Introduction
Maintaining genetic stability is an important
process in all organisms; failure to protect cells from genotoxic
stress caused by DNA-damaging agents, including ionizing radiation,
ultraviolet radiation and reactive oxygen species, may cause tumour
formation (1). The DNA damage
response (DDR) signaling pathway is the primary response mechanism
against these stresses (1,2). DDR serves a crucial function in DNA
repair since it protects the integrity of the genome from genotoxic
agents by controlling cell-cycle checkpoints, resulting in
cell-cycle arrest or apoptosis in eukaryotic cells (3). The fate of the cell, survival or death,
upon DNA damage is determined following DNA repair, cell-cycle
checks and the expression of apoptotic proteins (4). In arrested cells, polymerases are
inactivated to inhibit DNA replication and transcription and are
reactivated once DNA repair is complete (5). In a non-repaired or checkpoint-bypassed
cell, however, cell death, via mechanisms including apoptosis,
necrosis or mitotic catastrophe, depends on the balance between
pro-survival and pro-apoptotic factors (5,6). Proteins
that induce cell death via checkpoint mechanisms are known as tumor
suppressors; however, overactive proliferative proteins known as
oncogenes may induce cancer formation (7).
Tumor protein p53 (hereafter p53) is a tumor
suppressor protein that is occasionally referred to as ‘the
guardian of the genome’ (8,9). Previous research has shown that p53 is a
primary component of the intracellular signaling response against
genotoxic stress and that multiple types of human cancer exhibit
inactivated p53 owing to inherited mutations (9). Under normal conditions, p53 is commonly
inactivated and degraded in a ubiquitin-dependent manner; however,
it is upregulated and stabilized under genetically unstable
conditions, such as DNA damage, to suppress the progression of
tumorigenesis (10). The primary
function of p53 upon DNA damage is to regulate the expression of
target genes associated with cell fate (11). A previous study revealed that zinc
finger protein 224 (ZNF224) functions as an oncogene in breast
cancer cells and downregulates expression of p53 and cyclin
dependent kinase 1A (CDK1A, also known as p21) upon DNA damage
(12). Although the mRNA expression
levels of ZNF224 were unaffected by the DDR, ZNF224 protein
levels showed a gradual decrease (12). These results indicated that
post-translational modification to ZNF224 or the formation of a
complex with another regulatory protein may mediate its inhibition
upon DNA damage.
The human genome encodes ~30 mediator complex
subunit (MED) proteins that control intracellular signaling to the
nucleus (13). The major function of
these mediators is to transduce signals from transcription factors
to RNA polymerase II to aid the regulation of transcription; it has
recently been determined that mediators serve a function as
co-activators or co-repressors, depending on their interacting
proteins (14). MED28 has been
demonstrated to promote the migration and proliferation of breast
cancer cells through interactions with the mitogen-activated
protein kinase kinase-1 (MEK-1) signaling pathway (15–17).
However, the target ligands of MED28 have not been validated,
either in vitro or in vivo. To improve understanding
of the roles of MED28 in proliferating cancer cells, the present
study identified its interacting protein as ZNF224 using proteomic
and structural approaches. In addition, a cell-based fluorescence
assay revealed their functional co-localization in breast cancer
cells. The present study also investigated the role of MED28
targeting ZNF224, and the overexpression of MED28 demonstrated a
significant difference in terms of differentiation compared to the
control sample by colony forming experiment in breast cancer
cells.
Materials and methods
Cell culture
Cells from the 293 and human adenoma breast cancer
MCF-7 cell lines were cultured in Dulbecco's modified Eagle's
medium (DMEM; Hyclone; GE Healthcare Life Sciences, Logan, UT, USA)
supplemented with 10% fetal bovine serum (FBS; Hyclone; GE
Healthcare Life Sciences) and 1% penicillin/streptomycin (Hyclone;
GE Healthcare Life Sciences).
Plasmids
Constructs expressing Flag-ZNF224 (12), Myc-MED28 (18), and 6× His-MED28 (18) were previously verified. HA-tagged
N-terminus of ZNF224 construct was kindly provided by Dr. Constanzo
(University of Naples Federico II, Naples, Italy). Flag-ZNF224 and
Myc-MED28 constructs were subcloned into pE green fluorescence
protein (GFP) and pmCherry vectors, respectively. The following
primers were used to synthesize the glutathione S-transferase
(GST)-MED28 deletion mutants: Full-length forward,
5′-CGCGGATCCATGGCGGCTC-3′ and reverse,
5′-CCGCTCGAGTCACGTTGGCTTCAG-3′; amino acid (aa) 1–71 forward,
5′-CGCGGATCCATGGCGGCTC-3′ and reverse,
5′-CCGCTCGAGTCATTCCTGATCGGTGCC-3′; aa 1–150 forward,
5′-CGCGGATCCATGGCGGCTC-3′ and reverse,
5′-CCGCTCGAGTCAGTGCTGCACGTTGAT-3′; aa 43–176 forward,
5′-GGAATTCCATATGACTTTGGTGGACGAG-3′ and reverse,
5′-CCGCTCGAGTCACTTCAGAGGTGC-3′; aa 61–176 forward,
5′-GGAATTCCATATGAGTCAGGACTATGTCAATGG-3′ and reverse,
5′-CCGCTCGAGTCACTTCAGAGGTGC-3′; aa 72–176 (MED domain) forward,
5′-CGCGGATCCAAGAAATTCGAACCG-3′ and reverse,
5′-CCGCTCGAGTCACTTCAGAGGTGCA-3′. The above sequences were amplified
by PCR using AccuPower PCR premix (Bioneer Corporation, Daejeon,
Korea) and inserted into the pGEX-4T3 vector using the BamHI
and XhoI restriction sites (GE Healthcare Life Sciences,
Little Chalfont, UK) according to the manufacturer's protocol. The
thermocycler conditions were as follows: Initial denaturation at
95°C for 5 min, followed by 30 cycles of denaturation at 95°C,
annealing at melting temperature (Tm) 55–58°C and extension at 72°C
for 1 min each. ZNF224 and MED28 cDNA clones were
subcloned into pBiCF-VN173-Flag and pBiFC-VC155-HA vectors,
respectively (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany).
Mammalian cell transfection
Plasmid DNA constructs 1 µg DNA were transfected
into cells using a mixture of 150 mM NaCl and polyethylenimine
(Polysciences, Inc., Warrington, PA, USA) according to the
manufacturer's protocol, and cells were plated in a humidified
chamber atmosphere comprising 95% air and 5% CO2 at 37°C
for 24 h prior to transfection. All subsequent experiments were
performed 24 h after transfection.
Colony-forming assay
MCF-7 cells (3×103 cell/well) were
transfected with Myc-MED28 and/or Flag-ZNF224, and seeded on 6-well
plates and maintained in a humidified chamber atmosphere comprising
95% air and 5% CO2 at 37°C for 24 h. After transfection,
10 µM CPT (Camptothecin; Sigma-Aldrich; Merck KGaA) which dissolved
in dimethyl sulfoxide (DMSO) was treated for 2 weeks. Following CPT
treatment, cells were fixed with 4% paraformaldehyde at 25°C for 10
min and stained using crystal violet solution (0.05% crystal
violet, 1% formaldehyde, 1% methanol and 1X PBS) at 25°C for 30
min. Stained cells were washed with water by dropping gently, and
air dried at room temperature. The number of colonies were
quantified using the Nikon digital photo camera COOLPIX P310 and
ImageJ program (version 1.6.0; National Institutes of Health,
Bethesda, MD, USA).
Western blotting and
immunoprecipitation
The 293 cells were co-transfected with Flag-ZNFF224,
HA-ZNF224-N, Flag-ZNF255 and Myc-MED28 vectors and maintained in a
humidified chamber atmosphere comprising 95% air and 5%
CO2 at 37°C for 24 h. For immunoprecipitation, cells
were harvested and lysed with lysis buffer (50 mM HEPES, 150 mM
NaCl, 10% glycerol, 1% Triton X-100, 10 mM NaF, 1 mM
NaOV3, 1 mM PMSF, 1 mM EDTA) containing a protease
inhibitor cocktail (Thermo Fisher Scientific, Inc., Waltham, MA,
USA) at 4°C for 30 min, and centrifuged at 25,000 × g for 10 min at
4°C. Purified 1 µg anti-Flag (cat. no. F3165; Sigma-Aldrich; Merck
KGaA) and 1 µg anti-Myc antibodies (cat no. sc-40; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) were incubated with 500 µg of
protein extracts at 4°C for 4 h. Then, protein A/G PLUS agarose
beads (cat. no. sc-2003; Santa Cruz Biotechnology, Inc.) were added
and rotated at 0.04 × g at 4°C for 2 h on Rotator (FINEPCR,
Gunpo-si, Korea). The beads were collected by centrifugation at
3,000 × g for 1 min, and washed three times in lysis buffer and
resuspended in sodium dodecyl sulphate polyacrylamide gel
electrophoresis (SDS-PAGE) sample buffer [120 mM Tris-HCl (pH 6.8),
20% glycerol, 4% SDS, 28.8 mM 2-mercaptoethanol, 0.01% bromophenol
blue]. The samples were boiled in SDS-PAGE sample buffer [120 mM
Tris-HCl (pH 6.8), 20% glycerol, 4% SDS, 28.8 mM 2-mercaptoethanol,
0.01% bromophenol blue] at 100°C for 10 min and separated by
SDS-PAGE on a 12.5% gel. For western blotting, the whole cell
lysates were quantified using a Bradford protein assay (Bio-Rad
Laboratories, Inc., Hercules, CA, USA) through SpectroStar (BMG
Labtech GmbH, Ortenberg, Germany). The whole cell lysates (30 µg)
and immunoprecipitation sample (30 µl) were subjected to SDS-PAGE,
and transferred to a polyvinyldene fluoride membrane (EMD
Millipore, Billerica, MA, USA). Each transferred membrane was
blocked with TBS with Tween-20 (TBS-T) buffer [20 mM Tris-HCl (pH
7.4), 150 mM NaCl, 0.2% Tween-20] containing 5% skim milk at 25°C
for 1 h, and then incubated with the appropriate primary antibodies
diluted in 1% bovine serum albumin (BSA; Bovogen, Victoria,
Australia) in TBS-T buffer at 4°C for overnight. The following
primary antibodies were used: M2 anti-Flag (cat. no. F3165;
1:3,000; Sigma-Aldrich; Merck KGaA), anti-Myc (cat. no. sc-40;
1:1,000; Santa Cruz Biotechnology, Inc.), anti-HA (cat. no.
sc-57592; 1:1,000; Santa Cruz Biotechnology, Inc.), anti-ZNF224
(cat. no. ab168669; 1:1,000; Abcam, Cambridge, UK), anti-MED28
(1:10,000; laboratory-made), and anti-alpha-tubulin (1:20,000;
laboratory-made). The membranes were washed three times in TBS-T
buffer, and subjected to incubation with horseradish
peroxidase-conjugated goat anti-mouse immunoglobulin G as a
secondary antibody (cat. no. 31430; 1:25,000; Thermo Fisher
Scientific, Inc.) at 25°C for 1 h. An enhanced chemiluminescence
detection system (Santa Cruz Biotechnology, Inc.) was used for
detection according to the manufacturer's protocol, and the
membranes were exposed to the X-ray film (Fujifilm, Tokyo, Japan)
in a dark room, and then developed by Vivid X-ray developer and
Fixer solution (Duksan, Seoul, Korea) according to the
manufacturer's protocol.
GST fusion protein purification and
pull-down assays
BL-21 cells expressing the recombinant protein of
GST-tagged MED28 full length and deletion mutants were lysed in a
lysis buffer [10 mM Tris-HCl (pH 8.0), 150 mM NaCl, 1 mM EDTA, 200
mM PMSF, 5 mM DTT, 1% Triton X-100, 100 µg/ml lysozyme and PIC] at
4°C for 15 min. Cell lysates expressing GST-tagged proteins were
purified using Glutathione Separose™ 4B (GE Healthcare
Bio-Sciences, Pittsburgh, PA, USA) at 4°C for 2 h according to the
manufacturer's protocol. Purified 10 µg GST-tagged proteins were
incubated with Flag-ZNF224 transfected 293 cells lysate at 4°C for
1 h. The beads were collected by centrifugation at 3,000 × g for 1
min, and extensively washed three times with cell lysis buffer (50
mM HEPES, 150 mM NaCl, 10% glycerol, 1% Triton X-100, 10 mM NaF, 1
mM NaOV3, 1 mM PMSF, 1 mM EDTA) containing a protease
inhibitor cocktail (Thermo Fisher Scientific, Inc.). The bound
proteins were eluted by boiling in the SDS-PAGE sample buffer [120
mM Tris-HCl (pH 6.8), 20% glycerol, 4% SDS, 28.8 mM
2-mercaptoethanol, 0.01% bromophenol blue] at 100°C for 10 min, and
30 µg of whole cell lysate and 30 µl of samples were subjected to
SDS-PAGE and western blotting analysis as described above with an
M2 anti-Flag (cat. no. F3165; 1:3,000; Sigma-Aldrich; Merck KGaA)
antibody.
Matrix assisted laser desorption
ionization-time of flight mass spectrometry (MALDI-TOF MS) and
liquid chromatography-mass spectrometry (LC-MS/MS) analysis
MED28 overexpressed cells were lysed with lysis
buffer (50 mM HEPES, 150 mM NaCl, 10% glycerol, 1% Triton X-100, 10
mM NaF, 1 mM NaOV3, 1 mM PMSF, 1 mM EDTA) at 4°C for 30
min, and precipitated with an anti-MED28 antibody and protein A/G
PLUS-agarose (Santa Cruz Biotechnology, Inc.). The precipitates
were then boiled in SDS-PAGE sample buffer at 100°C for 10 min and
subjected to SDS-PAGE. Bands of interest on the SDS-PAGE gel were
in-gel digested with trypsin (Promega Corporation, Madison, WI,
USA) for 4°C for 30 min. The peptides were loaded on an Agilent
1100 Series nano-LC and LTQ-mass spectrometer (Thermo Electron;
Thermo Fisher Scientific, Inc.) for LC-MS/MS analysis. For LC
separation, 0.1% formic acid in deionized water and 0.1% formic
acid in acetonitrile were used for mobile phase A and B,
respectively. Mass spectra were acquired on a full mass scan
(400–1,800 m/z) by MS/MS and LTQ. The database search criteria were
as follows: taxonomy, Homo sapiens; fixed modification;
carboxyamidomethylated (+57) at cysteine residues; variable
modification, oxidized (+16) at methionine residues; maximum
allowed missed cleavage, 1; and MS tolerance, 100 ppm. Common
contaminants including trypsin and keratin were excluded.
Analysis of ZNF224 protein
stability
MCF-7 cells were transfected with the 1 µg Myc-empty
vector and 1 µg Myc-MED28 maintained in a humidified chamber
atmosphere comprising 95% air and 5% CO2 at 37°C for 24
h. Cells are treated with 100 µg cycloheximide for 24 h
post-transfection. Cells were harvested at 0, 1, 2, 4 and 8 h
following cycloheximide treatment, and the levels of ZNF224, MED28
and α-tubulin were determined by western blotting of total cell
lysates as described above.
Fluorescence detection
For the fluorescence assay, MCF-7 cells were
co-transfected with pEGFP-ZNF224 (0.5 µg) and pmCherry-MED28 (0.5
µg), and seeded (2×105 cells/well) onto sterile
coverslips in 12-well plates the day prior to transfection. MCF-7
cells were washed three times with 1× PBS and fixed with 4%
paraformaldehyde (cat. no. CNP015-1000; CellNest, Gyeonggi-do,
Korea) at 25°C for 10 min. The coverslips were mounted on glass
slides using mounting solution (Biomeda Corp., Foster City, CA,
USA) and images were captured under a fluorescence confocal
microscope under a ×60 magnification (Carl Zeiss AG, Oberkochen,
Germany).
Bimolecular fluorescence
complementation (BiFC) assay
MCF-7 cells were co-transfected with 1 µg
pBiFC-VN173-ZNF224-Flagand and 1 µg pBiFC-VC155-MED28-HA plasmids,
and seeded (3×105 cells/well) onto 6-well plates. Then,
the cells were maintained in a humidified chamber atmosphere
comprising 95% air and 5% CO2 at 37°C for 24 h.
BiFC-induced GFP images were captured under a fluorescence
microscope at a ×20 magnification (Nikon Corporation, Tokyo,
Japan).
Surface plasmon resonance (SPR)
assay
SPR experiments were performed using the SR7500DC
Dual Channel system (Reichert, Inc., Depew, NY, USA) according to
the manufacturer's protocol. Flag-ZNF224 proteins (50 µg) were
immobilized on CMDH chips (Reichert Technologies) according to the
manufacturer's protocol. For the Flag-ZNF224-6xHis-MED28 binding
analysis, each concentration (0, 1.25, 2.5 and 5 µM) of 6X
His-MED28 protein in 1X PBS buffer was injected as an analyte. BSA
was used as a control. The scrubber2 program (version 2.0.0004;
BioLogic Software Pty., Ltd., Campbell, ACT, Australia) was used to
evaluate the dissociation constant (KD).
Statistical analysis
The bands from all western blot analysis were
adjusted by ImageJ program (version 1.6.0; National Institutes of
Health) for statistical analysis. All experiments were performed at
least three times and analyses were performed with Microsoft Excel
(Microsoft Corporation, Redmond, WA, USA) and GraphPad Prism 5.0
software (GraphPad Software, Inc., La Jolla, CA, USA). When two
groups were compared, statistical differences were assessed with an
unpaired two-tailed Student's t-test. P<0.05 was considered to
indicate a statistically significant difference.
Results
MED28 is a binding partner of
ZNF224
It was previously revealed that ZNF224 regulates the
transcription of TP53 and p21 upon DNA damage
(12); however, the exact molecular
mechanism of this interaction is, to the best of our knowledge,
unknown. It was hypothesized that the physical interaction of
ZNF224 with its binding partner protein in the nucleus affected the
expression of downstream genes. Therefore, a pulldown assay was
performed using 293 cell lysates in the presence of purified GST or
GST-MED28. Mass analysis using MALDI-TOF identified ZNF224 as a
binding candidate of MED28 (data not shown). Sequence analysis
revealed that ZNF224 has a canonical Krüppel-associated box (KRAB)
domain and 19 tandem-repeat C2H2 zinc finger
domains; ZNF255, its alternative transcription product, lacks the
KRAB domain (19). In addition, MED28
possesses a proline-rich (aa 13–39) and MED domain (Fig. 1A) (20).
To determine whether MED28 is an essential binding partner of
ZNF224, a co-immunoprecipitation assay was performed following
transfection of Myc-MED28 with Flag-ZNF224 or Flag-ZNF255. A GST
pull-down assay using anti-Myc or anti-Flag antibodies revealed
that ZNF224, but not ZNF255, specifically associated with MED28,
indicating that the KRAB domain of ZNF224 may bind to MED28
(Fig. 1B and C). To investigate this
further, co-immunoprecipitation was performed following
transfection of Myc-MED28 with the HA-ZNF224 N-terminal KRAB domain
(aa 1–67), Flag-ZNF224 or Flag-ZNF255. This result confirmed that
the KRAB domain, which is found at the N-terminal of ZNF224,
specifically interacts with MED28 (Fig.
1D). To identify the ZNF224-interacting domain of MED28,
GST-fused MED28-deletion mutants were designed, designated aa 1–71,
aa 1–151, aa 43–176, aa 61–176, and aa 72–176 (identified as the
MED domain), and examined their interactions with ZNF224 using a
GST pull-down assay (Fig. 1E). The
results revealed that the MED domain of MED28 is critical for
association with ZNF224 (Fig. 1E).
Next, an SPR assay was performed to evaluate the binding affinity
of ZNF224 with MED28. ZNF224 bound to MED28 with affinity
(KD=3.4×10−6 M), but not to BSA used as
negative control. Collectively, these observations revealed that
MED28 is a ZNF224-interacting protein through specific binding
regions, and that ZNF224/MED28 interactions may be associated with
the regulation of downstream genes.
 | Figure 1.MED28 is a ZNF224-binding protein. (A)
Protein structure of ZNF224/ZNF255 and mediator complex subunit 28
(MED28). ZNF224 has conserved KRAB (aa 8–67) and zinc finger (aa
190–641) domains. MED28 has a PRD (aa 13–39) and MED domain (aa
72–178), which is shared between the paralogous mediator complex
subunit proteins. Myc-MED28 was transfected into 293 cells in the
presence of Flag-ZNF224 or Flag-ZNF255 for 24 h. Cell extracts were
subjected to an immunoprecipitation assay using (B) anti-Myc or (C)
anti-Flag antibodies; samples were separated by SDS-PAGE with WCL.
Immunoblotting was performed with the indicated antibodies, and
γ-tubulin was used as a loading control. (D) Myc-MED28 was
transfected into 293 cells in the presence of Flag-ZNF224,
Flag-ZNF255, or HA-ZNF224-N (aa 1–67). Cell lysates were
precipitated using an anti-Myc antibody and immunoblotted with the
indicated antibodies. (E) Ponceau-S stain showing purified GST
(lane 2), GST-MED28 full length (lane 3), aa 1–71 (lane 4), aa
1–151 (lane 5), aa 43–176 (lane 6), aa 61–176 (lane 7), and aa
72–176 (MED domain, lane 8). Cell lysates from
Flag-ZNF224-transfected 293 cells were incubated with GST fusion
proteins for 4 h, and samples were analyzed using SDS-PAGE and
immunoblotted with an anti-Flag antibody. MED28, mediator complex
subunit 28; ZNF224, zinc finger protein 224; aa, amino acid; KRAB,
Krüppel-associated box; PRD, proline rich domain; HA,
hemagglutinin; Myc, Myc proto-oncogene protein; WCL, whole cell
lysates; GST, glutathione S-transferase. |
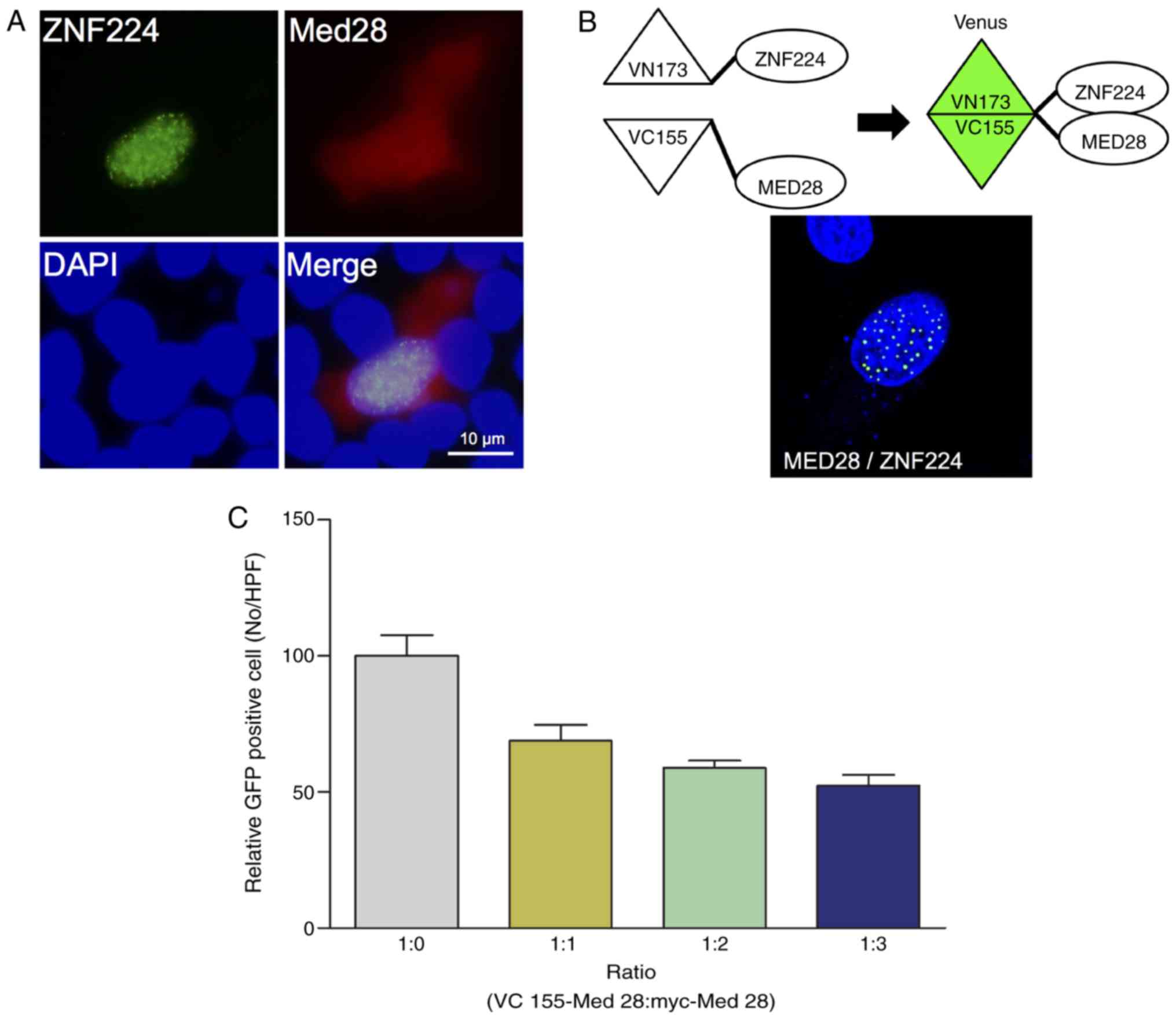
Nucleus-specific interactions of
ZNF224 with MED28
Previously, it was shown that ZNF224 downregulates
the transcription of p53 and p21, and that MED28 is
localized to the cytosol and nucleus (12,18). In
addition, it is known that ZNF224 primarily localizes to the
nucleus and forms a punctate pattern (12). Therefore, we hypothesized that the
interaction between ZNF224 and MED28 may occur in the nucleus. To
verify this hypothesis, the cellular co-localization of ZNF224 and
MED28 was examined using fluorescence microscopy. ZNF224
co-localized with MED28 in the nucleus (Fig. 2A). To confirm the nuclear interaction
of ZNF224 with MED28 further, a BiFC assay. ZNF224 was annealed to
the N-terminus of the Venus fluorescent protein (VN173-ZNF224) and
MED28 to the C-terminus of Venus (VC155-MED28) and co-transfected
them into HeLa cells. As shown in Fig.
2B, the BiFC signal was detected only in the nucleus of the
cells, confirming that the interaction of ZNF224 with MED28
primarily occurs in nucleus (Fig.
2A). To confirm the nuclear-specific interaction further, the
expression level of Myc-MED28 was increased in a dose-dependent
manner to act as a competitor for VC155-MED28, the number of GFP
positive cells was evaluated. The results showed that the number of
GFP-positive cells decreased as the concentration of Myc-MED28
increased (Fig. 2C), indicating that
the nuclear interaction between ZNF224 and MED28 is specific.
MED28 prevents the degradation of
ZNF224 upon DNA damage
Multiple KRAB-ZFPs function as transcriptional
repressors during DDR in cancer cells (21). In a previous study, it was found that
ZNF224, a KRAP-ZFP, served a function as a transcriptional
repressor upon DNA damage and that the overexpression of ZNF224
results in tumor formation (12). In
addition, it was observed that ZNF224 was degraded in a
time-dependent manner upon DNA damage (12). Previous studies have revealed that
MEDs possess functions as co-repressors or co-activators of
transcription in eukaryotic cells (14,22).
However, it is not clear whether MED28 functions as a co-repressor
or co-activator of ZNF224, or how MED28 affects ZNF224 in DDR. To
address this question, the degradation of ZNF224 was examined in
response to DNA damage in the presence or absence of MED28.
Notably, MED28 expression increased the stability of endogenous and
exogenous ZNF224 upon DNA damage (Fig.
3). Because the level of ZNF224 translation could affect
protein stability, ZNF224 protein stability in the presence or
absence of overexpressed MED28 was examined following treatment
with cycloheximide. The vector inducing overexpression of MED28
maintained higher levels of ZNF224 expression than the empty vector
(Fig. 3B). These results indicated
that ZNF224 is degraded by DDR (although the enzyme that regulates
the stability of ZNF224 has not been identified), and that MED28
may inhibit the turnover of ZNF224 by forming a MED28/ZNF224
complex or inhibiting the binding of DDR-mediated proteases.
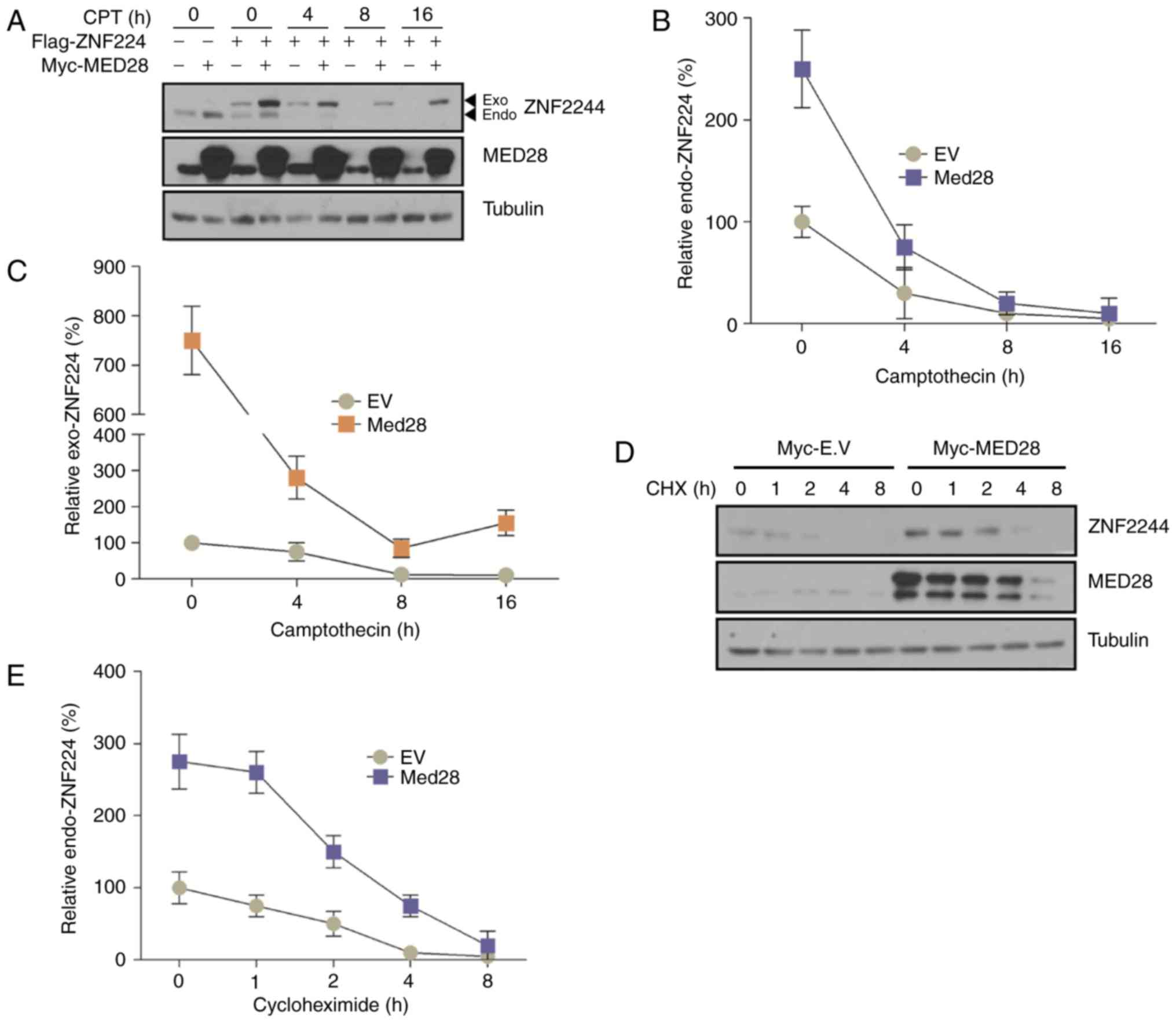 | Figure 3.ZNF224 is stabilized by MED28 upon
DNA damage. (A-C) Myc-MED28 and/or Flag-ZNF224 co-transfected MCF-7
cells were incubated with 100 µM CPT for the indicated time periods
prior to harvesting. Endogenous and exogenous ZNF224 and MED28 were
detected using an anti-ZNF224 and anti-MED28 antibodies,
respectively; α-tubulin was used as the loading control (n=3,
P<0.03 vs. EV control). (D and E) MCF-7 cells were transfected
with EV or Myc-MED28. After 24 h, cells were treated with
cycloheximide and harvested at the indicated times. Cell extracts
were examined by immunoblotting with the indicated antibodies (n=3,
P<0.001). MED28, mediator complex subunit 28; ZNF224, zinc
finger protein 224; EV, empty vector; CPT, camptothecin; CHX,
cycloheximide; Exo, exogenous; Endo, endogenous. |
MED28 increases cell proliferation
upon DNA damage
Cell-cycle progression is accompanied by checkpoint
phases (23). This phenomenon
generally occurs in proliferating cells through the activation of
checkpoint proteins to prevent genome instability; thereafter, the
cell will typically resume normal proliferation (24). Since p53 and p21 are activated during
DNA repair (25), it was hypothesized
that MED28 may disrupt DNA repair through ZNF224-mediated
repression of p53 and p21, leading to increased cell proliferation.
To quantify the function of MED28 in the recovery of cell
proliferation following DNA damage, the colony-forming ability of
MCF-7 cells was assessed following treatment with camptothecin
(CPT), a DNA-damaging agent known to be an inhibitor of
topoisomerase I (26). The
colony-forming assay revealed that co-transfection with MED28 and
ZNF224 considerably increased the proliferation rate of MCF-7 cells
(Fig. 4). This result indicated that
the interaction of MED28 with ZNF224 induced abnormal cell
proliferation following DNA damage.
Discussion
ZNF224 serves a function as a transcriptional
repressor in the regulation of gene expression through interactions
with multiple co-factors, including KRAB-associated protein-1,
protein arginine methyltransferase 5, DEP domain-containing 1 and
Wilms' tumor gene 1 (19,27–31). To
the best of our knowledge, previous studies investigating the
function of MED28 have focused only on phenotypes or relative
intracellular signaling in mammalian cells (17,20). In
the present study, MED28 was screened as a binding partner of
ZNF224. In addition, the present study revealed that MED28 was
associated with intracellular signals through a physical
interaction with ZNF224 upon DNA damage.
The KRAB domain is a core transcriptional repressor
domain in KRAB-ZFPs and mediates interactions with co-repressors
(21). Interaction-domain mapping
analysis using multiple deletion mutants revealed that the KRAB
domain of ZNF224 is critical for its association with MED28
(Fig. 1). A previous study
demonstrated that the zinc-finger domains of KRAB-ZFPs are critical
for binding to DNA at the promoter of the target gene, whereas the
remaining zinc-finger domains are required for protein-protein
interactions (32). However, the
present study indicated that the KRAB domain may also be required
for substrate binding.
Interaction analysis using multiple MED28-deletion
mutants derived from Escherichia coli also revealed
interactions between MED28 and ZNF224. In addition, the deletion
mutant containing the MED domain of MED28 exhibited an increased
binding affinity to ZNF224 compared with that of any other deletion
mutant. This result indicated that the post-translational
modification of MED28 is not required for interaction with ZNF224
and that the C-terminal MED domain of MED28 is essential for
mediating the association with ZNF224. In addition, co-localization
of ZNF224 with MED28 was observed in the nucleus. Therefore, the
results of the present study indicated that ZNF224 interacts with
MED28 in the nucleus to repress the transcription of target
genes.
In vitro colony-forming assays can be used to
predict the outcome of cancer therapies, and are often used to test
anticancer drugs (33). Therefore,
the present study assessed the association between ZNF224 and MED28
under chemo-sensitive conditions using a DNA-damaging agent in
cancer cells; their expression increased the incidence of colony
formation compared with the non-expressing group. This result
indicated that the interaction between ZNF224 and MED28 has
synergetic effects for cancer cell survival and proliferation,
against the cell-cycle checkpoint system upon DNA damage. ZNF224 is
highly expressed in breast and bladder cancer (12,30,34). In
addition, MED28 is abundantly expressed in breast, colon, and
prostate cancer (35). It was
determined that the interaction between MED28 and ZNF224
downregulates tumour suppressors, including p53 and p21, resulting
in increased cancer cell proliferation compared with normal
conditions. Further study is required to identify how the
interaction between ZNF224 and MED28 is regulated in cancer and how
the development of an inhibitor to suppress the interaction between
ZNF224 and MED28 may contribute to effective cancer treatments.
Collectively, the data suggests that the novel protein MED28
increases the proliferative ability of breast cancer cells by the
regulation of the ZNF224 protein against DNA damage responses.
Acknowledgements
The present study was supported by the Basic
Research Program through the National Research Foundation of Korea
funded by the Ministry of Education, Science and Technology (grant
no. NRF-2012R1A1A2040602).
References
|
1
|
Lord CJ and Ashworth A: The DNA damage
response and cancer therapy. Nature. 481:287–294. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jackson SP and Bartek J: The DNA-damage
response in human biology and disease. Nature. 461:1071–1078. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Weinert T: DNA damage and checkpoint
pathways: Molecular anatomy and interactions with repair. Cell.
94:555–558. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhou BB and Elledge SJ: The DNA damage
response: Putting checkpoints in perspective. Nature. 408:433–439.
2000. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Roos WP, Thomas AD and Kaina B: DNA damage
and the balance between survival and death in cancer biology. Nat
Rev Cancer. 16:20–33. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Vitale I, Galluzzi L, Castedo M and
Kroemer G: Mitotic catastrophe: A mechanism for avoiding genomic
instability. Nat Rev Mol Cell Biol. 12:385–392. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Freed-Pastor W and Prives C: Targeting
mutant p53 through the mevalonate pathway. Nat Cell Biol.
18:1122–1124. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lane DP: Cancer. p53, guardian of the
genome. Nature. 358:15–16. 1992. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Stracquadanio G, Wang X, Wallace MD,
Grawenda AM, Zhang P, Hewitt J, Zeron-Medina J, Castro-Giner F,
Tomlinson IP, Goding CR, et al: The importance of p53 pathway
genetics in inherited and somatic cancer genomes. Nat Rev Cancer.
16:251–265. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Muller PA and Vousden KH: Mutant p53 in
cancer: New functions and therapeutic opportunities. Cancer Cell.
25:304–317. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Menendez D, Inga A and Resnick MA: The
expanding universe of p53 targets. Nat Rev Cancer. 9:724–737. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cho JG, Park S, Lim CH, Kim HS, Song SY,
Roh TY, Sung JH, Suh W, Ham SJ, Lim KH and Park SG: ZNF224, Krüppel
like zinc finger protein, induces cell growth and
apoptosis-resistance by down-regulation of p21 and p53 via
miR-663a. Oncotarget. 7:31177–31190. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Malik S and Roeder RG: The metazoan
mediator co-activator complex as an integrative hub for
transcriptional regulation. Nat Rev Genet. 11:761–772. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Allen BL and Taatjes DJ: The mediator
complex: A central integrator of transcription. Nat Rev Mol Cell
Biol. 16:155–166. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lu M, Sartippour MR, Zhang L, Norris AJ
and Brooks MN: Targeted inhibition of EG-1 blocks breast tumor
growth. Cancer Biol Ther. 6:936–941. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lee MF, Pan MH, Chiou YS, Cheng AC and
Huang H: Resveratrol modulates MED28 (Magicin/EG-1) expression and
inhibits epidermal growth factor (EGF)-induced migration in
MDA-MB-231 human breast cancer cells. J Agric Food Chem.
59:11853–11861. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Huang CY, Chou YH, Hsieh NT, Chen HH and
Lee MF: MED28 regulates MEK1-dependent cellular migration in human
breast cancer cells. J Cell Physiol. 227:3820–3827. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yu MA, Cho JG, Kim KI, Jo YJ, Sung JH,
Yang HB and Park SG: Generation of med28 specific monoclonal
antibodies. Monoclon Antib Immunodiagn Immunother. 34:30–35. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Florio F, Cesaro E, Montano G, Izzo P,
Miles C and Costanzo P: Biochemical and functional interaction
between ZNF224 and ZNF255, two members of the Kruppel-like
zinc-finger protein family and WT1 protein isoforms. Hum Mol Genet.
19:3544–3556. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lu M, Zhang L, Maul RS, Sartippour MR,
Norris A, Whitelegge J, Rao JY and Brooks MN: The novel gene EG-1
stimulates cellular proliferation. Cancer Res. 65:6159–6166. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lupo A, Cesaro E, Montano G, Zurlo D, Izzo
P and Costanzo P: KRAB-Zinc finger proteins: A repressor family
displaying multiple biological functions. Curr Genomics.
14:268–278. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Myers LC and Kornberg RD: Mediator of
transcriptional regulation. Annu Rev Biochem. 69:729–749. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Otto T and Sicinski P: Cell cycle proteins
as promising targets in cancer therapy. Nat Rev Cancer. 17:93–115.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kastan MB and Bartek J: Cell-cycle
checkpoints and cancer. Nature. 432:316–323. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sperka T, Wang J and Rudolph KL: DNA
damage checkpoints in stem cells, ageing and cancer. Nat Rev Mol
Cell Biol. 13:579–590. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gokduman K: Strategies targeting DNA
topoisomerase i in cancer chemotherapy: Camptothecins, nanocarriers
for camptothecins, organic non-camptothecin compounds and metal
complexes. Curr Drug Targets. 17:1928–1939. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Medugno L, Florio F, De Cegli R, Grosso M,
Lupo A, Costanzo P and Izzo P: The Krüppel-like zinc-finger protein
ZNF224 represses aldolase A gene transcription by interacting with
the KAP-1 co-repressor protein. Gene. 359:35–43. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cesaro E, De Cegli R, Medugno L, Florio F,
Grosso M, Lupo A, Izzo P and Costanzo P: The Kruppel-like zinc
finger protein ZNF224 recruits the arginine methyltransferase PRMT5
on the transcriptional repressor complex of the aldolase A gene. J
Biol Chem. 284:32321–32330. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Montano G, Ullmark T, Jernmark-Nilsson H,
Sodaro G, Drott K, Costanzo P, Vidovic K and Gullberg U: The
hematopoietic tumor suppressor interferon regulatory factor 8
(IRF8) is upregulated by the antimetabolite cytarabine in leukemic
cells involving the zinc finger protein ZNF224, acting as a
cofactor of the Wilms' tumor gene 1 (WT1) protein. Leuk Res.
40:60–67. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Harada Y, Kanehira M, Fujisawa Y, Takata
R, Shuin T, Miki T, Fujioka T, Nakamura Y and Katagiri T:
Cell-permeable peptide DEPDC1-ZNF224 interferes with
transcriptional repression and oncogenicity in bladder cancer
cells. Cancer Res. 70:5829–5839. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lee TH, Lwu S, Kim J and Pelletier J:
Inhibition of Wilms tumor 1 transactivation by bone marrow zinc
finger 2, a novel transcriptional repressor. J Biol Chem.
277:44826–44837. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Moore M, Klug A and Choo Y: Improved DNA
binding specificity from polyzinc finger peptides by using strings
of two-finger units. Proc Natl Acad Sci USA. 98:pp. 1437–1441.
2001; View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Salmon SE: Human tumor colony assay and
chemosensitivity testing. Cancer Treat Rep. 68:117–125.
1984.PubMed/NCBI
|
|
34
|
Gur-Dedeoglu B, Konu O, Bozkurt B, Ergul
G, Seckin S and Yulug IG: Identification of endogenous reference
genes for qRT-PCR analysis in normal matched breast tumor tissues.
Oncol Res. 17:353–365. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang L, Maul RS, Rao J, Apple S, Seligson
D, Sartippour M, Rubio R and Brooks MN: Expression pattern of the
novel gene EG-1 in cancer. Clin Cancer Res. 10:3504–3508. 2004.
View Article : Google Scholar : PubMed/NCBI
|