Introduction
Osteosarcoma is one of the most common types of
primary malignant cancer in bone tissues, primarily affecting
adolescents and young adults (1).
The majority of osteosarcomas occur in the metaphysis of the long
bones of the extremities, whereas the involvement of the axial or
the craniofacial bone can also be observed in the adult population
(2). Despite the progress that has
been made in treatment strategies for osteosarcoma, including
surgical resection, chemotherapy and radiotherapy, the clinical
prognosis remains poor, and the 5-year survival rate is still
<20% due to the high rates of metastasis and recurrence
(3,4). Although substantial attention has
focused on the molecular mechanism behind osteosarcoma, the factors
behind its occurrence and development remain unclear. Therefore,
there is an urgent need to reveal the molecular mechanism of the
development of osteosarcoma, to identify new molecular markers for
early diagnosis, and to investigate new methods for treating this
disease.
Compared with normal cells, tumor cells tend to
undergo glycolysis despite the environment having sufficient
oxygen; this phenomenon was first reported by Otto Warburg and is
termed the Warburg effect (5).
Through this metabolic rearrangement, tumor cells obtain enough
energy to meet the requirements for growth and distant metastasis
(6,7). Several oncogenes that regulate the
Warburg effect and promote the development of osteosarcoma have
been reported. For example, sphingosine-1-phosphate has been
demonstrated to regulate the glycolytic enzyme phosphoglycerate
mutase 1 to promote osteosarcoma proliferation (8). In addition, slit guidance ligand 2 has
been reported to enhance the expression of
6-phosphofructo-2-kinase/fructose-2,6-biphosphatase 2 and to
increase osteosarcoma cell proliferation and migration (9). Therefore, targeting the glycolytic
pathway may be an appropriate strategy for the treatment of
osteosarcoma.
Keratins, one of the largest gene families in
humans, are a family of epithelial-specific intermediate filaments
forming an indispensable part of the cytoskeleton (10). Keratins can be classified into two
types, including 28 type I and 26 type II keratins (11). Keratins not only provide structural
support for the cytoskeleton and regulate cellular metabolic
processes, but also activate epithelial cell growth signaling
pathways (12). Their expression is
highly cell-type-specific, and mutation or loss of a keratin gene
can cause or lead to a predisposition to several human diseases,
induce the dysregulation of innate immunity, and produce increased
sensitivity to apoptosis (13).
Extensive research has been performed on the role of the keratin
family in human cancer, which has attracted broad attention in the
last two decades. For example, keratin 13 serves a role in
reprogramming bone and brain metastases of human prostate cancer
cells (14). In addition, keratin 5
and keratin 20 may be associated with urothelial bladder cancer
(15). Keratin 17 (KRT17), a type I
keratin, has also been identified to be expressed in several types
of normal human epithelium, including the respiratory epithelium
and urothelium (16). KRT17 was
first reported to serve a positive role in epithelial cell
proliferation and skin tumor growth (17). Furthermore, KRT17 has been reported
to be associated with the development of various types of cancer,
such as colorectal and breast cancer (18). However, its role in the progression
of osteosarcoma remains unclear.
The present study aimed to investigate the role of
KRT17 and its mechanism in osteosarcoma, and to determine whether
KRT17 may be a novel target for the diagnosis and treatment of
osteosarcoma.
Materials and methods
Patient samples
A total of 25 patients were enrolled in the present
study. All patients were pathologically diagnosed with osteosarcoma
and had received surgical resection at Guizhou Orthopedics Hospital
(Guizhou, China) between June 2017 and June 2019. The patients
included 17 males and 8 females, with a mean age of 19.15±13.17
years (range, 8–41 years). The inclusion criteria were as follows:
i) The tissues were obtained during surgery and osteosarcoma was
diagnosed by two pathologists; ii) the patients were diagnosed and
treated for the first time; and iii) the patients were willing to
participate. The exclusion criteria were as follows: i) Patients
with other malignancies; ii) patients with other systemic diseases;
iii) patients who had received treatment prior to admission; and
iv) patients (and/or their families) who refused to participate.
All 25 patients provided osteosarcoma tissue samples, of whom 10
also provided adjacent normal tissues. All specimens were obtained
from patients following written informed consent form the patients
or their parents/legal guardians. The study was approved by Guizhou
Provincial Orthopedics Hospital Ethics Committee and was performed
in accordance with the principles embodied in the Declaration of
Helsinki.
Cell lines and cell culture
Human osteosarcoma cell lines (MG63, Saos2, U20S and
143B) and normal human osteoblast hFOB cells were obtained from the
Cell Bank of the Chinese Academy of Sciences. The cells were
cultured in Dulbecco's modified Eagle's medium (Gibco; Thermo
Fisher Scientific, Inc.) containing 10% fetal bovine serum (Gibco;
Thermo Fisher Scientific, Inc.), 0.1% penicillin and 0.1%
streptomycin. MG63 (fibroblast morphology), Saos2 (epithelial
morphology), U20S (epithelial morphology) and 143B (mixed
morphology) cells were incubated at 37°C in a humidified atmosphere
with 5% CO2, whereas hFOB cells were incubated at 35°C
under the same atmosphere conditions.
Reverse transcription-quantitative PCR
(RT-qPCR)
TRIzol reagent [Yeasen Biotechnology (Shanghai) Co.,
Ltd.] was used to isolate total RNA from osteosarcoma tissues,
adjacent tissues, normal human osteoblast cells and osteosarcoma
cells. PrimeScript RT Reagent kit reagent (Takara Bio, Inc.) was
used to perform complementary DNA synthesis according to the
manufacturer's instructions. Hieff® qPCR
SYBR® Green Master Mix [Yeasen Biotechnology (Shanghai)
Co., Ltd.] was used for the fluorescence quantification. The
following primers were used: KRT17 forward,
5′-GGTGGGTGGTGAGATCAATGT-3′ and reverse,
5′-CGCGGTTCAGTTCCTCTGTC-3′; VEGF forward,
5′-AGGGCAGAATCATCACGAAGT-3′ and reverse,
5′-AGGGTCTCGATTGGATGGCA-3′; glucose transporter 1 (GLUT1) forward,
5′-GGCCAAGAGTGTGCTAAAGAA-3′ and reverse,
5′-ACAGCGTTGATGCCAGACAG-3′; MCL1 apoptosis regulator, Bcl2 family
member (MCL1) forward, 5′-TGCTTCGGAAACTGGACATCA-3′ and reverse,
5′-TAGCCACAAAGGCACCAAAAG-3′; HIF1α forward,
5′-GAACGTCGAAAAGAAAAGTCTCG-3′ and reverse,
5′-CCTTATCAAGATGCGAACTCACA-3′; β-actin forward,
5′-CATGTACGTTGCTATCCAGGC-3′ and reverse,
5′-CTCCTTAATGTCACGCACGAT-3′. The reaction conditions were as
follows: 95°C for 10 min, followed by 40 cycles of 95°C for 1 sec
and 60°C for 60 sec. β-actin was used for normalization to
determine the relative mRNA expression, which was analyzed using
the 2−ΔΔCq method (19).
Western blotting
RIPA lysis buffer (Boster Biological Technology)
containing a mixture of protease inhibitors was used to extract the
proteins. The lysates were centrifuged at 8,000 × g at 4°C for 10
min to separate the proteins from the cell debris. The BCA assay
kit (Boster Biological Technology) was used to measure the total
protein concentration. The proteins (30 µg/lane) were separated by
10% SDS-PAGE and then transferred to PVDF membranes (EMD
Millipore). Subsequently, 5% non-fat milk was used for blocking at
room temperature for 2 h, after which the membranes were incubated
overnight at 4°C with the following primary antibodies: Anti-KRT17
(1:1,000, cat no. 17516-1-AP), anti-cyclin D1 (1:1,000, cat no.
26939-1-AP), anti-CDK2 (1:1,000, cat no. 10122-1-AP), anti-CDK4
(1:1,000, cat no. 11026-1-AP, ProteinTech Group, Inc.), anti-CDK6
(1:1,000, cat no. 14052-1-AP, ProteinTech Group, Inc.), anti-AKT
(1:1,000, cat no. 10176-2-AP), anti-phosphorylated (p)-AKT
(1:1,000, cat no. 66444-1-Ig), anti-mTOR (1:1,000, cat no.
66888-1-Ig), anti-HIF1α (1:1,000, cat no. 20960-1-AP), anti-VEGF
(1:1,000, cat no. 19003-1-AP), anti-GLUT1 (1:1,000, cat no.
21829-1-AP), anti-MCL1 (1:1,000, cat no. 16225-1-AP), anti-β-actin
(1:1,000, cat no. 60008-1-Ig) (all from ProteinTech Group, Inc.)
and anti-p-mTOR (1:500, cat no. 5536, Cell Signaling Technology,
Inc.). After three washes with TBS containing 0.1% Tween-20, the
membranes were incubated with the secondary horseradish peroxidase
(HRP)-conjugated anti-mouse (1:3,000; cat no. BA1051) and
anti-rabbit (1:3,000; cat no. BA1050; both from Boster Biological
Technology) antibodies for 2 h at room temperature. Finally,
protein bands were visualized using ChemiDoc™ XRS+ with Image Lab™
software (version 5.2.1; Bio-Rad Laboratories, Inc.). The
expression levels of KRT17, cyclin D1, CDK2, CDK4, CDK6, HIF1α,
VEGF, MCL1 and GLUT1 were relative to β-actin, whereas those of
p-AKT and p-mTOR were analyzed as the ratios of p-AKT/AKT and
p-mTOR/mTOR.
Retroviral infection and
transfection
Human short hairpin RNA (shRNA) targeting KRT17 and
negative control shRNA oligonucleotide sequences were cloned into
pSuper-retro-puro viral vectors (Guangzhou RiboBio Co., Ltd.) to
generate pSuper-retro-KRT17-RNAi(s) and
pSuper-retro-scramble-RNAi(s). The shRNA sequences for KRT17 were
as follows: shRNA1, 5′-GCGTGACCAGTATGAGAAGAT-3′; shRNA2,
5′-TGGTGCAGAGTGGCAAGAGTGAGAT-3′; and shRNA3,
5′-CTGACTCAGTACAAGAAAGAA-3′. The scrambled shRNA sequence
5′-wasTTCTCCGAACGTGTCACGT-3′ (all synthesized by Guangzhou RiboBio
Co., Ltd). To obtain stable knockdown of KRT17 osteosarcoma cell
lines, the cells were cultured in an environment containing 0.5
µg/ml puromycin for 2 weeks after infection. Osteosarcoma cells
with the optimal stable knockdown of KRT17 and normal control
(sh-scramble) were used for further study.
Cell Counting Kit-8 (CCK-8) assay
MG63 and Saos-2 cells were seeded in a 96-well plate
at a density of 4×103 cells/well in a volume of 200 µl.
On days 0, 1, 2, 3 and 4, the cells were incubated with CCK-8
(Boster Biological Technology) for 2 h at 37°C, and absorbance was
measured at 450 nm using a microplate reader. For the rescue
experiment, the AKT activator SC79 (5 µg/ml; MedChemExpress), mTOR
activator MYH1485 (10 µM; MedChemExpress) and HIF activator ML228
(5 µM; MedChemExpress) were added after cell adherence, and the
absorbance was measured at 450 nm after culturing for the time
stated above.
Colony formation
MG63 and Saos-2 cells were seeded in a 6-well plate
at a density of 1,000 cells/well. After culturing for 7 days at
37°C in a humidified incubator with 5% CO2, 4%
paraformaldehyde was used to fix the cells for 15 min at room
temperature, which were then stained with 0.1% crystal violet for
30 min at room temperature. Images of stained colonies were
captured, and the number of colonies (≥10 mm2) was
counted under the naked eye.
Cell cycle analysis
MG63 and Saos-2 cells were washed with PBS three
times and fixed with 75% ethanol overnight at −20°C. The cells were
then treated with 50 µg/ml propidium iodide solution (Invitrogen;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
instructions and analyzed using a FACSCalibur flow cytometer (BD
Biosciences). The results of the cell cycle distribution were
analyzed using FlowJo software version 7.4.1 (FlowJo LLC).
Assays for measuring the extracellular
acidification rate (ECAR) and cellular oxygen consumption rate
(OCR)
The XFe Extracellular Flux Analyzer (Agilent
Technologies, Inc.) was used to detect the ECAR and OCR in the
present study. The ECAR was detected using an XF Glycolysis Stress
Test kit (Agilent Technologies. Inc.), and the OCR was detected
using an XF Cell Mito Stress Test kit. A total of 1×104
MG63 and Saos-2 cells were seeded in matched 96-well microplates.
Following baseline measurements, 30 mM glucose, 2 µM oligomycin (an
inhibitor of oxidative phosphorylation; Selleck Chemicals) and 10
mM 2-DG (an inhibitor of glycolysis; Selleck Chemicals) were added
to each well at the specified time points to detect the ECAR.
Similarly, 2 µM oligomycin, 5 µM FCCP (a reversible inhibitor of
oxidative phosphorylation; Selleck Chemicals) and 1 µM rotenone (an
inhibitor of mitochondrial complex I; Selleck Chemicals) plus 1 µM
antimycin A (an inhibitor of mitochondrial complex III; Selleck
Chemicals) were added in the appropriate order to detect the OCR
and ECAR according to the operating steps described in a previous
study (20). The data were
collected and analyzed using XF-96 Wave software version 2.1
(Agilent Technologies. Inc.). The data were normalized based on the
protein concentration.
Detection of glucose uptake, lactate
production and ATP synthesis
To detect glucose uptake in each group, the Glucose
Uptake Colorimetric assay kit (Invitrogen; Thermo Fisher
Scientific, Inc.) was used according to the manufacturer's
instruction. A total of 2,000 MG63 and Saos-2 cells were seeded in
a 96-well plate and treated with the Krebs-Ringer-Phosphate-HEPES
buffer containing 2% BSA (Wuhan Boster Biological Technology, Ltd.)
for 40 min to starve the cells. Subsequently, 10 ml 2-DG (10 mM)
was added to each well. Following culture for 20 min, the rate of
glucose uptake was determined. To determine the lactate production
and ATP synthesis of each group, Lactate assay kit II (Invitrogen;
Thermo Fisher Scientific, Inc.) and ATP assay kit (Invitrogen;
Thermo Fisher Scientific, Inc.), respectively, were used according
to the manufacturer's instructions. A total of 2×106
MG63 and Saos-2 cells were lysed in 100 µl of the corresponding
assay buffer and homogenized. Following centrifugation, the soluble
fractions of lactate and ATP were assayed using 530 and 636 nm
wavelengths in an ultraviolet spectrophotometer, respectively.
Animal experiments
All animal procedures were approved by the Ethics
Committee of Guizhou Provincial Orthopedics Hospital. Female BLBA/c
nude mice (6-week-old; n=10) were purchased from Beijing Huafukang
Biotechnology Co., Ltd. and housed under specific pathogen-free
conditions at 25°C with a 12-h light/dark cycle and free access to
food and water. A total of 1×106 MG63 cells transfected
with sh-KRT17 or sh-scramble were subcutaneously injected into the
flanks of nude mice. Each group contained five mice. The animal
health and behavior were monitored daily, and the volume of tumor
tissue was measured weekly. The volume of tumor tissues were
calculated using the following formula: Volume
(mm3)=(length × width2)/2. When the maximum
length of any tumor reached 15 mm or the volume of any tumor
reached 800 mm3, the experiment was terminated. The mice
were sacrificed in week 5 using cervical dislocation; death was
verified by the absence of a heartbeat and the onset of rigor
mortis, and the size of each tumor was determined.
Immunohistochemical staining
The tumor tissues isolated from nude mice were
dehydrated and embedded in paraffin (Wuhan Boster Biological
Technology, Ltd.) at room temperature. After cutting into 4-µm
slices, tissue sections were deparaffinized using xylene and
rehydrated in a descending alcohol series (100, 95, 90 and 80%) at
room temperature. Following restoration with sodium citrate, the
samples were treated with 3% H2O2 to block
endogenous peroxidase activity, and then blocked with 5% BSA (Wuhan
Servicebio Technology Co., Ltd.) for 30 min at room temperature.
The specimens were subsequently incubated with a primary anti-KI67
(1:400; cat. no. 27309-1-AP; ProteinTech Group, Inc.) and anti-PCNA
(1:400; cat. no. 10205-2-AP; ProteinTech Group, Inc.) antibodies
for 12 h at 4°C, followed by incubation with HRP-conjugated
secondary antibodies anti rabbit IgG (1:200; cat. no.
G1210-2-A-100; Wuhan Servicebio Technology Co., Ltd.) for 2 h at
room temperature. Following subsequent development using the Cell
and Tissue Staining HRP-DAB kit (Beyotime Institute of
Biotechnology) according to the manufacturer's protocol, images
were obtained under a light microscope (magnification, ×400).
Statistical analysis
SPSS 20.0 (IBM Corp.) was used to perform the
statistical analysis in the current study. Data are representative
of at least three independent experiments and presented as the mean
± standard deviation. Unpaired Student's t-test was used to analyze
the expression difference of KRT17 in osteosarcoma and
corresponding adjacent tissues. The co-expression relationship
between KRT17 and HIF1α in osteosarcoma tissues was analyzed using
Pearson's correlation analysis. Two-tailed unpaired Student's
t-test was used to analyze the statistical differences between two
different groups of continuous variables. One-way analysis of
variance (ANOVA) followed by Dunnett's post hoc test was performed
to test the differences between control group and multiple
experimental groups, while ANOVA followed by Bonferroni's post hoc
test was used to test the differences among multiple groups.
P<0.05 was considered to indicate a statistically significant
difference.
Results
KRT17 is expressed in osteosarcoma
tissues and cell lines
First, RT-qPCR was performed to analyze the
expression of KRT17 in osteosarcoma tissues. The results
demonstrated that the expression of KRT17 was significantly higher
in osteosarcoma tissues (n=25) compared with in adjacent normal
osteoblast tissues (n=10) (P<0.01; Fig. 1A). Similarly, the mRNA and protein
levels of KRT17 were markedly increased in osteosarcoma lines,
including MG63, Saos2, U2OS and 143B, compared with those in normal
bone hFOB cells (P<0.01; Fig.
1B-D). The mRNA and protein levels of KRT17 were 2–3-fold
higher in the MG63 and Saos2 cell lines compared with those in hFOB
cells; therefore, these two cell lines were used for further study.
143B cells were excluded due to their mixed morphology resulting in
large experimental error.
Knockdown of KRT17 decreases
osteosarcoma cell proliferation and induces G1 phase
arrest in vitro
To determine the role of KRT17 in osteosarcoma
proliferation in vitro, its expression was disrupted using
targeted KRT17 shRNA. Three potential KRT17-targeting shRNAs were
used, and the results demonstrated that sh-KRT17-3 exhibited the
strongest inhibition of KRT17 expression compared with the
sh-scramble group (P<0.01; Fig. 2A
and B). Therefore, sh-KRT17-3 was used to construct
osteosarcoma cells with stable KRT17 knockdown for further
experiments. CCK-8 assays were performed, and the results revealed
that KRT17 knockdown significantly decreased the proliferation rate
of MG63 and Saos2 cells compared with those transfected with
sh-scramble (P<0.01; Fig. 2C and
D). Similarly, it was identified that knockdown of KRT17
significantly decreased the numbers of MG63 and Saos2 cell colonies
compared with those in the corresponding sh-scramble groups
(P<0.01; Fig. 2E and F). Cell
cycle distribution analysis was performed to detect the differences
between the sh-scramble and sh-KRT17 groups; the results
demonstrated that the proportion of cells in the G1
phase significantly increased in the sh-KRT17 groups of both MG63
and Saos2 cells compared with the respective sh-scramble groups,
and a significant decrease of cells in the G2 phase was
observed in the sh-KRT17 groups (P<0.05; Fig. 2G and H). In addition, the expression
levels of cyclin D1, CDK2, CDK4 and CDK6 were examined in the
sh-KRT17 and sh-scramble groups; it was observed that inhibition of
KRT17 decreased the expression of cyclin D1, CDK2, CDK4 and CDK6
(Fig. 2I). These results indicated
that knockdown of KRT17 decreased osteosarcoma cell proliferation
and induced G1 phase arrest in vitro.
Knockdown of KRT17 decreases the
Warburg effect of osteosarcoma cells
Previous studies have demonstrated that the Warburg
effect serves a key role in the proliferation of osteosarcoma cells
(21,22). Therefore, the present study
investigated the effect of KRT17 on the Warburg effect of
osteosarcoma cells. OCR, ECAR, glucose uptake, lactate production
and ATP synthesis were analyzed in the sh-scramble and sh-KRT17
groups. The ECAR was significantly decreased in the sh-KRT17 group
in both MG63 (at 30, 35 and 40 min) and Saos-2 cells (at 35 and 40
min) (P<0.05; Fig. 3A and B),
whereas the basal and maximal OCR were significantly increased in
the sh-KRT17 group compared with the sh-scramble group (P<0.05;
Fig. 3C and D). In addition, the
glucose uptake rate, lactate production and ATP synthesis were
significantly decreased in MG63 and Saos-2 cells following KRT17
knockdown compared with the sh-scramble-transfected cells
(P<0.05; Fig. 3E-J). Taken
together, these results indicated that knockdown of KRT17 decreased
the Warburg effect in osteosarcoma cells.
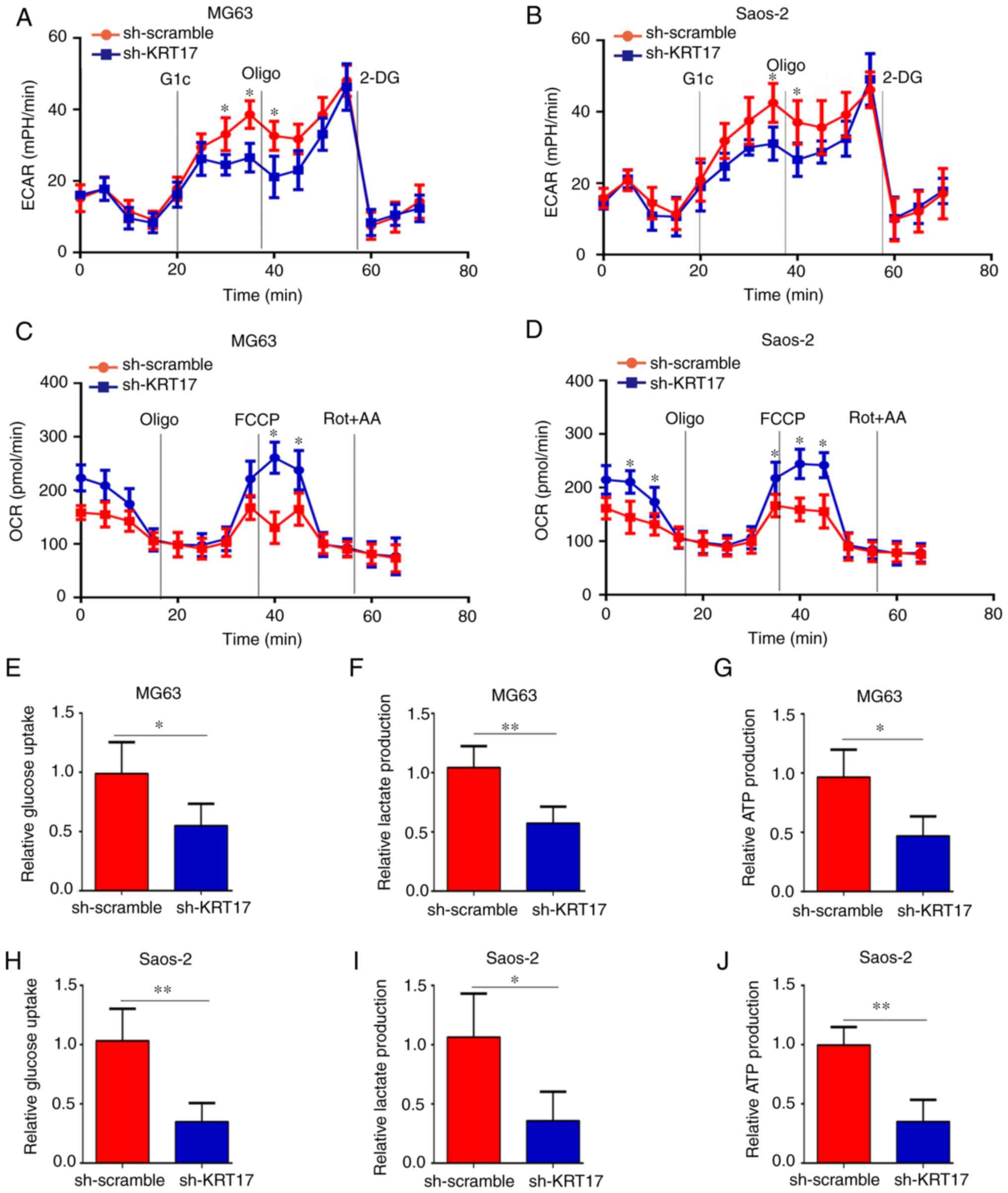 | Figure 3.Knockdown of KRT17 decreases the
Warburg effect of osteosarcoma cells. (A and B) Using a Seahorse
Bioscience XFp analyzer, the ECAR of MG63 and Saos2 cells in the
sh-KRT17 and sh-scramble groups was measured. (C and D) Using a
Seahorse Bioscience XFp analyzer, the OCR of MG63 and Saos2 cells
with stable knockdown of KRT17 and sh-scramble cells was measured.
(E-G) Glucose uptake, lactate production and ATP synthesis were
measured in MG63 cells with stable knockdown of KRT17 and those
transfected with sh-scramble. (H-J) Glucose uptake, lactate
production and ATP synthesis were measured in Saos2 sh-KRT17 and
sh-scramble cells. *P<0.05, **P<0.01 vs. sh-scramble. Glc,
glucose; Oligo, oligomycin; 2-DG, 2-deoxy-D-glucose; FCCP, carbonyl
cyanide 4-(trifluoromethoxy) phenylhydrazone; Rot, rotenone; AA,
antimycin A; KRT17, keratin 17; ECAR, extracellular acidification
rate; OCR, cellular oxygen consumption rate. |
Knockdown of KRT17 decreases tumor
growth in vivo
The effect of KRT17 on tumor growth in vivo
was determined using a subcutaneous tumorigenesis model in nude
mice. The results revealed that the rates of tumor growth were
slower and the weights of the tumor were lower in the sh-KRT17
group compared with those in the sh-scramble group (P<0.01;
Fig. 4A-C). In addition, the
expression levels of KI67 and PCNA in tumor tissues from the
sh-KRT17 group were significantly decreased compared with those in
tissues from the sh-scramble group. (P<0.05; Fig. 4D). In summary, these results
suggested that knockdown of KRT17 decreased osteosarcoma tumor
growth in vivo.
KRT17 regulates the AKT/mTOR/HIF1α
pathway
A previous study has demonstrated that KRT17 binds
to stratifin and recruits it to the cytoplasm, where this complex
activates the AKT/mTOR pathway (13). To determine whether KRT17 also
regulates the AKT/mTOR pathway in osteosarcoma, western blotting
was performed. The results demonstrated that total AKT and total
mTOR levels changed insignificantly (P>0.05), whereas p-AKT and
p-mTOR levels were significantly decreased in the sh-KRT17 group
compared with those in the sh-scramble group (Fig. 5A-C). HIF1α is a key downstream
protein of mTOR, which has been reported to serve a major role in
cell proliferation and metabolic rearrangement, such as by
increasing glycolysis (23).
Therefore, the present study determined the expression of HIF1α
following KRT17 inhibition and revealed that its level was also
significantly decreased in the sh-KRT17 group compared with that in
the sh-scramble group (P<0.05; Fig.
5A-C). Of note, correlation analysis revealed that KRT17 was
co-expressed with HIF1α in osteosarcoma tissues (r=0.61, P<0.01;
Fig. 5D). Furthermore, the
expression levels of HIF1α target genes, such as VEGF, GLUT1 and
MCL1, were determined, which demonstrated that the mRNA and protein
levels of these genes were significantly decreased following KRT17
inhibition compared with those in the sh-scramble group (P<0.05;
Fig. 5E-I). Among these target
genes of HIF1α, GlUT1, which serves an important role in
glycolysis, exhibited the strongest decrease.
 | Figure 5.Inhibition of KRT17 suppresses the
AKT/mTOR/HIF1α pathway. (A-C) The expression of AKT, p-AKT, mTOR,
p-mTOR and HIF1α in the sh-scramble and sh-KRT17 groups was
detected using western blotting. β-actin was used as a loading
control. (D) The co-expression relationship between KRT17 and HIF1α
in osteosarcoma tissues was determined using RT-qPCR and Pearson's
correlation analysis. (E and F) The mRNA levels of VEGF, GLUT1 and
MCL1 were determined using RT-qPCR in the sh-scramble and sh-KRT17
groups. (G-I) The protein levels of VEGF, GLUT1 and MCL1 were
detected using western blotting in the sh-scramble and sh-KRT17
groups. *P<0.05, **P<0.01. KRT17, keratin 17; sh, short
hairpin; p-, phosphorylated; HIF1α, hypoxia-inducible factor 1α;
MCL1, MCL1 apoptosis regulator, Bcl2 family member; GLUT1, glucose
transporter 1. |
Restoration of the AKT/mTOR/HIF1α
pathway decreases the inhibitory effect of KRT17 knockdown on
osteosarcoma cell proliferation
To detect whether the AKT/mTOR/HIF1α pathway was
involved in the KRT17 knockdown-induced inhibition of
proliferation, activators of AKT (SC79), mTOR (MYH1485) and HIF1α
(ML228) were employed. The results demonstrated that the expression
of GLUT1 was increased when cells in the sh-KRT17 group were
treated with SC79, MYH1485 or ML228 compared with the untreated
sh-KRT17 group (Fig. 6A and B). The
results of the CCK-8 assay revealed that the inhibitory effect of
KRT17 knockdown on cell proliferation was reversed by SC79, MYH1485
or ML228 in MG63 and Saos2 cells (P<0.05; Fig. 6C and D). Similarly, the number of
colonies was significantly increased when sh-KRT17 cells were
treated with SC79, MYH1485 or ML228 compared with that in untreated
sh-KRT17 cells (Fig. 6E and F).
These results indicated that restoration of the AKT/mTOR/HIF1α
pathway decreased the inhibitory effect of KRT17 knockdown on
osteosarcoma cell proliferation.
 | Figure 6.Restoration of the AKT/mTOR/HIF1α
pathway reverses the effects of KRT17 knockdown on osteosarcoma
cell proliferation and colony formation. (A and B) Western blotting
was used to detect the expression of KRT17, AKT, p-AKT, mTOR,
p-mTOR, HIF1α and GLUT1 in cells treated with sh-scramble or
sh-KRT17 and SC79, MYH1485 and ML228. (C and D) Cell Counting Kit-8
assay was used to detect the cell proliferation in each group. (E
and F) Colony formation assays were used to detect the colony
formation of osteosarcoma cells in each group. **P<0.01 vs.
sh-scramble; #P<0.05, ##P<0.01 vs.
sh-KRT17. KRT17, keratin 17; sh, short hairpin; p-, phosphorylated;
HIF1α, hypoxia-inducible factor 1α; GLUT1, glucose transporter
1. |
Restoration of the AKT/mTOR/HIF1α
pathway decreases the inhibitory effect of KRT17 knockdown on
osteosarcoma cell glycolysis
The association between KRT17, the AKT/mTOR/HIF1α
pathway and glycolysis was also investigated in the present study.
In the ECAR analysis, the ECAR at 40 and 45 min was increased in
sh-KRT17 cells upon treatment with activators of the AKT/mTOR/HIF1α
pathway compared with untreated sh-KRT17 cells (P<0.05; Fig. 7A and B). In addition, the maximal
OCR at 40 and 45 min was decreased in sh-KRT17 cells upon treatment
with activators of the AKT/mTOR/HIF1α pathway compared with
untreated sh-KRT17 cells (P<0.05; Fig. 7C and D). The rates of glucose
uptake, lactate production and ATP synthesis were significantly
increased when sh-KRT17 cells were treated with AKT, mTOR or HIF1α
activators compared with those in untreated sh-KRT17 cells
(P<0.05; Fig. 7E-J). These
results indicated that restoration of the AKT/mTOR/HIF1α pathway
decreased the inhibitory effect of KRT17 knockdown on osteosarcoma
cell glycolysis.
 | Figure 7.Restoration of the AKT/mTOR/HIF1α
pathway decreases the inhibitory effect of KRT17 knockdown on
osteosarcoma cell glycolysis. (A and B) The ECAR was detected in
MG63 and Saos2 cells treated with sh-scramble or sh-KRT17 and SD79,
MYH1485 and ML228. (C and D) The OCR was detected in each group of
MG63 and Saos2 cells. (E-G) Glucose uptake, lactate production and
ATP synthesis were measured in each group of MG63 cells. (H-J)
Glucose uptake, lactate production and ATP synthesis were measured
in each group of Saos2 cells. *P<0.05, **P<0.01 vs.
sh-scramble; #P<0.05, ##P<0.01 vs.
sh-KRT17. Glc, glucose; Oligo, oligomycin; 2-DG, 2-deoxy-D-glucose;
FCCP, carbonyl cyanide 4-(trifluoromethoxy) phenylhydrazone; Rot,
rotenone; AA, antimycin A; KRT-17, keratin 17; sh, short hairpin;
HIF1α, hypoxia-inducible factor 1α; ECAR, extracellular
acidification rate; OCR, cellular oxygen consumption rate. |
Discussion
The present study identified the role of KRT17 in
osteosarcoma and the underlying molecular mechanism. KRT17 is a
member of the keratin family that serves a key role in protecting
cells from damage or physical stress (24). Dysregulated expression of KRT17 has
been reported in numerous types of cancer. Hu et al
(25) have suggested that KRT17 is
highly expressed in gastric cancer and is associated with poor
outcome in those affected by this disease. In addition, Liu et
al (26) have demonstrated that
KRT17 has the potential to promote the proliferation, migration and
invasion of lung adenocarcinoma cells. Khanom et al
(13) have reported that the
inhibition of KRT17 decreases the proliferation of oral cancer
cells. Furthermore, Li et al (27) have demonstrated that KRT17 serves a
key role in the resistance to paclitaxel in cervical cancer cells.
Consistent with these previous studies, the present study
demonstrated that KRT17 is increased in osteosarcoma cell tissues
and cell lines. Knockdown of KRT17 significantly decreased the
proliferation of osteosarcoma cells in vitro and in
vivo. These results indicate that KRT17 may act as an oncogene
in osteosarcoma.
Glycolysis is a common hallmark for cancer tissues
as cancer cells utilize energy via glycolysis rather than by the
tricarboxylic acid cycle (21).
Based on glycolysis, cancer cells have enough energy for
proliferation, migration and metastasis (28). The results of the present study
demonstrated that inhibition of KRT17 significantly increased the
OCR and decreased the ECAR, ATP production, lactate production and
glucose uptake of osteosarcoma cells compared with those in the
control group.
Previous studies have reported that the AKT/mTOR
pathway is activated in various types of cancer, including
osteosarcoma (29,30). Activated mTOR promotes cell
proliferation by promoting the phosphorylation of downstream
proteins (31). A previous study
has demonstrated that KRT17 can bind with stratifin and increase
the phosphorylation level of AKT (13). In Ewing's sarcoma, KRT17 has also
been reported to have the capacity to activate the AKT pathway
(32). Therefore, the present study
determined the expression of proteins in the AKT pathway, with the
results revealing that the levels of p-AKT and p-mTOR were
decreased in KRT17-knockdown cells compared with those in the
normal control group.
HIF1α is one of the downstream proteins of mTOR
(33). Previous studies have
demonstrated that activated mTOR can maintain the stability of
HIF1α (34,35). Increased HIF1α translocates into the
nucleus and binds to the promoters of its target genes, such as
VEGF, GLUT1 and MCL1 (36–38). Through the regulation of its target
genes, HIF1α serves roles in cancer cell proliferation,
angiopoiesis and glycolysis (39).
Based on the significant effects of KRT17 on osteosarcoma
glycolysis, the present study considered whether HIF1α was
regulated by KRT17 via the AKT/mTOR pathway; consistent with this
speculation, it was identified that the expression of HIF1α was
significantly decreased in sh-KRT17 osteosarcoma cells, as was that
of its target genes, such as VEGF, MCL1 and GLUT1. Among these,
GLUT1, which serves a key role in cell glycolysis, was decreased
the most significantly. In addition, the results of the correlation
analysis demonstrated that KRT17 was co-expressed with HIF1α. In
summary, these results indicate that there may be a regulatory
relationship between KRT17 and HIF1α via the AKT/mTOR pathway. To
confirm these conclusions, AKT, mTOR and HIF1α activators were
used, and the results demonstrated that the restoration of the
AKT/mTOR/HIF1α pathway reversed the effects of KRT17 knockdown on
osteosarcoma cell function, including proliferation and
glycolysis.
In conclusion, the results of the present study
demonstrated that KRT17 was highly expressed in osteosarcoma
tissues and osteosarcoma cell lines. Knockdown of KRT17 decreased
osteosarcoma cell proliferation and glycolysis by inhibiting the
AKT/mTOR/HIF1α pathway. Therefore, KRT17 may be a novel biomarker
for osteosarcoma diagnosis, as well as an effective target for
treatment.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XY, CY and WH performed the experiments and were
responsible for data collection, analysis and interpretation of the
results. XY, TC, QW, BQ and FP provided clinical samples for the
experiments and validated the data. XY and BT were responsible for
the experimental design, analysis and interpretation of data. All
authors have read and approved the final version of the
manuscript.
Ethics approval and consent to
participate
The study of clinical samples was approved by the
Ethics Committee of Guizhou Provincial Orthopedics Hospital and was
performed in accordance with the principles embodied in the
Declaration of Helsinki. All patients provided written informed
consent to participate in the study. All animal procedures were
approved by the Ethics Committee of Guizhou Provincial Orthopedics
Hospital.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Melim C, Jarak I, Veiga F and Figueiras A:
The potential of micelleplexes as a therapeutic strategy for
osteosarcoma disease. 3 Biotech. 10:1472020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Moore DD and Luu HH: Osteosarcoma. Cancer
Treat Res. 162:65–92. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bishop MW, Janeway KA and Gorlick R:
Future directions in the treatment of osteosarcoma. Curr Opin
Pediatr. 28:26–33. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fan TM, Roberts RD and Lizardo MM:
understanding and modeling metastasis biology to improve
therapeutic strategies for combating osteosarcoma progression.
Front Oncol. 10:132020. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Baltazar F, Afonso J, Costa M and Granja
S: Lactate beyond a waste metabolite: Metabolic affairs and
signaling in malignancy. Front Oncol. 10:2312020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Liberti MV and Locasale JW: Correction to:
‘The warburg effect: How does it benefit cancer cells?’: [Trends in
Biochemical Sciences, 41 (2016) 211]. Trends Biochem Sci.
41:2872016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Schwartz L, Supuran CT and Alfarouk KO:
The warburg effect and the hallmarks of cancer. Anticancer Agents
Med Chem. 17:164–170. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shen Y, Zhao S, Wang S, Pan X, Zhang Y, Xu
J, Jiang Y, Li H, Zhang Q, Gao J, et al: S1P/S1PR3 axis promotes
aerobic glycolysis by YAP/c-MYC/PGAM1 axis in osteosarcoma.
Ebiomedicine. 40:210–223. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhao SJ, Shen YF, Li Q, He YJ, Zhang YK,
Hu LP, Jiang YQ, Xu NW, Wang YJ, Li J, et al: SLIT2/ROBO1 axis
contributes to the Warburg effect in osteosarcoma through
activation of SRC/ERK/c-MYC/PFKFB2 pathway. Cell Death Dis.
9:3902018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Rajabi M, Ali A, McConnell M and Cabral J:
Keratinous materials: Structures and functions in biomedical
applications. Mater Sci Eng C, Mater Biol Appl. 110:1106122020.
View Article : Google Scholar
|
|
11
|
Donato RK and Mija A: Keratin associations
with synthetic, biosynthetic and natural polymers: An extensive
review. Polymers (Basel). 12(pii): E322019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yi H, Yoon HN, Kim S and Ku NO: The role
of keratins in the digestive system: Lessons from transgenic mouse
models. Histochem Cell Biol. 150:351–359. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Khanom R, Nguyen CT, Kayamori K, Zhao X,
Morita K, Miki Y, Katsube K, Yamaguchi A and Sakamoto K: Keratin 17
Is induced in oral cancer and facilitates tumor growth. PLoS One.
11:e1611632016. View Article : Google Scholar
|
|
14
|
Li Q, Yin L, Jones LW, Chu GC, Wu JB,
Huang JM, Li Q, You S, Kim J, Lu YT, et al: Keratin 13 expression
reprograms bone and brain metastases of human prostate cancer
cells. Oncotarget. 7:84645–84657. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Eckstein M, Wirtz RM, Gross-Weege M,
Breyer J, Otto W, Stoehr R, Sikic D, Keck B, Eidt S, Burger M, et
al: mRNA-Expression of KRT5 and KRT20 defines distinct prognostic
subgroups of muscle-invasive urothelial bladder cancer correlating
with histological variants. Int J Mol Sci. 19(pii): E33962018.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li D, Ni XF, Tang H, Zhang J, Zheng C, Lin
J, Wang C, Sun L and Chen B: KRT17 Functions as a tumor promoter
and regulates proliferation, migration and invasion in pancreatic
cancer via mTOR/S6k1 pathway. Cancer Manag Res. 12:2087–2095. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Depianto D, Kerns ML, Dlugosz AA and
Coulombe PA: Keratin 17 promotes epithelial proliferation and tumor
growth by polarizing the immune response in skin. Nat Genet.
42:910–914. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yang L, Zhang S and Wang G: Keratin 17 in
disease pathogenesis: From cancer to dermatoses. J Pathol.
247:158–165. 2019.PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2-(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Pan C, Liu Q and Wu X:
HIF1α/miR-520a-3p/AKT1/mTOR feedback promotes The proliferation and
glycolysis of gastric cancer cells. Cancer Manag Res.
11:10145–10156. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kobliakov VA: The mechanisms of regulation
of aerobic glycolysis (warburg effect) by oncoproteins in
carcinogenesis. Biochemistry (Mosc). 84:1117–1128. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Abbaszadeh Z, Cesmeli S and Biray Avci C:
Crucial players in glycolysis: Cancer progress. Gene.
726:1441582020. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liao Z, She C, Ma L, Sun Z, Li P, Zhang X,
Wang P and Li W: KDELR2 Promotes glioblastoma tumorigenesis
targeted by HIF1a via mTOR signaling pathway. Cell Mol Neurobiol.
39:1207–1215. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen P, Shen Z, Fang X, Wang G, Wang X,
Wang J and Xi S: Silencing of keratin 17 by lentivirus-mediated
short hairpin RNA inhibits the proliferation of PANC-1 human
pancreatic cancer cells. Oncol Lett. 19:3531–3541. 2020.PubMed/NCBI
|
|
25
|
Hu H, Xu DH, Huang XX, Zhu CC, Xu J, Zhang
ZZ and Zhao G: Keratin 17 promotes tumor growth and is associated
with poor prognosis in gastric Cancer. J Cancer. 9:346–357. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu J, Liu L, Cao L and Wen Q: Keratin 17
promotes lung adenocarcinoma progression by enhancing cell
proliferation and invasion. Med Sci Monit. 24:4782–4790. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li J, Chen Q, Deng Z, Chen X, Liu H, Tao
Y, Wang X, Lin S and Liu N: KRT17 confers paclitaxel-induced
resistance and migration to cervical cancer cells. Life Sci.
224:255–262. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liu L, Chai L, Ran J, Yang Y and Zhang L:
BAI1 acts as a tumor suppressor in lung cancer A549 cells by
inducing metabolic reprogramming via the SCD1/HMGCR module.
Carcinogenesis. Apr 7–2020.([Epub ahead of print). View Article : Google Scholar
|
|
29
|
Ma H, Su R, Feng H, Guo Y and Su G: Long
noncoding RNA UCA1 promotes osteosarcoma metastasis through
CREB1-mediated epithelial-mesenchymal transition and activating
PI3K/AKT/mTOR pathway. J Bone Oncol. 16:1002282019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mei L, Sang W, Cui K, Zhang Y, Chen F and
Li X: Norcantharidin inhibits proliferation and promotes apoptosis
via c-Met/Akt/mTOR pathway in human osteosarcoma cells. Cancer Sci.
110:582–595. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Xu F, Na L, Li Y and Chen L: Roles of the
PI3K/AKT/mTOR signalling pathways in neurodegenerative diseases and
tumours. Cell Biosci. 10:542020. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sankar S, Tanner JM, Bell R, Chaturvedi A,
Randall RL, Beckerle MC and Lessnick SL: A novel role for keratin
17 in coordinating oncogenic transformation and cellular adhesion
in Ewing sarcoma. Mol Cell Biol. 33:4448–4460. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Spirina LV, Kondakova IV, Yurmazov ZA,
Usynin EA, Slonimskaya EM, Lushnikova NA and Podnebesnova DV: VHL
expression in kidney cancer: Relation to metastasis development,
transcription and growth factors and component of Akt/m-TOR
signaling pathway. Bull Exp Biol Med. 167:671–675. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lyu X, Wang J, Guo X, Wu G, Jiao Y, Faleti
OD, Liu P, Liu T, Long Y, Chong T, et al: EBV-miR-BART1-5P
activates AMPK/mTOR/HIF1 pathway via a PTEN independent manner to
promote glycolysis and angiogenesis in nasopharyngeal carcinoma.
PLoS Pathog. 14:e10074842018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Cheng SC, Quintin J, Cramer RA, Shepardson
KM, Saeed S, Kumar V, Giamarellos-Bourboulis EJ, Martens JH, Rao
NA, Aghajanirefah A, et al: mTOR- and HIF-1alpha-mediated aerobic
glycolysis as metabolic basis for trained immunity. Science.
345:12506842014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang Y, Huang Y, Liu H, Su D, Luo F and
Zhou F: Long noncoding RNA CDKN2B-AS1 interacts with miR-411-3p to
regulate ovarian cancer in vitro and in vivo through
HIF-1a/VEGF/P38 pathway. Biochem Biophys Res Commun. 514:44–50.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Shi L, He C, Li Z, Wang Z and Zhang Q:
FBP1 modulates cell metabolism of breast cancer cells by inhibiting
the expression of HIF-1α. Neoplasma. 64:535–542. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lou S, Wang Y, Yu Z, Guan K and Kan Q:
Curcumin induces apoptosis and inhibits proliferation in infantile
hemangioma endothelial cells via downregulation of MCL-1 and
HIF-1α. Medicine (Baltimore). 97:e95622018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wang H, Li ZY, Xu ZH, Chen YL, Lu ZY, Shen
DY, Lu JY, Zheng QM, Wang LY, Xu LW, et al: The prognostic value of
miRNA-18a-5p in clear cell renal cell carcinoma and its function
via the miRNA-18a-5p/HIF1A/PVT1 pathway. J Cancer. 11:2737–2748.
2020. View Article : Google Scholar : PubMed/NCBI
|

















