Introduction
Lung cancer is a global health issue and the leading
cause of cancer-related mortality. Non-small cell lung cancer
(NSCLC) accounts for 80–85% of all lung cancer cases (1). Despite the optimization of
chemotherapy regimens, treatment outcomes for advanced NSCLC remain
disappointing.
Gefitinib and erlotinib are orally administered,
small-molecule epidermal growth factor receptor (EGFR)-tyrosine
kinase inhibitors (TKIs) that improve the survival of NSCLC
patients and caused a paradigm shift for the treatment of NSCLC.
Patients with EGFR-activating mutations greatly benefit from
treatment with EGFR-TKIs (2–4).
However, the presence of K-Ras mutation is associated with primary
resistance to EGFR-TKIs (5,6). In NSCLC, 15–30% of adenocarcinoma
patients possess a gain of function mutation in the K-Ras gene,
meaning that for these patients, their tumors fail to respond to
EGFR-TKIs (7,8). Thus, clinical research of new
treatment strategies for NSCLC patients is urgently needed.
Angiogenesis is a complex process regulated by
several pro- and anti-angiogenic factors. Vascular endothelial
growth factor (VEGF) and platelet-derived growth factor (PDGF) are
critical factors in the promotion of angiogenesis in NSCLC
(9,10). Activation of VEGF and PDGF
stimulated downstream signaling pathways, including
phosphatidylinositol-3-kinase (PI3K) and extracellular
signal-regulated kinase (ERK) (11–13).
The Ras/Raf/MEK/ERK and Ras/PI3K/PTEN/Akt pathways interact to
regulate growth and play key roles in the transmission of
proliferative signals. Therefore, the overexpression of VEGF and
PDGF is correlated with tumor progression of NSCLC patients and is
a strong prognostic indicator in NSCLC (14–16).
In NSCLC, activation of K-Ras leads to ERK1/2 overexpression
through the Raf/MEK/ERK signaling pathway (17–19).
Hence, inhibition of the Ras/RAF/MEK/ERK signaling pathway is an
important strategy in anticancer drug development.
Sorafenib (BAY 43-9006) is an oral multikinase
inhibitor that decreases the activity of C-RAF and B-RAF in the
RAF/MEK/ERK signaling pathway and targets the VEGF receptor family
(VEGFR-2 and VEGFR-3) and PDGF receptor familyβ (PDGFRβ) (20). Single-agent sorafenib showed
preclinical and clinical activity against NSCLC (21–23).
In xenograft models administered a combination of sorafenib and
anticancer agents, such as vinorelbine, cisplatin and gefitinib,
the anti-proliferative effect is at least as efficacious as
sorafenib alone and the treatment is well-tolerated (24). The safety profile of sorafenib in
previous trials has increased the feasibility of using the drug in
combination with cytotoxic and cytostatic agents. However,
sorafenib administered concurrently with chemotherapy does not
improve patient outcomes compared with chemotherapy alone in
advanced-stage NSCLC. The ESCAPE trial assessed the efficacy and
safety of sorafenib in combination with carboplatin and paclitaxel
in 926 patients with advanced NSCLC. There was no clinical benefit
observed from adding sorafenib to carboplatin and paclitaxel (CP)
chemotherapy as first-line treatment for NSCLC. Patients with
squamous cell histology had greater mortality (25). The subsequent NExUS trial of
sorafenib in combination with gemcitabine/cisplatin in a planned
900 patients with non-squamous advanced NSCLC (NCT00449033) was
also stopped early as it failed to meet its primary endpoint of OS
(26).
One potential explanation for this lack of benefit
is a negative interaction or antagonism between chemotherapy and
sorafenib when delivered concomitantly. Support for this line of
reasoning is provided by preclinical data demonstrating that
sorafenib induce primarily a cytostatic effect resulting from a G1
cell cycle arrest in NSCLC cell lines (27,28),
reducing cell cycle phase-dependent (S and G2/M phase) cytotoxicity
of chemotherapy. At present, sequential administration is
considered to be a promising therapeutic approach in NSCLC as well
as in other types of cancer. Sequential administration avoids
potential negative interactions between the two drugs and has been
explored with EGFR-TKIs and chemotherapy (29,30).
Gemcitabine is a pyrimidine nucleoside
antimetabolite agent with a favorable toxicity profile, which is
active against a variety of human malignancies, including NSCLC
(31), and has been frequently used
in combinatorial treatments with other anticancer agents.
In the present study, we used NSCLC cells harboring
EGFR and K-Ras mutations to investigate the effect of sorafenib and
gemcitabine as single agents and in different sequences on
proliferation and cell cycle progression in vitro. We also
evaluated the molecular mechanisms of the different effects.
Materials and methods
Drugs
Sorafenib (BAY 43-9006) was obtained from Bayer
(Leverkusen, Germany) and was dissolved in dimethyl sulfoxide
(DMSO) to a stock concentration of 10 mmol/l. Gemcitabine was
purchased as a commercial product from the pharmacy at The Third
Affiliated Hospital of Anhui Medical University, Hefei, China, and
was dissolved in DMSO at 100 mmol/l, as stock solution. The drugs
were stored at −20°C and diluted with culture medium prior to
use.
Cell lines
The EGFR-TKI-sensitive PC-9 (mutant EGFR/wild-type
K-Ras) and EGFR-TKI-resistant A549 (wild-type EGFR/mutant K-Ras)
human NSCLC cell lines were purchased from American Type Culture
Collection (ATCC, Manassas, VA, USA) and maintained in RPMI-1640
medium (Hyclone, Logan, UT, USA), supplemented with 10%
heat-inactivated fetal bovine serum (Hyclone), penicillin (100
U/ml), streptomycin (100 μg/ml) and L-glutamine (2 mM) at
37°C in a 5% CO2 atmosphere, and then harvested with
trypsin-EDTA when the cells reached exponential growth.
Anti-proliferative effects of single
agents
The anti-proliferative effects of sorafenib and
gemcitabine as single agents on A549 and PC-9 cells were evaluated
by MTT assay, as previously described (32). Cells were cultured in 96-well
plates, in which the number of A549 and PC-9 cells was 4,000 and
6,000 per well, respectively. The IC50 value, indicating the
concentration resulting in inhibition of 50% of the maximal cell
growth, was determined following 72 h exposure to the drug compared
with unexposed control cells. After cells were exposed to each drug
for 72 h in 96-well plates, 20 ml MTT solution was added to each
well. The optical density (OD) of each well was measured at 490 nm
following incubation for 4 h. The percentage of cell growth
inhibition resulting from each drug was calculated as: [(OD 490
control cells – OD 490 treated cells)/OD 490 control cells] × 100.
This assay was repeated in more than three independent
experiments.
Anti-proliferative effects of different
sequences of sorafenib and gemcitabine
The anti-proliferative effects of three different
sequences of sorafenib and gemcitabine were evaluated. In the first
schedule, cells were pretreated with gemcitabine for 24 h, followed
by a washout with phosphate-buffered saline (PBS) and an additional
exposure to sorafenib for 72 h. In the second schedule, the reverse
sequence of sorafenib followed by gemcitabine was performed.
Thirdly, cells were concurrently treated with sorafenib and
gemcitabine for 72 h and incubated in a drug-free medium for 24 h.
The combination drug doses using constant ratios of the IC50 values
were calculated from the previous cytotoxicity tests. Thus, the
combination index (CI) value was calculated using 0.125, 0.25, 0.5,
1, 2 and 4 times (A549) or 0.296, 0.444, 0.667, 1, 1.5 and 2.25
times (PC-9) the IC50 of sorafenib and gemcitabine combination
doses. The CI values of interactions between sorafenib and
gemcitabine were analyzed according to the Chou and Talaly method
using CompuSyn software (ComboSyn, Inc., Paramus, NJ, USA):
CI>1, CI=1 and CI<1 indicate antagonistic, additive and
synergistic effects, respectively (33).
Cell cycle analysis of single agents and
of different sequences of sorafenib and gemcitabine
Cells (1×105/well) were plated into
six-well plates and exposed to sorafenib and gemcitabine as single
agents and in different sequences at the concentration of IC50
levels for the interval as described above. At the end of each
exposure, cells were collected and fixed with 70% cold ethanol at
4°C overnight. DNA staining was performed using a solution with
propidium iodide (0.05 mg/ml) and RNase (2 mg/ml) for 30 min at
room temperature. Cells were analyzed using a FACScan cytometer and
the percentage of cells in G1, S and G2/M phases of the cell cycle
was estimated by Cell Lab Quanta SC Software.
Western blot analysis
Cells (5×105/well) were treated with
sorafenib and gemcitabine as single agents and in different
sequences for the desired time. Cells were washed with ice-cold PBS
solution and scraped in lysis buffer. The lysates were centrifuged
at 14,000 rpm for 30 min at 4°C and the supernatant was collected.
Equivalent amounts of protein were analyzed by sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and
transferred to PVDF membranes. Appropriate primary antibodies to
pPDGFRβ, PDGFRβ, pAKT, AKT, pERK1/2, ERK1/2, Bcl-2 and β-actin
purchased from Cell Signaling Technology (Beverly, MA, USA) were
used. Proteins were visualized with a horseradish
peroxidase-coupled secondary antibody from Cell Signaling
Technology. Specific bands were detected using the enhanced
chemiluminescence reagent (ECL; PerkinElmer Life Sciences, Inc.,
Boston, MA, USA) on autoradiographic film and quantitated by
densitometry.
Statistical analysis
The results obtained from at least three independent
experiments are expressed as mean ± standard deviation (SD).
Student’s t-test and one-way ANOVA test were used to determine the
differences between control and treatment groups. P<0.05 was
considered to indicate a statistically significant result.
Results
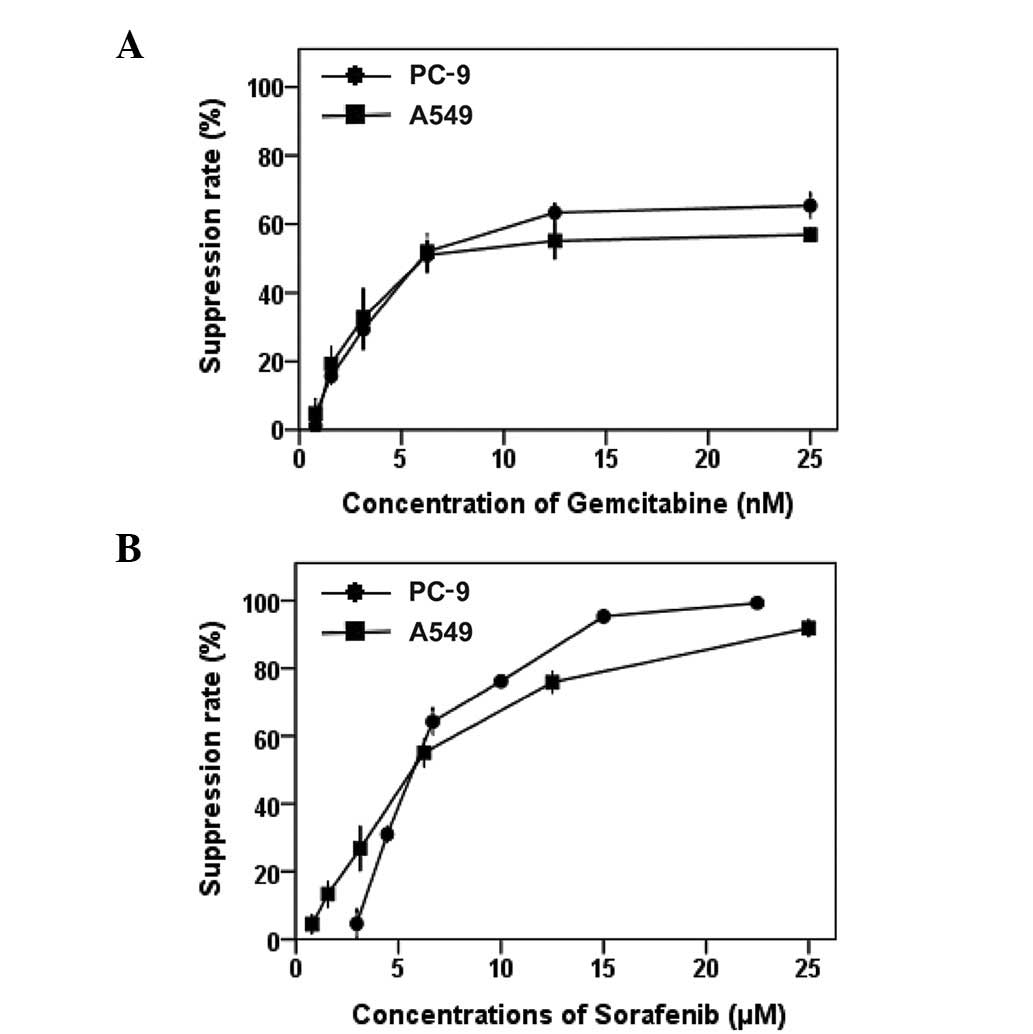
Dose-dependent anti-proliferative
activity of sorafenib and gemcitabine
MTT assays were used to evaluate the
anti-proliferative effects of sorafenib and gemcitabine as single
agents on EGFR-TKI-sensitive PC-9 (mutant EGFR/wild-type K-Ras) and
EGFR-TKI-resistant A549 (wild-type EGFR/mutant K-Ras) NSCLC cell
lines. Dose-dependent growth inhibitory effects of sorafenib
(0.78–25 μM) and gemcitabine (0.78–25 nM) were observed in
the two NSCLC cell lines (Fig. 1).
We demonstrated that the sensitivity of PC-9 and A549 cells to
sorafenib or gemcitabine are similar. Table I summarizes the IC50 of the two
drugs. The IC50 values of sorafenib in the two cell lines are
within the clinically relevant concentration range for this drug
(8.5–15.7 μmol/l) (34).
 | Table I.IC50 values of sorafenib and
gemcitabine were determined by MTT. |
Table I.
IC50 values of sorafenib and
gemcitabine were determined by MTT.
| IC50 | A549 | PC-9 |
|---|
| Gemcitabine | 10.38±0.80 nM | 8.38±0.64 nM |
| Sorafenib | 5.91±0.22
μM | 6.13±0.14
μM |
Schedule-dependent anti-proliferative
activity of sorafenib and gemcitabine
We evaluated the anti-proliferative effects of
sorafenib and gemcitabine in three different sequences on A549 and
PC-9 cell lines. As shown in Figs.
2 and 3, the anti-proliferative
effects observed in A549 cells following the administration of
gemcitabine followed by sorafenib were more noticeable than the
reversed sequence of sorafenib followed by gemcitabine (P<0.05)
and the concurrent administration of the two drugs (P<0.05).
Similar results were also found in PC-9 cells. In the A549 and PC-9
cell lines, the calculation of CI values revealed that the sequence
of gemcitabine followed by sorafenib produced synergistic effects
(Fig. 4), with mean CI values of
0.67 in A549 and 0.76 in PC-9 cells. Concomitant administration of
the drugs resulted in synergistic effects, with mean CI values of
0.9 in A549 and 0.95 in PC-9 cells. The sorafenib followed by
gemcitabine sequence resulted in an antagonistic interaction with
mean CI values of 1.31 in A549 and 1.45 in PC-9 cells. Therefore,
regardless of the mutation status of EGFR or K-Ras in NSCLC cells,
exposure to gemcitabine followed by sorafenib was shown to exert
synergistic effects, whereas the effect of the reversed sequence is
antagonistic. These results illustrate that the sequential
administration of gemcitabine followed by sorafenib is superior to
sorafenib followed by gemcitabine and concurrent
administration.
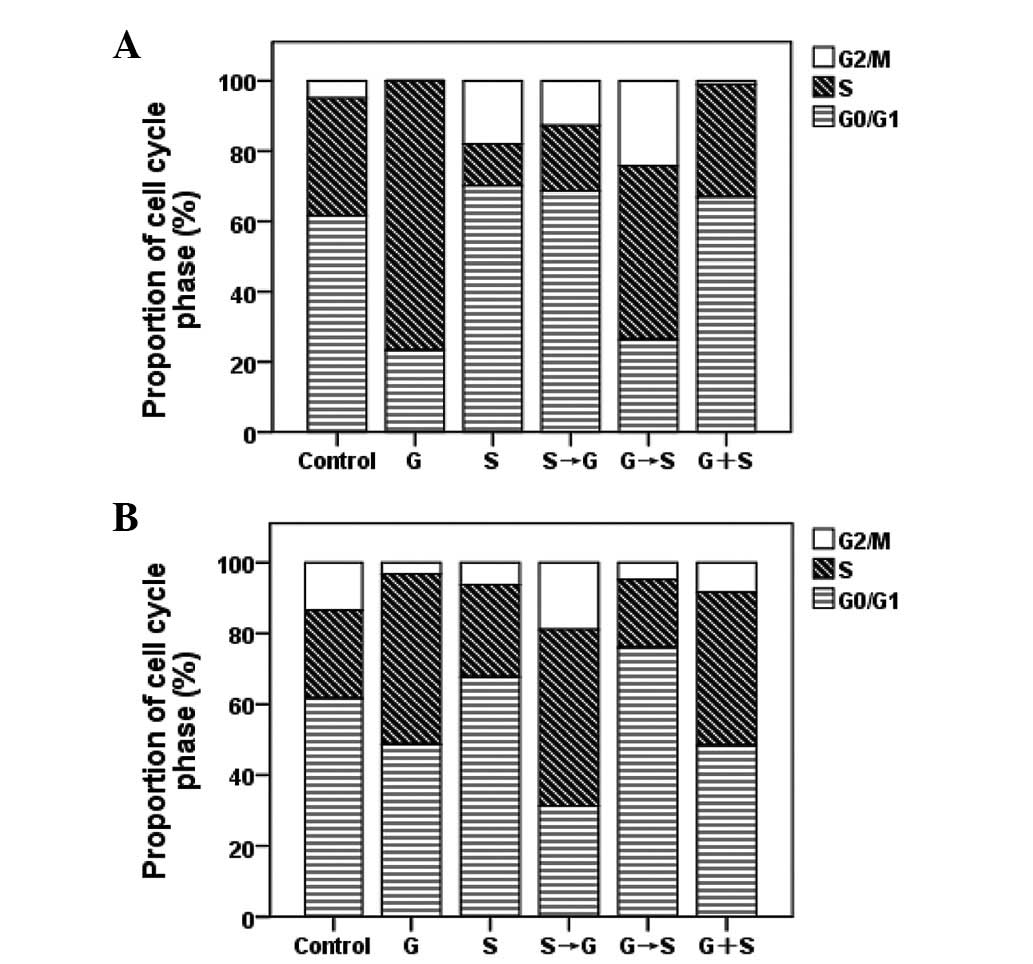
Cell cycle effects of sorafenib and
gemcitabine
Flow cytometry was applied to evaluate the cell
cycle phase distributions in EGFR-TKI-sensitive and
EGFR-TKI-resistant cells following single-drug, sequential and
concurrent administration of gemcitabine and sorafenib (Fig. 5). Following sorafenib treatment, the
proportion of A549 and PC-9 cells in G0/G1 phase increased relative
to control cells (P<0.05). Following treatment with gemcitabine
alone, the fraction of A549 and PC-9 cells in S phase increased
(P<0.05). Treatment with gemcitabine followed by sorafenib
resulted in an increase in cells in the S and G2/M phases
(P<0.05). By contrast, when cells were exposed to the reversed
sequence, the proportion of cells in the S phase decreased
(P<0.05).
Gemcitabine-mediated activation of
downstream signaling pathways
To further evaluate the potential synergistic
mechanisms of gemcitabine and sorafenib, the effects of gemcitabine
on the downstream AKT and ERK signaling pathways and the
anti-apoptotic Bcl-2 protein were detected by western blot analysis
in PC-9 and A549 cells. The MEK/ERK and PI3K/AKT pathways are
critical for proliferation and survival. In the two cell lines, we
found that the level of p-AKT gradually increased from 0 to 24 h
and lasted for 72 h when cells were exposed to gemcitabine at three
times the IC50 concentration. Similarly, we observed an increase in
p-ERK level in the two cell lines induced by gemcitabine alone. We
also observed gemcitabine induced an increase in the Bcl-2 level in
PC-9 and A549 cells. The effects of exposure to gemcitabine were
not significantly different in EGFR-mutated and K-Ras-mutated cells
(Fig. 6).
Enhanced anti-proliferative effects of
the schedule of gemcitabine followed by sorafenib
The effects of sorafenib and gemcitabine as single
agents and in different exposure schedules on cell signaling
pathways in A549 and PC-9 cells were evaluated. As shown in
Fig. 6, after 24 h exposure to
gemcitabine at IC50 concentration, the levels of p-AKT were
upregulated, but p-ERK did not change significantly, compared with
Fig. 6. This result implies that
gemcitabine increased the levels of p-ERK at a certain
concentration and exposure time. We found that sorafenib
downregulated the levels of p-PDGFRβ, p-AKT, p-ERK and Bcl-2 in
A549 and PC-9 cells compared with unex-posed cells. We also found
that the decreased Bcl-2 level in PC-9 cells was more significant
than in the A549 cells. When the two cell lines were exposed to the
sequence of gemcitabine followed by sorafenib, the levels of
p-PDGFRβ, p-AKT, p-ERK and Bcl-2 were downregulated compared with
sorafenib followed by gemcitabine. Upregulation of p-PDGFRβ, p-AKT,
p-ERK and Bcl-2 expression levels was observed following the
exposure sequence of sorafenib followed by gemcitabine compared
with the reversed sequence (Fig.
7). However, compared with the control cells, there was no
significant variation in the total PDGFR, ERK and AKT
expression.
Discussion
Platinum-based chemotherapy has become the mainstay
treatment for advanced NSCLC (35).
Although traditional cytotoxic chemotherapy improves patient
survival, treatment options remain limited for patients with
advanced NSCLC. Recently, targeted anticancer drugs, including
EGFR-TKIs, have been approved for the treatment of lung cancer
(36). However, EGFR-TKIs, such as
gefitinib and erlotinib, have no effect in the majority of K-Ras
mutation NSCLC tumors (37). The
development of new treatment strategies for NSCLC patients is thus
an important clinical goal.
There is multilevel cross-stimulation among the
targets in lung cancer. When only one pathway is blocked, others
act as salvage or escape mechanisms for cancer cells. Anticancer
agents that interfere at different stages and avoid escape or
salvage mechanisms may be more effective than single targeted
agents (38). Therefore,
multi-targeted TKIs that block multiple signaling pathways are
considered to be more effective therapeutic agents for cancer.
Sorafenib is a novel, multi-kinase inhibitor that targets tumor
proliferation and tumor angiogenesis (20). It has been approved for the
treatment of advanced renal cell cancer and is currently being
evaluated for the treatment of other tumors (20).
The present study was performed in
EGFR-TKI-sensitive PC-9 (EGFR mutant/wild-type K-Ras) and
EGFR-TKI-resistant A549 (wild-type EGFR/mutant K-Ras) human lung
cancer cell lines to investigate the anti-proliferative effects of
sorafinib as a single agent and in different schedules in
combination with gemcitabine. We found that sorafenib and
gemcitabine exhibited dose-dependent growth inhibition of cell
growth as single agent treatment in PC-9 and A549 lung cancer
cells. This result suggests that sorafenib is efficacious for
growth inhibition in EGFR-mutated and K-Ras-mutated NSCLC cell
lines. The strongest synergism was observed upon administration of
gemcitabine followed by sorafenib in the two cell lines, whereas
antagonistic interactions were observed upon administration of
sorafenib followed by gemcitabine. Similar to our observations, a
previous in vivo study demonstrated that the administration
of gemcitabine followed by sorafenib had synergistic effects in
NSCLC cells (39). Therefore, this
sequential administration of sorafenib and gemcitabine may benefit
patients with K-Ras mutations.
In our study, A549 and PC-9 cells were arrested by
sorafenib at G1 phase, while gemcitabine caused cells to accumulate
in S phase. The synergism and antagonism effect may be explained by
these effects between the two drugs. A549 and PC-9 cells were first
arrested at the G1 phase by sorafenib, thereby the proportion of
cells in S phase decreased, resulting in weakened cell
cycle-specific cytotoxicity of gemcitabine. In the reverse
sequence, gemcitabine arrested cells in the S phase, inhibiting
cell mitosis and multiplication, then the subsequent sorafenib
suppressed the cell growth, thereby increasing the proportion of
apoptotic cells. Similar to our observation, previous studies
concerning antagonism between sorafenib and chemotherapy have
reported that cells were arrested firstly by sorafenib at G1 phase,
interfering with the cytotoxicity effects in S phase of the cell
cycle-specific drugs (40,41).
The difference in the sequence-dependent
anti-proliferative effects of sorafenib and gemcitabine may also
result from growth signaling pathways. We found that gemcitabine
enhanced the expression levels of molecules in downstream signaling
pathways, for example, increasing the levels of p-AKT and p-ERK in
A549 and PC-9 cells. Similar to our results, a previous study
reported that cell signaling pathways may be gradually activated by
chemotherapy (42). p-ERK and AKT
play important roles in tumor cell proliferation, but gemcitabine
induced ERK and AKT phosphorylation, leading to the prevention of
apoptosis.
We have shown that sorafenib inhibited the activity
of the upstream receptor PDGFRβ and decreased the levels of the
downstream p-AKT, p-ERK and Bcl-2 in the A549 and PC-9 cell lines
after 72 h exposure. Sorafenib inhibited the PDGFRβ-dependent
activation of the PI3K/AKT and MAPK pathways, thereby decreasing
the levels of p-AKT and p-ERK (43). Sorafenib decreased the activity of
C-RAF and B-RAF in the RAF/MEK/ERK signaling pathway, meaning that
the level of p-ERK may also be downregulated directly. However, a
previous study has reported that sorafenib failed to inhibit p-ERK
in NSCLC cell lines with K-Ras mutations (28). The conflicting results may be
attributed to a shorter exposure to sorafenib. It means that
prolonging the exposure time of sorafenib may decreased the level
of p-ERK.
Sorafenib inhibited the expression of Bcl-2 in the
two cell lines, mainly by the simultaneous inhibition of the ERK
and AKT downstream pathways. We also found that the decreased Bcl-2
level in PC-9 cells was more significant than in the A549 cells. We
conclude that sorafenib has stronger cytotoxicity effects in lung
cancer cells with K-Ras mutation.
We also found that the expression of p-ERK and p-AKT
differed in response to each combinatorial treatment. When
gemcitabine was administered first, a significant decrease in p-AKT
and p-ERK levels was observed in the A549 and PC-9 cell lines.
Conversely, when sorafenib was administered first, the levels of
p-AKT and p-ERK were decreased and then upregulated by subsequent
exposure to gemcitabine. The sorafenib-induced decrease in p-ERK
and p-AKT appears to be reversible. These observations of p-AKT and
p-ERK in NSCLC cells may explain the synergistic and antagonist
growth inhibitory effects observed in A549 and PC-9 cells treated
with sorafenib and gemcitabine.
In conclusion, we found that sorafenib exhibited
significant growth inhibition in EGFR-TKI-sensitive and
EGFR-TKI-resistant NSCLC cells. Moreover, regardless of the
mutation status of EGFR and K-Ras, the sequential administration of
gemcitabine followed by sorafenib was an optimum schedule against
NSCLC. These data encourage the development of sorafenib as a
single targeted therapy or in combination with cytotoxic
chemotherapy drugs for treatment of NSCLC. Our study used an in
vitro model and was unable to test the antiangiogenic effects
of sorafenib, so further in vivo studies are required to
explore sorafenib as a single agent and the schedule-dependent
administration of sorafenib plus gemcitabine in NSCLC.
Acknowledgements
This study was supported by a grant
from the Anhui Provincial Science and Technology Agency Foundation
of China (No. 09020303042) and supported by The Central Laboratory
of The Third Affiliated Hospital of Anhui Medical University.
References
|
1.
|
Jemal A, Siegel R, Xu J and Ward E: Cancer
statistics, 2010. Cancer J Clin. 60:277–300. 2010. View Article : Google Scholar
|
|
2.
|
Mok TS, Wu YL, Thongprasert S, et al:
Gefitinib or carboplatin-paclitaxel in pulmonary adenocarcinoma. N
Engl J Med. 361:947–957. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Maemondo M, Inoue A, Kobayashi K, et al:
Gefitinib or chemotherapy for non-small-cell lung cancer with
mutated EGFR. N Engl J Med. 362:2380–2388. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Masago K, Fujita S, Togashi Y, et al:
Clinicopathologic factors affecting the progression-free survival
of patients with advanced non-small-cell lung cancer after
gefitinib therapy. Clin Lung Cancer. 12:56–61. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Pao W, Wang TY, Riely GJ, et al: KRAS
mutations and primary resistance of lung adenocarcinomas to
gefitinib or erlotinib. PLoS Med. 2:e172005. View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Gazdar AF: Activating and resistance
mutations of EGFR in non-small-cell lung cancer: role in clinical
response to EGFR tyrosine kinase inhibitors. Oncogene. 28:S24–S31.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Rodenhuis S, Slebos RJ, Boot AJ, et al:
Incidence and possible clinical significance of K-ras oncogene
activation in adenocarcinoma of the human lung. Cancer Res.
48:5738–5741. 1988.PubMed/NCBI
|
|
8.
|
Mitsudomi T, Kosaka T, Endoh H, et al:
Mutations of the epidermal growth factor receptor gene predict
prolonged survival after gefitinib treatment in patients with
non-small-cell lung cancer with postoperative recurrence. J Clin
Oncol. 23:2513–2520. 2005. View Article : Google Scholar
|
|
9.
|
Kosaka T, Yatabe Y, Endoh H, et al:
Analysis of epidermal growth factor receptor gene mutation in
patients with non-small cell lung cancer and acquired resistance to
gefitinib. Clin Cancer Res. 12:5764–5769. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Toi M, Matsumoto T and Bando H: Vascular
endothelial growth factor:its prognostic, predictive, and
therapeutic implications. Lancet Oncol. 2:667–673. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
Alvarez RH, Kantarjian HM and Cortes JE:
Biology of platelet-derived growth factor and its involvement in
disease. Mayo Clinic Proceedings. 81:1241–1257. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Ferrara N, Gerber HP and LeCouter J: The
biology of VEGF and its receptors. Nat Med. 9:669–676. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Wu E, Palmer N, Tian Z, et al:
Comprehensive dissection of PDGF-PDGFR signaling pathways in PDGFR
genetically defined cells. PLoS One. 3:e37942008. View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Erber R, Thurnher A, Katsen AD, et al:
Combined inhibition of VEGF and PDGF signaling enforces tumor
vessel regression by interfering with pericyte-mediated endothelial
cell survival mechanisms. FASEB J. 18:338–340. 2004.PubMed/NCBI
|
|
15.
|
Shikada Y, Yonemitsu Y, Koga T, et al:
Platelet-derived growth factor-AA is an essential and autocrine
regulator of vascular endothelial growth factor expression in
non-small cell lung carcinomas. Cancer Res. 65:7241–7248. 2005.
View Article : Google Scholar
|
|
16.
|
Donnem T, Al-Saad S, Al-Shibli K, Busund
LT and Bremnes RM: Co-expression of PDGF-B and VEGFR-3 strongly
correlates with lymph node metastasis and poor survival in
non-small-cell lung cancer. Ann Oncol. 21:223–231. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Linardou H, Dahabreh IJ, Kanaloupiti D, et
al: Assessment of somatic k-Ras mutations as a mechanism associated
with resistance to EGFR-targeted agents: a systematic review and
meta-analysis of studies in advanced non-small cell lung cancer and
colorectal cancer. Lancet Oncol. 9:962–972. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Rodenhuis S, van de Wetering ML, Mooi WJ,
Evers SG, van Zandwijk N and Bos JL: Mutational activation of the
K-ras oncogene. A possible pathogenetic factor in adenocarcinoma of
the lung. N Engl J Med. 317:929–935. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Lopez-Chavez A, Carter CA and Giaccone G:
The role of KRAS mutations in resistance to EGFR inhibition in the
treatment of cancer. Curr Opin Investig Drugs. 10:1305–1314.
2009.PubMed/NCBI
|
|
20.
|
Wilhelm SM, Carter C, Tang L, et al: BAY
43-9006 exhibits broad spectrum oral antitumor activity and targets
the RAF/MEK/ERK pathway and receptor tyrosine kinases involved in
tumor progression and angiogenesis. Cancer Res. 64:7099–7109. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Dal Lago L, D’Hondt V and Awada A:
Selected combination therapy with sorafenib: a review of clinical
data and perspectives in advanced solid tumors. Oncologist.
13:845–858. 2008.PubMed/NCBI
|
|
22.
|
Blumenschein GR Jr, Gatzemeier U, Fossella
F, et al: Phase II, multicenter, uncontrolled trial of single-agent
sorafenib in patients with relapsed or refractory, advanced
non-small-cell lung cancer. J Clin Oncol. 27:4274–4280. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Dy GK, Hillman SL, Rowland KM Jr, et al
North Central Cancer Treatment Group Study N0326: A front-line
window of opportunity phase 2 study of sorafenib in patients with
advanced nonsmall cell lung cancer: North Central Cancer Treatment
Group Study N0326. Cancer. 116:5686–5693. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24.
|
Carter CA, Chen C, Brink C, et al:
Sorafenib is efficacious and tolerated in combination with
cytotoxic or cytostatic agents in preclinical models of human
non-small cell lung carcinoma. Cancer Chemother Pharmacol.
59:183–195. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25.
|
Scagliotti G, Novello S, von Pawel J, et
al: Phase III study of carboplatin and paclitaxel alone or with
sorafenib in advanced non-small cell lung cancer. J Clin Oncol.
28:1835–1842. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26.
|
Paz-Ares LG, Biesma B, Heigener D, et al:
Phase III, randomized, double-blind, placebo-controlled trial of
gemcitabine/cisplatin alone or with sorafenib for the first-line
treatment of advanced, nonsquamous non-small-cell lung cancer. J
Clin Oncol. 30:3084–3092. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27.
|
Plastaras JP, Kim SH, Liu YY, et al: Cell
cycle dependent and schedule-dependent antitumor effects of
sorafenib combined with radiation. Cancer Res. 67:9443–9454. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
28.
|
Takezawa K, Okamoto I, Yonesaka K,
Hatashita E, Yamada Y, Fukuoka M and Nakagawa K: Sorafenib inhibits
non-small cell lung cancer cell growth by targeting B-RAF in KRAS
wild-type cells and C-RAF in KRAS mutant cells. Cancer Res.
69:6515–6521. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29.
|
Mahaffey CM, Davies AM, Lara PN Jr, et al:
Schedule-dependent apoptosis in K-ras mutant non-small-cell lung
cancer cell lines treated with docetaxel and erlotinib: rationale
for pharmacodynamic separation. Clin Lung Cancer. 8:548–553. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
30.
|
Cheng H, An SJ, Zhang XC, et al: In vitro
sequence-dependent synergism between paclitaxel and gefitinib in
human lung cancer cell lines. Cancer Chemother Pharmacol.
67:637–646. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31.
|
Dougherty DW and Friedberg JW: Gemcitabine
and other new cytotoxic drugs: will any find their way into primary
therapy? Curr Hematol Malig Rep. 5:148–156. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32.
|
Mosmann T: Rapid colorimetric assay for
cellular growth and survival: application to proliferation and
cytotoxicity assays. J Immunol Methods. 65:55–63. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
33.
|
Chou TC and Talalay P: Quantitative
analysis of dose-effect relationships: the combined effects of
multiple drugs or enzyme inhibitors. Adv Enzyme Regul. 22:27–55.
1984. View Article : Google Scholar : PubMed/NCBI
|
|
34.
|
Strumberg D, Clark JW, Awada A, et al:
Safety, pharmacokinetics, and preliminary antitumor activity of
sorafenib: a review of four phase I trials in patients with
advanced refractory solid tumors. Oncologist. 12:426–437. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
35.
|
Rajeswaran A, Trojan A, Burnand B and
Giannelli M: Efficacy and side effects of cisplatin- and
carboplatin-based doublet chemotherapeutic regimens versus
non-platinum-based doublet chemotherapeutic regimens as first line
treatment of meta-static non-small cell lung carcinoma: a
systematic review of randomized controlled trials. Lung Cancer.
59:1–11. 2008.
|
|
36.
|
Maemondo M: Timing the change of
chemotherapy for non-small cell lung cancer. Gan To Kagaku Ryoho.
39:1316–1319. 2012.(in Japanese).
|
|
37.
|
Ayoola A, Barochia A, Belani K and Belani
CP: Primary and acquired resistance to epidermal growth factor
receptor tyrosine kinase inhibitors in non-small cell lung cancer:
an update. Cancer Invest. 30:433–446. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38.
|
Petrelli A and Giordano S: From single- to
multi-target drugs in cancer therapy: when aspecificity becomes an
advantage. Curr Med Chem. 15:422–432. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39.
|
Pasqualetti G, Ricciardi S, Mey V, Del
Tacca M and Danesi R: Synergistic cytotoxicity, inhibition of
signal transduction pathways and pharmacogenetics of sorafenib and
gemcitabine in human NSCLC cell lines. Lung Cancer. 74:197–205.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
40.
|
Zhang XH, Shin JY, Kim JO, Oh JE, Yoon SA,
Jung CK and Kang JH: Synergistic antitumor efficacy of sequentially
combined paclitaxel with sorafenib in vitro and in vivo NSCLC
models harboring KRAS or BRAF mutations. Cancer Lett. 322:213–222.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
41.
|
Ulivi P, Arienti C, Zoli W, et al: In
vitro and in vivo antitumor efficacy of docetaxel and sorafenib
combination in human pancreatic cancer cells. Curr Cancer Drug
Targets. 10:600–610. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42.
|
Torres K and Horwitz SB: Mechanisms of
taxol-induced cell death are concentration dependent. Cancer Res.
58:3620–3626. 1998.PubMed/NCBI
|
|
43.
|
Li QL, Gu FM, Wang Z, et al: Activation of
PI3K/AKT and MAPK pathway through a PDGFRβ-dependent feedback loop
is involved in rapamycin resistance in hepatocellular carcinoma.
PLoS One. 7:e333792012.
|