Introduction
The vinca alkaloid, vinblastine, is a chemical
analogue of vincristine. Vinblastine binds to tubulin, which
inhibits microtubule assembly. Vinblastine is an anti-tumor drug
that is widely used in cancer chemotherapy, as it inhibits cell
proliferation via a mitotic block (1,2). This
anti-microtubule agent is used to treat various types of cancer,
including Hodgkin’s lymphoma, Kaposi sarcoma, breast cancer, head
and neck cancer, Langerhans cell histiocytosis and testicular
cancer (3–6). In addition, it is used to treat
non-small cell lung cancer (7).
Although the mechanism behind it is not entirely
understood, one characteristic of vinblastine treatment is the
formation of giant paracrystalline aggregates in the cell
cytoplasm. These paracrystals consist of tubulin molecules and can
be detected by tubulin immunostaining under a light microscope or
by transmission electron microscopy (8–13). The
mechanism of paracrystal formation has been ascribed to the
capacity of vinblastine to depolymerize mitotic spindle
microtubules. To understand the mechanism of paracrystalline
aggregate formation in more detail, an experimental system for
observing their formation is required.
RBM8A, a member of the RNA binding motif (RBM)
family (14), localizes to mRNA
through its RNA binding domain (15). RBM8A binds to spliced mRNA and
maintains this binding until the bound mRNA is degraded (16). It was previously demonstrated that
depleting RBM8A proteins in Drosophila SL2 cells resulted in
impaired cell growth (17).
Furthermore, Sudo et al performed loss-of-function screening
for genes involved in apoptosis and growth for a human mesothelioma
cell line (18). In addition to the
COPA gene, RBM8A was also shown to contribute to cell
growth, as observed by gene silencing experiments using RNAi. In
addition, these results were confirmed using human tumor cell lines
and the contribution of G2/M phase progression was
indicated. In our previous study, an essential function was
identified for cell cycle progression for the RNA binding protein
RBM8A in A549 cells (19).
Knockdown of the RBM8A gene resulted in arrest at the
G2/M phase, concomitant with aberrant centrosome
formation. In addition, these cells underwent apoptosis following
RBM8A knockdown. On the other hand, in our recent study,
immunostaining experiments showed that RBM8A proteins were
localized at centrosomes and microtubules (20). This was confirmed by the presence of
exogenous tagged RBM8A in A549 cells. These results prompted the
study of the localization of RBM8A proteins with respect to
paracrystals in the present study.
Recent progress in using the exogenous expression of
fluorescent proteins that are conjugated with polypeptides via
baculovirus infection has enabled simple, rapid visualization of
target proteins in living cells (21–23).
This can be combined with time-lapse microscopy and can be used to
make movies of living cells (24,25).
The present study aimed to develop vinblastine-induced
paracrystalline aggregate formation in a human lung tumor cell line
and establish a time-lapse analysis system.
Materials and methods
Cell culture and introduction of labeled
proteins
The human non-small cell lung cancer A549 cell line
(Riken Tsukuba Institute, Tsukuba, Japan) was maintained in
Dulbecco’s modified Eagle’s medium (Sigma-Aldrich, St. Louis, MO,
USA), supplemented with 10% fetal bovine serum (Sigma-Aldrich) and
antibiotics [penicillin (100 units/ml) and streptomycin (100
units/ml) solution; Wako Pure Chemicals Co., Ltd., Osaka, Japan]. A
total of 40,000 cells were seeded onto a glass-bottomed dish (Asahi
Glass Co., Ltd., Tokyo, Japan). The cells were allowed to adhere
and grow for two days at 37°C in 5% CO2 prior to
applying Cellular Lights™ (Invitrogen Life Technologies, Carlsbad,
CA, USA) transduction.
Introducing Cellular Lights
To introduce fluorescent proteins conjugated with
proteins, Cellular Lights Red Fluorescent Protein (RFP)-Tubulin and
Cellular Lights Green Fluorescent Protein (GFP)-Actin were
introduced at the same time. Cellular Lights Null (empty control)
was used as a negative control. All of these regents were purchased
from Invitrogen Life Technologies. The reagents contained a
baculovirus that enables the expression of autofluorescent proteins
upon entry into insect cells. The use of baculovirus to deliver
genes into mammalian cells, referred to as BacMam technology, was
developed and became commercially available fairly recently
(21,22). BacMam technology has the following
significant features: i) High transduction efficiency, ii) minimal
cytotoxic effects, iii) high expression levels, iv) safety, as it
cannot replicate in mammalian cells, and v) easy delivery of
multiple different genes. Thus, BacMam technology is a method of
gene delivery with few or no observable side-effects. The reagents
used combine fluorescent protein-tagged target proteins with the
viral delivery used with BacMam technology, which results in
extensive expression in mammalian cells. All reagents were used
according to the manufacturer’s protocol.
In brief, the BacMam enhancer solution was prepared
by reconstituting an entire vial of enhancer in dimethyl sulfoxide
(Sigma-Aldrich). The transduction solution was prepared by
combining a Cellular Lights reagent with Dulbecco’s
phosphate-buffered saline (D-PBS, Wako Pure Chemicals Co., Ltd.).
To simultaneously transduce cells with RFP-Tubulin and GFP-Actin,
the additional volume of the reagents was substituted for PBS.
Subsequent to aspirating the transduction solution from the cell
culture dish, culture medium with or without serum plus enhancer
was added. The cells were subsequently incubated for 2 h.
Furthermore, the cells were incubated at 37°C in 5% CO2
for 16 h, during which time the introduced fluorescent proteins
were expressed.
Expression of cell components and
time-lapse analysis
Time-lapse imaging was performed using an incubator
microscope system (LCV110; Olympus Corp., Tokyo, Japan).
Vinblastine sulfate (30 μM; Calbiochem, La Jolla, CA, USA) was
added to the cells in which the Cellular Lights reagents had been
introduced. The glass-based dishes were set up in a tray for the
LCV110 system and preincubated for 60 min. Image acquisition was
begun using a time interval of 15–20 min between each acquisition.
These images were processed using the MetaMorph software (Molecular
Devices, Inc., Sunnyvale, CA, USA).
Immunostaining
The cells were fixed in 4% paraformaldehyde,
followed by treatment with 0.2% Triton X-100 (Sigma-Aldrich). Next,
either a polyclonal primary goat antibody against tubulin (1:200;
Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA) or a mouse
anti-human RBM8A monoclonal antibody (1:1,000; clone 4C4,
Sigma-Aldrich) was incubated with the fixed cells. Alexa Fluor 488
(green) or 589 (red) conjugated secondary antibodies (Molecular
Probes; Invitrogen Life Technologies) were used as appropriate.
Cell nuclei were stained with DAPI. Prolong Gold anti-fade reagent
(Invitrogen Life Technologies) was used to avoid fading. Images
were acquired using an Axiovert 200M microscope (Carl Zeiss,
Oberkochen, Germany) and processed using ZEN software (Carl
Zeiss).
Results
Vinblastine-induced paracrystal formation
in A549 cells
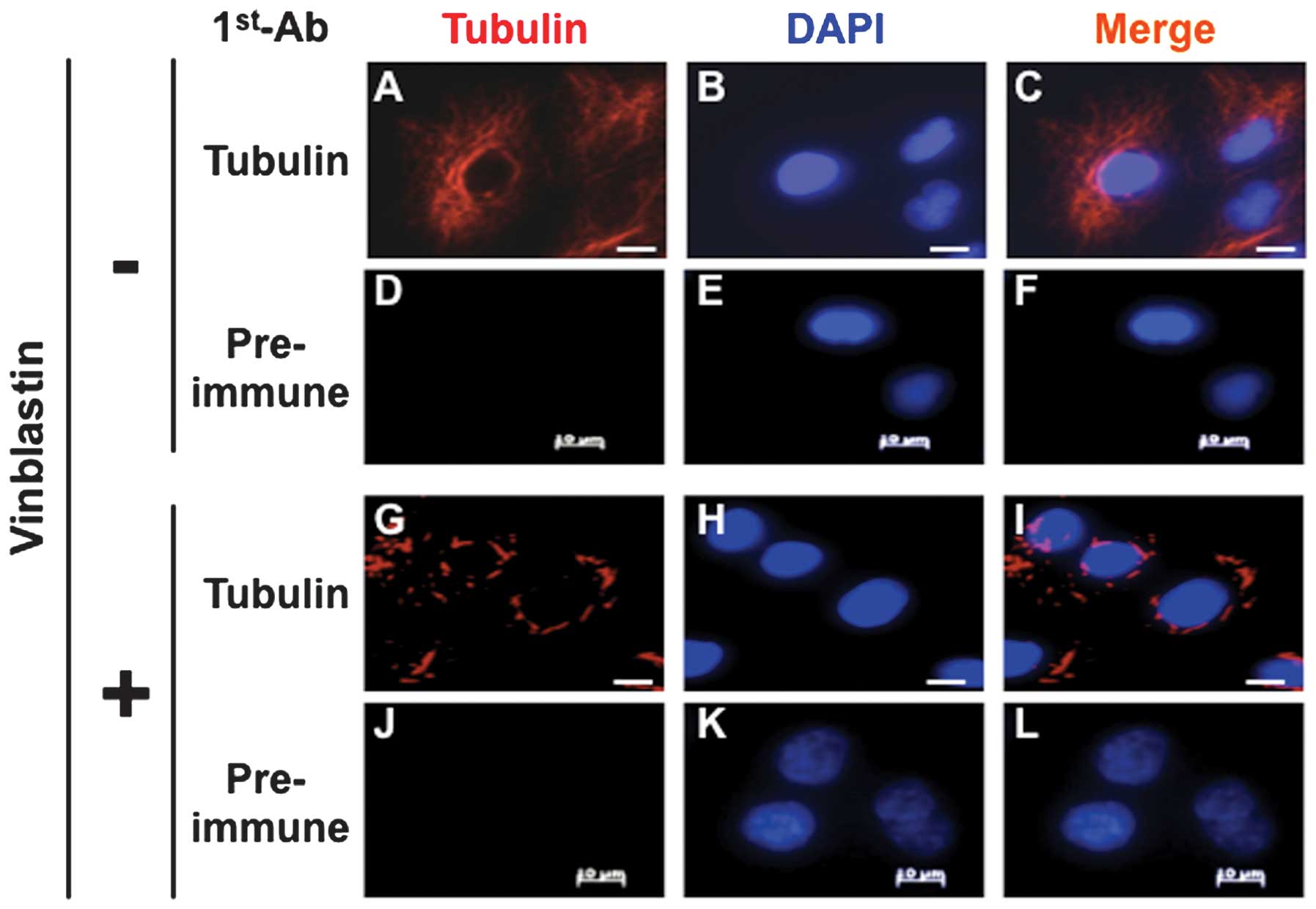
To confirm paracrystal formation in the A549 cells,
the cells were treated with vinblastine for 12 h and the
paracrystals that formed were stained with an anti-tubulin
antibody, as described in previous studies (26,27).
As shown in Fig. 1A–C, tubulin
cytoplasmic localization was verified in the cells without
vinblastine treatment. In the vinblastine-treated cells,
characteristic paracrystal tube-like structure formation could
clearly be detected (Fig. 1G–I).
Thus, vinblastine treatment caused paracrystal formation in the
A549 cells.
RFP-Tubulin co-localizes with tubulin
staining
RFP-Tubulin was introduced and immunostaining was
performed with an anti-tubulin antibody. As shown in Fig. 2, the RFP-Tubulin was expressed in
the A549 cells and was localized to the centrosomes (Fig. 2A–D). The signals from immunostaining
with an anti-tubulin antibody co-localized with those of
RFP-Tubulin. Thus, RFP-Tubulin could be used to detect the
formation of paracrystals, which were identified by an anti-tubulin
antibody (Fig. 2I–L).
At the same time, a mouse anti-RBM8A antibody
(Sigma-Aldrich) was used, as our previous study had indicated that
this protein localized to centrosomes. Our preliminary results
showed vivid staining of the spindle fibers, in addition to the
centrosomes, when using this antibody (20). Thus, we speculated that the
anti-RBM8A antibody from Sigma-Aldrich could co-localize with
paracrystals. As shown in Fig.
2M–P, the signals from immunostaining with the anti-RBM8A
antibody overlapped with the RFP-Tubulin signals. Thus, this
monoclonal antibody was useful for the detection of paracrystals.
In addition, in place of the antibody from Sigma-Aldrich, a
self-made rabbit antiserum against the N-terminal region of RBM8A
was used and similar results were obtained (data not shown). Thus,
it was concluded that RNA-binding RBM8A proteins that were
localized to paracrystals formed due to vinblastine treatment.
Observations of vinblastine-induced
paracrystals by Cellular Lights
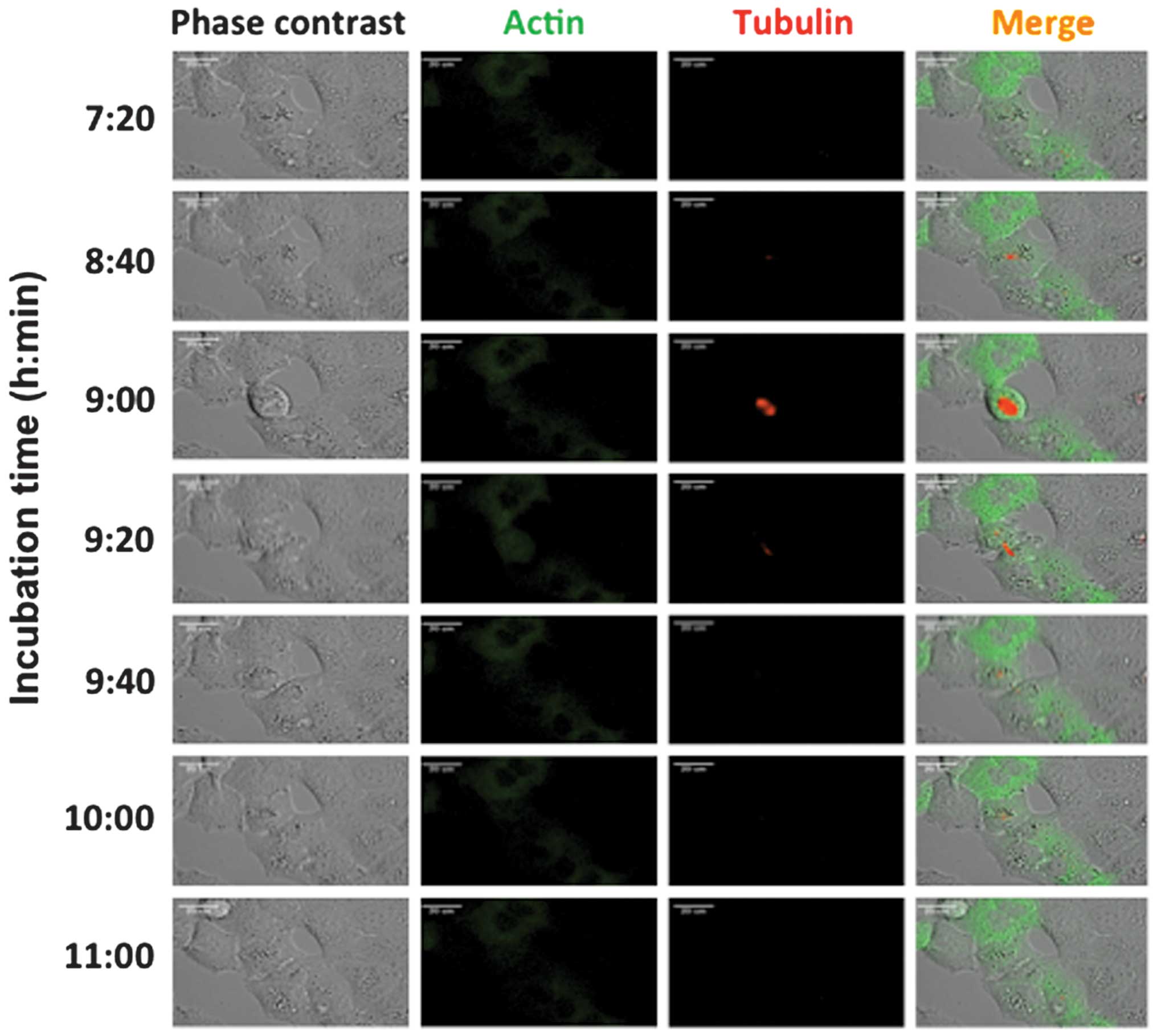
To obtain time-lapse images of paracrystal
formation, we introduced Cellular Lights RFP-Tubulin into the A549
cells and observed the cells using an Olympus LCV110 system. To
determine the outline of each cell, GFP-Actin was also introduced.
Fig. 3 shows the results of the
time-lapse analysis of the cells without vinblastine treatment
using the LCV110 system and the progression of the mitotic phases.
Green signals were derived from GFP-Actin and red signals were
derived from RFP-Tubulin. Microtubule formation during mitotic
phases was observed by RFP-Tubulin at the 9 h 00 min time-point,
and GFP-Actin was spread through the entire cytoplasm.
In comparison, the time-lapse results for the
vinblastine-treated cells are shown in Fig. 4 (images obtained on different days).
Tube-like structures were clearly demonstrated by the red signals
derived from RFP at the 7 h 11 min time-point. These paracrystals
gradually grew until the 12 h 11 min time-point. During paracrystal
formation, cell cycle progression was arrested around mitosis, as
the cell shapes were round. At the same time, the green signals
derived from GFP spread as shown in the negative control cells.
Subsequent to the 15 h 41 min time-point, rounded cells
predominantly appeared and cell cycle progression was inhibited.
These cells retained paracrystals and appeared to be arrested at
the mitotic phase.
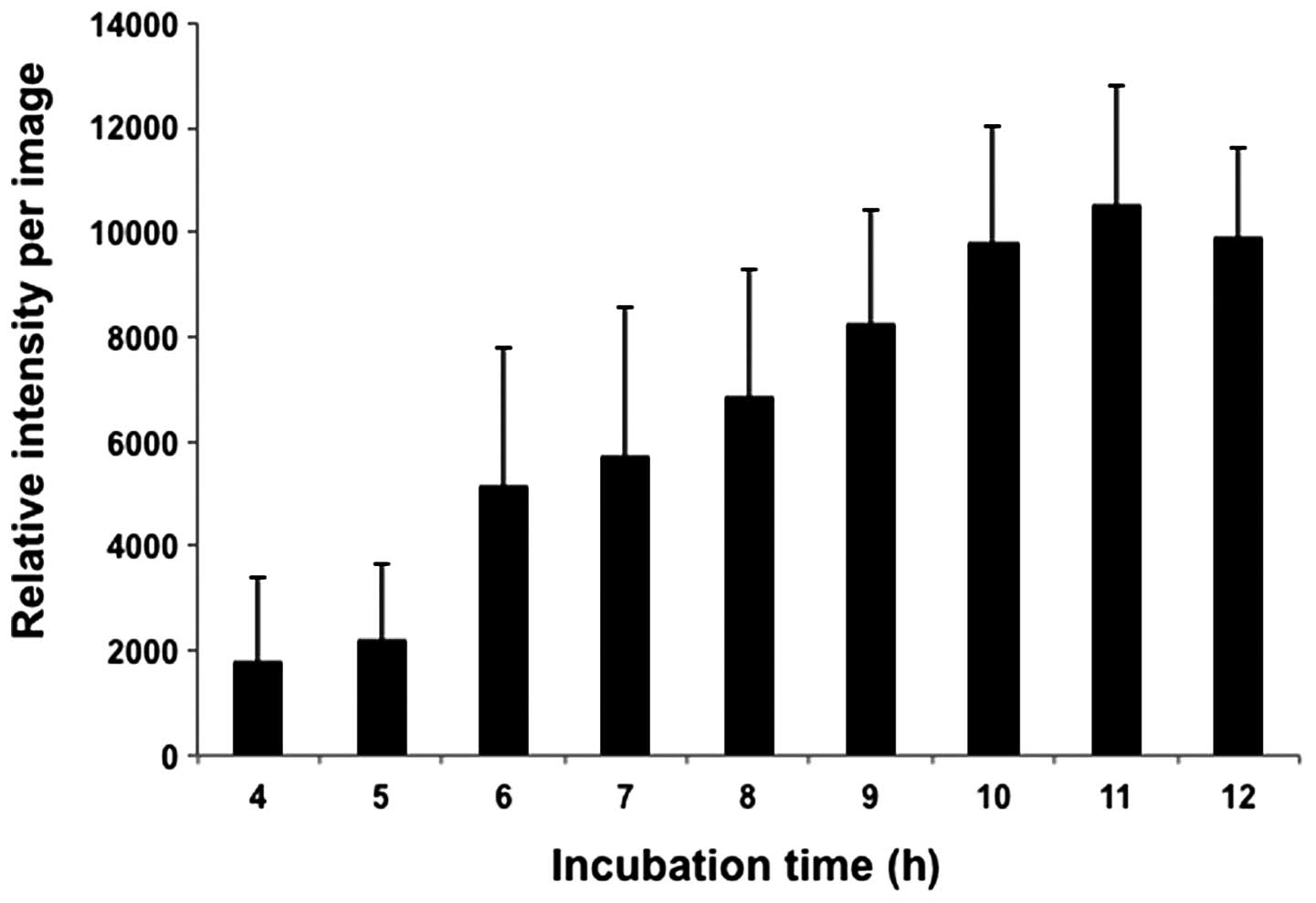
Finally, paracrystalline RFP fluorescence intensity
was measured on these time-lapse images, which showed the time
dependency for their formation (Fig.
5). Based on the images in Fig.
4, RFP fluorescence intensity gradually increased and the green
signals were stably expressed.
Discussion
The immunostaining results of the present study
revealed overlapping signals from an anti-tubulin antibody and an
anti-RBM8A antibody in A549 cells. In a previous study,
localization of syndecan proteins to tubulin was reported on the
basis of its localization to vinblastine-induced paracrystals
(13). Similar to this study, the
co-localization of RBM8A to paracrystals in the present study
supports our previous observations that RBM8A localizes to
microtubules and centrosomes (20).
As mRNA-protein complexes are known to move via their binding to
microtubules (28), RBM8A
localization may indicate that certain mRNA molecules on
microtubules bind to RBM8A. To examine whether RBM8A is required
for paracrystalline formation, we repeatedly performed knockdown
experiments in A549 cells, but these trials resulted in rapid cell
death and did not obtain clear results (data not shown).
In the present study, a time-lapse analysis system
was successfully established. The combination of time-lapse
analysis with RFP-tubulin expression is the main technique analyzed
in the present study. This method was anticipated to obtain more
detailed information compared with conventional immunostaining
methods with fixed cells. Staining became visible following spindle
fiber or paracrystal formation. RFP-Tubulin was expressed by
Cellular Lights and the molecules spread through the cytoplasm.
RFP-tubulin was not evident as the proteins were diffuse. When
RFP-tubulin forms microtubules or paracrystals due to
polymerization, it becomes clearly visible.
As immunostaining with an RBM8A antibody
successfully detected paracrystals in these cells, the expression
of GFP-tagged RBM8A proteins was attempted in the A549 cells. The
fluorescent protein-tagged RBM8A also appeared to be useful for the
detection of paracrystals, similar to RFP-tubulin in the present
study. However, transient expression of this modified RBM8A protein
caused rapid cell death similar to that observed in the knockdown
experiments (data not shown) and it was therefore concluded that
this was not suitable for detection. In the case of time-lapse
analysis, in future studies, combinations with other cell cycle
progression markers, such as a fluorescent ubiquitination-based
cell cycle indicator system (29,30),
will enable the analysis of cell cycle-dependent mechanisms.
Acknowledgments
This study was supported by grants from Kanazawa
Medical University (S2013-2 and SR2012-02) and a Grant-in-Aid for
Scientific Research in Japan (KAKENHI, 25460376).
References
|
1
|
Moudi M, Go R, Yien CY and Nazre M: Vinca
alkaloids. Int J Prev Med. 4:1231–1235. 2013.
|
|
2
|
Nelson RL: The comparative clinical
pharmacology and pharmacokinetics of vindesine, vincristine, and
vinblastine in human patients with cancer. Med Pediatr Oncol.
10:115–127. 1982.
|
|
3
|
Duggan DB, Petroni GR, Johnson JL, et al:
Randomized comparison of ABVD and MOPP/ABV hybrid for the treatment
of advanced Hodgkin’s disease: report of an intergroup trial. J
Clin Oncol. 21:607–614. 2003.
|
|
4
|
Ladisch S, Gadner H, Aricò M, et al:
LCH-I: a randomized trial of etoposide vs. vinblastine in
disseminated Langerhans cell histiocytosis. Med Pediatr Oncol.
23:107–110. 1994.
|
|
5
|
Epstein JB: Treatment of oral Kaposi
sarcoma with intralesional vinblastine. Cancer. 71:1722–1725.
1993.
|
|
6
|
Einhorn LH: Curing metastatic testicular
cancer. Proc Natl Acad Sci USA. 99:4592–4595. 2002.
|
|
7
|
Woodcock TM, Blumenreich MS, Richman SP,
Kubota TT, Gentile PS and Allegra JC: Combination chemotherapy with
cis-diamminedichloroplatinum and vinblastine in advanced non-small
cell lung cancer. J Clin Oncol. 1:247–250. 1983.
|
|
8
|
Kovacs K and Horvath E:
Vinblastine-induced ultrastructural changes in perisinusoidal cells
of the rat liver. Res Exp Med (Berl). 165:245–249. 1975.
|
|
9
|
Shiino M: Ultrastructural observation of
paracrystalline aggregates of microtubules in anterior pituitary
cells of the chinchilla (Chinchilla laniger). Cell Tissue
Res. 213:433–440. 1980.
|
|
10
|
De Brabander M, De May J, Joniau M and
Geuens G: Ultrastructural immunocytochemical distribution of
tubulin in cultured cells treated with microtubule inhibitors. Cell
Biol Int Rep. 1:177–183. 1977.
|
|
11
|
Shiino M and Rennels EG:
Vinblastine-induced microtubular paracrystals in prolactin cells of
anterior pituitary gland of lactating rats. Am J Anat. 144:399–405.
1975.
|
|
12
|
Nogales E, Medrano FJ, Diakun GP, Mant GR,
Towns-Andrews E and Bordas J: The effect of temperature on the
structure of vinblastine-induced polymers of purified tubulin:
detection of a reversible conformational change. J Mol Biol.
254:416–430. 1995.
|
|
13
|
Brockstedt U, Dobra K, Nurminen M and
Hjerpe A: Immunoreactivity to cell surface syndecans in cytoplasm
and nucleus: tubulin-dependent rearrangements. Exp Cell Res.
274:235–245. 2002.
|
|
14
|
Salicioni AM, Xi M, Vanderveer LA, et al:
Identification and structural analysis of human RBM8A and RBM8B:
two highly conserved RNA-binding motif proteins that interact with
OVCA1, a candidate tumor suppressor. Genomics. 69:54–62. 2000.
|
|
15
|
Lau CK, Diem MD, Dreyfuss G and Van Duyne
GD: Structure of the Y14-Magoh core of the exon junction complex.
Curr Biol. 13:933–941. 2003.
|
|
16
|
Chuang TW, Chang WL, Lee KM and Tarn WY:
The RNA-binding protein Y14 inhibits mRNA decapping and modulates
processing body formation. Mol Biol Cell. 24:1–13. 2013.
|
|
17
|
Le Hir H, Gatfield D, Braun IC, Forler D
and Izaurralde E: The protein Mago provides a link between splicing
and mRNA localization. EMBO Rep. 2:1119–1124. 2001.
|
|
18
|
Sudo H, Tsuji AB, Sugyo A, et al:
Knockdown of COPA, identified by loss-of-function screen, induces
apoptosis and suppresses tumor growth in mesothelioma mouse model.
Genomics. 95:210–216. 2010.
|
|
19
|
Ishigaki Y, Nakamura Y, Tatsuno T, et al:
Depletion of RNA-binding protein RBM8A (Y14) causes cell cycle
deficiency and apoptosis in human cells. Exp Biol Med (Maywood).
238:889–897. 2013.
|
|
20
|
Ishigaki Y, Nakamura Y, Tatsuno T,
Hashimoto M, Iwabuchi K and Tomosugi N: RNA-binding protein RBM8A
(Y14) and MAGOH localize to centrosome in human A549 cells.
Histochem Cell Biol. 141:101–109. 2014.
|
|
21
|
Kost TA, Condreay JP and Jarvis DL:
Baculovirus as versatile vectors for protein expression in insect
and mammalian cells. Nat Biotechnol. 23:567–575. 2005.
|
|
22
|
Kost TA and Condreay JP: Recombinant
baculoviruses as mammalian cell gene-delivery vectors. Trends
Biotechnol. 20:173–180. 2002.
|
|
23
|
Shukla S, Schwartz C, Kapoor K, Kouanda A
and Ambudkar SV: Use of baculovirus BacMam vectors for expression
of ABC drug transporters in mammalian cells. Drug Metab Dispos.
40:304–312. 2012.
|
|
24
|
Araki R, Jincho Y, Hoki Y, et al:
Conversion of ancestral fibroblasts to induced pluripotent stem
cells. Stem Cells. 28:213–220. 2010.
|
|
25
|
Sakaue-Sawano A, Kobayashi T, Ohtawa K and
Miyawaki A: Drug-induced cell cycle modulation leading to
cell-cycle arrest, nuclear mis-segregation, or endoreplication. BMC
Cell Biol. 12:22011.
|
|
26
|
Ben-Ze’ev A, Farmer SR and Penman S:
Mechanisms of regulating tubulin synthesis in cultured mammalian
cells. Cell. 17:319–325. 1979.
|
|
27
|
Wehland J, Henkart M, Klausner R and
Sandoval IV: Role of microtubules in the distribution of the Golgi
apparatus: effect of taxol and microinjected anti-alpha-tubulin
antibodies. Proc Natl Acad Sci USA. 80:4286–4290. 1983.
|
|
28
|
Sharp JA, Plant JJ, Ohsumi TK, Borowsky M
and Blower MD: Functional analysis of the microtubule-interacting
transcriptome. Mol Biol Cell. 22:4312–4323. 2011.
|
|
29
|
Sakaue-Sawano A, Ohtawa K, Hama H, Kawano
M, Ogawa M and Miyawaki A: Tracing the silhouette of individual
cells in S/G2/M phases with fluorescence. Chem Biol. 15:1243–1248.
2008.
|
|
30
|
Sakaue-Sawano A, Kurokawa H, Morimura T,
et al: Visualizing spatiotemporal dynamics of multicellular
cell-cycle progression. Cell. 132:487–498. 2008.
|