Introduction
The Wnt signaling pathway is able to function via
two routes: A canonical (Wnt/β-catenin) pathway or a non-canonical
Wnt pathway (1). In the canonical Wnt
signaling pathway, when the Wnt ligand does not bind to its
receptors, the pathway is ‘switched off’. In this case, β-catenin
is degraded by a destruction complex comprising Axin, adenomatous
polyposis coli and glycogen synthase kinase 3β (Gsk3β). β-catenin
is phosphorylated in the cytosol, an event which is followed by
ubiquitination, resulting in proteasomal degradation (2). When the Wnt pathway is ‘switched on’,
cytosolic β-catenin is stabilized, resulting in β-catenin
accumulation in the cytoplasm, and subsequent nuclear entry. Once
inside the nucleus, β-catenin activates Wnt target genes by binding
to transcription factors of the T-cell factor (TCF)/ lymphoid
enhancer factor (LEF) family (3).
Wnt signaling is a crucial mechanism involved in the
regulation of cell proliferation, differentiation and morphogenesis
(4). Numerous studies have suggested
that abnormal activation of the Wnt/β-catenin signaling pathway may
have a significant role in human tumorigenesis, including in
colorectal cancer (CRC), melanoma and lung cancer (5–7).
Therefore, the screening of inhibitors of the Wnt signaling pathway
is significant in the search for potential anticancer drugs.
Apigenin is one of the most commonly occurring
flavonoids, and is found in numerous plants, including parsley,
onions, orange, tea, chamomile, wheat sprouts and certain
condiments (8). Apigenin has
exhibited potential for use in cancer prevention and therapy, and
suppresses cell growth in a number of human cancer cell lines,
including skin, colon, thyroid, breast, leukemia and prostate
cancer cells (9–13). Although previous studies have
indicated that the anti-carcinogenic mechanism of apigenin occurs
via certain signaling pathways, including the nuclear factor-κB,
P53 and mitogen activated protein kinase pathways, to the best of
our knowledge there have been no previous reports indicating a
contributory role of apigenin in the Wnt/β-catenin signaling
pathway. Therefore, the present study aimed to investigate the
effect of apigenin on the migration and invasion of CRC cells, as
well as the inhibition of the Wnt/β-catenin signaling pathway.
Materials and methods
Cell lines, cell culture and
reagents
Apigenin was purchased from Sigma-Aldrich (St.
Louis, MO, USA). A 10 mM solution of apigenin was prepared in
sterile water, stored at −20°C and protected from light, and
diluted to the required concentrations for each experiment. Human
CRC cell lines (SW480 and HCT15) were purchased from the American
Type Culture Collection (Manassas, VA, USA), cultured and
maintained in Dulbecco's modified Eagle's medium (DMEM; HyClone,
Logan, UT, USA), supplemented with 10% heated-inactivated fetal
bovine serum (FBS), 100 U/ml penicillin, 10 µg/ml streptomycin
sulfate and then held at 37°C in a humidified 5% CO2
incubator. HEK293T cells were purchased from the Cell Bank of the
Chinese Academy of Sciences (Shanghai, China). Mitomycin C was
purchased from Roche Diagnostics (Shanghai, China).
Cell viability assay
SW480 and HCT15 human colorectal cancer cells
(2×104 cells⁄well) were treated with various
concentrations (0, 5, 10, 20, 40 and 80 µM) of apigenin for 48 h.
Cell viability was determined by MTT assay (Sigma-Aldrich) using a
Bio-Rad microplate reader (Model 680; Bio-Rad Laboratories, Inc.,
Hercules, CA, USA) (14).
Wound healing assay
SW480 cells were cultured to full confluence in
6-well plates and subsequently incubated with 10 µg/ml mitomycin C
for 2 h to inactivate cell proliferation. Cells were washed with
phosphate-buffered saline (PBS) and wounds were made with sterile
pipette tips. DMEM supplemented with 0.5% FBS was added into wells
with various concentrations (0, 20 and 40 µM) of apigenin. Images
of the cells were captured using an Olympus DX41 inverted
microscope (Olympus Corp., Tokyo, Japan), following 10 h of
incubation with the aforementioned reagents.
Transwell migration assay
Transwells (Corning Inc., Corning, NY, USA) were
coated with 50 µl growth factor reduced Matrigel (1:8 dilution; BD
Biosciences, Franklin Lakes, NJ, USA) for 30 min in a cell
incubator. The bottom chambers of the Transwells were filled with
DMEM containing 0.5% FBS and the top chambers were seeded with
mitomycin C inactivator, SW480 cells (4×104 cells/well)
in 100 µl DMEM (0.5% FBS) and various concentrations (0, 20 and 40
µM) of apigenin. Following an overnight incubation, the cells on
the top surface of the membrane (non-migrated cells) were removed
with a cotton swab and the cells which had migrated onto the bottom
sides of the membrane (invasive cells) were then fixed with cold 4%
paraformaldehyde and stained with 2% crystal violet (Beyotime
Institute of Biotechnology, Inc., Shanghai, China). Images were
captured using an Olympus DX41 inverted microscope.
Wnt/β-catenin signal reporter
assay
HEK293T and SW480 cells were plated at a
concentration of 5,000 cells/well on white-bottomed 96-well plates.
Cells were serum-starved overnight and co-transfected with 0.2 µg
TOPflash or FOPflash expression plasmids (EMD Millipore, Billerica,
MA, USA) and 0.1 µg pRL-TK (Renilla TK-luciferase vector;
Promega Corp., Madison, WI, USA) as a control, using
Lipofectamine® 2000 (Invitrogen Life Technologies,
Carlsbad, CA, USA). Cells were subsequently treated with LiCl (20
mM/ml) for 48 h prior to the measurement of luciferase activity.
Luciferase activity was measured using a Glomax 96 Microplate
Luminometer (Turner BioSystems, Sunnyvale, CA, USA). Firefly
luciferase activity was normalized for transfection efficiency by
dividing the results by the Renilla luciferase activity. The
TOP/FOP ratio was used as a measure of β-catenin-driven
transcription. Average activity and standard deviations were
derived from 8 transfected samples.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
RNA was extracted using the RNeasy mini kit (Qiagen,
Valencia, CA, USA) according to the manufacturer's instructions.
Final concentrations were determined using the Nanodrop DA-1000
Spectrophotometer (Thermo Fisher Scientific, Wilmington, DE, USA).
RNA (1 µg) was purified from genomic DNA using DNase I (Invitrogen
Life Technologies) and reverse transcribed to complementary DNA
(cDNA) using the QuantiTect W Reverse Transcription kit (Qiagen)
according to the manufacturer's instructions. The resulting cDNA
product was then used as a template for PCR amplification. A 25 µl
qPCR reaction mix, consisting of 25 ng diluted cDNA, SYBR Green Dye
(Qiagen) and 0.1 µM of each qPCR primer pair, was amplified to
obtain quantifiable expressions of Wnt downstream genes in SW480
cells. All qPCR was conducted in a Stratagene MxPro™ 3005P (Agilent
Technologies, Inc., Santa Clara, CA, USA). Primer sequences were as
follows: Axin2 forward, 5′-CAACACCAGGCGGAACGAA-3′ and reverse,
5′-GCCCAATAAGGAGTGTAAGGACT-3′; C-myc forward,
5′-ATGGCCCATTACAAAGCCG-3′ and reverse,
5′-TTTCTGGAGTAGCAGCTCCTAA-3′; CylinD1 forward,
5′-GCTGCGAAGTGGAAACCATC-3′ and reverse,
5′-CCTCCTTCTGCACACATTTGAA-3′; Ephb2 forward,
5′-AGAAACGCTAATGGACTCCACT-3′ and reverse,
5′-GTGCGGATCGTGTTCATGTT-3′; Ephb3 forward,
5′-TGGGTAACATCTGAGTTGGCG-3′ and reverse,
5′-TGGTATGTGCGGATGGGATTC-3′.
Protein isolation and western
blotting
Standard western blotting was performed for protein
expression analyses (15). Cells were
washed twice with cold PBS and lysed on ice in
radioimmunoprecipitation assay buffer containing proteinase
inhibitors (Roche Diagnostics). Protein lysates were resolved on
10% SDS polyacrylamide gel, transferred to nitrocellulose membranes
(Beyotime Institute of Biotechnology, Inc.,)and blocked in 0.1%
Tween 20 and 5% bovine serum albumin (BSA) in Tris-buffered saline.
Proteins were probed with rabbit anti-β-catenin monoclonal antibody
(1:1,000; Cell Signaling Technology, Inc., Danvers, MA, USA) and
rabbit anti-β-actin antibody (1:10,000; Sigma-Aldrich) overnight at
4°C. The membrane was washed and visualized by incubation with
horseradish peroxidase-conjugated anti-rabbit IgG polyclonal
secondary antibodies for 1 h. Signals were detected by enhanced
chemiluminescence (Odyssey, LI-COR Biosciences, Lincoln, NE,
USA).
Isolation of intestinal crypts and
organoid culture
C57BL/6 mice were purchased from the Laboratory
Animal Center (Shanghai, China). An equal number of male (n=3) and
female (n=3) mice were kept in specific pathogen-free housing, fed
a normal diet and drinking water, and submitted to a 12-h
light/dark cycle and temperatures of 18–23°C. All animal
experiments conformed to the regulations drafted by Association for
Assessment and Accreditation of Laboratory Animal Care in Shanghai
and were approved by the Ethics Committee of Suzhou University
(Suzhou, China). The small intestine was separated from mice aged
3–5 weeks following sacrifice by cervical dislocation. After
several washes with cold PBS, tissues were divided into 5 mm
sections. Intestinal crypts were selectively isolated as previously
described (16); however, certain
modifications were made to the subsequent procedures and media in
the organoid culture. Briefly, crypts were further dissociated into
single cells using Accutase® (Innovative Cell
Technologies, Inc., San Diego, CA, USA) treatment for 10 min at
37°C and seeded on polymerized Matrigel (BD Biosciences).
Serum-free media optimized for organoid culture containing
epidermal growth factor (EGF; Peprotech, Inc., Rocky Hill, NJ,
USA), R-Spondin1 (R&D Systems, Inc., Minneapolis, MN, USA) and
Noggin (Peprotech, Inc.) were routinely supplemented with Y27632
(Wako, Osaka, Japan) and Jagged-1 (AnaSpec, Inc., Fremont, CA, USA)
to support the survival of single cells and the proliferation of
stem cells (14). The medium was
replaced every 2 days.
Immunofluorescent staining and
confocal microscopy
Cells were fixed using methanol and blocked with 5%
BSA. Cells were subsequently stained with anti-β-catenin monoclonal
antibody (1:1,000, Cell Signaling Technology, Inc.), followed by
fluorescein isothiocyanate (FITC)-conjugated goat anti-mouse
immunoglobulin G (Merck Millipore, Hong Kong, China). Cells were
then washed four times with PBS. Nuclear staining was achieved with
DAPI. Cells were imaged with a TCS SP2 spectral confocal system
(Leica Microsystems GmbH, Wetzlar, Germany). All experiments were
conducted according to the antibody manufacturer's
instructions.
Statistical analysis
Data are presented as the mean ± standard error, and
statistical comparisons between groups were made using Student's
t-test. P≤0.05 was considered to indicate a statistically
significant difference.
Results
Apigenin inhibits CRC cell line
proliferation
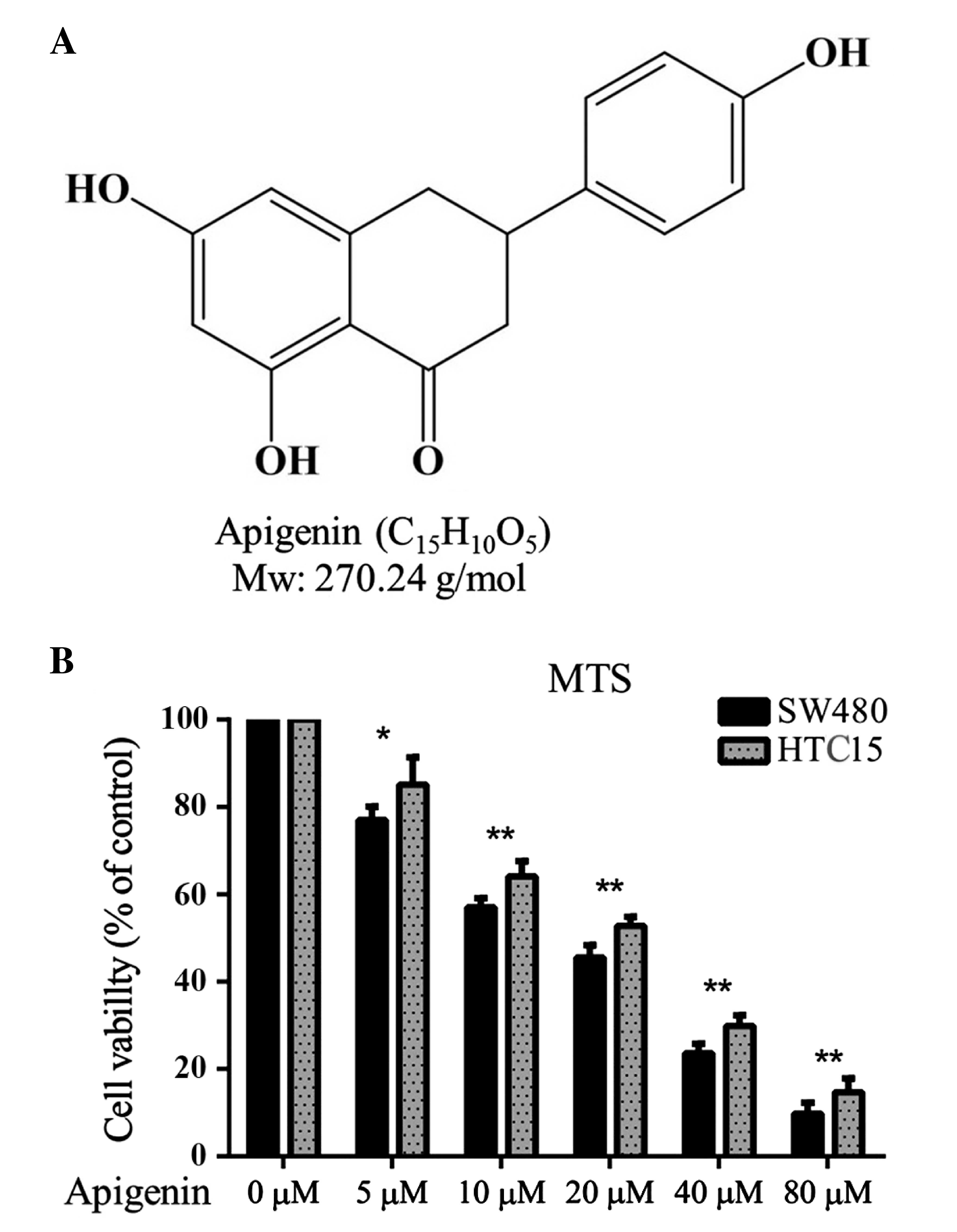
Apigenin is one of the most common flavonoids and
has a molecular weight of 270.24 g⁄mol (Fig. 1A). The in vitro
anti-proliferative effect of apigenin on the SW480 and HCT15
colorectal cancer cell lines was determined using a standard MTT
proliferation assay. The results revealed that apigenin
significantly reduced SW480 and HCT15 cell proliferation at an
IC50 of 18.17 µM and 23.57 µM, respectively. These data
suggest that apigenin may exhibit anti-CRC effects (Fig. 1B). The inhibitive action of apigenin
was greater on SW480 cells and therefore these cells were used in
subsequent experiments.
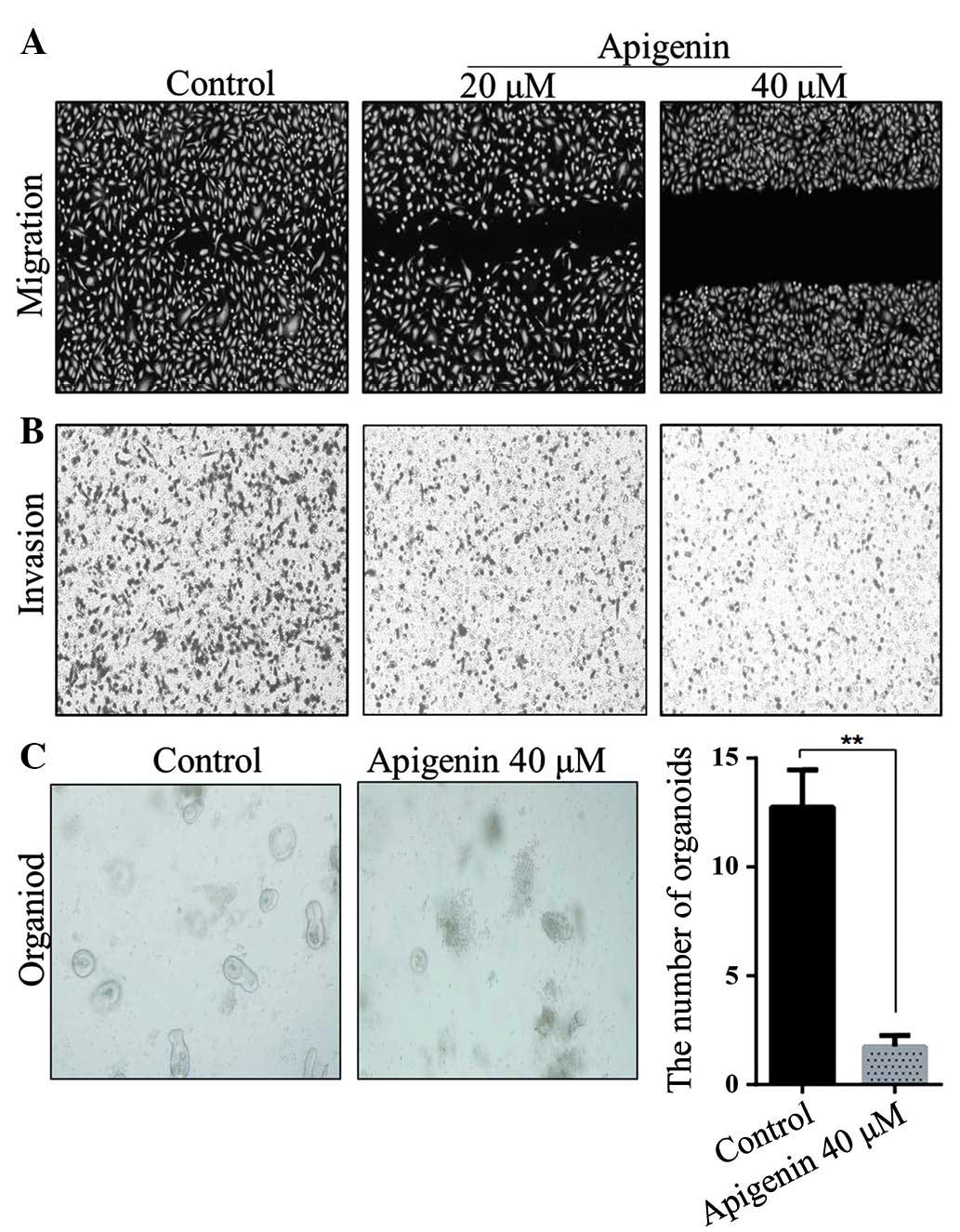
Apigenin inhibits migration and
invasion of SW480 cells and growth of intestinal organoids
Tumor migration and invasion are essential steps in
tumorigenesis (17). The effects of
apigenin on the chemotactic motility of SW480 cells were determined
using wound-healing migration and Transwell cell invasion assays.
As shown in Fig. 2A and B, apigenin
significantly inhibited SW480 cell migration and invasion at 20 and
40 µM. Furthermore, the inhibition of apigenin on the migration of
SW480 cells occurred in a dose-dependent manner. Intestinal
organoid culture in vitro requires multiple cytokines,
particularly Wnt signaling pathway cytokines. Without Wnt signaling
pathway activation, organoids are unable to grow effectively
(18). Therefore, 40 µM apigenin was
added to the intestinal organoid culture system and it was observed
that the growth of organoids was significantly inhibited (Fig. 2C).
Apigenin inhibits β-catenin/TCF/LEF
signal activation
The TOPflash/FOPflash luciferase assay is frequently
used for the analysis of β-catenin/TCF/LEF signal activation
(19). The TOPflash construct
contains two repeats of three optimal copies of the TCF/LEF binding
sites, upstream of a thymidine kinase minimal promoter that is able
to direct the transcription of the luciferase reporter gene.
FOPflash, which contains mutated TCF/LEF binding sites, is used as
a negative control (20). The Gsk3β
inhibitor LiCl activates the Wnt pathway; therefore, stimulation
with LiCl (20 mM) significantly increased the transcription of the
Wnt luciferase reporter in HEK293T and SW480 cells. Subsequently,
the effect of apigenin treatment on HEK293T and SW480 cells and
expression of the Wnt luciferase reporter was investigated.
Treatment with various concentrations of apigenin and LiCl (20 mM)
inhibited β-catenin/TCF/LEF signal activation in a dose-dependent
manner (Fig. 3A and B).
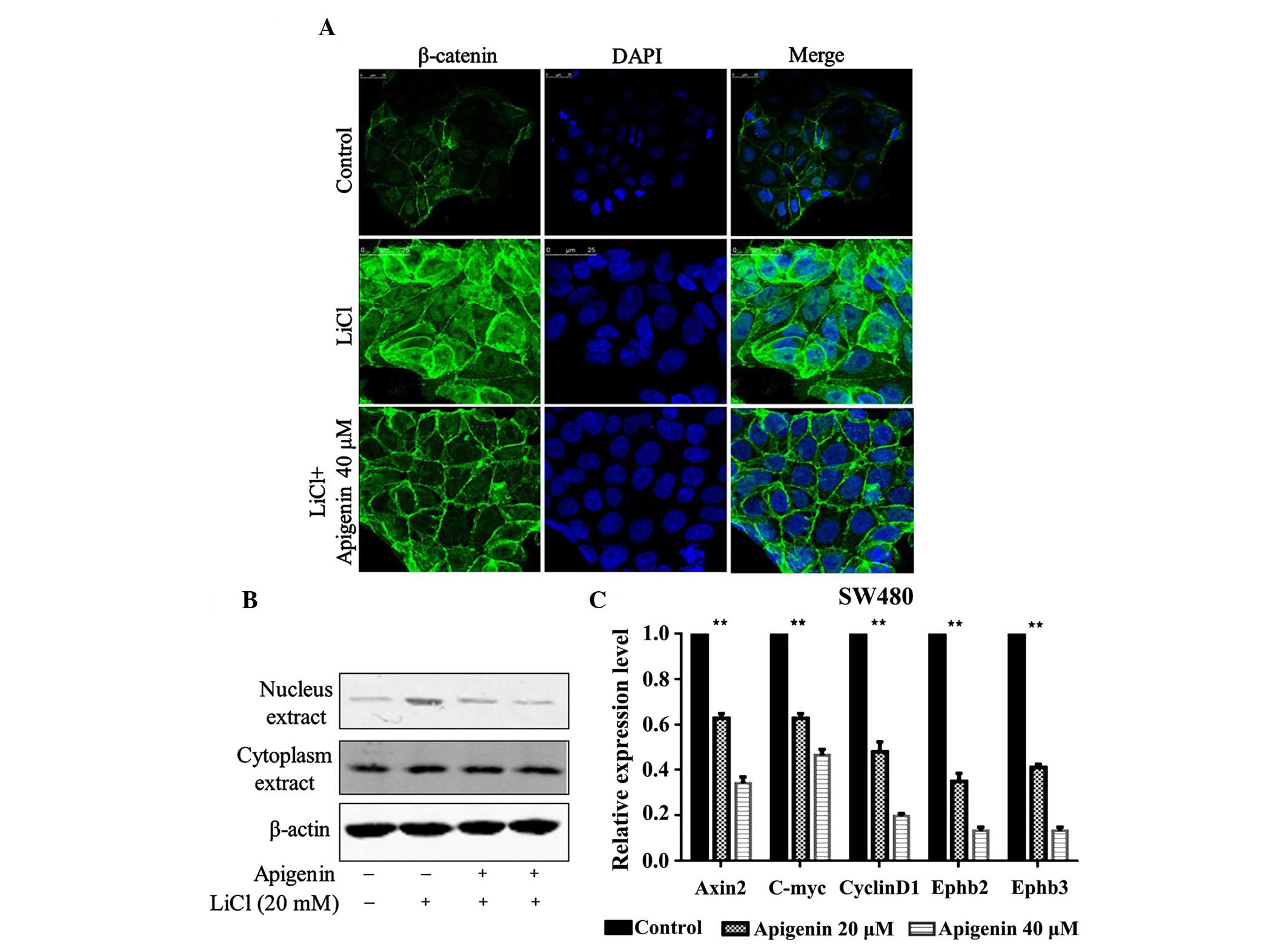
Apigenin suppresses β-catenin nuclear
entry and the expression of Wnt downstream genes
Wnt signaling activation requires β-catenin nuclear
entry (1). In order to determine how
apigenin inhibits the Wnt signaling pathway, immunofluorescent
staining was used to examine β-catenin nuclear entry in HCT15 cells
and western blotting was used to examine the expression of
β-catenin in SW480 nuclear and cytoplasmic extracts following LiCl
treatment, as well as treatment with various concentrations of
apigenin. Apigenin was found to effectively inhibit LiCl-activated
β-catenin nuclear entry (Fig. 4A) and
markedly decrease LiCl-activated expression of β-catenin in nuclear
extracts. However, apigenin had no effect on β-catenin expression
in cytosolic extracts (Fig. 4B). The
expression levels of Wnt downstream target genes, including Axin2,
C-myc, CylinD1, Ephb2 and Ephb3, were also detected. Apigenin
significantly decreased the expression levels of Wnt downstream
target genes in a dose-dependent manner (Fig. 4C).
Discussion
In the United States CRC is the third most common
type of cancer, and is second only to lung cancer in terms of
mortality, resulting in ~50,000 mortalities/year (21). Tumorigenesis in sporadic CRC is
typically initiated by an inactivating mutation in the APC gene
which induces aberrant activation of the Wnt signaling pathway
(22). Therefore, inhibition of Wnt
signaling may contribute to the treatment of CRC.
Numerous Wnt signaling inhibitors, each of which
function at various stages in the Wnt/β-catenin pathway, have been
reported to be used for the treatment of cancer (23–26). Mesd
C-terminal region peptide, similarly to the full-length Mesd
protein, was reported to inhibit Wnt 3A- and Rspodin1-induced
Wnt/β-catenin signaling in LRP5- and LRP6-expressing cells,
suppress Wnt/β-catenin signaling in HS578T human breast cancer
cells and PC-3 prostate cancer cells, and inhibit cancer cell
proliferation (27). Calcimycin, a
Wnt signaling inhibitor, targets the expression of S100A4 and
provides a functional strategy for the restriction of cell motility
in colon cancer cells (28). Thus,
the results of the present study suggested that apigenin, as a
natural compound, may inhibit β-catenin/TCF/LEF signaling
activation, which was induced by LiCl in a dose-dependent manner,
preventing β-catenin nuclear entry and therefore the expression of
Wnt downstream target genes.
Apigenin has been identified in numerous plants.
Recently, it was reported that apigenin may have potential
anti-tumor effects on multiple human cancer cell lines with low
cytotoxicity and no mutagenic activity (20,29,30). In a
study of apigenin as a potential treatment for CRC, apigenin
demonstrated inhibition of colorectal cellular migration and
invasion. It was shown to suppress the growth of CRC xenografts
through decreased phosphorylation of AKT, as well as
phosphorylation and upregulation of FADD expression (31,32). In
the present study, apigenin significantly suppressed CRC cell
proliferation, migration, invasion and intestinal organoid
growth.
In conclusion, apigenin significantly suppressed CRC
cell proliferation, migration and invasion in a dose-dependent
manner. Additionally, apigenin inhibited β-catenin/TCF/LEF
signaling activation, which was induced by LiCl in a dose-dependent
manner. Finally, apigenin inhibited β-catenin nuclear entry and
thereby prevented the expression of Wnt downstream target
genes.
Acknowledgements
The present study was financially supported by a
grant from the Open Projects of Jiangsu Provincial Commission of
Health and Family Planning (no. 2103).
References
|
1
|
Arend RC, Londoño-Joshi AI, Straughn JM Jr
and Buchsbaum DJ: The Wnt/β-catenin pathway in ovarian cancer: A
review. Gynecol Oncol. 131:772–779. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Noguti J, De Moura CF, Hossaka TA, Franco
M, Oshima CT, Dedivitis RA and Ribeiro DA: The role of canonical
WNT signaling pathway in oral carcinogenesis: A comprehensive
review. Anticancer Res. 32:873–878. 2012.PubMed/NCBI
|
|
3
|
Pecina-Slaus N: Wnt signal transduction
pathway and apoptosis: A review. Cancer Cell Int. 10:222010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Moon RT, Brown JD and Torres M: WNTs
modulate cell fate and behavior during vertebrate development.
Trends Genet. 13:157–162. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Behrens J: Control of beta-catenin
signaling in tumor development. Ann NY Acad Sci. 910:21–35. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chesire DR, Ewing CM, Sauvageot J, Bova GS
and Isaacs WB: Detection and analysis of beta-catenin mutations in
prostate cancer. Prostate. 45:323–334. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Polakis P: The oncogenic activation of
beta-catenin. Curr Opin Genet Dev. 9:15–21. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Birt DF, Mitchell D, Gold B, Pour P and
Pinch HC: Inhibition of ultraviolet light induced skin
carcinogenesis in SKH-1 mice by apigenin, a plant flavonoid.
Anticancer Res. 17:85–91. 1997.PubMed/NCBI
|
|
9
|
Caltagirone S, Rossi C, Poggi A,
Ranelletti FO, Natali PG, Brunetti M, Aiello FB and Piantelli M:
Flavonoids apigenin and quercetin inhibit melanoma growth and
metastatic potential. Int J Cancer. 87:595–600. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang IK, Lin-Shiau SY and Lin JK:
Induction of apoptosis by apigenin and related flavonoids through
cytochrome c release and activation of caspase-9 and
caspase-3 in leukaemia HL-60 cells. Eur J Cancer. 35:1517–1525.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang W, Heideman L, Chung CS, Pelling JC,
Koehler KJ and Birt DF: Cell-cycle arrest at G2/M and growth
inhibition by apigenin in human colon carcinoma cell lines. Mol
Carcinog. 28:102–110. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yin F, Giuliano AE, Law RE and Van Herle
AJ: Apigenin inhibits growth and induces G2/M arrest by modulating
cyclin-CDK regulators and ERK MAP kinase activation in breast
carcinoma cells. Anticancer Res. 21:413–420. 2001.PubMed/NCBI
|
|
13
|
Zhu Y, Mao Y, Chen H, Lin Y, Hu Z, Wu J,
Xu X, Xu X, Qin J and Xie L: Apigenin promotes apoptosis, inhibits
invasion and induces cell cycle arrest of T24 human bladder cancer
cells. Cancer Cell Int. 13:542013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang YJ, Zhou SM, Xu G and Gao YQ:
Interference of Phenylethanoid Glycosides from Cistanche tubulosa
with the MTT Assay. Molecules. 20:8060–8071. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Feliciangeli S, Bendahhou S, Sandoz G,
Gounon P, Reichold M, Warth R, Lazdunski M, Barhanin J and Lesage
F: Does sumoylation control K2P1/TWIK1 background K+ channels?
Cell. 130:563–569. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sato T, Vries RG, Snippert HJ, van de
Wetering M, Barker N, Stange DE, van Es JH, Abo A, Kujala P, Peters
PJ and Clevers H: Single Lgr5 stem cells build crypt-villus
structures in vitro without a mesenchymal niche. Nature.
459:262–265. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Miao L, Xiong X, Lin Y, Cheng Y, Lu J,
Zhang J and Cheng N: MiR-203 inhibits tumor cell migration and
invasion via caveolin-1 in pancreatic cancer cells. Oncol Lett.
7:658–662. 2014.PubMed/NCBI
|
|
18
|
Schuijers J, Junker JP, Mokry M, Hatzis P,
Koo BK, Sasselli V, van der Flier LG, Cuppen E, van Oudenaarden A
and Clevers H: Ascl2 acts as an R-spondin/Wnt-responsive switch to
control stemness in intestinal crypts. Cell stem cell. 16:158–170.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Osada T, Chen M, Yang XY, Spasojevic I,
Vandeusen JB, Hsu D, Clary BM, Clay TM, Chen W, Morse MA and Lyerly
HK: Antihelminth compound niclosamide downregulates Wnt signaling
and elicits antitumor responses in tumors with activating APC
mutations. Cancer Res. 71:4172–4182. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shen X, Ying H and Yue BY: Wnt activation
by wild type and mutant myocilin in cultured human trabecular
meshwork cells. PLoS One. 7:e449022012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Haggar FA and Boushey RP: Colorectal
cancer epidemiology: Incidence, mortality, survival and risk
factors. Clin Colon Rectal Surg. 22:191–197. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Blair V, Kahokehr A and Sammour T: Cancer
in Māori: Lessons from prostate, colorectal and gastric cancer and
progress in hereditary stomach cancer in New Zealand. ANZ J Surg.
83:42–48. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lazarova DL, Chiaro C, Wong T, Drago E,
Rainey A, O'Malley S and Bordonaro M: CBP Activity Mediates Effects
of the histone deacetylase inhibitor butyrate on WNT activity and
apoptosis in colon cancer cells. J Cancer. 4:481–490. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chong KY, Hsu CJ, Hung TH, Hu HS, Huang
TT, Wang TH, Wang C, Chen CM, Choo KB and Tseng CP: Wnt pathway
activation and ABCB1 expression account for attenuation of
proteasome inhibitor-mediated apoptosis in multidrug-resistant
cancer cells. Cancer Biol Ther. 16:149–159. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jang GB, Hong IS, Kim RJ, Lee SY, Park SJ,
Lee ES, Park JH, Yun CH, Chung JU, Lee KJ, et al: Wnt/beta-catenin
small-molecule inhibitor CWP232228 preferentially inhibits the
growth of breast cancer stem-like cells. Cancer Res. 75:1691–1702.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hall CL, Zhang H, Baile S, Ljungman M,
Kuhstoss S and Keller ET: p21CIP-1/WAF-1 induction is required to
inhibit prostate cancer growth elicited by deficient expression of
the Wnt inhibitor Dickkopf-1. Cancer research. 70:9916–9926. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lin C, Lu W, Zhang W, Londoño-Joshi AI,
Buchsbaum DJ, Bu G and Li Y: The C-terminal region Mesd peptide
mimics full-length Mesd and acts as an inhibitor of Wnt/β-catenin
signaling in cancer cells. PLoS One. 8:e581022013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sack U, Walther W, Scudiero D, Selby M,
Aumann J, Lemos C, Fichtner I, Schlag PM, Shoemaker RH and Stein U:
S100A4-induced cell motility and metastasis is restricted by the
Wnt/β-catenin pathway inhibitor calcimycin in colon cancer cells.
Mol Biol Cell. 22:3344–3354. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Seo HS, Ku JM, Choi HS, Woo JK, Jang BH,
Shin YC and Ko SG: Induction of caspase-dependent apoptosis by
apigenin by inhibiting STAT3 signaling in HER2-overexpressing
MDA-MB-453 breast cancer cells. Anticancer Res. 34:2869–2882.
2014.PubMed/NCBI
|
|
30
|
Wu DG, Yu P, Li JW, Jiang P, Sun J, Wang
HZ, Zhang LD, Wen MB and Bie P: Apigenin potentiates the growth
inhibitory effects by IKK-β-mediated NF-κB activation in pancreatic
cancer cells. Toxicol Lett. 224:157–164. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chunhua L, Donglan L, Xiuqiong F, Lihua Z,
Qin F, Yawei L, Liang Z, Ge W, Linlin J, Ping Z, et al: Apigenin
up-regulates transgelin and inhibits invasion and migration of
colorectal cancer through decreased phosphorylation of AKT. J Nutr
Biochem. 24:1766–1775. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang QR, Yao XQ, Wen G, Fan Q, Li YJ, Fu
XQ, Li CK and Sun XG: Apigenin suppresses the growth of colorectal
cancer xenografts via phosphorylation and up-regulated FADD
expression. Oncol Lett. 2:43–47. 2011.PubMed/NCBI
|