and Song J: LncRNA THOR increases the stemness of gastric
cancer cells via enhancing SOX9 mRNA stability. Biomed
Pharmacother. 108:338–346. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Loiselle AE, Bragdon GA, Jacobson JA,
Hasslund S, Cortes ZE, Schwarz EM, Mitten DJ, Awad HA and O'Keefe
RJ: Remodeling of murine intrasynovial tendon adhesions following
injury: MMP and neotendon gene expression. J Orthop Res.
27:833–840. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fernandez JW, Akbarshahi M, Crossley KM,
Shelburne KB and Pandy MG: Model predictions of increased knee
joint loading in regions of thinner articular cartilage after
patellar tendon adhesion. J Orthop Res. 29:1168–1177. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jagodzinski M, Hankemeier S, van Griensven
M, Bosch U, Krettek C and Zeichen J: Influence of cyclic mechanical
strain and heat of human tendon fibroblasts on HSP-72. Eur J Appl
Physiol. 96:249–256. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu J, Wang R, Liu W, Zhang H, Guo Y and
Wen R: Genome-wide characterization of heat-shock protein 70s from
chenopodium quinoa and expression analyses of cqhsp70s in response
to drought stress. Genes (Basel). 9:E352018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Peterson C, Driskell E, Wilkie D,
Premanandan C and Hamor R: Heat-shock protein 70 expression in the
equine cornea. Vet Ophthalmol. 20:344–348. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kampinga HH and Craig EA: The HSP70
chaperone machinery: J proteins as drivers of functional
specificity. Nat Rev Mol Cell Biol. 11:579–592. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Stankiewicz AR, Lachapelle G, Foo CP,
Radicioni SM and Mosser DD: Hsp70 inhibits heat-induced apoptosis
upstream of mitochondria by preventing Bax translocation. J Biol
Chem. 280:38729–38739. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Inoue T, Suzuki S, Hagiwara R, Iwata M,
Banno Y and Okita M: Effects of passive stretching on muscle injury
and HSP expression during recovery after immobilization in rats.
Pathobiology. 76:253–259. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Son YO, Kim HE, Choi WS, Chun CH and Chun
JS: RNA-binding protein ZFP36L1 regulates osteoarthritis by
modulating members of the heat shock protein 70 family. Nat Commun.
10:772019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tonomura H, Takahashi KA, Mazda O, Arai Y,
Shin-Ya M, Inoue A, Honjo K, Hojo T, Imanishi J and Kubo T: Effects
of heat stimulation via microwave applicator on cartilage matrix
gene and HSP70 expression in the rabbit knee joint. J Orthop Res.
26:34–41. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Vlaisavljevich E, Xu Z, Arvidson A, Jin L,
Roberts W and Cain C: Effects of thermal preconditioning on tissue
susceptibility to histotripsy. Ultrasound Med Biol. 41:2938–2954.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hagiwara S, Iwasaka H, Matsumoto S,
Noguchi T and Yoshioka H: Association between heat stress protein
70 induction and decreased pulmonary fibrosis in an animal model of
acute lung injury. Lung. 185:287–293. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Li SQ, Wang DM, Shu YJ, Wan XD, Xu ZS and
Li EZ: Proper heat shock pretreatment reduces acute liver injury
induced by carbon tetrachloride and accelerates liver repair in
mice. J Toxicol Pathol. 26:365–373. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kasperska-Zajac A, Damasiewicz-Bodzek A,
Bieniek K, Skrzypulec-Frankel A, Tyrpien-Golder K and Grzanka A:
Elevated circulating heat shock protein 70 and its antibody
concentrations in chronic spontaneous urticaria. Int J Immunopathol
Pharmacol. 31:3946320177504402018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sevin M, Girodon F, Garrido C and de
Thonel A: HSP90 and HSP70: Implication in inflammation processes
and therapeutic approaches for myeloproliferative neoplasms.
Mediators Inflamm. 2015:9702422015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Tan Y, Wu QF, Wu Q, Tan XT, Chen LB and
Wang X: Thermal preconditioning may prevent tendon adhesion by
up-regulating hsp72 in rats. Cell Physiol Biochem. 42:1623–1634.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Li Y, Zhao Q, Duan X, Song C and Chen M:
Transcription of four Rhopalosiphum padi (L.) heat shock protein
genes and their responses to heat stress and insecticide exposure.
Comp Biochem Physiol A Mol Integr Physiol. 205:48–57. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kojima A, Goto K, Morioka S, Naito T,
Akema T, Fujiya H, Sugiura T, Ohira Y, Beppu M, Aoki H and Yoshioka
T: Heat stress facilitates the regeneration of injured skeletal
muscle in rats. J Orthop Sci. 12:74–82. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Tsubone T, Moran SL, Subramaniam M, Amadio
PC, Spelsberg TC and An KN: Effect of TGF-beta inducible early gene
deficiency on flexor tendon healing. J Orthop Res. 24:569–575.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wang R, Xu B and Xu HG: Up-regulation of
tgf-beta promotes tendon-to-bone healing after anterior cruciate
ligament reconstruction using bone marrow-derived mesenchymal stem
cells through the tgf-beta/mapk signaling pathway in a new zealand
white rabbit model. Cell Physiol Biochem. 41:213–226. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chen YJ, Wang CJ, Yang KD, Kuo YR, Huang
HC, Huang YT, Sun YC and Wang FS: Extracorporeal shock waves
promote healing of collagenase-induced achilles tendinitis and
increase TGF-beta1 and IGF-I expression. J Orthop Res. 22:854–861.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Berglund ME, Hart DA, Reno C and Wiig M:
Growth factor and protease expression during different phases of
healing after rabbit deep flexor tendon repair. J Orthop Res.
29:886–892. 2011. View Article : Google Scholar : PubMed/NCBI
|