Introduction
Colorectal cancer (CRC) is one of the most commonly
diagnosed types of cancer and is the second leading cause of
cancer-associated mortality worldwide (1,2).
Etiological studies report that dietary factors serve an important
role in CRC carcinogenesis (3–5). CRC risk
is increased by a high intake of red and processed meats (6), as red meat is a contributing factor in
the initiation of colorectal carcinogenesis (7). However, milk and other dairy products
reportedly have a protective effect against CRC, due to their high
calcium content and bioactive constituents, including vitamin D
(8–10). Fermented dairy products, including
yogurt, also exhibit protective effects against CRC, possibly due
to lactic acid bacteria and their reported ability to inactivate
intestinal carcinogens and therefore reduce CRC risk (11). Although the present epidemiological
evidence is insufficient (12–16),
diet-associated preventative measures may be an important strategy
for CRC reduction.
Maple syrup is a natural sweetener produced by
boiling down sap, which is collected from the sugar maple, Acer
saccharum, and is consumed worldwide by individuals of all ages
(17,18). The sugar maple is distributed
throughout North America, and maple trees serve an important role
in traditional medicine among Native Americans (19). A number of previous studies have
examined the chemical composition and biological properties of
maple-derived products, including maple syrup (20–26).
The climatic conditions during production season
influence maple sap composition, including the color, the aroma and
the taste of the maple syrup, which vary based on the season of sap
collection (27–29). Maple syrup is primarily graded
according to its flavor and transmittance, including visual color
differences, ranging from light-colored and delicately flavored, to
dark-colored and strongly flavored (18). Although the variation in composition
may further lead to different grades of maple syrup, along with
different biological effects, the differences in composition of
ingredients among each grade of maple syrup remain unknown and
require further investigation. In our previous study of the
anticancer effects of different grades of maple syrup, it was
reported that dark-colored maple syrup reduced AKT, also termed
protein kinase B, activation, and therefore significantly inhibited
proliferation and invasion in CRC cells (30). In addition, another previous study
showed that it significantly inhibited growth in other types of
gastrointestinal cancer cell (31).
This suggests that dark-colored maple syrup may be a useful dietary
factor for potentially preventing cancer progression.
In the present study, a shotgun liquid
chromatography-tandem mass spectrometry (LC-MS/MS)-based global
proteomic analysis was performed on human CRC cells treated with
different grades of maple syrup, in order to examine the underlying
mechanism behind dark-colored maple syrup inhibiting CRC
proliferation. Two types of maple syrup, which indicated the
strongest and weakest anticancer effects in our previous study of
colon cancer cells (30), were
selected. A total of 388 proteins were identified that were
differentially expressed in CRC cells treated with dark-grade maple
syrup compared with extra-light grade maple syrup. The current
study focused on the expression of proliferating cell nuclear
antigen (PCNA), which is a key factor of cell cycle regulation.
Therefore, further investigations were conducted on whether changes
of PCNA expression following dark grade maple syrup treatment may
be involved in cell cycle regulation in human CRC cells.
Materials and methods
Materials
Urea was purchased from GE Healthcare Life Sciences
(Little Chalfont, UK), and thiourea and Triton X-100 were obtained
from Nacalai Tesque, Inc., (Kyoto, Japan). All other chemicals and
reagents were purchased from Wako Pure Chemical Industries, Ltd.,
(Osaka, Japan). Maple syrups were purchased at a local grocery
store in Osaka, Japan, in March 2015.
Based on Canadian standards, maple syrup is
classified into the following five grades: AA, extra light; grade
A, light; grade B, medium; grade C, amber, and grade D dark. Since
the present study was performed the maple syrup classification has
changed according to the following: Golden, delicate Taste; amber,
rich Taste; dark, robust taste; and very dark, strong taste
(32). In Japan, these new grading
maple syrup grades have been used since April 2017. Since the
differences in ingredient composition among each grade of maple
syrup are not yet fully understood, in the present study, two
grades of maple syrup were selected: The extra light grade maple
syrup, which has a slightly golden tint and a delicate flavor with
>75% light transmission, and the dark grade maple syrup, which
has a much darker brown color and a strong flavor with <25%
light transmission.
The colorectal cancer DLD-1 cell line was purchased
from the American Type Culture Collection (Manassas, VA, USA). All
cells were cultured in RPMI-1640 medium supplemented with 10% fetal
bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc., Waltham,
MA, USA) at 37°C in an atmosphere containing 5% CO2.
Protein preparation
The DLD-1 cells were plated at a density of
2×105 cells/60-mm dish with RPMI-1640 medium. The next
day, the culture medium was replaced with culture medium with 1%
(v/v) extra light grade maple syrup (extra), dark grade maple syrup
(dark) or without syrup (control). This concentration was selected
due to results from our previous study indicating a lack of
cytotoxic effects against DLD-1 cells due to the high concentration
of sucrose (30). After 72 h, the
cells were solubilized in urea lysis buffer (7 M urea, 2 M
thiourea, 5% 3-[(3-Cholamidopropyl) dimethylammonio]
propanesulfonate and 1% Triton X-100), and the protein
concentration was measured with the Bio-Rad Protein assay (cat. no.
5000006JA; Bio-Rad Laboratories Inc., Hercules, CA, USA), according
to the manufacturer's protocols.
Gel-free digestion was subsequently performed, as
described previously (33). Briefly,
10 µg protein extract from each sample was reduced by adding 45 mM
dithiothreitol and 20 mM tris (2-carboxyethyl) phosphine. The
proteins were subsequently alkylated with 100 mM iodoacetic acid.
Following alkylation, the samples were digested at 37°C for 24 h
using MS-grade trypsin gold (Promega Corporation, Madison, WI,
USA). Finally, the digests were purified using PepClean C-18 Spin
Columns (Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol.
Liquid chromatography tandem-mass
spectrometry (LC-MS/MS) analysis for protein identification
Peptide samples (~2 µg) were injected into a peptide
L-trap column (Chemicals Evaluation and Research Institute, Tokyo,
Japan) with an HTC PAL autosampler (CTC Analytics AG, Zwingen,
Switzerland). The samples were subsequently further separated
through a Paradigm MS4 (AMR Inc., Tokyo, Japan) with a
reverse-phase C18-column (L-column, 3-µm-diameter gel particles,
120 Å pore size, 0.2×150 mm; Chemicals Evaluation and Research
Institute, Tokyo, Japan). The column flow rate was 1 µl/min, and
the mobile phase consisted of 0.1% formic acid in water (solution
A) and acetonitrile (solution B), with a concentration gradient of
5% solution B to 40% solution B over 120 min. Gradient-eluted
peptides were introduced into the mass spectrometer through the
nanoelectrospray ionization (NSI) interface that had a separation
column outlet directly connected with an NSI needle. The peptides
were analyzed with an LTQ ion-trap mass spectrometer (Thermo Fisher
Scientific, Inc.). No sheath or auxiliary gas was used. The MS scan
sequence used was full-scan MS in the normal/centroid mode and
sequential MS/MS in the normal/centroid mode. The positive ion mass
spectra were acquired in a data-dependent manner, with MS/MS
fragmentation performed on the two most intense peaks of every full
MS scan with an isolation width of 1.0 m/z and a collisional
activation amplitude of 35% in the m/z range of 300–2,000.
All MS/MS spectral data were searched against the
SwissProt Homo Sapiens database (https://www.uniprot.org/) using Mascot version 2.4.01
(Matrix Science, Ltd., London, UK). The search criteria were
‘enzyme’ and ‘trypsin’, with the following allowances: ≤2 missed
cleavage peptides; mass tolerance, ± 2.0 Da; MS/MS tolerance, ± 0.8
Da; cysteine carbamidomethylation; and methionine oxidation
modifications.
Semi-quantitative analysis of
identified proteins
The fold-change in expression was calculated as the
log2 ratio of protein abundance (Rsc), evaluated by spectral
counting (34). For comparisons, the
relative amounts of identified proteins were calculated using the
normalized spectral abundance factor (NSAF) (35). Differentially expressed proteins were
considered significant when the Rsc was >0.585 or <-0.585,
corresponding to fold-changes of >1.5 or <0.66,
respectively.
Gene ontology (GO) analysis
The functions of proteins that indicated altered
expression with maple syrup treatment were additionally
investigated. Their sequences were assigned to Kyoto Encyclopedia
of Genes and Genomes (https://www.genome.jp/kegg/kegg_ja.html) signaling
pathway terms to examine their functional annotations using the
Database for Annotation, Visualization, and Integrated Discovery
(DAVID) version 6.8 (http://david.abcc.ncifcrf.gov/home.jsp) (36–38).
P<0.05 was considered to indicate a significant category.
Western blot analysis
Total protein (5 µg) that had been prepared as
aforementioned was mixed with loading buffer and boiled at 95°C for
10 min. The proteins were then separated on a 12% SDS-PAGE gel. The
separated proteins were transferred to polyvinylidene fluoride
membranes (Merck KGaA, Darmstadt, Germany) for 30 min at 15 V.
Following blocking in TBS-Tween-20 (0.1%) buffer (Cell Signaling
Technology, Inc., Danvers, MA, USA) with 5% skimmed milk for 2 h at
room temperature, the membranes were incubated with an anti- PCNA
antibody (1:20,000; cat. no. 13110; Cell Signaling Technology,
Inc.) at 4°C overnight. The membranes were subsequently washed and
incubated with horseradish peroxidase-conjugated anti-rabbit
immunoglobulin(Ig)G antibody (1:4,000; cat. no. A106PU; American
Qualex, San Clemente, CA, USA) at room temperature for 1 h. The
blots were washed and visualized with SuperSignal West Dura
Extended Duration substrate (Thermo Fisher Scientific, Inc.). The
bands were analyzed with the myECL Imager system 2.0 software
(Thermo Fisher Scientific, Inc.). The membranes were subsequently
stripped by Restore Western Blot Stripping buffer (Thermo Fisher
Scientific, Inc.), and the same membranes were re-probed with an
anti-β-actin antibody (1:5,000 dilution; cat. no. sc-47778; Santa
Cruz Biotechnology, Inc., Dallas, TX, USA) at 4°C overnight, which
served as the protein loading control. The relative quantities of
PCNA over β-actin were used to evaluate PCNA expression under
different conditions. All western blot analyses were performed as
three independent experiments.
Cell cycle analysis by flow
cytometry
To analyze cell cycle distribution, DNA was stained
with propidium iodide (PI; Nacalai Tesque, Inc., Kyoto, Japan).
Briefly, DLD-1 cells were plated at a density of 2×105
cells/100-mm dish in culture medium. The next day, the culture
medium was replaced with 1% (v/v) extra light grade maple syrup
(Extra), dark grade maple syrup (Dark) or without syrup (control).
Following 72 h of incubation at 37°C, the cells were washed with
PBS and fixed in ice-cold 70% ethanol at 4°C for 2 h. The cells
were subsequently treated with 0.25 mg/ml RNase in PBS for 60 min
at 37°C, followed by staining with 50 µg/ml PI in PBS for 30 min at
4°C in the dark. Cell proportion in different phases of the cell
cycle was determined using a BD LSRFortessa flow cytometer (BD
Biosciences, San Jose, CA, USA) and analyzed using FACS DIVA
software v8.0.1 (BD Biosciences).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) using TaqMan array
analysis
Total RNA was extracted from treated DLD-1 cells
using the GenElute Mammalian Total RNA Miniprep kit (cat. no.
RTN70-1KT; Sigma-Aldrich; Merck KGaA), according to the
manufacturer's protocols. From the extracted RNA, cDNA was
synthesized using the High Capacity cDNA Reverse Transcription kit
(cat. no. 4368814; Thermo Fisher Scientific, Inc.), according to
the manufacturer's protocols. Analysis was performed using TaqMan
Array Human Cyclins & Cell Cycle Regulation 96-Well Plates
(cat. no. 4414123; Thermo Fisher Scientific, Inc.) in the 7500
system (Applied Biosystems; Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocols. The thermocycling
conditions were as follows: Denaturation at 95°C for 20 sec,
followed by 40 cycles of amplification at 95°C for 3 sec and 60°C
for 30 sec. The relative gene expression was calculated using the
2−ΔΔCq method (39–43). The
ΔΔCq method uses the normalized ΔCq value of each sample, which was
calculated with 18S rRNA as the endogenous control gene. The ΔΔCq
value is the difference between treated and control samples.
Finally, the fold-change was determined as 2−ΔΔCq.
Statistical analysis
All experiments were repeated at a minimum of three
times. All data are presented as the mean ± standard error of the
mean (SEM). The data were analyzed by one-way analysis of variance
followed by Tukey's post hoc test. P<0.05 was considered to
indicate a statistically significant difference. Computations were
performed using GraphPad Prism version 5 (GraphPad Software Inc.,
La Jolla, CA, USA).
Results
Identification and semi-quantitative
comparisons of differentially expressed proteins in maple
syrup-treated DLD-1 cells
To investigate the inhibitory effects of maple syrup
on CRC proliferation, shotgun proteomics was used to examine the
molecular profile of proteins that were regulated by maple syrup
treatment. Applying the aforementioned search parameters, a total
of 575 proteins were identified in DLD-1 cells treated with extra
light grade maple syrup, and 549 proteins in DLD-1 cells treated
with dark-colored maple syrup. Proteins were categorized into the
‘Extra’ and ‘Dark’ groups accordingly.
The proteins expressed in maple syrup-treated DLD-1
cells were further evaluated using a label-free semi-quantitative
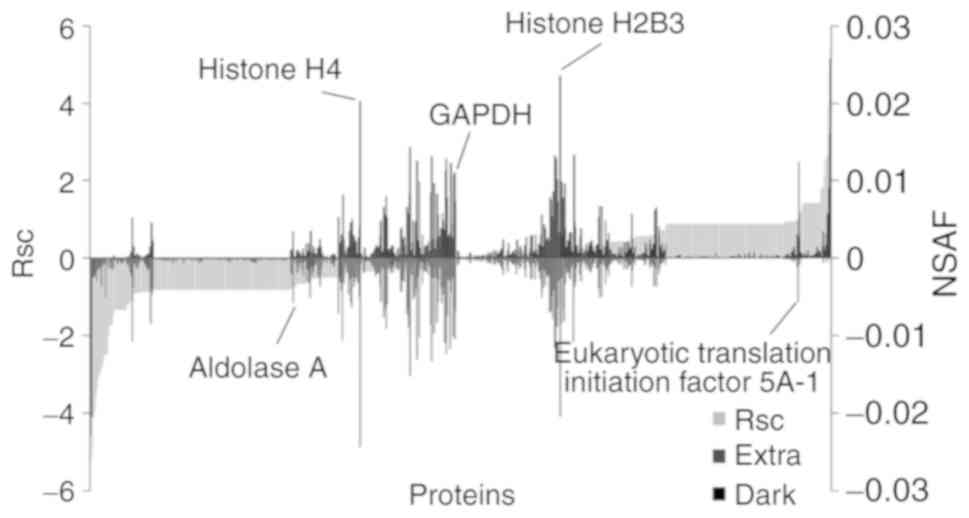
method based on spectral counting. The Rsc values were calculated
for the proteins identified in the Extra and Dark groups. A
positive Rsc value indicated increased expression with dark-colored
maple syrup treatment, and a negative value indicated reduced
expression with dark-colored maple syrup treatment (Fig. 1; light grey area). The NSAF value was
also calculated for each protein identified in the Extra and Dark
groups. Proteins with a >0.585 and <-0.585 Rsc value were
considered candidate dark-colored maple syrup-regulated
proteins.
This semi-quantitative procedure resulted in the
identification of 388 proteins that were differentially expressed
with dark-colored maple syrup treatment (data not shown). Maple
syrup treatment did not alter the expression of housekeeping
proteins, including glyceraldehyde-3-phosphate dehydrogenase and
histone H4 (Fig. 1).
Functional annotation of proteins
regulated by maple syrup
GO analysis of the candidate dark-colored maple
syrup-regulated proteins was performed. GO terms associated with
‘pathway’ were searched for in DAVID (Fig. 2), and the focus was on proteins
classified as associated with the ‘cell cycle’ (Table I).
 | Table I.Proteins categorized as associated
with the ‘cell cycle’ in Gene Ontology analysis. |
Table I.
Proteins categorized as associated
with the ‘cell cycle’ in Gene Ontology analysis.
| Accession
number | Description | Fold-change
(Rsc) |
|---|
| P33992 | DNA replication
licensing factor MCM5 | −0.812 |
| P49736 | DNA replication
licensing factor MCM2 | −0.812 |
| P33991 | DNA replication
licensing factor MCM4 | −0.812 |
| P12004 | Proliferating cell
nuclear antigen | −0.781 |
| P62258 | 14-3-3 protein
ε | −0.648 |
| Q14566 | DNA replication
licensing factor MCM6 | 0.884 |
| Q13547N | Histone deacetylase
1 | 0.884 |
| P61981 | 14-3-3 protein
γ | 2.107 |
| P27348 | 14-3-3 protein
τ | 2.573 |
Effects of maple syrup on PCNA
expression in DLD-1 cells
The expression of PCNA protein in maple
syrup-treated DLD-1 cells was examined. The results of the present
study indicated a significant decrease in PCNA expression with
dark-colored maple syrup treatment (Dark) compared with that in the
cells treated with extra light grade maple syrup treatment (Extra)
and the untreated (control) cells (Fig.
3).
Effects of maple syrup on cell cycle
progression of CRC cells
The present study also investigated whether the
dark-colored maple syrup-induced decrease in PCNA expression
affected cell cycle progression in DLD-1 cells. Flow cytometry
analysis revealed a significantly increased cell population in the
S phase (P<0.01), and a decreased population in the
G2/M phase (P<0.05) in the Dark group compared with
that in the Extra group and the control group (Fig. 4A and B).
Effects of maple syrup on the
expression of cell cycle-associated factors in CRC cells
Using the TaqMan Array Human Cyclins & Cell
Cycle Regulation 96-Well Plate, qPCR was conducted to examine the
molecular profile of cell cycle-associated mRNAs that were
regulated by maple syrup treatment. RT-qPCR analysis was performed
with DLD-1 cells treated with extra light or dark-colored maple
syrup. In the Dark group, fold-changes were induced in the relative
expression of cell cycle-associated factors. Expression levels in
the Extra group were set as 1, and the fold-changes following
dark-colored maple syrup treatment were evaluated using the DDCq
method. Among the 44 tested cell cycle-associated genes, 12 genes
indicated changes in expression of >2-fold in DLD-1 cells
treated with dark-colored maple syrup treatment compared with extra
light grade maple syrup (Fig. 5). It
was indicated that dark-colored maple syrup treatment reduced
cyclin-dependent kinase 4 (CDK4), CDK6 and transforming growth
factor β1 (TGFB1) expression, and induced cyclin-dependent kinase
inhibitor 2B (CDKN2B) expression (Fig.
5).
 | Figure 5.Expression profile of cell
cycle-associated genes in maple syrup-treated DLD-1 cells, based on
polymerase chain reaction array analysis. Extra, extra light maple
syrup; Dark, dark-colored maple syrup; CCNB1, cyclin B1; CDK6,
cyclin-dependent kinase 6; CDK7, cyclin-dependent kinase 7; GSK3B,
glycogen synthase kinase 3 beta; HDAC4, histone deacetylase 4; ATR,
ATR serine/threonine kinase; CDC25A, cell division cycle 25A; E2F1,
E2F transcription factor 1; E2F4, E2F transcription factor 4;
TGFB1, transforming growth factor, beta 1; CDK4, cyclin-dependent
kinase 4; CDKN2B, cyclin-dependent kinase inhibitor 2B. |
Discussion
In the present study, a gel-free LC-MS/MS-based
proteomics approach was applied to examine the underlying mechanism
of CRC cell growth inhibition by dark-colored maple syrup. Using a
semi-quantitative method of spectral counting, a total of 388
proteins were identified that indicated >1.5-fold changes in
expression following maple syrup treatment. The roles of these
identified proteins were examined with GO analysis, focusing on the
functions of proteins classified as associated with the ‘cell
cycle’, since these proteins serve important roles in the
proliferation system. The study also focused on PCNA, which is a
member of this pathway. A western blot analysis was subsequently
performed to validate the spectral counting results, and to
determine whether dark-colored maple syrup treatment led to
decreased PCNA expression in DLD-1 cells.
PCNA has been reported to exhibit different
behaviors depending on the cell cycle phase, and serves important
roles in DNA replication and DNA repair (44–47). PCNA
also recruits chromatin remodeling and epigenetic modification
factors (48). During the S phase of
the cell cycle, PCNA is reportedly localized in the active
replication site, and has the ability to differentiate between
early, mid and late S phase (49,50).
Therefore, PCNA expression is considered to be associated with
S-phase cell cycle progression.
The present study observed that dark-colored maple
syrup treatment decreased PCNA expression, which induced the
S-phase cell cycle arrest of DLD-1 cells. Previous reports also
indicate that induction of S-phase cell cycle arrest is accompanied
by decreased PCNA expression in a number of tumor cells, including
in CRC cells treated with functional compounds, such as resveratrol
and oxoaporphine metal complexes (51,52).
Previous data also indicate that the induction of S-phase cell
cycle arrest is associated with changes in the expression of
cyclins and CDKs in these cells. Therefore, maple syrup treatment
may influence the cell cycle-associated gene expression in CRC
cells. Accordingly, we hypothesized that dark-colored maple syrup
induced S-phase cell cycle arrest in DLD-1 cells, which may affect
the expression of cell cycle-associated genes. The study findings
supported this hypothesis.
CDK4/6 is a key kinase in cell cycle promotion, and
is currently considered a molecular target for anticancer drug
development (53,54). The CDK4/6 inhibitor palbociclib is
clinically used for the treatment of advanced breast cancer
(55,56). The results of the present study
support the possibility that the bioactive compounds in
dark-colored maple syrup may be useful in the development of novel
anticancer drugs for treatment of CRC and other types of advanced
cancer. In addition, CDKN2B is also known as a cell-cycle regulator
via its interaction with CDK4 and CDK6, and tumor suppressor genes,
including p53 and p18 (57).
Therefore, it can by hypothesized that the bioactive compounds,
which have been indicated to upregulate CDKN2B expression may be
present in dark grade maple syrup and may be a useful resource in
developing novel anticancer drugs. In addition, these bioactive
compounds may have an inhibitory effect on the TGF signaling
pathway by suppressing TGF-β1 expression by upregulating CDNK2B
expression, since previous reports demonstrated that downregulation
of CDKN2B expression induced an increase of TGFβ1 expression in
human smooth muscle cells and umbilical vein endothelial cells
(58). A previous study reported that
the ginnalins-polyphenols that are present in maple syrup inhibit
proliferation through S-phase cell cycle arrest (59). However, the effects of ginnalins on
PCNA, CDK4/6 and CDKN2B expression in CRC cells are not well
understood. Further studies are required to identify the bioactive
compounds in dark-colored maple syrup that are responsible for
inhibiting the expression of PCNA, CDK4/6 and/or CDKN2B, and
therefore, inducing cell cycle arrest. Further clarification is
required in order to examine whether dark-colored maple syrup
inhibits proliferation through S-phase cell cycle arrest by
regulating cell cycle-associated gene expression in other types of
cancer cells.
In conclusion, the present study indicated that
dark-colored maple syrup induced S-phase cell cycle arrest in CRC
cells by reducing PCNA expression and regulating cell
cycle-associated genes. These findings suggest that dark-colored
maple syrup may be a useful dietary factor, with a potential
preventative effect against CRC. Compounds in dark-colored maple
syrup may also be useful for the development of novel anticancer
drugs for colorectal cancer treatment.
Acknowledgements
Not applicable.
Funding
The present study was supported in part by the
MEXT-Supported Program of the Strategic Research Foundation at
Private Universities (grant no. S1411037).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
TY and AT designed the study and analyzed the data.
TY and TN performed the experiments. TY drafted the manuscript. AT
critically evaluated the study and the final version of the
manuscript. All authors participated in discussion of the work and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
CRC
|
colorectal cancer
|
|
LC-MS/MS
|
liquid chromatography-tandem mass
spectrometry
|
|
NSAF
|
normalized spectral abundance
factor
|
|
PBS
|
phosphate-buffered saline
|
|
SEM
|
standard error of measurement
|
|
GO
|
Gene Ontology
|
|
PCNA
|
proliferating cell nuclear antigen
|
|
CDK
|
cyclin-dependent kinase
|
References
|
1
|
Tariq K and Ghias K: Colorectal cancer
carcinogenesis: A review of mechanisms. Cancer Biol Med.
13:120–135. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mármol I, Sánchez-de-Diego C, Pradilla
Dieste A, Cerrada E and Rodriguez Yoldi MJ: Colorectal carcinoma: A
general overview and future perspectives in colorectal cancer. Int
J Mol Sci. 18(pii): E1972017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Armstrong B and Doll R: Environmental
factors and cancer incidence and mortality in different countries,
with special reference to dietary practices. Int J Cancer.
15:617–631. 1975. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Doll R and Peto R: The causes of cancer:
Quantitative estimates of avoidable risks of cancer in the United
States today. J Natl Cancer Inst. 66:1191–1308. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Song M, Garrett WS and Chan AT: Nutrients,
foods, and colorectal cancer prevention. Gastroenterology.
148:1244–1260.e16. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chan DS, Lau R, Aune D, Vieira R,
Greenwood DC, Kampman E and Norat T: Red and processed meat and
colorectal cancer incidence: Meta-analysis of prospective studies.
PLoS One. 6:e204562011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chao A, Thun MJ, Connell CJ, McCullough
ML, Jacobs EJ, Flanders WD, Rodriguez C, Sinha R and Calle EE: Meat
consumption and risk of colorectal cancer. JAMA. 293:172–182. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Holt PR, Atillasoy EO, Gilman J, Guss J,
Moss SF, Newmark H, Fan K, Yang K and Lipkin M: Modulation of
abnormal colonic epithelial cell proliferation and differentiation
by low-fat dairy foods: A randomized controlled trial. JAMA.
280:1074–1079. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Glinghammar B, Venturi M, Rowland IR and
Rafter JJ: Shift from a dairy product-rich to a dairy product-free
diet: Influence on cytotoxicity and genotoxicity of fecal
water-potential risk factors for colon cancer. Am J Clin Nutr.
66:1277–1282. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Norat T and Riboli E: Dairy products and
colorectal cancer. A review of possible mechanisms and
epidemiological evidence. Eur J Clin Nutr. 57:1–17. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wollowski I, Ji ST, Bakalinsky AT,
Neudecker C and Pool-Zobel BL: Bacteria used for the production of
yogurt inactivate carcinogens and prevent DNA damage in the colon
of rats. J Nutr. 129:77–82. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kesse E, Boutron-Ruault MC, Norat T,
Riboli E and Clavel-Chapelon F; E3N Group, : Dietary calcium,
phosphorus, vitamin D, dairy products and the risk of colorectal
adenoma and cancer among French women of the E3N-EPIC prospective
study. Int J Cancer. 117:137–144. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cho E, Smith-Warner SA, Spiegelman D,
Beeson WL, van den Brandt PA, Colditz GA, Folsom AR, Fraser GE,
Freudenheim JL, Giovannucci E, et al: Dairy foods, calcium, and
colorectal cancer: A pooled analysis of 10 cohort studies. J Natl
Cancer Inst. 96:1015–1022. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Giovannucci E, Rimm EB, Stampfer MJ,
Colditz GA, Ascherio A and Willett WC: Intake of fat, meat, and
fiber in relation to risk of colon cancer in men. Cancer Res.
54:2390–2397. 1994.PubMed/NCBI
|
|
15
|
Park Y, Leitzmann MF, Subar AF, Hollenbeck
A and Schatzkin A: Dairy food, calcium, and risk of cancer in the
NIH-AARP diet and health study. Arch Intern Med. 169:391–401. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lee SA, Shu XO, Yang G, Li H, Gao YT and
Zheng W: Animal origin foods and colorectal cancer risk: A report
from the Shanghai Women's health study. Nutr Cancer. 61:194–205.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ball DW: The chemical composition of maple
syrup. J Chem Educ. 84:16472007. View Article : Google Scholar
|
|
18
|
Perkins TD and van den Berg AK: Maple
syrup-production, composition, chemistry, and sensory
characteristics. Adv Food Nutr Res. 56:101–143. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Arnason T, Hebda RJ and Johns T: Use of
plants for food and medicine by Native People's of eastern Canada.
Can J Bot. 59:2189–2325. 1981. View
Article : Google Scholar
|
|
20
|
Apostolidis E, Li L, Lee C and Seeram NP:
In vitro evaluation of phenolic-enriched maple syrup extracts for
inhibition of carbohydrate hydrolyzing enzymes relevant to type 2
diabetes management. J Funct Foods. 3:100–106. 2011. View Article : Google Scholar
|
|
21
|
Legault J, Girard-Lalancette K, Grenon C,
Dussault C and Pichette A: Antioxidant activity, inhibition of
nitric oxide overproduction, and in vitro antiproliferative effect
of maple sap and syrup from Acer saccharum. J Med Food. 13:460–468.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
González-Sarrías A, Li L and Seeram NP:
Effects of maple (Acer) plant part extracts on proliferation,
apoptosis and cell cycle arrest of human tumorigenic and
non-tumorigenic colon cells. Phytother Res. 26:995–1002. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Theriault M, Caillet S, Kermasha S and
Lacroix M: Antioxidant, antiradical and antimutagenic activities of
phenolic compounds present in maple products. Food Chem.
98:490–501. 2006. View Article : Google Scholar
|
|
24
|
Hawco CL, Wang Y, Taylor M and Weaver DF:
A maple syrup extract prevents β-amyloid aggregation. Can J Neurol
Sci. 43:198–201. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Nagai N, Ito Y and Taga A: Comparison of
the enhancement of plasma glucose levels in type 2 diabetes otsuka
long-evans tokushima fatty rats by oral administration of sucrose
or maple syrup. J Oleo Sci. 62:737–743. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Nagai N, Yamamoto T, Tanabe W, Ito Y,
Kurabuchi S, Mitamura K and Taga A: Changes in plasma glucose in
otsuka long-evans tokushima fatty rats after oral administration of
maple syrup. J Oleo Sci. 64:331–335. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kim YT and Leech RH: Effects of climatic
conditions on sap flow in sugar maple. Forest Chron. 61:303–307.
1985. View Article : Google Scholar
|
|
28
|
Marvin JW and Erickson RO: A statistical
evaluation of some of the factors responsible for the flow of sap
from the sugar maple. Plant Physiol. 31:57–61. 1956. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Houle D, Paquette A, Côté B, Logan T,
Power H, Charron I and Duchesne L: Impacts of climate change on the
timing of the production season of maple syrup in Eastern Canada.
PLoS One. 10:e01448442015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yamamoto T, Uemura K, Moriyama K, Mitamura
K and Taga A: Inhibitory effect of maple syrup on the cell growth
and invasion of human colorectal cancer cells. Oncol Rep.
33:1579–1584. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yamamoto T, Sato K, Kubota Y, Mitamura K
and Taga A: Effect of dark-colored maple syrup on cell
proliferation of human gastrointestinal cancer cell. Biomed Rep.
7:6–10. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Maple products regulation. https://laws-lois.justice.gc.ca/eng/regulations/C.R.C.,_c._289/page-8.html#h-24
|
|
33
|
Bluemlein K and Ralser M: Monitoring
protein expression in whole-cell extracts by targeted label- and
standard-free LC-MS/MS. Nat Protoc. 6:859–869. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Old WM, Meyer-Arendt K, Aveline-Wolf L,
Pierce KG, Mendoza A, Sevinsky JR, Resing KA and Ahn NG: Comparison
of label-free methods for quantifying human proteins by shotgun
proteomics. Mol Cell Proteomics. 4:1487–1502. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zybailov B, Coleman MK, Florens L and
Washburn MP: Correlation of relative abundance ratios derived from
peptide ion chromatograms and spectrum counting for quantitative
proteomic analysis using stable isotope labeling. Anal Chem.
77:6218–6224. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Dennis G Jr, Sherman BT, Hosack DA, Yang
J, Gao W, Lane HC and Lempicki RA: DAVID: Database for annotation,
visualization, and integrated discovery. Genome Biol. 4:P32003.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Huang da W, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Huang da W, Sherman BT and Lempicki RA:
Bioinformatics enrichment tools: Paths toward the comprehensive
functional analysis of large gene lists. Nucleic Acids Res.
37:1–13. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Parikh P, Bai H, Swartz MF, Alfieris GM
and Dean DA: Identification of differentially regulated genes in
human patent ductus arteriosus. Exp Biol Med (Maywood).
241:2112–2118. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Carbotti G, Nikpoor AR, Vacca P, Gangemi
R, Giordano C, Campelli F, Ferrini S and Fabbi M: IL-27 mediates
HLA class I up-regulation, which can be inhibited by the IL-6
pathway, in HLA-deficient small cell lung cancer cells. J Exp Clin
Cancer Res. 36:1402017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Adnan M, Morton G and Hadi S: Analysis of
rpoS and bolA gene expression under various stress-induced
environments in planktonic and biofilm phase using 2(−ΔΔCT) method.
Mol Cell Biochem. 357:275–282. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Soejima M and Koda Y: TaqMan-based
real-time polymerase chain reaction for detection of FUT2 copy
number variations: Identification of novel Alu-mediated deletion.
Transfusion. 51:762–769. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kelman Z: PCNA: Structure, functions and
interactions. Oncogene. 14:629–640. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Majka J and Burgers PM: The PCNA-RFC
families of DNA clamps and clamp loaders. Prog Nucleic Acid Res Mol
Biol. 78:227–260. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Moldovan GL, Pfander B and Jentsch S:
PCNA, the maestro of the replication fork. Cell. 129:665–679. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Bártová E, Suchánková J, Legartová S,
Malyšková B, Hornáček M, Skalníková M, Mašata M, Raška I and
Kozubek S: PCNA is recruited to irradiated chromatin in late
S-phase and is most pronounced in G2 phase of the cell cycle.
Protoplasma. 254:2035–2043. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Budhavarapu VN, Chavez M and Tyler JK: How
is epigenetic information maintained through DNA replication?
Epigenetics Chromatin. 6:322013. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Celis JE and Celis A: Cell cycle-dependent
variations in the distribution of the nuclear protein cyclin
proliferating cell nuclear antigen in cultured cells: Subdivision
of S phase. Proc Natl Acad Sci USA. 82:3262–3266. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Schönenberger F, Deutzmann A, Ferrando-May
E and Merhof D: Discrimination of cell cycle phases in
PCNA-immunolabeled cells. BMC Bioinformatics. 16:1802015.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Liu B, Zhou Z, Zhou W, Liu J, Zhang Q, Xia
J, Liu J, Chen N, Li M and Zhu R: Resveratrol inhibits
proliferation in human colorectal carcinoma cells by inducing
G1/S-phase cell cycle arrest and apoptosis through
caspase/cyclin-CDK pathways. Mol Med Rep. 10:1697–1702. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Qin JL, Shen WY, Chen ZF, Zhao LF, Qin QP,
Yu YC and Liang H: Oxoaporphine metal complexes (CoII,
NiII, ZnII) with high antitumor activity by
inducing mitochondria-mediated apoptosis and S-phase arrest in
HepG2. Sci Rep. 7:460562017. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Zhao H, Hu X, Cao K, Zhang Y, Zhao K, Tang
C and Feng B: Synthesis and SAR of
4,5-dihydro-1H-pyrazolo[4,3-h]quinazoline derivatives as potent and
selective CDK4/6 inhibitors. Eur J Med Chem. 157:935–945. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Goel S, DeCristo MJ, Watt AC, BrinJones H,
Sceneay J, Li BB, Khan N, Ubellacker JM, Xie S, Metzger-Filho O, et
al: CDK4/6 inhibition triggers anti-tumour immunity. Nature.
548:471–475. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Finn RS, Crown JP, Lang I, Boer K,
Bondarenko IM, Kulyk SO, Ettl J, Patel R, Pinter T, Schmidt M, et
al: The cyclin-dependent kinase 4/6 inhibitor palbociclib in
combination with letrozole versus letrozole alone as first-line
treatment of oestrogen receptor-positive, HER2-negative, advanced
breast cancer (PALOMA-1/TRIO-18): A randomised phase 2 study.
Lancet Oncol. 16:25–35. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Cristofanilli M, Turner NC, Bondarenko I,
Ro J, Im SA, Masuda N, Colleoni M, DeMichele A, Loi S, Verma S, et
al: Fulvestrant plus palbociclib versus fulvestrant plus placebo
for treatment of hormone-receptor-positive, HER2-negative
metastatic breast cancer that progressed on previous endocrine
therapy (PALOMA-3): Final analysis of the multicentre,
double-blind, phase 3 randomised controlled trial. Lancet Oncol.
17:425–439. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Krimpenfort P, Ijpenberg A, Song JY, van
der Valk M, Nawijn M, Zevenhoven J and Berns A: p15Ink4b is a
critical tumour suppressor in the absence of p16Ink4a. Nature.
448:943–946. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Nanda V, Downing KP, Ye J, Xiao S, Kojima
Y, Spin JM, DiRenzo D, Nead KT, Connolly AJ, Dandona S, et al:
CDKN2B regulates TGFβ signaling and smooth muscle cell investment
of hypoxic neovessels. Circ Res. 118:230–240. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
González-Sarrías A, Ma H, Edmonds ME and
Seeram NP: Maple polyphenols, ginnalins A-C, induce S- and
G2/M-cell cycle arrest in colon and breast cancer cells mediated by
decreasing cyclins A and D1 levels. Food Chem. 136:636–642. 2013.
View Article : Google Scholar : PubMed/NCBI
|