Introduction
Wnt signaling is important in embryonic development,
tissue homeostasis regulation and tissue regeneration upon injury
(1), and the deregulation of Wnt
signaling is implicated in the initiation and development of
various types of cancer (2).
β-catenin is a key component of Wnt signaling, and its stability
and sub-cellular localization are tightly controlled. In the
absence of Wnt ligands, cytoplasmic β-catenin protein is constantly
degraded by a destruction complex composed of the scaffolding
protein AXIN, adenomatous polyposis coli (APC), casein kinase 1
(CK1) and glycogen synthase kinase 3β (GSK3β) (3). CK1 and GSK3β sequentially phosphorylate
β-catenin, promoting its degradation through an
ubiquitination-proteasome pathway (4). The binding of Wnt proteins to frizzled
(FZD) receptor and lipoprotein receptor-related protein 5 or 6
(LRP5/6) co-receptors trigger the phosphorylation of LRP5 or LRP6.
Phosphorylated LRP5 or LRP6 recruits AXIN to the plasma membrane
and inactivates the β-catenin destruction complex, leading to the
accumulation and nuclear translocation of β-catenin (5). In the nucleus, β-catenin forms a complex
with the lymphoid enhancer factor/T-cell factor and activates the
transcription of Wnt target genes, including AXIN2, cyclin D1
(CCND1), B-cell lymphoma 2 (BCL2), naked cuticle homolog 1 (NKD1),
and leucine-rich repeat-containing G-protein coupled receptor 5
(LGR5) (6).
Glioma is the most common form of neural malignancy
and contributes to ~70% of malignant primary brain tumors each year
(7). Despite therapeutic advances,
the prognosis of malignant gliomas remains poor and the majority of
patients eventually relapse (8).
Increasing evidence demonstrates that the aberrant activation of
Wnt signaling is crucial in gliomagenesis (9,10).
Hyperactive Wnt signaling in glioma is mainly attributed to the
overexpression of positive regulators and the silencing of negative
regulators. WNT1, evenness interrupted (EVI), dishevelled 3,
forkhead box M1 (FoxM1) and β-catenin, primary positive regulators
of Wnt signaling, show higher expression in glioma specimens,
compared with non-tumor brain tissues, and their expression levels
are associated positively with the degree of glioma (11–15).
Dickkopf (DKK1 and DKK3), antagonists of Wnt signaling, are
frequently silenced through promoter hypermethylation (16). Functional studies have demonstrated
that the ectopic expression of positive regulators of Wnt
signaling, including EVI (12) and
FoxM1 (14), promotes glioma
tumorigenesis, whereas DKK3 induces cell death in human malignant
glioma (17). Therefore, targeting
the Wnt signaling may be an effective therapeutic approach to treat
patients with glioma.
Propofol is widely used as an intravenous anesthetic
agent clinically due to its limited side effects and rapid recovery
(18). In addition to its anesthetic
advantages, accumulating clinical evidence demonstrates that cancer
patients receiving propofol-paravertebral anesthesia during cancer
surgery have reduced risk of recurrence and metastasis (19). In addition, studies have shown that
propofol exhibits potent antitumor activity in a variety of human
cancer cell lines, including lung cancer cells (20), colon carcinoma cells (21), cervical cancer cells (22), pancreatic cancer cells (23) and gastric cancer cells (24), through different molecular mechanisms.
However, the effect of propofol on Wnt signaling and glioma cell
growth remains to be elucidated. In present study, propofol was
identified an inhibitor of Wnt signaling, and it was shown that
propofol suppressed the expression of Wnt target genes and growth
of glioma cells in vitro and in vivo.
Materials and methods
Cell culture and reagents
The 293T cells and LN299 glioma cell line were
obtained from Cobioer Biosciences Co., Ltd. (Nanjing, China). The
LN299 glioma cell line and 293T cells were cultured in Dulbecco's
modified Eagle's medium (DMEM; Hyclone; GE Healthcare Life
Sciences, Logan, UT, USA) supplemented with 10% fetal bovine serum
(FBS; Hyclone; GE Healthcare Life Sciences), 100 U/ml penicillin
and 100 mg/l streptomycin. The cells were maintained in humidified
incubators at 37°C, 5% CO2. Propofol was purchased from
Sigma-Aldrich; Merck Millipore (Darmstadt, Germany) and dissolved
in either DMSO (Sigma-Aldrich; Merck Millipore) or soyabean oil
(Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA).
Luciferase reporter assay
The cells (3×104 cells/per well) were
plated into 24-well plates. The following day, the cells were
co-transfected with 300 ng of the TOPFlash luciferase construct and
500 ng of control, Wnt1 or Wnt3 plasmid. The pCMXβ gal plasmid (30
ng) was co-transfected as an internal control. Wnt3a-conditioned
medium (Wnt3a-CM) was prepared as previously described (25). At 24 h post-transfection, the cells
were treated the indicated concentrations (1–10 µg/ml) of propofol.
Following treatment with propofol for 48 h in humidified incubators
at 37°C in 5% CO2, the cells were harvested for
luciferase activity measurement. The luciferase assays were
performed using the Dual-Luciferase Reporter Assay system (Promega,
Madison, WI, USA) according to the manufacturer's protocol.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA was extracted from the 293T and LN299
cells using TRIzol (Qiagen, Inc., Valencia, CA, USA) following the
manufacturer's protocol. The RNA (1 µg per sample) was reverse
transcribed into cDNA using Reverse Transcriptase M-MLV (Takara
Biotechnology Co., Ltd., Dalian, China) according to the
manufacturer's protocol. RT-qPCR analysis was performed in an
Applied Biosystems 7900 Real-Time PCR system (Applied Biosystems;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) using the
QuantiTect SYBR Green PCR kit (Qiagen, Inc.) according to the
manufacturer's protocols. A total of 0.5 µM each primer was added
to a 20-µl RT-qPCR reaction mix containing 10 µl 2× QuantiTect SYBR
Green, 500 ng cDNA, 0.5 µl QuantiTect RT Mix (Qiagen, Inc.) and
RNase-free water. The thermocycling conditions were as follows:
95°C for 10 min, followed by 40 cycles of 95°C for 30 sec and 60°C
for 1 min. The average cycle quantitation (Cq), from triplicate
assays, was used for further calculations. The endogenous GAPDH
used as the normalization control. The 2−ΔΔCq Method
(26) was used to quantify the
relative levels of mRNA. The primer sequences were as follows:
AXIN2, forward: 5′-CCAACACCAGGCGGAACGAAG-3′ and reverse
5′-CGCCCAATAAGGAGTGTAAGGAC-3′; CCND1, forward
5′-AATGACCCCGCACGATTTC-3′ and reverse 5′-TCAGGTTCAGGCCTTGCAC-3′;
NKD1, forward 5′-GGGAAACTTCACTCCAAGCCG′ and reverse
5′-GTCTCCCGATCCACTCCTCG-3′; LGR5, forward
5′-CTCTTCCTCAAACCGTCTGC-3′ and reverse 5′-GATCGGAGGCTAAGCAACTG-3′;
BCL2, forward 5′-ATGTGTGTGGAGAGCGTCAA-3′ and reverse
5′-ACAGTTCCACAAAGGCATCC-3′; GAPDH, forward
5′-CCAGAACATCATCCCTGCCTCTACT-3′ and reverse
5′-GGTTTTTCTAGACGGCAGGTCAGGT-3′.
Western blot analysis
The cells were pelleted, washed twice in cold PBS
and resuspended in 100 µl lysis buffer (50 mM Tris-HCl, pH 7.4, 150
mM NaCl, 1 % Nonidet P-40, 1 mM EDTA, 0.25 % sodium deoxycholate
and 1 mM NaF) supplemented with 1X protease inhibitors (Roche
Diagnostics, Basel, Switzerland) on ice for 30 min. Lysate was
centrifuged for 5 min at 12,000 × g, 4°C to remove cell debris and
the supernatant was harvested. The protein concentration was
measured using the Pierce BCA protein assay kit (Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol. The
detailed protocol is described previously (27). An equal amount of protein per lane (50
µg) were resolved on 10% SDS-PAGE gels and transferred onto
nitrocellulose membranes (Bio-Rad Laboratories, Inc., Richmond, CA,
USA). The membranes were blocked with 5% nonfat milk in PBS
containing 0.1% Tween 20 and then incubated with primary antibodies
overnight at 4°C. Anti-tubulin antibody (cat. no. sc-101527) was
purchased from Santa Cruz Biotechnology, Inc.; anti-β-catenin
antibody (cat. no. 9857), anti-phosphorylated (p)LRP6 (Ser1490)
antibody (cat. no. 2568), anti-LRP6 antibody (cat. no. 2560),
anti-CCND antibody (cat. no. 2922) and anti-BCL2 antibody (cat no.
2872) were obtained from Cell Signaling Technology, Inc. (Beverly,
MA, USA). Anti-pLRP6 antibody was used at a dilution of 1:500,
anti-tubulin antibody was used at a dilution of 1:200, and other
antibodies were used at a dilution of 1:1,000. Following incubation
with the horseradish peroxidase-conjugated secondary antibody (cat.
no. HAF007 for mouse and cat. no. HAF008 for rabbit; R&D
Systems, Inc., Minneapolis, MN, USA) for 1 h at room temperature,
the proteins were visualized using ECL detection reagent (Amersham;
GE Healthcare Life Sciences).
Cell viability assays
The cells were seeded in 96-well plates at a density
of 2,000 per well. The following day, the cells were treated with
different concentrations (0, 1, 2, 5 and 10 µg/ml) of propofol for
48 h. Cell survival was determined using a Cell Counting Kit-8
(CCK-8) assay (Dojindo Molecular Technologies, Inc., Kumamoto,
Japan) according to the manufacturer's protocols. The absorbance at
450 nm was detected using a multilabel plate reader. Each
experiment was performed with three replicates per sample.
Analysis of cell proliferation and
apoptosis
Ki67 quantification was performed to determine the
effects of propofol treatment on cell proliferation. Briefly, the
cells were harvested 48 h following propofol treatment and stained
with anti-human Ki67 antibody (Abcam, Cambridge, MA) at a dilution
of 1:200 overnight at 4°C, followed by a FITC-conjugated mouse
anti-rabbit secondary antibody (Thermo Fisher Scientific, Inc.) at
a dilution of 1:2,000 for 1 h at room temperature prior to flow
cytometric analysis. Cell apoptosis was assessed using an Annexin
V-FITC apoptosis detection kit (BD Pharmingen, San Diego CA, USA)
according to the manufacturer's protocols. Briefly, the cells were
harvested 48 h following propofol treatment and washed twice with
ice-cold phosphate-buffered saline (PBS). The cells were stained
with FITC-labeled Annexin V and PI for 15 min in the dark, followed
by flow cytometric analysis. Data were analyzed using FlowJo
software version 10 (Tree Star, Inc., Ashland, OR, USA).
Xenograft animal model
Animal experiments were approved by the Ethics
Committee of Yangtze University (Jingzhou, China). Male,
5–6-week-old immune-deficient BALB/c nude mice were purchased from
Vital River Experimental Animal Center (Beijing, China). The mice
were housed in specific pathogen-free conditions in an environment
with a 12-h light-dark cycle, a temperature of 22±0.5°C and a
relative humidity of 50±1%. The mice had ad libitum access
to autoclaved rodent diet and deionized water treated by reverse
osmosis. LN299 cells (5×106) in 0.1 ml serum-free medium
were injected subcutaneously in the right flank of the nude mice.
When the tumors reached ~100 mm3, the mice were divided
randomly into two groups (n=6/group) according to tumor volumes and
body weights, and were treated with the vehicle (0.5 ml soybean
oil) or 0.5 ml propofol (50 mg/kg) by intraperitoneal injection for
20 days (once each day). The tumor sizes and body weights were
measured at indicated times. At the end of the experiments, the
mice were sacrificed and tumors were harvested and weighed,
followed by RT-qPCR and western blot analyses.
Statistical analysis
All data were analyzed using SPSS 17.0 software
(SPSS, Inc., Chicago, IL, USA). All values are presented as the
mean ± standard deviation. Student's t-test (unpaired) was used to
detect the statistical significance between two groups. One-way
analysis of variance followed by Dunnett's test was used to
determine the statistical differences when comparing more than two
groups. P<0.05 was considered to indicate a statistically
significant difference.
Results
Inhibition of Wnt signaling by
propofol
The present study first investigated the effect of
propofol on the Wnt signaling in 293T cells. Due to the low
endogenous Wnt signaling activity in 293T cells (28), the 293T cells were transfected with
the SuperTopFlash reporter plasmid together with Wnt1 or Wnt3
expression plasmids, following which they were treated with
increasing concentrations of propofol. As shown in Fig. 1A and B, Wnt1 or Wnt3 transfection
significantly increased the transcriptional activity of the
SuperTOPflash reporter, compared with the basal levels, and
propofol treatment suppressed the Wnt signaling induced by Wnt1 or
Wnt3 in a dose-dependent manner. In addition, it was found that
propofol dose-dependently inhibited the transcriptional activity of
the SuperTopFlash reporter induced with Wnt3A-CM (Fig. 1C). To confirm the inhibitory role of
propofol on Wnt signaling, the effect of propofol treatment on Wnt
signaling target genes was examined in the 293T cells transfected
with Wnt3 expression plasmids using RT-qPCR analysis. Compared with
DMSO treatment (as a control), propofol treatment significantly
reduced the expression levels of AXIN2, CCND1, NKD1 and LGR5, which
are four well-known Wnt targets (Fig.
1D). Taken together, these data suggested that propofol
inhibited Wnt signaling.
 | Figure 1.Inhibition of Wnt signaling by
propofol. Luciferase reporter assays of 293T cells transfected with
the SuperTopFlash reporter plasmid together with (A) Wnt1 or (B)
Wnt3 expression plasmids. Following 24 h of transfection, the cells
were treated with indicated concentrations of propofol for 48 h and
luciferase activities were measured. The results are shown as the
mean ± standard deviation of three independent experiments and were
analyzed using one-way analysis of variance followed by Dunnett's
test. *P<0.05, **P<0.01 and ***P<0.001, vs. 0 µg/ml
propofol. (C) Luciferase reporter assays of 293T cells transfected
with the SuperTopFlash reporter plasmid. After 24 h, the cells were
treated with indicated concentrations of propofol with or without
Wnt3A-CM for 48 h and luciferase activities were measured. Results
are shown as the mean ± standard deviation of three independent
experiments. Data were analyzed using one-way analysis of variance
followed by Dunnett's test. *P<0.05, **P<0.01, ***P<0.001,
vs. 0 µg/ml propofol. (D) RT-qPCR analysis. 293T cells were
transfected with Wnt3 expression plasmids. After 24 h, the cells
were treated with DMSO or 5 µg/ml propofol for 48 h, and the mRNA
expression of Wnt target genes (AXIN2, CCND1, NKD1 and LGR5) were
analyzed using RT-qPCR analysis. DMSO treatment was used as the
control. The expression of Wnt target genes was normalized to
GAPDH. Data were analyzed using Student's t-test (paired)
*P<0.05, vs. DMSO. Wnt3A-CM, Wnt3A-conditioned medium; RT-qPCR,
reverse transcription-quantitative polymerase chain reaction;
CCND1, cyclin D1; NKD1, naked cuticle homolog 1, LGR5, leucine-rich
repeat-containing G-protein coupled receptor 5. |
Propofol suppresses Wnt signaling in
glioma cells
The present study then determined the effect of
propofol on the Wnt signaling in glioma cells. The LN229 cells were
transfected with the SuperTopFlash reporter plasmid and treated
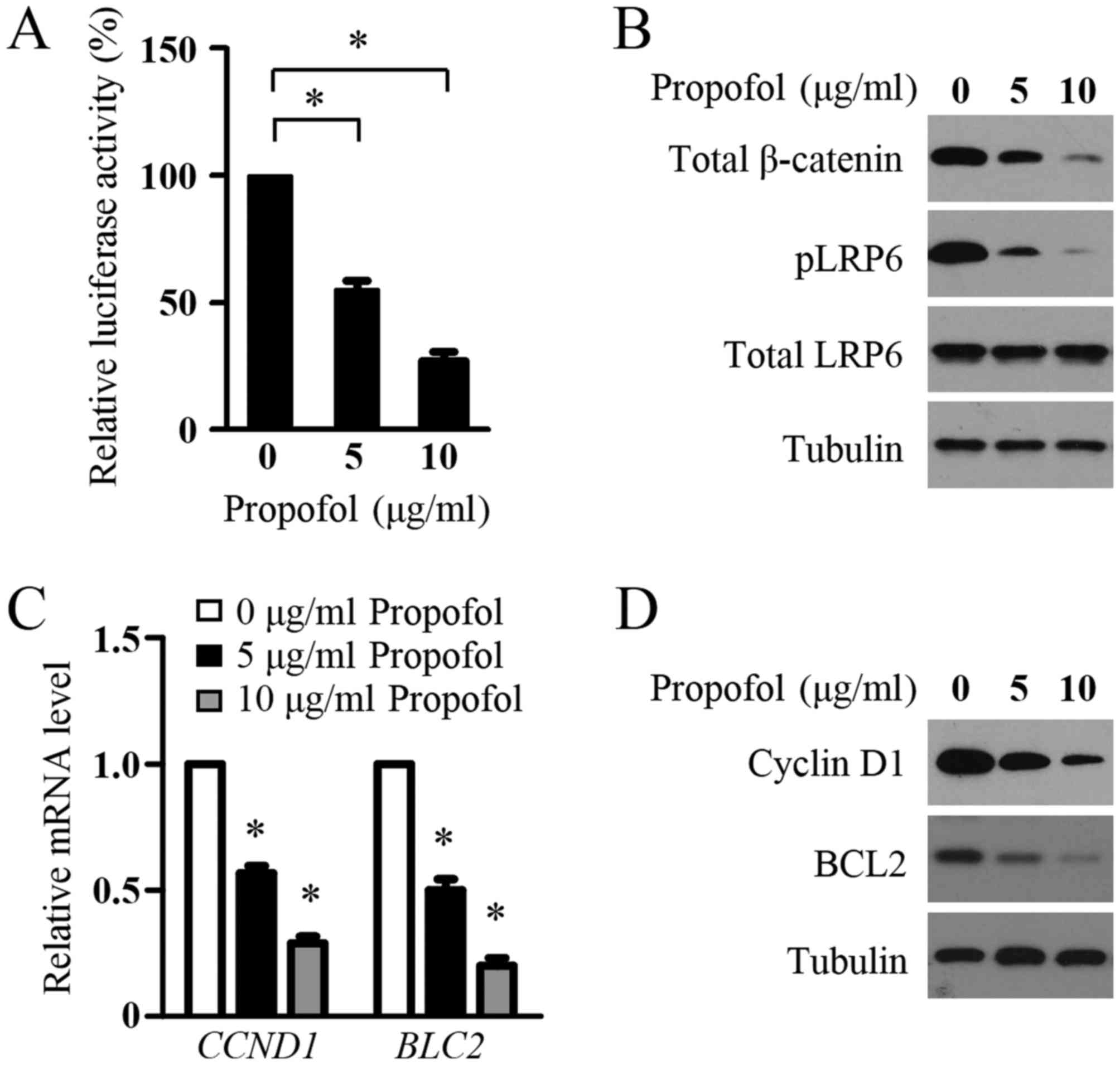
with propofol. As shown in Fig. 2A,
propofol decreased the transcriptional activity of the
SuperTOPflash reporter. In addition, significant decreases in the
protein levels of pLRP6, total LRP6 and total β-catenin were
detected in the LN299 cells following propofol treatment,
indicating the inhibition of Wnt signaling in the glioma cells by
propofol (Fig. 2B). To confirm this
inhibitory role, the present study examined the effect of propofol
treatment on the expression of Wnt target genes, CCND1 and BLC2,
which are two key genes involved in glioma cell proliferation and
survival. Using RT-qPCR analysis, it was found that propofol
treatment resulted in a marked decrease in the mRNA expression
levels of CCND1 and BLC2 in LN299 cells (Fig. 2C). In agreement with the mRNA data,
propofol markedly inhibited the protein levels of CCND1 and BCL2 in
the glioma cells (Fig. 2D). These
results suggested that propofol inhibited Wnt signaling in the
glioma cells.
Propofol inhibits proliferation and
induces apoptosis in glioma cells
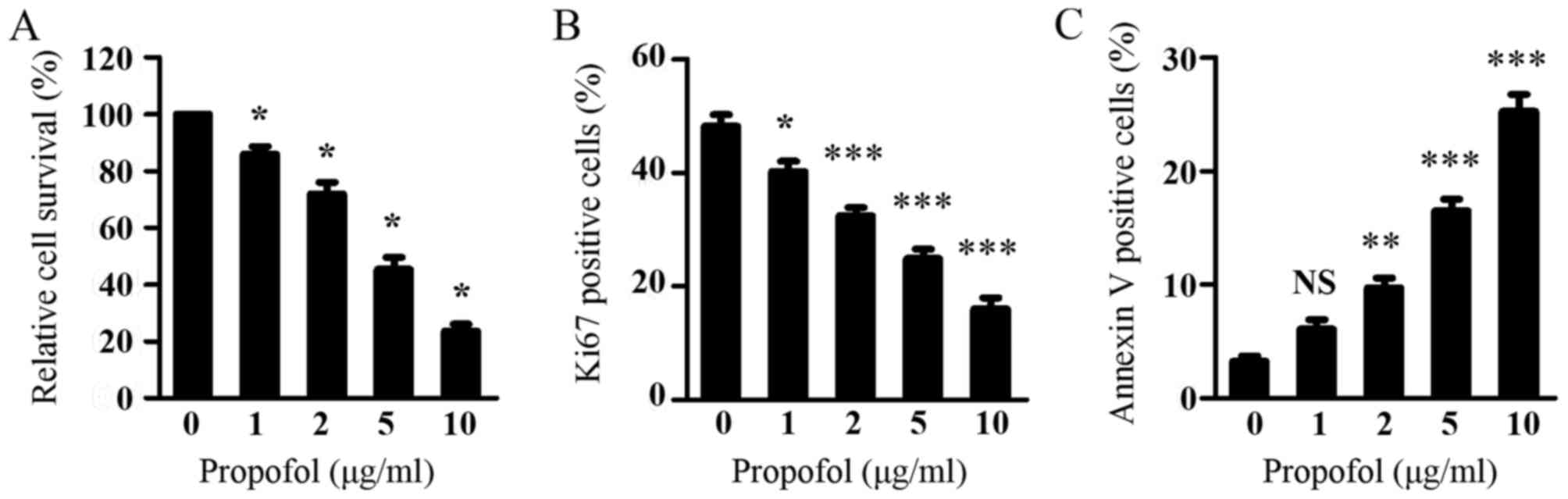
As Wnt signaling is critical in glioma cell
viability and it was shown that propofol suppressed Wnt signaling
in glioma cells, the present study subsequently investigated the
effect of propofol on glioma cell viability using a CCK-8 assay. As
shown in Fig. 3A, propofol treatment
dose-dependently inhibited the viability of the LN299 cells.
Consistent with the inhibitory effect of propofol on the expression
of CCND1, propofol treatment resulted in a dose-dependent decrease
in the percentage of proliferating LN299 cells, as assessed by Ki67
staining (Fig. 3B). Additionally,
propofol treatment led to a significant increase in the percentage
of apoptotic LN299 cells, as measured by Annexin V staining
(Fig. 3C). These results indicated
that propofol inhibited the proliferation and induced the apoptosis
of glioma cells, leading to suppressed glioma cell viability.
Propofol treatment inhibits the growth
of xenograft tumors in nude mice and suppresses Wnt signaling in
tumors
The above results demonstrated the inhibitory role
of propofol on Wnt signaling and cell growth of glioma cells in
vitro, therefore, the present study examined whether such an
inhibitory effect occurs in vivo. A xenograft tumor model
was established by injecting LN299 cells subcutaneously into the
right dorsal flank of nude mice. When the average tumor volume
reached ~100 mm3, the mice were randomly divided to two
groups (n=6/group) and treated with an intraperitoneal injection
with propofol (50 mg/kg). Soybean oil was used as a vehicle and to
dissolve propofol for treating the xenograft animal, as it is
non-toxic and safe for use in mice (29). Propofol treatment significantly
suppressed tumor growth (Fig. 4A) but
had no effect on body weight (Fig.
4B). Propofol also significantly decreased tumor weight
(Fig. 4C). To examine whether
propofol treatment suppressed Wnt signaling in the xenograft
tumors, the expression levels of Wnt target genes were measured in
primary tumors by RT-qPCR analysis. As shown in Fig. 4D, AXIN2, CCND1 and BCL2 were
significantly downregulated in the LN299 xenografts treated with
propofol. In accordance with the mRNA results, propofol treatment
also suppressed the protein levels of CCND1 and BCL2 in the LN299
xenografts (Fig. 4E). The protein
levels of total β-catenin in the LN299 xenografts were also
repressed by propofol treatment (Fig.
4E), confirming the inhibition of Wnt signaling in LN299
xenografts by propofol treatment.
Discussion
Propofol is one of the most widely used intravenous
anesthetic agents for pain relief in patients in clinical surgery.
In addition to its well-known anesthetic advantages, the anticancer
effect of propofol has attracted increasing attention (18). In lung cancer cells, propofol can
promote cell apoptosis by inducing endoplasmic reticulum stress
(20). It has been reported that
propofol inhibits cervical carcinoma cell growth via decreasing the
HOTAIR-mediated mammalian target of rapamycin pathway (22). Propofol has also been found to
attenuate prostate cancer cell malignancy through modulating the
hypoxia-inducible factor-1α pathway (30). Li et al demonstrated that
propofol reduced the level of matrix metalloproteinase, and
suppressed the migration and invasion ability of breast cancer
cells via inhibition of the nuclear factor-κB pathways (31). These findings demonstrate that
propofol exerts antitumor activity in several types of cancer
through various molecular mechanisms. However, whether propofol has
any effects on Wnt signaling and glioma cell growth remains to be
elucidated.
Wnt signaling is critical in the initiation and
development of glioma. Wnt ligands (Wnt1, Wnt3 and Wnt5) and FZD
receptors (FZD2, FZD4, FZD6, FZD7 and FZD9) are overexpressed in
human gliomas (9,10). Multiple negative regulators of Wnt
signaling, including FAT atypical cadherin 1, dkk1, NKD2, secreted
FZD-related protein (sFRP)1 and sFRP2, are downregulated in gliomas
(9). Although β-catenin mutations are
rarely detected in gliomas, elevated levels of β-catenin and high
nuclear accumulation are frequently found in glioma tissue and are
positively correlated with poor prognosis (32). The transcriptional knockdown of Wnt
pathway regulators, including Wnt2 and β-catenin, in glioma cells
significantly represses cell proliferation and invasiveness, and
induces apoptotic cell death (33).
SEN461, a potent WNT signaling inhibitor, exhibits anticancer
activity in gliomas (34). These
findings indicate that WNT signaling is a potential therapeutic
target in gliomas.
In the present study, the effect of propofol on WNT
signaling was determined using a SuperTopFlash reporter assay and
it was found that propofol treatment suppressed the transcriptional
activity of the SuperTOPflash reporter in a dose-dependent manner.
In addition, propofol significantly inhibited the protein levels of
pLRP6 and total β-catenin and the expression of Wnt target genes in
glioma cells, supporting the inhibition of Wnt signaling by
propofol. Propofol treatment also inhibited the growth of glioma
cells in vitro and in vivo, but had no significant
effect on the body weight of propofol-treated mice. Consistent with
the inhibitory role of propofol on Wnt signaling in vitro,
propofol treatment significantly suppressed Wnt signaling in the
xenograft tumors. These data demonstrated that propofol inhibited
Wnt signaling and suppressed glioma cell growth. As Wnt signaling
is also important in the migration and invasiveness of gliomas, and
it was found that propofol decreased Wnt signaling in glioma cells
(35), there is merit in examining
whether propofol treatment affects glioma cell migration or
invasiveness in future investigations.
In conclusion, the data obtained revealed that
propofol is a novel potential Wnt signaling inhibitor, which
suppressed the growth and survival of glioma cells in vitro
and in vivo. The results indicate a novel mechanism of
anticancer activity for propofol and provide novel insights into
the signaling pathways regulated by propofol. The results suggested
the possibility for the potential use of propofol as a novel agent
for the treatment of patients with glioma.
Acknowledgements
The authors would like to thank for Dr. Wei Zhang
for his revision of the manuscript.
Funding
This study was supported by Jingzhou City Science
and Technology Funding (grant no. 2016043).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Author's contributions
RX coordinated and designed the project. WX, JZ, SB,
LK, QM, WL, JG and JL performed the experiments. WX and JZ analyzed
the data. WX, JZ and RX wrote the manuscript.
Ethics approval and consent to
participate
Animal experiments were approved by the Ethics
Committee of Yangtze University (Jingzhou, China).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
APC
|
adenomatous polyposis coli
|
|
CK1
|
casein kinase 1
|
|
GSK3β
|
glycogen synthase kinase 3β
|
|
LRP5
|
lipoprotein receptor-related protein
5
|
|
LRP6
|
lipoprotein receptor-related protein
6
|
References
|
1
|
Zhou Y, Wang Y, Tischfield M, Williams J,
Smallwood PM, Rattner A, Taketo MM and Nathans J: Canonical WNT
signaling components in vascular development and barrier formation.
J Clin Invest. 124:3825–3846. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Anastas JN and Moon RT: WNT signalling
pathways as therapeutic targets in cancer. Nat Rev Cancer.
13:11–26. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chiurillo MA: Role of the Wnt/β-catenin
pathway in gastric cancer: An in-depth literature review. World J
Exp Med. 5:84–102. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kahn M: Can we safely target the WNT
pathway? Nat Rev Drug Discov. 13:513–532. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhou L and Liu Y: Wnt/β-catenin signalling
and podocyte dysfunction in proteinuric kidney disease. Nat Rev
Nephrol. 11:535–545. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Inestrosa NC and Varela-Nallar L: Wnt
signalling in neuronal differentiation and development. Cell Tissue
Res. 359:215–223. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Louis DN: Molecular pathology of malignant
gliomas. Ann Rev Pathol. 1:97–117. 2006. View Article : Google Scholar
|
|
8
|
Omuro A and DeAngelis LM: Glioblastoma and
other malignant gliomas: A clinical review. JAMA. 310:1842–1850.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lee Y, Lee JK, Ahn SH, Lee J and Nam DH:
WNT signaling in glioblastoma and therapeutic opportunities. Lab
Invest. 96:137–150. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang K, Zhang J, Han L, Pu P and Kang C:
Wnt/beta-catenin signaling in glioma. J Neuroimmune Pharmacol.
7:740–749. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu C, Tu Y, Sun X, Jiang J, Jin X, Bo X,
Li Z, Bian A, Wang X, Liu D, et al: Wnt/beta-Catenin pathway in
human glioma: Expression pattern and clinical/prognostic
correlations. Clin Exp Med. 11:105–112. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Augustin I, Goidts V, Bongers A, Kerr G,
Vollert G, Radlwimmer B, Hartmann C, Herold-Mende C, Reifenberger
G, von Deimling A and Boutros M: The Wnt secretion protein
Evi/Gpr177 promotes glioma tumourigenesis. EMBO Mol Med. 4:38–51.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sareddy GR, Panigrahi M, Challa S,
Mahadevan A and Babu PP: Activation of Wnt/beta-catenin/Tcf
signaling pathway in human astrocytomas. Neurochem Int. 55:307–317.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang N, Wei P, Gong A, Chiu WT, Lee HT,
Colman H, Huang H, Xue J, Liu M, Wang Y, et al: FoxM1 promotes
β-catenin nuclear localization and controls Wnt target-gene
expression and glioma tumorigenesis. Cancer Cell. 20:427–442. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang J, Huang K, Shi Z, Zou J, Wang Y,
Jia Z, Zhang A, Han L, Yue X, Liu N, et al: High β-catenin/Tcf-4
activity confers glioma progression via direct regulation of AKT2
gene expression. Neuro Oncol. 13:600–609. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Foltz G, Yoon JG, Lee H, Ma L, Tian Q,
Hood L and Madan A: Epigenetic regulation of wnt pathway
antagonists in human glioblastoma multiforme. Genes Cancer.
1:81–90. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mizobuchi Y, Matsuzaki K, Kuwayama K,
Kitazato K, Mure H, Kageji T and Nagahiro S: REIC/Dkk-3 induces
cell death in human malignant glioma. Neuro Oncol. 10:244–253.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Marik PE: Propofol: Therapeutic
indications and side-effects. Curr Pharm Des. 10:3639–3649. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen X, Lu P, Chen L, Yang SJ, Shen HY, Yu
DD, Zhang XH, Zhong SL, Zhao JH and Tang JH: Perioperative
propofol-paravertebral anesthesia decreases the metastasis and
progression of breast cancer. Tumour Biol. 36:8259–8266. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cui WY, Liu Y, Zhu YQ, Song T and Wang QS:
Propofol induces endoplasmic reticulum (ER) stress and apoptosis in
lung cancer cell H460. Tumour Biol. 35:5213–5217. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Miao Y, Zhang Y, Wan H, Chen L and Wang F:
GABA-receptor agonist, propofol inhibits invasion of colon
carcinoma cells. Biomed Pharmacother. 64:583–588. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang D, Zhou XH, Zhang J, Zhou YX, Ying
J, Wu GQ and Qian JH: Propofol promotes cell apoptosis via
inhibiting HOTAIR mediated mTOR pathway in cervical cancer. Biochem
Biophys Res Commun. 468:561–567. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen X, Wu Q, You L, Chen S, Zhu M and
Miao C: Propofol attenuates pancreatic cancer malignant potential
via inhibition of NMDA receptor. Eur J Pharmacol. 795:150–159.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yang C, Gao J, Yan N, Wu B, Ren Y, Li H
and Liang J: Propofol inhibits the growth and survival of gastric
cancer cells in vitro through the upregulation of ING3. Oncol Rep.
37:587–593. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Willert K, Shibamoto S and Nusse R:
Wnt-induced dephosphorylation of axin releases beta-catenin from
the axin complex. Genes Dev. 13:1768–1773. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-delta delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Roberts KP, Ensrud KM and Hamilton DW: A
comparative analysis of expression and processing of the rat
epididymal fluid and sperm-bound forms of proteins D and E. Biol
Reprod. 67:525–533. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang Z, Li B, Zhou L, Yu S, Su Z, Song J,
Sun Q, Sha O, Wang X, Jiang W, et al: Prodigiosin inhibits
Wnt/β-catenin signaling and exerts anticancer activity in breast
cancer cells. Proc Nat Acad Sci USA. 113:13150–13155. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chen H, Khemtong C, Yang X, Chang X and
Gao J: Nanonization strategies for poorly water-soluble drugs. Drug
Discov Today. 16:354–360. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Huang H, Benzonana LL, Zhao H, Watts HR,
Perry NJ, Bevan C, Brown R and Ma D: Prostate cancer cell
malignancy via modulation of HIF-1α pathway with isoflurane and
propofol alone and in combination. Br J Cancer. 111:1338–1349.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li Q, Zhang L, Han Y, Jiang Z and Wang Q:
Propofol reduces MMPs expression by inhibiting NF-κB activity in
human MDA-MB-231 cells. Biomed Pharmacother. 66:52–56. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu X, Wang L, Zhao S, Ji X, Luo Y and
Ling F: β-Catenin overexpression in malignant glioma and its role
in proliferation and apoptosis in glioblastma cells. Med Oncol.
28:608–614. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Pu P, Zhang Z, Kang C, Jiang R, Jia Z,
Wang G and Jiang H: Downregulation of Wnt2 and beta-catenin by
siRNA suppresses malignant glioma cell growth. Cancer Gene Ther.
16:351–361. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sareddy GR, Kesanakurti D, Kirti PB and
Babu PP: Nonsteroidal anti-inflammatory drugs diclofenac and
celecoxib attenuates Wnt/β-catenin/Tcf signaling pathway in human
glioblastoma cells. Neurochem Res. 38:2313–2322. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Paul I, Bhattacharya S, Chatterjee A and
Ghosh MK: Current understanding on EGFR and Wnt/β-Catenin signaling
in glioma and their possible crosstalk. Genes Cancer. 4:427–446.
2013. View Article : Google Scholar : PubMed/NCBI
|