For many years, angiogenesis via the sprouting of
new vessels from existing ones was considered to be the exclusive
method of tumor vascularization (1), and anti-angiogenesis therapies were
applied as a promising method to ‘starve’ tumors. However, with the
administration of angiogenesis inhibitors primarily targeting
endothelial cells, it was identified that the effect of these types
of drugs was limited. This indicated that there may be other
supplementary blood supply patterns used to nourish tumors. In
1999, Maniotis et al(2)
first reported that highly aggressive and metastatic melanoma cells
are able to form highly patterned vascular channels lined
externally by tumor cells, without the existence of endothelial
cells. This process was termed vasculogenic mimicry (VM), which is
independent of angiogenesis, and is composed of tumor cells and a
basement membrane. VM was categorized into two distinctive types:
the patterned matrix type (2) and
the tubular type (3). Blood plasma
and red blood cells are able to flow in the nonendothelial
cell-lined vessel-like structures (2,4), and a
VM-angiogenesis junction in the central area of the inflammatory
breast cancer (IBC) xenografts has been observed (5). This evidence suggests that VM in the
tumor mass is connected with host vessels for blood supply and is
part of the functional microcirculation.
Subsequently, VM has been observed in numerous types
of aggressive tumors, such as colorectal cancer (6), head and neck squamous cell carcinoma
(HNSCC) (7–9), glioblastoma (3,10),
breast cancer (11,12), ovarian carcinoma (13,14),
astrocytoma (15) and Ewing sarcoma
(4). Increasing evidence has
suggested that patients with tumors undergoing VM have a worse
prognosis and VM may be used as an independent predictor of
prognosis (16–18). Lin et al(9) analyzed the clinical and pathological
significance of VM in 168 cases of laryngeal squamous cell
carcinoma (LSCC) and found that VM occurred in LSCC, and LSCC with
VM had increased potential for invasion and metastasis. Upile et
al(8) showed that HN2b
metastatic HNSCC cells lines have higher VM formation properties
when compared with the HN2a primary tumor cell line, and
endothelial growth factor antibodies discouraged VM formation.
Additionally, certain studies demonstrated that
administration of angiogenesis inhibitors did not suppress the
formation of VM, and even induced extracellular matrix-rich tubular
network formation in vitro(19). Conceivably, VM may play a pivotal
role as an alternative pathway for blood supply when the pattern of
angiogenesis is inhibited.
Epithelial-mesenchymal transition (EMT) is a
reversible dedifferentiation process that converts epithelial
cancer cells into dedifferentiated cells with additional
mesenchymal features. This process is characterized by the loss of
epithelial traits and the acquisition of mesenchymal phenotypes
(20–23). Activation of EMT triggers tumor cell
invasion and metastasis to distant organs. Recently, EMT has been
reported to contribute to the formation of VM, and the upregulation
of EMT-associated transcription factors has been demonstrated in
VM-forming tumor cells (24,25).
Normal tissues and tumors contain a small subset of
cells, known as stem cells, with the capacity for self-renewal and
the multipotency to differentiate into diverse committed lineages
(26,27). Tumors are composed of diverse types
of cells (28) and cancer stem
cells (CSCs) are at the top of the hierarchical pyramid (26,29–31).
Mounting evidence demonstrates that CSCs have the capacity for
differentiation along tumor and endothelial lineages (32,33),
as well as vascular smooth muscle-like cells (3). VM-engaging tumor cells show a
significant expression of both endothelial and tumor phenotypes
(14,34), and thus VM may represent the
incomplete progress of CSC differentiation into endothelial
lineages. Additionally, it has been observed that epithelial cancer
cells may be endowed with the self-renewal stem cell phenotype via
EMT (35,36). Therefore, in this review, we discuss
the correlation between CSCs, EMT and VM formation.
CSCs, as defined by the American Association of
Cancer Research, are a small subset of cells with the capability of
self-renewal and differentiation into the heterogeneous lineages
that constitute the tumor mass (26). In reality, this is only a functional
definition. Due to the lack of specific markers, the so-called
‘CSCs’ obtained in almost all experiments are actually a mixture of
real CSCs and progenitor cells. In that sense, it is also
reasonable to call these cells tumor stem-like cells. Although
there is controversy regarding the accurate definition of CSCs,
increasing evidence supports the existence of CSCs and the validity
of the CSC hypothesis (37). CSCs
were first demonstrated in human acute myeloid leukemia (AML) when
investigators found that the ability to initiate tumors by
transplantation of AML cells into NOD/SCID mice was limited to a
CD34+/CD38− subpopulation of leukemic cells
(38). CSCs have been further
observed in several solid tumors, such as breast (28), brain (39,40),
melanoma (41,42), prostate (43), ovarian (44,45)
and pancreatic cancers (46), as
well as HNSCC (47–53). In addition to the capability of
tumor initiation, CSCs have also been implicated in tumor invasion
and metastasis. In breast cancer, the CSCs sorted by a number of
markers have a higher capability of invasion and metastasis. Balic
et al(54) found that the
majority of early disseminated cancer cells in bone marrow have a
breast CSC phenotype (CD44+/CD24−). From a
retrospective study of 109 patients with IBC, the patient prognosis
and metastasis trends showed a significant correlation with
aldehyde dehydrogenase 1 (ALDH1) expression, a specific marker of
CSCs. Both in vitro and xenograft assays showed that
invasion and metastasis in IBC are mediated by a cellular component
that exhibits ALDH activity (55).
In HNSCC cell lines, based on an invasive assay in vitro and
injection of tumor cells into the tail vein of mice, Davis et
al(56) found that
CD44+ cells have an increased ability to invade through
the basement membrane and to form lung metastases. In the
peripheral blood of patients with HNSCC, a greater number of
CD44+ tumor cells were also observed compared with that
of the healthy control group (57).
Song et al(58) demonstrated
that side population (SP) cells in HNSCC were highly invasive, and
the highly metastatic M3a2 and M4e HNSCC cell lines contained a
greater number of SP cells in comparison with the 686LN parental
HNSCC cell line that has low metastatic potential. It was deduced
that SP cells may be a major driving force in head and neck tumor
formation and metastasis. Goldie et al(59) reported that upregulation of FRMD4A,
a human epidermal stem cell marker, occurs in primary human HNSCCs,
where high expression levels correlate with increased risk of
relapse. Additionally, FRMD4A silencing was shown to decrease the
growth and metastasis of human squamous cell carcinoma xenografts
in the skin and tongue.
With increasing knowledge of CSC phenotypes and
functions, the evidence suggests that CSCs are involved in VM
formation. In human breast cancer, by injecting human breast CSCs
into SCID mice, Bussolati et al(64) found that a number of the intratumor
vessels were of human origin, indicating the involvement of breast
CSCs in vessel formation. In melanoma, there is evidence showing
that the VM-forming tumor cells express phenotypes that are usually
expressed in other cell types, such as endothelial or epithelial
cells (11). This indicated that
these cells may revert to an undifferentiated, stem-like phenotype
(65). Recently, in glioblastoma,
Ricci-Vitiani et al(32) and
Wang et al(33) found that
CD133+ glioblastoma stem-like cells are pluripotent and
capable of differentiation along tumor and endothelial lineages
(33,66), as well as mixed endothelial cell
lineages, with co-expression of the tumor phenotype (32). On the analysis of tumor xenografts
obtained by orthotopic and subcutaneous injection of human
glioblastoma in immunocompromised mice, the authors observed that
the vessels in the transplanted tumor were primarily composed of
tumor cells with an aberrant endothelial phenotype. The findings
indicate that these cells are derived from CSCs, and thus VM may
represent an incomplete differentiation of cancer stem-like cells
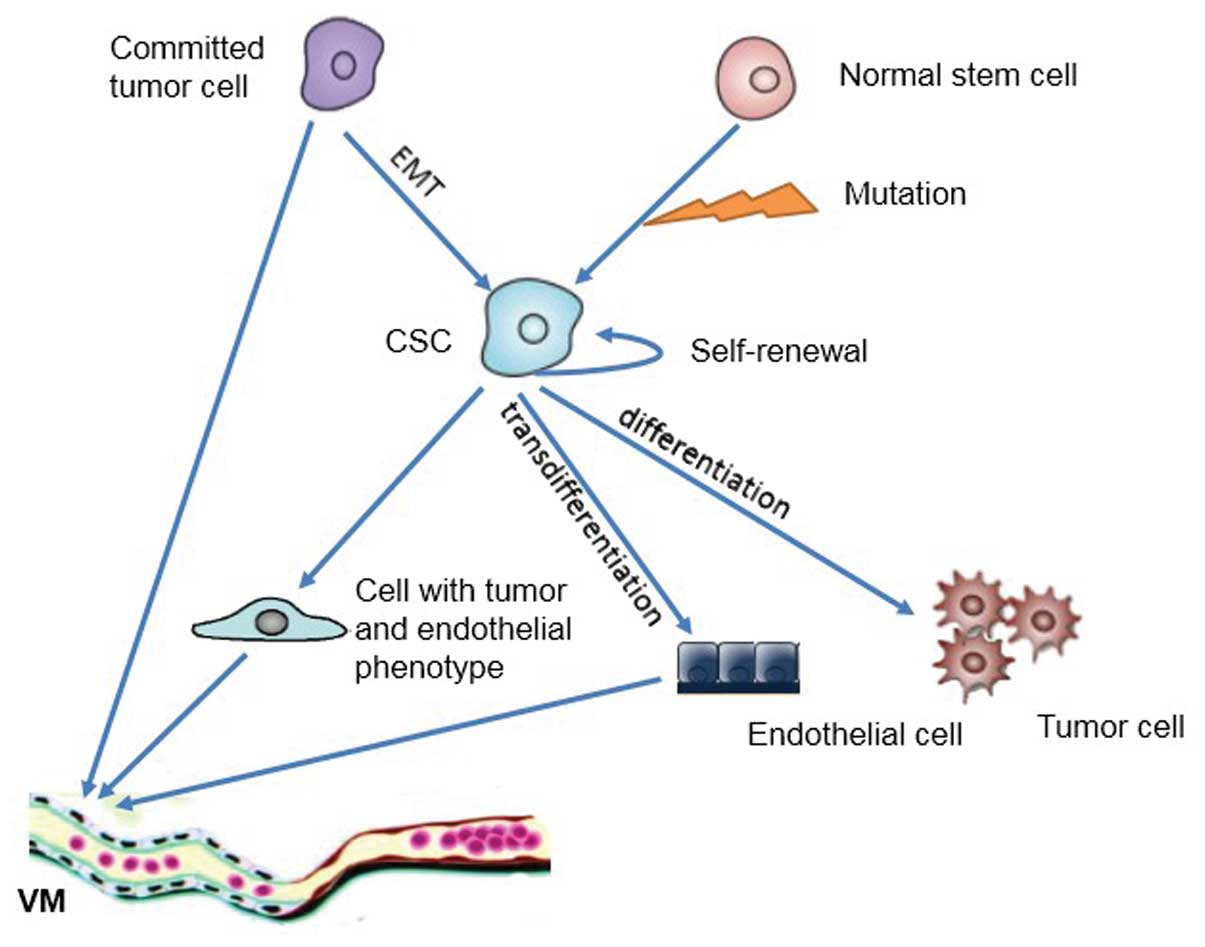
towards the endothelial lineage (Fig.
1). It has been reported that CD133+ and
ABCB5+ subpopulations are colocalized in melanomas in
perivascular niches that contain vascular endothelial
(VE)− cadherin+ melanoma cells, which have
the ability to form VM (67). Frank
et al(68) found that
vascular endothelial growth factor 1 (VEGF-1) signaling plays an
important role in this process and knockdown of VEGF receptor 1
blocked the development of ABCB5+ VM morphology. In oral
squamous cell carcinoma, Dang and Ramos (69) observed that
TRA-1-60+/β6+ tumor cells with CSC attributes
are able to form vascular-like structures in vivo. However,
based on the observation of the melanoma xenograft model, Zhang
et al(70) found that VM was
the dominant blood supply pattern in the early stage of tumor
growth. During tumor growth progression, the level of VM decreased
and the number of endothelial-dependent vessels increased. The
authors proposed a three-stage blood supply pattern consisting of
VM, mosaic vessels and endothelium-dependent vessels. This inverse
change tendency between VM and endothelium-dependent vessels may be
due to the persistent differentiation of CSCs to endothelial cells
without tumor cell phenotypes.
EMT is a crucial process in cancer progression,
providing cancer cells with the ability to escape from the primary
site, invade stromal tissues and migrate to distant regions of the
body. Epithelial cells undergoing EMT are characterized by
downregulation of epithelial makers (such as cytokeratin), loss of
cell polarity and intercellular adhesion molecules (for instance
E-cadherin and occludin), which is concomitant with upregulation of
mesenchymal markers (vimentin, N-cadherin and fibronectin) and
acquisition of fibroblast-like morphology with cytoskeleton
reorganization (20,22,71).
The loss of E-cadherin and the gain of N-cadherin expression are
known as cadherin switching, a major hallmark of EMT. Cadherin
switching was observed in 30 out of 80 HNSCC cases and was closely
correlated with histological differentiation, pattern of invasion
and lymph node metastasis in HNSCC cases (72). According to analysis of HNSCC
specimens and cell lines, Mandal et al(73) demonstrated a close correlation
between EMT and aggressive tumor features, including penetrating
invasive fronts, high-grade sarcomatoid transformation and lymph
node metastasis. A variety of transcription factors such as Snai1
(Snail1) (74–76), Slug (Snail2) (77), Twist (78), SOX4 (79) and ZEB (80), and several signaling pathways,
involving TGF-β, Wnt, Notch and Hedgehog, have been reported to
play significant roles in the process of EMT (81–83).
Snail and Slug repress E-cadherin transcription to degrade
cell-to-cell adhesion by binding the E-box in the E-cadherin
promoter, and inducing tumor cell migration (74,79,84).
The transcription factor Twist, a master regulator of embryonic
morphogenesis, contributes to metastasis of mammary carcinoma by
promoting an EMT (78). Twist1
induced invasion and metastasis of hepatocellular carcinoma (HCC)
via downregulation of E-cadherin and increased activity of matrix
metalloproteinase (MMP), specifically MMP2 and MMP9 (85). In a spontaneous skin squamous cell
carcinoma mouse model, Tsai et al(86) demonstrated that activation of Twist1
is sufficient to promote carcinoma cells to undergo EMT and
disseminate into blood circulation. Aigner et al(87) identified that the transcription
factor ZEB1 is able to induce the repression of certain polarity
genes (Crumbs3, PATJ and HUGL2) to improve tumor cell invasion.
Moreover, by virtue of repression of the miR-200 and miR-34a
families, respectively, ZEB1 contributes to metastasis by
maintenance of the dedifferentiation status and remodeling of
cytoskeletal actin (88,89). Furthermore, there is evidence to
suggest that these transcription factors function together, but not
independently, in order to induce EMT (84,90).
Recently, evidence has shown that EMT is involved in
the process of VM formation. In VM-positive colorectal carcinoma
samples, Liu et al(24)
found that expression of ZEB1 was upregulated. Downregulation of
E-cadherin and upregulation of vimentin in the ZEB1-positive group
were detected. Knockdown of ZEB1 resulted in a decrease in VM and
the restoration of certain epithelial phenotypes, such as
VE-cadherin and Flk-1. In HCC, the inhibition of Twist1 expression
by the short hairpin RNA markedly reduced VM formation (25). Furthermore, the Bcl-2/Twist1 complex
facilitates the nuclear transport of Twist1 and leads to
transcriptional activation of a wide range of genes that may
increase the tumor cell plasticity, metastasis and VM formation of
hepatocellular carcinoma (91).
Lirdprapamongkol et al(92)
have reported that the poorly-differentiated HCC cell line,
SK-Hep-1, with mesenchymal features (high invasiveness and
expressing vimentin, with no E-cadherin) could form VM in
vitro, while the well-differentiated cell line HepG2 did not
form VM. These findings indicated that EMT is involved in VM
formation.
Researchers have been engaged in discovering the
origin of CSCs for a number of years. It is widely accepted that
tumor formation is due to the multistep mutation of genomes.
Considering the longer lifespan of stem cells, normal stem cells
suffer from the accumulation of mutations over time. Thus, it is
hypothesized that CSCs derive from normal stem cells with genetic
mutations, and this has been demonstrated by independent
investigators (93,94). In addition to this mechanism,
mounting data suggest that differentiated tumor cells may reacquire
stemness (95), particularly via
EMT induction (35). The endowment
of stem cell traits by EMT provided another source for the origin
of CSCs. Biddle et al(96)
classified the CSCs into two types, namely, non-EMT CSCs and EMT
CSCs, based on EMT progression.
An increasing body of evidence shows that EMT is
associated with the acquisition of CSC properties. In 2008, Mani
et al(35) reported that the
induction of EMT in immortalized human mammary epithelial cells
results in the acquisition of mesenchymal traits, as well as the
expression of stem cell markers. Stem-like cells isolated either
from mouse or human mammary glands or mammary carcinomas express
EMT markers (35,97). Morel et al(36) also indicated that cells possessing
both stem and tumorigenic characteristics of ‘CSCs’ may be derived
from human mammary epithelial cells following the activation of the
Ras-MAPK pathway. The acquisition of these stem cell and
tumorigenic characteristics is driven by EMT induction. Santisteban
et al(98) found that breast
tumor cells undergoing EMT induced by CD8+ T cells
acquired certain characteristics of breast CSCs, including potent
tumorigenicity, resistance to conventional treatment, and the
ability to form spheroids. Fang et al(99) demonstrated that Twist2 is
overexpressed in breast cancer cells. Ectopic overexpression of
Twist2 results in the induction of EMT and increases the number of
CD44+/CD24− cells. Breast cancer cells
exposed to TGF-β and TNF-α lead to the generation of breast cancer
cells with stem-like characteristics by induction of EMT (100). In addition to breast cancer, Ryu
et al(101) identified that
the gastric CSC marker CD44 was significantly associated with the
protein expression of Snail-1, ZEB-1 and E-cadherin. In colorectal
cancer, EMT induced by brachyury increased the nanog expression and
endowed the colorectal cells with stem cell attributes (102). In HNSCC, EMT conferring to stem
cell phenotypes has also been observed. Xia et al(103) found that miR-200a regulates the
acquisition of stem-like traits by the induction of EMT in
nasopharyngeal carcinoma. Knockdown of miR-200a induced EMT
progression and resulted in stem cell attributes, including an
increasing proportion of SP, sphere formation capacity, in
vivo tumorigenicity in nude mice and stem cell marker
expression. Chen et al(104) revealed that
HNSCC-ALDH1+ cells exhibit a high level of expression of
Snail, and knockdown of Snail significantly decreased the
expression of ALDH1. These data suggest that epithelial cells
within tumors are able to convert into CSCs via EMT (Fig. 1). Moreover, Chen et
al(105) demonstrated that
upregulation of CD133 increased the phosphorylation of Src coupled
with EMT transformation, and CD133/Src signaling is a regulatory
switch resulting in EMT and stemness properties in HNSCC. This
knowledge provides an improved understanding of the origin of CSCs
and is a basis for novel cancer therapeutic strategies targeting
EMT and CSCs.
VM allows tumor cells to express the endothelial
phenotype and play a similar functional role to endothelial cells
in forming blood vessel-like structures. In fact, both epithelial
and mesenchymal markers have been observed in tumor cells engaged
in VM formation (14,106,107). Therefore, in view of the crucial
role of EMT in the acquisition of stemness, it is plausible that
CSCs are implicated in VM formation by induction of EMT (Fig. 1). Signal transducers and activators
of transcription 3 transcription factor plays a critical role in
the development and progression of a variety of tumors, including
HNSCC, by regulating cell proliferation, cell cycle progression,
apoptosis, angiogenesis, immune evasion and EMT, and through
effects in CSCs. Garnier et al(108) found that tissue factor
overexpression accompanies features of cellular aggressiveness,
such as markers of CSCs (CD133), EMT and expression of the
angiogenic and prometastatic phenotype. Recently, Gill et al
demonstrated that Snail promotes the induction of Flk1+
endothelial cells in an early subset of differentiating mouse
embryonic stem cells, depending on fibroblast growth factor
signaling as well as the repression of the miR-200 family (109). Hypoxia is one of the fundamental
changes in the development and aggressiveness of a variety of solid
tumors. It has been recognized to play critical roles in tumor
invasion, metastasis, angiogenesis and chemo-radiation resistance.
In addition to tumor angiogenesis, HIF-1a is closely associated
with VM formation (4,110–112). Recently, Misra et
al(113) found that
hypoxia-exposure resulted in an upregulation of c-Myc and OCT3/4,
and contributed to VM formation. Hypoxia was also recognized as an
important regulator of CSCs and EMT through NF-κB, PI3K/Akt/mTOR,
Notch, Wnt/β-catenin and Hedgehog signaling pathways (114,115). Thus, the hypoxia microenvironment
may be important in VM formation through stemness maintenance and
EMT induction.
In reality, the coexistence of angiogenesis and VM
is common within aggressive tumors. Angiogenesis inhibitors have
little or even no effect on VM (10,19)
and VM may replace the effect of angiogenesis to provide the tumor
with oxygen and nutrients. Moreover, Qu et al(119) reported that anti-angiogenesis
therapy may even induce the formation of VM. Clearly, the
combination of several treatments targeting angiogenesis and VM is
required.
For quite some time, the survival rate of patients
with aggressive tumors has remained at a low level, despite the
administration of surgery, chemotherapy and radiotherapy. The
existence of CSCs was thought to be an underlying cause. Although
CSCs comprise only a small proportion of tumor cell populations,
CSCs have high resistance to multiple chemotherapeutics and
ionizing radiation. Remaining CSCs are able to induce recurrence
following treatment with chemotherapy and radiotherapy.
Furthermore, it has been demonstrated that CSCs are implicated in
VM formation. In this context, CSCs have been considered as a
promising treatment target in cancer patients with VM. It has been
observed that tumors undergoing the process of EMT acquire
resistance to chemotherapy (120).
EMT is also involved in the acquisition of CSC properties (35,36,98),
and EMT-inducing CSCs have been considered as an important origin
of CSCs and another target of VM formation in cancer. A combination
of targeting EMT and CSCs may be beneficial for anti-VM formation
therapy, decreasing invasion and metastasis, and improving the
survival rate of patients.
This study was supported by the National Natural
Science Foundation of China (grant nos. 81072215, 81172580 and
81272961) and by the Fundamental Research Funds of the Central
Universities of China (2011).
|
1
|
Hillen F and Griffioen AW: Tumour
vascularization: sprouting angiogenesis and beyond. Cancer
Metastasis Rev. 26:489–502. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Maniotis AJ, Folberg R, Hess A, et al:
Vascular channel formation by human melanoma cells in vivo and in
vitro: vasculogenic mimicry. Am J Pathol. 155:739–752. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
El Hallani S, Boisselier B, Peglion F, et
al: A new alternative mechanism in glioblastoma vascularization:
tubular vasculogenic mimicry. Brain. 133:973–982. 2010.PubMed/NCBI
|
|
4
|
van der Schaft DW, Hillen F, Pauwels P, et
al: Tumor cell plasticity in Ewing sarcoma, an alternative
circulatory system stimulated by hypoxia. Cancer Res.
65:11520–11528. 2005.PubMed/NCBI
|
|
5
|
Shirakawa K, Kobayashi H, Heike Y, et al:
Hemodynamics in vasculogenic mimicry and angiogenesis of
inflammatory breast cancer xenograft. Cancer Res. 62:560–566.
2002.PubMed/NCBI
|
|
6
|
Ricci-Vitiani L, Lombardi DG, Pilozzi E,
et al: Identification and expansion of human
colon-cancer-initiating cells. Nature. 445:111–115. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang W, Lin P, Han C, Cai W, Zhao X and
Sun B: Vasculogenic mimicry contributes to lymph node metastasis of
laryngeal squamous cell carcinoma. J Exp Clin Cancer Res.
29:602010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Upile T, Jerjes W, Radhi H, et al:
Vascular mimicry in cultured head and neck tumour cell lines. Head
Neck Oncol. 3:552011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lin P, Wang W, Sun BC, et al: Vasculogenic
mimicry is a key prognostic factor for laryngeal squamous cell
carcinoma: a new pattern of blood supply. Chin Med J (Engl).
125:3445–3449. 2012.PubMed/NCBI
|
|
10
|
Francescone R, Scully S, Bentley B, et al:
Glioblastoma-derived tumor cells induce vasculogenic mimicry
through Flk-1 protein activation. J Biol Chem. 287:24821–24831.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shirakawa K, Wakasugi H, Heike Y, et al:
Vasculogenic mimicry and pseudo-comedo formation in breast cancer.
Int J Cancer. 99:821–828. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ponti D, Costa A, Zaffaroni N, et al:
Isolation and in vitro propagation of tumorigenic breast cancer
cells with stem/progenitor cell properties. Cancer Res.
65:5506–5511. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tang HS, Feng YJ and Yao LQ: Angiogenesis,
vasculogenesis, and vasculogenic mimicry in ovarian cancer. Int J
Gynecol Cancer. 19:605–610. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Su M, Feng YJ, Yao LQ, et al: Plasticity
of ovarian cancer cell SKOV3ip and vasculogenic mimicry in vivo.
Int J Gynecol Cancer. 18:476–486. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liu Z, Li Y, Zhao W, Ma Y and Yang X:
Demonstration of vasculogenic mimicry in astrocytomas and effects
of Endostar on U251 cells. Pathol Res Pract. 207:645–651. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Baeten CI, Hillen F, Pauwels P, de Bruine
AP and Baeten CG: Prognostic role of vasculogenic mimicry in
colorectal cancer. Dis Colon Rectum. 52:2028–2035. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Warso MA, Maniotis AJ, Chen X, et al:
Prognostic significance of periodic acid-Schiff-positive patterns
in primary cutaneous melanoma. Clin Cancer Res. 7:473–477.
2001.PubMed/NCBI
|
|
18
|
Folberg R, Rummelt V, Parys-Van
Ginderdeuren R, et al: The prognostic value of tumor blood vessel
morphology in primary uveal melanoma. Ophthalmology. 100:1389–1398.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
van der Schaft DW, Seftor RE, Seftor EA,
et al: Effects of angiogenesis inhibitors on vascular network
formation by human endothelial and melanoma cells. J Natl Cancer
Inst. 96:1473–1477. 2004.PubMed/NCBI
|
|
20
|
Kalluri R and Neilson EG:
Epithelial-mesenchymal transition and its implications for
fibrosis. J Clin Invest. 112:1776–1784. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Eccles SA and Welch DR: Metastasis: recent
discoveries and novel treatment strategies. Lancet. 369:1742–1757.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Thiery JP: Epithelial-mesenchymal
transitions in tumour progression. Nat Rev Cancer. 2:442–454. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu Z, Sun B, Qi L, Li H, Gao J and Leng
X: Zinc finger E-box binding homeobox 1 promotes vasculogenic
mimicry in colorectal cancer through induction of
epithelial-to-mesenchymal transition. Cancer Sci. 103:813–820.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sun T, Zhao N, Zhao XL, et al: Expression
and functional significance of Twist1 in hepatocellular carcinoma:
its role in vasculogenic mimicry. Hepatology. 51:545–556. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Clarke MF, Dick JE, Dirks PB, et al:
Cancer stem cells - perspectives on current status and future
directions: AACR Workshop on cancer stem cells. Cancer Res.
66:9339–9344. 2006. View Article : Google Scholar
|
|
27
|
Shipitsin M and Polyak K: The cancer stem
cell hypothesis: in search of definitions, markers, and relevance.
Lab Invest. 88:459–463. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Al-Hajj M, Wicha MS, Benito-Hernandez A,
Morrison SJ and Clarke MF: Prospective identification of
tumorigenic breast cancer cells. Proc Natl Acad Sci USA.
100:3983–3988. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
O’Brien CA, Pollett A, Gallinger S and
Dick JE: A human colon cancer cell capable of initiating tumour
growth in immunodeficient mice. Nature. 445:106–110.
2007.PubMed/NCBI
|
|
30
|
Rosen JM and Jordan CT: The increasing
complexity of the cancer stem cell paradigm. Science.
324:1670–1673. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Singh SK, Hawkins C, Clarke ID, et al:
Identification of human brain tumour initiating cells. Nature.
432:396–401. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ricci-Vitiani L, Pallini R, Biffoni M, et
al: Tumour vascularization via endothelial differentiation of
glioblastoma stem-like cells. Nature. 468:824–828. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang R, Chadalavada K, Wilshire J, et al:
Glioblastoma stem-like cells give rise to tumour endothelium.
Nature. 468:829–833. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Dong J, Zhao Y, Huang Q, et al: Glioma
stem/progenitor cells contribute to neovascularization via
transdifferentiation. Stem Cell Rev. 7:141–152. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Mani SA, Guo W, Liao MJ, et al: The
epithelial-mesenchymal transition generates cells with properties
of stem cells. Cell. 133:704–715. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Morel AP, Lièvre M, Thomas C, Hinkal G,
Ansieau S and Puisieux A: Generation of breast cancer stem cells
through epithelial-mesenchymal transition. PLoS One. 3:e28882008.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Chen J, Li Y, Yu TS, et al: A restricted
cell population propagates glioblastoma growth after chemotherapy.
Nature. 488:522–526. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lapidot T, Sirard C, Vormoor J, et al: A
cell initiating human acute myeloid leukaemia after transplantation
into SCID mice. Nature. 367:645–648. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hemmati HD, Nakano I, Lazareff JA, et al:
Cancerous stem cells can arise from pediatric brain tumors. Proc
Natl Acad Sci USA. 100:15178–15183. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Singh SK, Clarke ID, Terasaki M, et al:
Identification of a cancer stem cell in human brain tumors. Cancer
Res. 63:5821–5828. 2003.PubMed/NCBI
|
|
41
|
Fang D, Nguyen TK, Leishear K, et al: A
tumorigenic subpopulation with stem cell properties in melanomas.
Cancer Res. 65:9328–9337. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Schatton T, Murphy GF, Frank NY, et al:
Identification of cells initiating human melanomas. Nature.
451:345–349. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Collins AT, Berry PA, Hyde C, Stower MJ
and Maitland NJ: Prospective identification of tumorigenic prostate
cancer stem cells. Cancer Res. 65:10946–10951. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Bapat SA, Mali AM, Koppikar CB and Kurrey
NK: Stem and progenitor-like cells contribute to the aggressive
behavior of human epithelial ovarian cancer. Cancer Res.
65:3025–3029. 2005.PubMed/NCBI
|
|
45
|
Alvero AB, Chen R, Fu HH, et al: Molecular
phenotyping of human ovarian cancer stem cells unravel the
mechanisms for repair and chemo-resistance. Cell Cycle. 8:158–166.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Hermann P, Huber S, Herrler T, et al:
Distinct populations of cancer stem cells determine tumor growth
and metastatic activity in human pancreatic cancer. Cell Stem Cell.
1:313–323. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Sun S and Wang Z: Head neck squamous cell
carcinoma c-Met(+) cells display cancer stem cell properties and
are responsible for cisplatin-resistance and metastasis. Int J
Cancer. 129:2337–2348. 2011.
|
|
48
|
Monroe MM, Anderson EC, Clayburgh DR and
Wong MH: Cancer stem cells in head and neck squamous cell
carcinoma. J Oncol. 2011:7627802011. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Sayed SI, Dwivedi RC, Katna R, et al:
Implications of understanding cancer stem cell (CSC) biology in
head and neck squamous cell cancer. Oral Oncol. 47:237–243. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Zito G, Richiusa P, Bommarito A, et al: In
vitro identification and characterization of CD133(pos) cancer
stem-like cells in anaplastic thyroid carcinoma cell lines. PLoS
One. 3:e35442008. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Wang J, Guo LP, Chen LZ, Zeng YX and Lu
SH: Identification of cancer stem cell-like side population cells
in human nasopharyngeal carcinoma cell line. Cancer Res.
67:3716–3724. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Prince ME, Sivanandan R, Kaczorowski A, et
al: Identification of a subpopulation of cells with cancer stem
cell properties in head and neck squamous cell carcinoma. Proc Natl
Acad Sci USA. 104:973–978. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Prince ME and Ailles LE: Cancer stem cells
in head and neck squamous cell cancer. J Clin Oncol. 26:2871–2875.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Balic M, Lin H, Young L, et al: Most early
disseminated cancer cells detected in bone marrow of breast cancer
patients have a putative breast cancer stem cell phenotype. Clin
Cancer Res. 12:5615–5621. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Charafe-Jauffret E, Ginestier C, Iovino F,
et al: Aldehyde dehydrogenase 1-positive cancer stem cells mediate
metastasis and poor clinical outcome in inflammatory breast cancer.
Clin Cancer Res. 16:45–55. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Davis SJ, Divi V, Owen JH, et al:
Metastatic potential of cancer stem cells in head and neck squamous
cell carcinoma. Arch Otolaryngol Head Neck Surg. 136:1260–1266.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Faber A, Barth C, Hörmann K, et al: CD44
as a stem cell marker in head and neck squamous cell carcinoma.
Oncol Rep. 26:321–326. 2011.PubMed/NCBI
|
|
58
|
Song J, Chang I, Chen Z, Kang M and Wang
CY: Characterization of side populations in HNSCC: highly invasive,
chemoresistant and abnormal Wnt signaling. PLoS One. 5:e114562010.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Goldie SJ, Mulder KW, Tan DW, Lyons SK,
Sims AH and Watt FM: FRMD4A upregulation in human squamous cell
carcinoma promotes tumor growth and metastasis and is associated
with poor prognosis. Cancer Res. 72:3424–3436. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
La Fleur L, Johansson AC and Roberg K: A
CD44high/EGFRlow subpopulation within head and neck cancer cell
lines shows an epithelial-mesenchymal transition phenotype and
resistance to treatment. PLoS One. 7:e440712012.PubMed/NCBI
|
|
61
|
Tamara Marie-Egyptienne DT, Lohse I and
Hill RP: Cancer stem cells, the epithelial to mesenchymal
transition (EMT) and radioresistance: potential role of hypoxia.
Cancer Lett. Nov 28–2012.Epub ahead of print. View Article : Google Scholar
|
|
62
|
Todaro M, Alea MP, Di Stefano AB, et al:
Colon cancer stem cells dictate tumor growth and resist cell death
by production of interleukin-4. Cell Stem Cell. 1:389–402. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Cheung ST, Cheung PF, Cheng CK, Wong NC
and Fan ST: Granulin-epithelin precursor and ATP-dependent binding
cassette (ABC)B5 regulate liver cancer cell chemoresistance.
Gastroenterology. 140:344–355. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Bussolati B, Grange C, Sapino A and
Camussi G: Endothelial cell differentiation of human breast tumour
stem/progenitor cells. J Cell Mol Med. 13:309–319. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Hendrix MJ, Seftor EA, Meltzer PS, et al:
Expression and functional significance of VE-cadherin in aggressive
human melanoma cells: role in vasculogenic mimicry. Proc Natl Acad
Sci USA. 98:8018–8023. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Soda Y, Marumoto T, Friedmann-Morvinski D,
et al: Transdifferentiation of glioblastoma cells into vascular
endothelial cells. Proc Natl Acad Sci USA. 108:4274–4280. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Lai CY, Schwartz BE and Hsu MY: CD133+
melanoma subpopulations contribute to perivascular niche
morphogenesis and tumorigenicity through vasculogenic mimicry.
Cancer Res. 72:5111–5118. 2012.
|
|
68
|
Frank NY, Schatton T, Kim S, et al:
VEGFR-1 expressed by malignant melanoma-initiating cells is
required for tumor growth. Cancer Res. 71:1474–1485. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Dang D and Ramos DM: Identification of
αvβ6-positive stem cells in oral squamous cell carcinoma.
Anticancer Res. 29:2043–2049. 2009.
|
|
70
|
Zhang S, Guo H, Zhang D, et al:
Microcirculation patterns in different stages of melanoma growth.
Oncol Rep. 15:15–20. 2006.PubMed/NCBI
|
|
71
|
Polyak K and Weinberg RA: Transitions
between epithelial and mesenchymal states: acquisition of malignant
and stem cell traits. Nat Rev Cancer. 9:265–273. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Nguyen PT, Kudo Y, Yoshida M, Kamata N,
Ogawa I and Takata T: N-cadherin expression is involved in
malignant behavior of head and neck cancer in relation to
epithelial-mesenchymal transition. Histol Histopathol. 26:147–156.
2011.PubMed/NCBI
|
|
73
|
Mandal M, Myers JN, Lippman SM, et al:
Epithelial to mesenchymal transition in head and neck squamous
carcinoma: association of Src activation with E-cadherin
down-regulation, vimentin expression, and aggressive tumor
features. Cancer. 112:2088–2100. 2008. View Article : Google Scholar
|
|
74
|
Batlle E, Sancho E, Franci C, et al: The
transcription factor snail is a repressor of E-cadherin gene
expression in epithelial tumour cells. Nat Cell Biol. 2:84–89.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Cano A, Pérez-Moreno MA, Rodrigo I, et al:
The transcription factor snail controls epithelial-mesenchymal
transitions by repressing E-cadherin expression. Nat Cell Biol.
2:76–83. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Zhang A, Chen G, Meng L, et al:
Antisense-Snail transfer inhibits tumor metastasis by inducing
E-cadherin expression. Anticancer Res. 28:621–628. 2008.PubMed/NCBI
|
|
77
|
Hajra KM, Chen DY and Fearon ER: The SLUG
zinc-finger protein represses E-cadherin in breast cancer. Cancer
Res. 62:1613–1618. 2002.PubMed/NCBI
|
|
78
|
Yang J, Mani SA, Donaher JL, et al: Twist,
a master regulator of morphogenesis, plays an essential role in
tumor metastasis. Cell. 117:927–939. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Zhang J, Liang Q, Lei Y, et al: SOX4
induces epithelial-mesenchymal transition and contributes to breast
cancer progression. Cancer Res. 72:4597–4608. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Eger A, Aigner K, Sonderegger S, et al:
DeltaEF1 is a transcriptional repressor of E-cadherin and regulates
epithelial plasticity in breast cancer cells. Oncogene.
24:2375–2385. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Huber MA, Kraut N and Beug H: Molecular
requirements for epithelial-mesenchymal transition during tumor
progression. Curr Opin Cell Biol. 17:548–558. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Porsch H, Bernert B, Mehic M, Theocharis
AD, Heldin CH and Heldin P: Efficient TGFbeta-induced
epithelial-mesenchymal transition depends on hyaluronan synthase
HAS2. Oncogene. Oct 29–2012.(Epub ahead of print). View Article : Google Scholar
|
|
83
|
Wu ZQ, Li XY, Hu CY, Ford M, Kleer CG and
Weiss SJ: Canonical Wnt signaling regulates Slug activity and links
epithelial-mesenchymal transition with epigenetic Breast Cancer 1,
Early Onset (BRCA1) repression. Proc Natl Acad Sci USA.
109:16654–16659. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Guaita S, Puig I, Franci C, et al: Snail
induction of epithelial to mesenchymal transition in tumor cells is
accompanied by MUC1 repression and ZEB1 expression. J Biol Chem.
277:39209–39216. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Zhao XL, Sun T, Che N, et al: Promotion of
hepatocellular carcinoma metastasis through matrix
metalloproteinase activation by epithelial-mesenchymal transition
regulator Twist1. J Cell Mol Med. 15:691–700. 2011. View Article : Google Scholar
|
|
86
|
Tsai JH, Donaher JL, Murphy DA, Chau S and
Yang J: Spatiotemporal regulation of epithelial-mesenchymal
transition is essential for squamous cell carcinoma metastasis.
Cancer Cell. 22:725–736. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Aigner K, Dampier B, Descovich L, et al:
The transcription factor ZEB1 (deltaEF1) promotes tumour cell
dedifferentiation by repressing master regulators of epithelial
polarity. Oncogene. 26:6979–6988. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Wellner U, Schubert J, Burk UC, et al: The
EMT-activator ZEB1 promotes tumorigenicity by repressing
stemness-inhibiting microRNAs. Nat Cell Biol. 11:1487–1495. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Ahn YH, Gibbons DL, Chakravarti D, et al:
ZEB1 drives prometastatic actin cytoskeletal remodeling by
downregulating miR-34a expression. J Clin Invest. 122:3170–3183.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Casas E, Kim J, Bendesky A, Ohno-Machado
L, Wolfe CJ and Yang J: Snail2 is an essential mediator of
Twist1-induced epithelial mesenchymal transition and metastasis.
Cancer Res. 71:245–254. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Sun T, Sun BC, Zhao XL, et al: Promotion
of tumor cell metastasis and vasculogenic mimicry by way of
transcription coactivation by Bcl-2 and Twist1: a study of
hepatocellular carcinoma. Hepatology. 54:1690–1706. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Lirdprapamongkol K, Chiablaem K, Sila-Asna
M, Surarit R, Bunyaratvej A and Svasti J: Exploring stemness gene
expression and vasculogenic mimicry capacity in well- and
poorly-differentiated hepatocellular carcinoma cell lines. Biochem
Biophys Res Commun. 422:429–435. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Passegué E, Jamieson CH, Ailles LE and
Weissman IL: Normal and leukemic hematopoiesis: are leukemias a
stem cell disorder or a reacquisition of stem cell characteristics?
Proc Natl Acad Sci USA. 100(Suppl 1): 11842–11849. 2003.
|
|
94
|
Prindull G: Hypothesis: cell plasticity,
linking embryonal stem cells to adult stem cell reservoirs and
metastatic cancer cells? Exp Hematol. 33:738–746. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Notta F, Mullighan CG, Wang JC, et al:
Evolution of human BCR-ABL1 lymphoblastic leukaemia-initiating
cells. Nature. 469:362–367. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Biddle A, Liang X, Gammon L, et al: Cancer
stem cells in squamous cell carcinoma switch between two distinct
phenotypes that are preferentially migratory or proliferative.
Cancer Res. 71:5317–5326. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Damonte P, Gregg JP, Borowsky AD, Keister
BA and Cardiff RD: EMT tumorigenesis in the mouse mammary gland.
Lab Invest. 87:1218–1226. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Santisteban M, Reiman JM, Asiedu MK, et
al: Immune-induced epithelial to mesenchymal transition in vivo
generates breast cancer stem cells. Cancer Res. 69:2887–2895. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Fang X, Cai Y, Liu J, et al: Twist2
contributes to breast cancer progression by promoting an
epithelial-mesenchymal transition and cancer stem-like cell
self-renewal. Oncogene. 30:4707–4720. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Asiedu MK, Ingle JN, Behrens MD, Radisky
DC and Knutson KL: TGFbeta/TNF(alpha)-mediated
epithelial-mesenchymal transition generates breast cancer stem
cells with a claudin-low phenotype. Cancer Res. 71:4707–4719. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Ryu HS, Park do J, Kim HH, Kim WH and Lee
HS: Combination of epithelial-mesenchymal transition and cancer
stem cell-like phenotypes has independent prognostic value in
gastric cancer. Hum Pathol. 43:520–528. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Sarkar D, Shields B, Davies ML, Muller J
and Wakeman JA: BRACHYURY confers cancer stem cell characteristics
on colorectal cancer cells. Int J Cancer. 130:328–337. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Xia H, Cheung WK, Sze J, et al: miR-200a
regulates epithelial-mesenchymal to stem-like transition via ZEB2
and beta-catenin signaling. J Biol Chem. 285:36995–37004. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Chen YC, Chen YW, Hsu HS, et al: Aldehyde
dehydrogenase 1 is a putative marker for cancer stem cells in head
and neck squamous cancer. Biochem Biophys Res Commun. 385:307–313.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Chen YS, Wu MJ, Huang CY, et al: CD133/Src
axis mediates tumor initiating property and epithelial-mesenchymal
transition of head and neck cancer. PLoS One. 6:e280532011.
View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Hendrix MJ, Seftor EA, Hess AR and Seftor
RE: Vasculogenic mimicry and tumour-cell plasticity: lessons from
melanoma. Nat Rev Cancer. 3:411–421. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Pisacane AM, Picciotto F and Risio M: CD31
and CD34 expression as immunohistochemical markers of endothelial
transdifferentiation in human cutaneous melanoma. Cell Oncol.
29:59–66. 2007.PubMed/NCBI
|
|
108
|
Garnier D, Milsom C, Magnus N, et al: Role
of the tissue factor pathway in the biology of tumor initiating
cells. Thromb Res. 125(Suppl 2): S44–S50. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Gill JG, Langer EM, Lindsley RC, Cai M,
Murphy TL and Murphy KM: Snail promotes the cell-autonomous
generation of Flk1(+) endothelial cells through the repression of
the microRNA-200 family. Stem Cells Dev. 21:167–176.
2012.PubMed/NCBI
|
|
110
|
Sun B, Zhang D, Zhang S, Zhang W, Guo H
and Zhao X: Hypoxia influences vasculogenic mimicry channel
formation and tumor invasion-related protein expression in
melanoma. Cancer Lett. 249:188–197. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Ma JL, Han SX, Zhu Q, et al: Role of Twist
in vasculogenic mimicry formation in hypoxic hepatocellular
carcinoma cells in vitro. Biochem Biophys Res Commun. 408:686–691.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Comito G, Calvani M, Giannoni E, et al:
HIF-1alpha stabilization by mitochondrial ROS promotes
Met-dependent invasive growth and vasculogenic mimicry in melanoma
cells. Free Radic Biol Med. 51:893–904. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Misra RM, Bajaj MS and Kale VP:
Vasculogenic Mimicry of HT1080 Tumour Cells In Vivo: Critical Role
of HIF-1alpha-Neuropilin-1 Axis. PLoS One. 7:e501532012. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Jiang J, Tang YL and Liang XH: EMT: a new
vision of hypoxia promoting cancer progression. Cancer Biol Ther.
11:714–723. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Bao B, Azmi AS, Ali S, et al: The
biological kinship of hypoxia with CSC and EMT and their
relationship with deregulated expression of miRNAs and tumor
aggressiveness. Biochim Biophys Acta. 1826:272–296. 2012.PubMed/NCBI
|
|
116
|
Carmeliet P and Jain RK: Angiogenesis in
cancer and other diseases. Nature. 407:249–257. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Xu X and Prestwich GD: Inhibition of tumor
growth and angiogenesis by a lysophosphatidic acid antagonist in an
engineered three-dimensional lung cancer xenograft model. Cancer.
116:1739–1750. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Keunen O, Johansson M, Oudin A, et al:
Anti-VEGF treatment reduces blood supply and increases tumor cell
invasion in glioblastoma. Proc Natl Acad Sci USA. 108:3749–3754.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Qu B, Guo L, Ma J and Lv Y:
Antiangiogenesis therapy might have the unintended effect of
promoting tumor metastasis by increasing an alternative circulatory
system. Med Hypotheses. 74:360–361. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Cao G, Fehrenbach ML, Williams JT,
Finklestein JM, Zhu JX and Delisser HM: Angiogenesis in platelet
endothelial cell adhesion molecule-1-null mice. Am J Pathol.
175:903–915. 2009. View Article : Google Scholar : PubMed/NCBI
|