Introduction
Ceramide, sphingosine and shingosine-1-phosphate
(S1P) are three major metabolites of the sphingolipid signaling
pathway. In response to various stresses, ceramide has been shown
to mediate differentiation, growth arrest and apoptosis (1–5);
sphingosine acts similarly to ceramide (6), and S1P has the opposite effects
(7). Therefore, the balance between
the intracellular levels of ceramide/sphingosine and S1P may
determine the fate of a cell (8).
It is well known that ceramide can be metabolized by ceramidases to
generate sphingosine, which in turn can be phosphorylated by
sphingosine kinases to form S1P. In addition to sphingosine
kinases, ceramidases may play a pivotal role in the regulation of
the intracellular ratio between ceramide, sphingosine and S1P
(6). Based on optimal pHs and
structures, ceramidase are classified into acid, neutral and
alkaline ceramidase. Thus far, three alkaline ceramidases, such as
alkaline ceramidase 1, 2 and 3, have been identified in humans
(9). Human alkaline ceramidase 2
(haCER2), cloned by us (7),
regulates sphingosine and S1P by controlling ceramides hydrolysis.
Different expression levels of haCER2 have opposite effects; high
expression of haCER2 caused sphingosine-induced growth arrest and
low expression of haCER2 promoted S1P-mediated cell proliferation
(7). Thus, regulation of haCER2 may
affect the fate of a cell by controlling the balance of
ceramide/sphingosine and S1P. In our previous study, serum
deprivation upregulated haCER2 mRNA and activity, but the
mechanism remains unknown. In the present study, the HepG2 human
hepatoma cell line was used as a cell model and it was found that
p38/activator protein-1 (AP-1) signaling is involved in serum
deprivation-induced haCER2 transcriptional activation and
subsequently caused an increase in haCER2 enzymatic activity.
Materials and methods
Cells and materials
HepG2 cells were cultured in Dulbecco's modified
Eagle's medium (HyClone Corp., Logan, UT, USA) supplemented with
10% heat-inactivated fetal bovine serum (FBS; HyClone Corp.), 100
U/ml penicillin and 100 µg/ml streptomycin in a humidified
atmosphere at 37˚C and 5% CO2.
o-Phthalaldehyde was from Fluka (Milwaukee,
WI, USA). D-e-C24:1-ceramide and D-e-C17-sphingosine were purchased
from Avanti Polar Lipids, Inc., (Pelham, AL, USA). Actinomycin D
and 12-O-tetradecanoylphorbol-13-acetate (TPA; an activator
of AP-1) were purchased from Sigma-Aldrich (St. Louis, MO, USA).
PD-98059 [inhibitor of the extracellular signal-regulated kinase
(ERK) pathway], SP-600125 [inhibitor of the c-Jun
NH2-terminal kinase (JNK) pathway], SB-203580 [inhibitor
of p38 mitogen-activated protein kinase (MAPK) pathway] and
Bay117085 [inhibitor of nuclear factor-κ B (NF-κ B) pathway] were
from Calbiochem/EMD Biosciences, Inc., (San Diego, CA, USA). AP-1
and NF-κ B Cignal Reporter were purchased from Qiagen (Valencia,
CA, USA). Renilla luciferase reporter plasmid pRL-TK and the
Dual-Luciferase Reporter Assay system were from Promega Corp.,
(Madison, WI, USA). SR11302 (AP-1 specific inhibitor) were
purchased from Tocris Bioscience (Ellisville, MO, USA). Human p38
small interfering RNA (siRNA) (sc-29433), p38 primers for
quantitative polymerase chain reaction (qPCR) (sc-29433-PR) and p38
antibody (sc-7972) were purchased from Santa Cruz Biotechnology,
Inc., (Santa Cruz, CA, USA). The RNeasy Mini kit was from Qiagen
(Santa Clarita, CA, USA) and the iScript cDNA Synthesis kit was
purchased from Bio-Rad Laboratories, Inc., (Hercules, CA, USA).
Other unlisted chemicals were purchased from Sigma-Aldrich Co.,
LLC.
haCER2 activity assays
haCER2 activity was determined by the release of
sphingosine from ceramide as described previously (10). In brief, D-e-C24:1-ceramide was
selected as a substrate dispersed into A buffer (pH 9.0), including
25 mmol/1 glycine-NaOH, 5 mmol/1 CaCl2 and 0.3% Triton
X-100, by water bath sonication. The microsomes containing haCER2
from HepG2 cells were suspended in B buffer (pH 9.0), including 25
mmol/1 glycine-NaOH and 5 mmol/1 CaCl2. Incubation of
the microsomes with ceramide substrate at 37˚C for 20 min initiated
the enzymatic reactions, which were stopped by boiling the mixture.
After Bligh-Dye extraction, sphingosine was assayed by
high-performance liquid chromatography analysis with
D-e-C17-sphingosine as an internal standard. HPLC was conducted
using the Agilent 1050-HPLC model fitted with an eclipse XDB-C18
column (Agilent Technologies, Palo Alto, CA, USA). The solvent was
methanol-potassium phosphate buffer (90:10 v/v) and the flow rate
was 0.7 ml/min. A HP1046 fluorescence detector with an excitation
at 345 nm and emission at 455 nm was used.
Reverse transcription-qPCR
(RT-qPCR)
Total RNA was isolated from HepG2 cells using the
RNeasy Mini kit according to the manufacturer's instructions. RNA
was reverse-transcribed into first-strand cDNA with the iScript
cDNA Synthesis kit using 20 µ1 of the reaction mixture containing 1
µ1 iScript reverse transcriptase, 4 µ1 5X iScript reaction mixture
and 0.5 µg total RNA. The complete reaction was cycled for 5 min at
25˚C, 30 min at 42˚C and 5 min at 85˚C using a PTC-200 DNA Engine
(MJ Research Inc., Waltham, MA, USA) (11). cDNA was subjected to RT-qPCR
analysis, which was performed on an iCycler system (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). The initial PCR step was 3
min at 95˚C, followed by 40 cycles of a 10-sec melting at 95˚C and
a 45-sec annealing/extension at 60˚C. The final step was 1-min
incubation at 60˚C. All the reactions were performed in triplicate.
Data are expressed as the mean normalized expression, which is
directly proportional to the amount of haCER2 mRNA relative
to the amount of β-actin mRNA. The primers used were
forward, 5′-AGTGTCCTGTCTGCGGTTACG-3′; and reverse,
5′-TGTTGTTGATGGCAGGCTTGAC-3′ for haCER2; and forward,
5′CAATGTTCGGTGCAATTCAGAG-3′ and reverse,
5′-GGATCCCATTCCTACCACTGTG-3′ for β-actin (9).
mRNA stability analysis
HepG2 cells were plated in 12-well plates at a
density of 0.4×106 cells/well overnight and treated with
serum deprivation, followed by the addition of 10 µg/ml actinomycin
D. HepG2 cells were harvested 2 h after the addition of actinomycin
D and haCER2 mRNA was quantified using RT-qPCR as above.
Transfection and luciferase activity
assay
HepG2 cells were transiently transfected with 1 µg
AP-1 or NF-κ B Cignal Reporter for 24 h using FuGENE HD as the
transfection reagent. The cells were cotransfected with the
Renilla luciferase reporter plasmid pRL-TK (50
ng/106 cells) as an internal control. The cells were
subsequently treated with serum deprivation for 8 h. Following the
treatment, the cells were rinsed with cold PBS and lysed with the
buffer from the Dual-Luciferase Reporter Assay system. Firefly and
renilla luciferase levels were measured in a luminometer
using the dual-luciferase reporter assay reagents according to the
manufacturer's instructions. The firefly luciferase levels were
normalized to the renilla luciferase levels.
Treatment of cells with the inhibitors
of the signaling pathways
HepG2 cells were treated with serum deprivation in
the absence or presence of 10 µM PD-98059, SP-600125, SB-203580 for
8 h. Following the treatment, haCER2 mRNA was quantified
using qPCR.
Transfection of siRNA
In order to silence the p38 MAPK gene
expression, the siRNA approach was used. The HepG2 cells were
cultured for 24 h before the transfection was processed in 24-well
plates with 40 nM siRNAs using Lipofectamine® 2000
(Invitrogen Corp., Carlsbad, CA, USA). After a 24-h incubation with
p38 siRNA or Con-siRNA, the HepG2 cells were cultured with
or without serum deprivation for an additional 8 h and subsequently
the cells were collected and haCER2 mRNA was quantified
using RT-qPCR. p38 MAPK knockdown was confirmed by western
blot analysis (12) and qPCR.
Statistical analysis
Data are expressed as the mean ± standard deviation,
with a minimum of three independent experiments analyzed by
Student's t-test or analysis of variance. P<0.05 was considered
to indicate a statistically significant difference.
Results
haCER2 is upregulated by serum
deprivation in HepG2 cells
Our previous study revealed that haCER2 is
upregulated by serum deprivation in HeLa cells; this prompted the
investigation of whether the expression of the endogenous
haCER2 was upregulated upon serum deprivation in HepG2
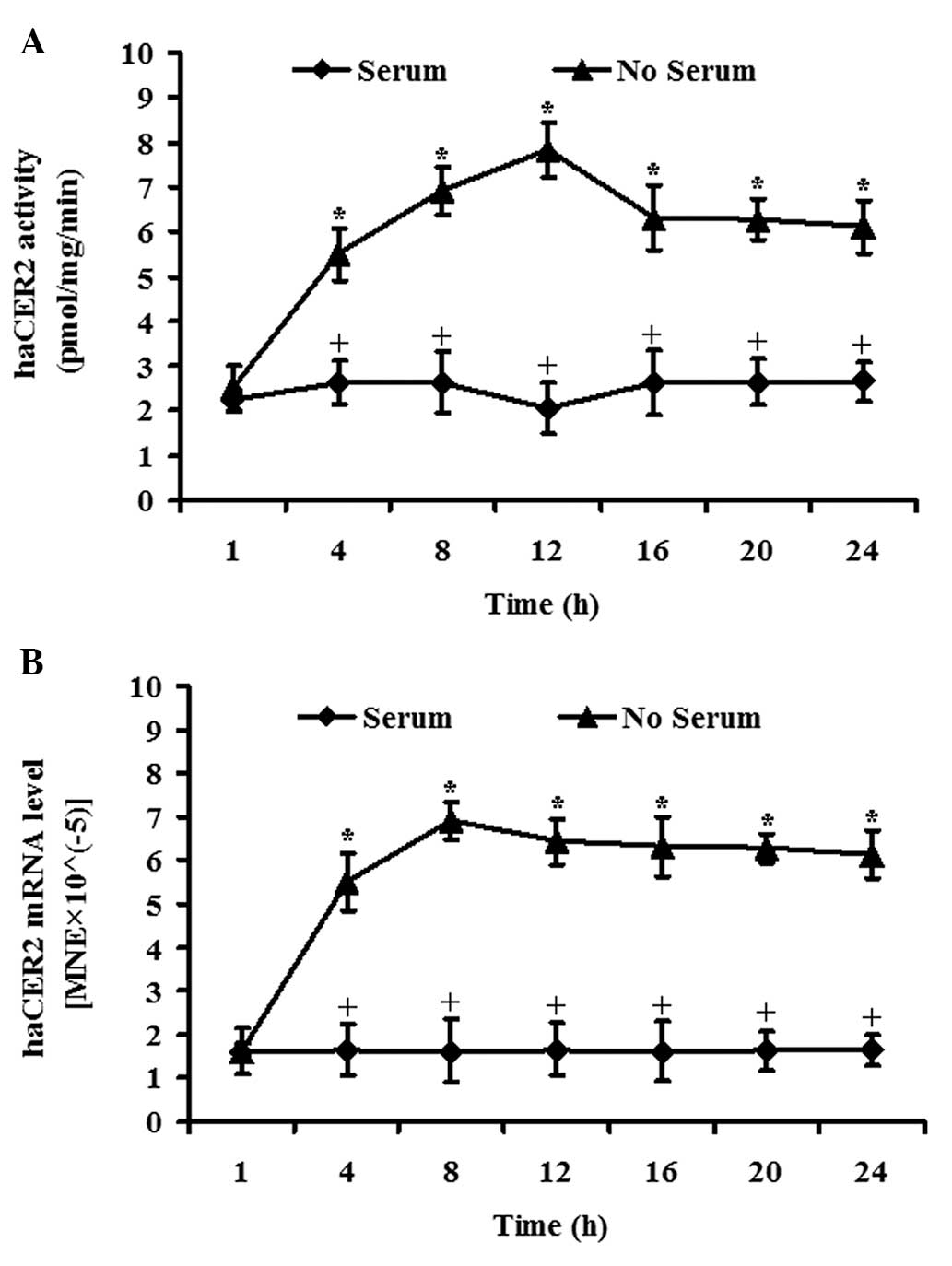
cells. In vitro activity assays showed that serum
deprivation markedly increased alkaline ceramidase activity on
D-e-C24:1-ceramide in a time-dependent manner, reaching a peak at
12 h (Fig. 1A). RT-qPCR analysis
demonstrated that haCER2 mRNA was significantly upregulated
in response to serum deprivation in a time-dependent manner and the
increase plateaued at 8 h (Fig.
1B). These results indicate that serum deprivation upregulates
haCER2 activity and mRNA. Comparing the kinetics of haCER2 activity
and mRNA, it is indicated that the serum deprivation induced-haCER2
activity increase in HepG2 happens at mRNA level. Based on this
kinetics study, haCER2 mRNA was measured at 8 h and haCER2
activity at 12 h in the subsequent studies.
Transcription, but not mRNA stability,
is involved in haCER2 upregulation in HepG2 cells by serum
deprivation
The data above showed that haCER2 mRNA was
upregulated by serum deprivation and it is well known that gene
expression can be regulated at the transcriptional level or the
post-transcriptional level. In the last years, it became evident
that mRNA stability/turnover provides an important mechanism for
post-transcriptional control of gene expression. Therefore, whether
mRNA stability was involved in the regulation of haCER2
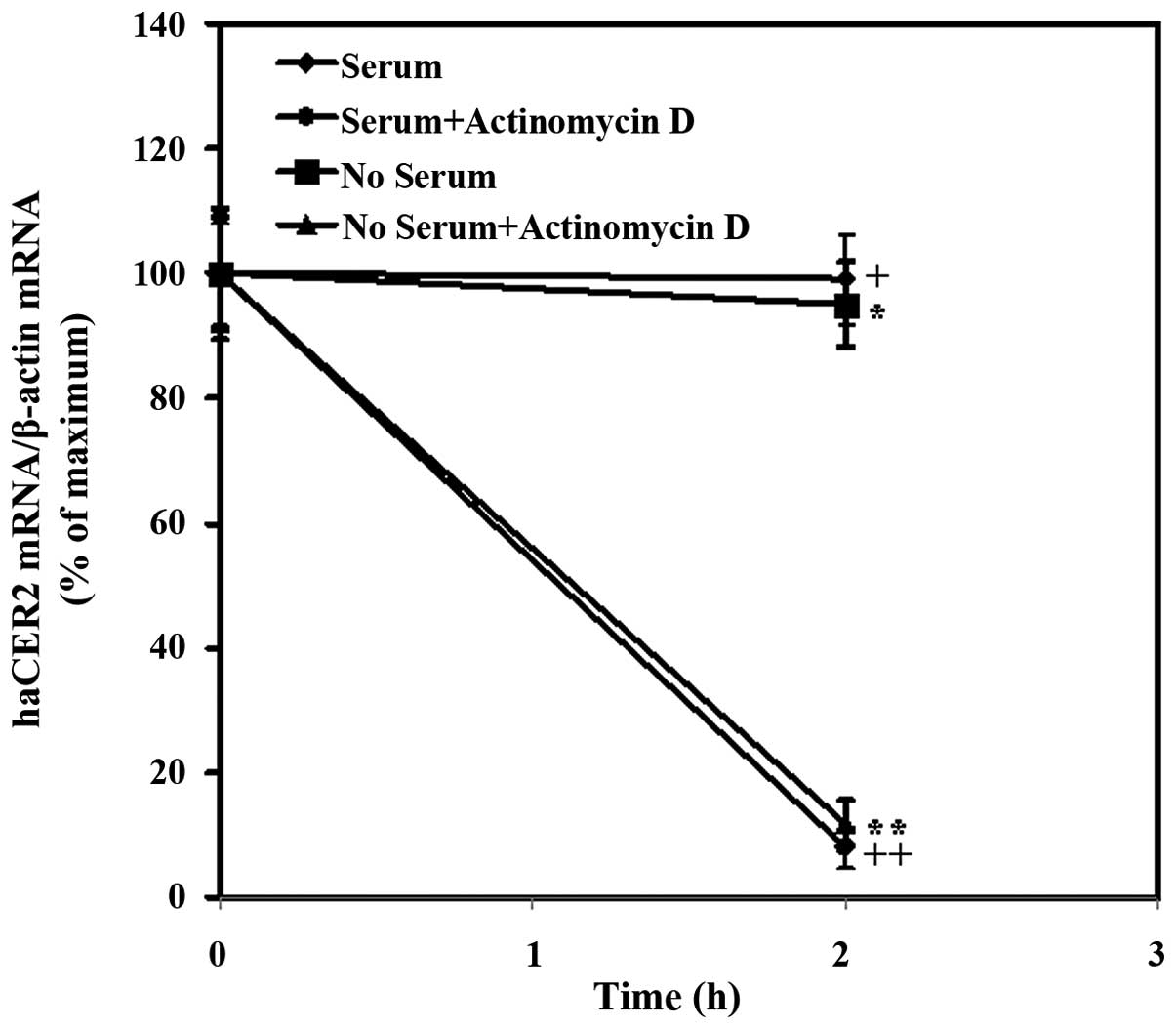
expression by serum deprivation was determined. In the present
study, the inhibitory effect of actinomycin D, an inhibitor of
transcription, on haCER2 transcription was confirmed by the
finding that the treatment of cells with actinomycin D for 2 h
reduced the haCER2 mRNA level by 92% in medium containing
10% FBS (Fig. 2). Furthermore, a
result showed that in the presence of actinomycin D for 2 h,
haCER2 mRNA level was observed to reduce by 88% in HepG2
cells treated with serum starvation (Fig. 2). Comparing the mRNA level in HepG2
cells exposure to actinomycin D for 2 h between serum starvation
and 10% FBS treatment, it was indicated that serum
deprivation-stimulated haCER2 upregulation was due to the
transcriptional regulation and not mRNA stability.
AP-1 signaling is involved in haCER2
upregulation in HepG2 cells by serum deprivation
A previous study revealed that transcriptional
factor AP-1 signaling regulated human neutral ceramidase gene
transcription and the degree of decrease in ceramide mass was
significantly greater compared to the reduction of human neutral
ceramidase transcription, raising the possibility that AP-1 may
regulate other ceramide metabolizing enzymes, including haCER2
(13). Therefore, in the present
study, whether AP-1 regulated serum deprivation-induced haCER2
upregulation or not was investigated. In the first experiment,
HepG2 cells were transfected with the luciferase reporter vectors
constructed with either AP-1 or NF-κ B-binding element in the
promoter. The results showed that serum deprivation stimulated AP-1
but not NF-κ B activity at 12 h (Fig.
3A). Consistent with the above results, haCER2 mRNA
could be upregulated by serum deprivation in HepG2 cells
transfected with AP-1 or NF-κ B (Fig.
3B). In addition, the AP-1 specific inhibitor SR11302 (14, 15)
completely blocked the serum deprivation-induced haCER2 mRNA
upregulation (Fig. 3C), and TPA, as
an activator of AP-1 (16), has an
additive effect on serum deprivation-induced haCER2 mRNA
upregulation (Fig. 3D). Notably, in
10% FBS medium, TPA also stimulates haCER2 mRNA expression
(Fig. 3D).
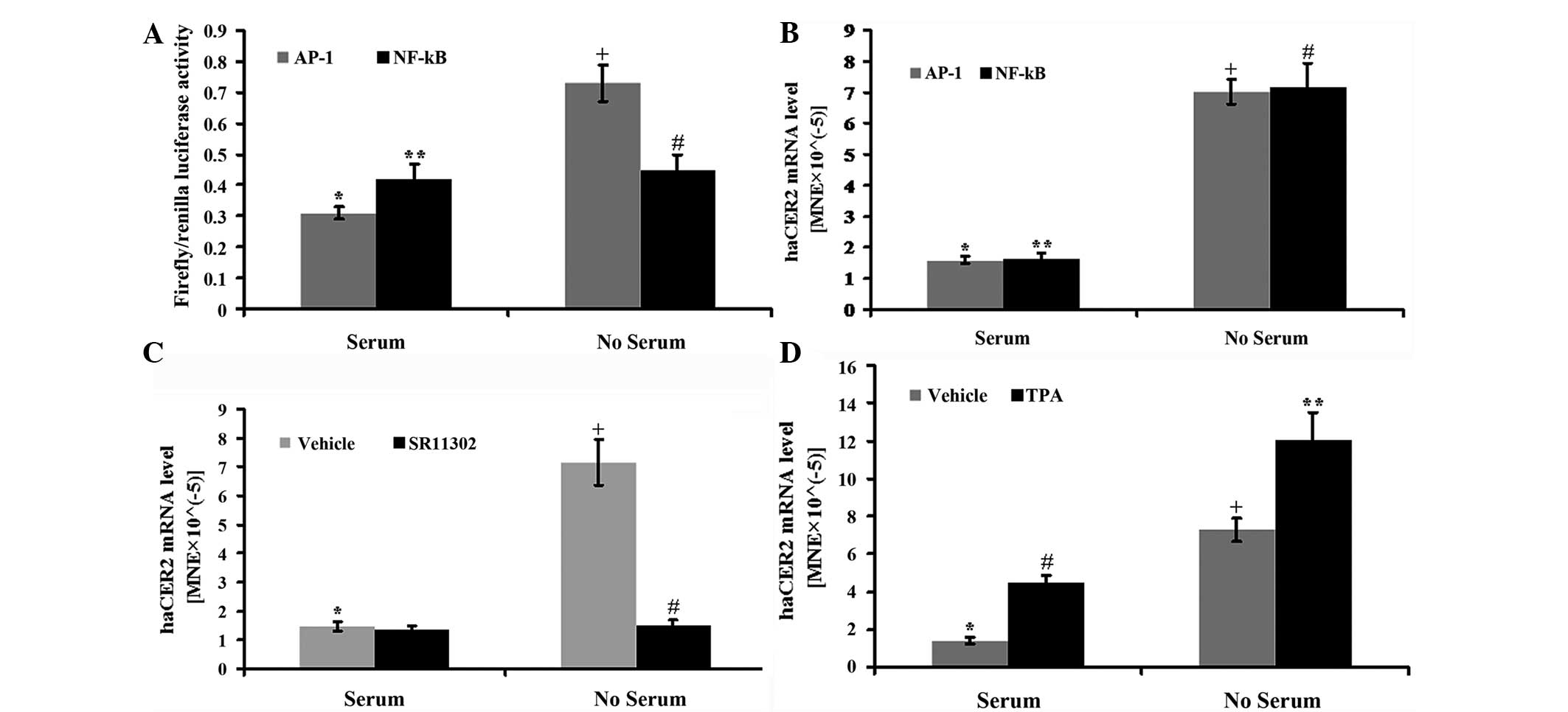 | Figure 3.Activator protein-1 (AP-1) signaling
is involved in human alkaline ceramidase 2 (haCER2)
upregulation in HepG2 cells by serum deprivation. HepG2 cells were
transfected with the DNA vectors constructed with AP-1 or the
nuclear factor-κ B (NF-κ B)-binding element in the presence of
FuGENE HD as a transfection reagent for 24 h. After the
transfection, the cells were treated with serum deprivation for 8 h
(for haCER2 mRNA) or 12 h (for luciferase activity) and
collected for further analysis. (A) The AP-1 or NF-κ B activity was
presented as the ratio of firefly luciferase vs. renilla
luciferase activity
(*,+P<0.05;**,#P>0.05) and (B)
haCER2 mRNA was quantified using reverse
transcription-quantitative PCR (RT-qPCR),
(*,+P<0.05;**,#P<0.05). HepG2 cells
were treated with (C) 1 µM SR11302
(*,+P<0.05;+,#P<0.05) or (D) 10 ng/ml
12-O-tetradecanoylphorbol-13-acetate (TPA)
(*,+P<0.05;#,**P<0.05;*,#P<0.05;+,**P<0.05)
in serum deprived medium or 10% fetal bovine serum medium for 8 h
and haCER2 mRNA was measured by RT-qPCR. MNE, mean
normalized expression. |
p38 MAPK is involved in haCER2
upregulation in HepG2 cells by serum deprivation
AP-1 transcriptional activity can be regulated by
MAPK, including ERK, JNK and p38 MAPK signal transduction pathways
(17). Thus, which one was involved
in haCER2 upregulation by serum deprivation in HepG2 cells
was determined. In the study, HepG2 cells were treated with serum
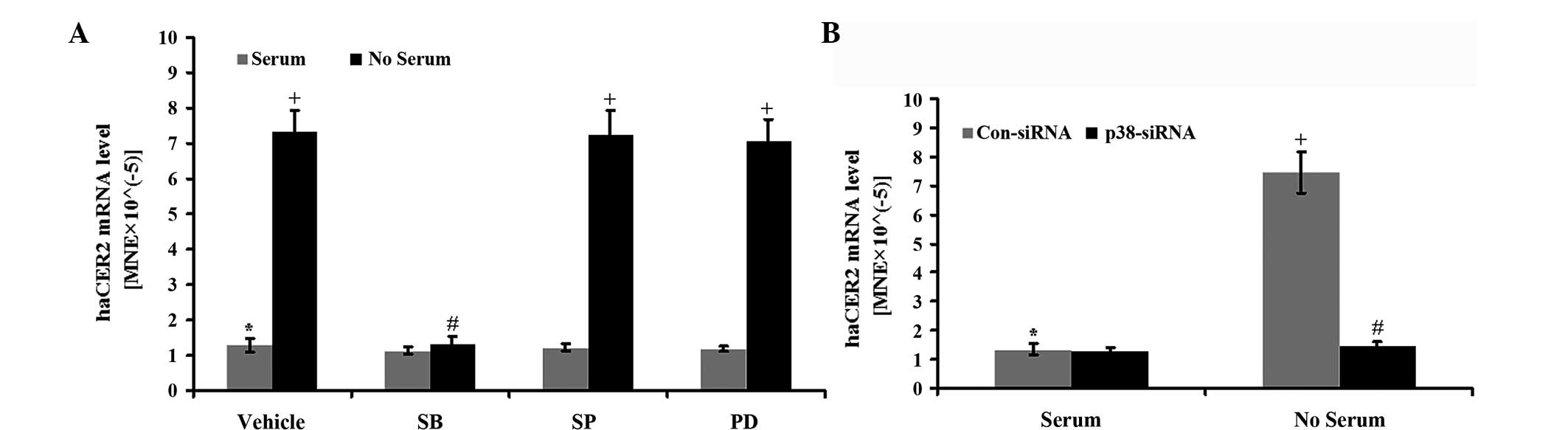
deprivation in the presence of the pharmacological inhibitors of
the MAPK pathways. The results showed that haCER2
upregulation by serum deprivation were inhibited by SB-203580, an
inhibitor for the p38 MAPK pathway, but not by SP-600125 (an
inhibitor for JNK) and PD-98059 (an inhibitor for ERK) (Fig. 4A). Consistent with this result,
downregulation of p38 by specific siRNA completely inhibited
haCER2 upregulation by serum deprivation (Fig. 4B). p38 MAPK knockdown was confirmed
by western blot analysis and qPCR (data not shown). These data
indicate that the p38 MAPK pathway is involved in haCER2
upregulation by serum deprivation.
Discussion
haCER2 plays an important role in cellular responses
by regulating the hydrolysis of ceramides in cells (18). High and low ectopic expression of
haCER2 leads to different cell fates through changing the
ratio of ceramide/sphingosine and S1P (7). Therefore, the regulation of
haCER2 expression by physiological or pathological stimuli
is extremely important, but little is known regarding it. In the
present study, firstly, serum deprivation was found to cause haCER2
activation in HepG2 cells in a time-dependent manner, which is in
agreement with our previous finding conducted in HeLa cells
(7). Comparing the kinetics of
haCER2 activity and mRNA expression, it is indicated that haCER2
activity elevation was caused by increasing haCER2 mRNA. The
level of an mRNA within a cell depends on its rate of synthesis and
rate of decay (19). mRNA stability
assay showed that actinomycin D, an inhibitor of transcription,
completely inhibited the haCER2 mRNA increase caused by
serum deprivation. Taking the above results together, it was
indicated that serum deprivation-stimulated haCER2 mRNA
elevation was due to the transcriptional regulation.
In response to a plethora of physiological and
pathological stimuli, numerous gene regulation is mediated by AP-1
(20). A previous study suggested
that human neutral ceramidase gene transcription is regulated by
AP-1 signaling, it also raised the possibility that other
ceramidases may be regulated by AP-1 as the degree of decrease in
ceramide mass was significantly greater than the reduction of human
neutral ceramidase transcription (13). Therefore, whether haCER2 gene
transcription by serum deprivation is regulated by AP-1 signaling
pathway was investigated. The results showed that serum deprivation
stimulated AP-1 luciferase activity, whereas NF-κ B activity was
not stimulated by serum deprivation. Notably, SR11302 (a specific
inhibitor of AP-1) blocked serum deprivation-induced haCER2
mRNA increase, whereas TPA (an activator of AP-1) additively
increased haCER2 mRNA with serum deprivation. All these data
confirmed that transcription factor AP-1 is involved in
haCER2 gene transcription by serum deprivation.
Three different types of MAPKs, including ERK, JNK
and p38, contribute to the induction of AP-1 activity in response
to a diverse array of extracellular stimuli (21). In the present study, the
pharmacological inhibitors experiment revealed that SB-203580
blocked haCER2 transcription induced by serum deprivation,
whereas SP-600125 and PD-98059 could not. These indicated that the
p38 MAPK, and not JNK and ERK, pathway is involved in serum
deprivation-induced haCER2 transcription. The involvement of
the p38 MAPK pathway in haCER2 transcription mediated by
serum starvation was further confirmed by p38-specific siRNA
downregulation.
Notably, AP-1 is composed of the Jun protein family
(c-Jun, Jun B and Jun-D) and the Fos protein family (such as c-Fos,
FosB, Fra-1 and Fra-2) (20), and
the present study does not identify which subunit(s) of AP-1 play a
pivotal role in haCER2 transcription mediated by serum
starvation and this is an area for future investigation.
A low-nutrient environment is commonly found in the
central region of solid tumor, such as liver cancer (22, 23).
Serum starvation mimics the tumor growth environment in vivo
(24). In HepG2 cells, serum
deprivation upregulated haCER2 expression in which the p38
MAPK/AP-1 signaling pathway is involved, and this mechanism may
explain why haCER2 is upregulated in solid liver cancer (7, 25).
The regulation of mRNA stability is an important
factor in modulating gene expression, and the p38 MAPK pathway has
been indicated in the regulation of the mRNA half-lives of a number
of genes (26). Based on the
aforementioned studies, we hypothesized that mRNA stability may be
involved in serum starvation-induced haCER2 mRNA increase,
but mRNA decay experiments exclude this possibility and future
research is required to explore this reason.
In conclusion, the present study conducted in the
human HepG2 hepatoma cell line indicates that serum deprivation
affects the p38 MAPK signaling pathway leading transcription factor
AP-1 activation and subsequently regulates haCER2 mRNA
expression. This mechanism may interpret why haCER2 is
upregulated in liver cancer.
Acknowledgements
The present study was supported in part by the
National Natural Science Foundation of China (grant no. 81360309),
‘Sphingolipids and Related Diseases’ Program for Innovative
Research Team of Guilin Medical University, and Hundred Talents
Program of Universities and Colleges Directly under the Guangxi
Zhuang Autonomous Region.
References
|
1
|
Spiegel S and Merrill AH Jr: Sphingolipid
metabolism and cell growth regulation. FASEB J. 10:1388–1397.
1996.PubMed/NCBI
|
|
2
|
Merrill AH Jr, Schmelz EM, Wang E, et al:
Importance of sphingolipids and inhibitors of sphingolipid
metabolism as components of animal diets. J Nutr. 127 (Suppl
5):S830–S833. 1997.
|
|
3
|
Hannun YA and Obeid LM: The
Ceramide-centric universe of lipid-mediated cell regulation: stress
encounters of the lipid kind. J Biol Chem. 277:25847–25850. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Levade T, Malagarie-Cazenave S, Gouaze V,
et al: Ceramide in apoptosis: a revisited role. Neurochem Res.
27:601–607. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kolesnick R and Fuks Z: Radiation and
ceramide-induced apoptosis. Oncogene. 22:5897–5906. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cuvillier O: Sphingosine in apoptosis
signaling. Biochim Biophys Acta. 1585:153–162. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Xu R, Jin J, Hu W, et al: Golgi alkaline
ceramidase regulates cell proliferation and survival by controlling
levels of sphingosine and S1P. FASEB J. 20:1813–1825. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cuvillier O, Pirianov G, Kleuser B, et al:
Suppression of ceramide-mediated programmed cell death by
sphingosine-1-phosphate. Nature. 381:800–803. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Xu R, Sun W, Jin J, Obeid LM and Mao C:
Role A of alkaline ceramidases in the generation of sphingosine and
its phosphate in erythrocytes. FASEB J. 24:2507–2515. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sun W, Hu W, Xu R, et al: Alkaline
ceramidase 2 regulates beta1 integrin maturation and cell adhesion.
FASEB J. 23:656–666. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jin J, Zhang X, Lu Z, et al: Acid
sphingomyelinase plays a key role in palmitic acid-amplified
inflammatory signaling triggered by lipopolysaccharide at low
concentrations in macrophages. Am J Physiol Endocrinol Metab.
305:E853–E867. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhu Q, Lin L, Cheng Q, et al: The role of
acid sphingomyelinase and caspase 5 in hypoxia-induced HuR cleavage
and subsequent apoptosis in hepatocytes. Biochim Biophys Acta.
1821:1453–1461. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
O'Neill SM, Houck KL, Yun JK, Fox TE and
Kester M: AP-1 binding transcriptionally regulates human neutral
ceramidase. Arch Biochem Biophys. 511:31–39. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Huang C, Ma WY, Dawson MI, Rincon M,
Flavell RA and Dong Z: Blocking activator protein-1 activity, but
not activating retinoic acid response element, is required for the
antitumor promotion effect of retinoic acid. Proc Natl Acad Sci
USA. 94:5826–5830. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li JJ, Westergaard C, Ghosh P and Colburn
NH: Inhibitors of both nuclear factor-kappa B and activator
protein-1 activation block the neoplastic transformation response.
Cancer Res. 57:3569–3576. 1997.PubMed/NCBI
|
|
16
|
Fisch TM, Prywes R and Roeder RG: An
AP1-binding site in the c-fos gene can mediate induction by
epidermal growth factor and 12-O-tetradecanoyl phorbol-13-acetate.
Mol Cell Biol. 9:1327–1331. 1989.PubMed/NCBI
|
|
17
|
Whitmarsh AJ and Davis RJ: Transcription
factor AP-1 regulation by mitogen-activated protein kinase signal
transduction pathways. J Mol Med (Berl). 74:589–607. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sun W, Jin J, Xu R, et al: Substrate
specificity, membrane topology, and activity regulation of human
alkaline ceramidase 2 (ACER2). J Biol Chem. 285:8995–9007. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bellofatto V and Wilusz J: Transcription
and mRNA stability: parental guidance suggested. Cell.
147:1438–1439. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hess J, Angel P and Schorpp-Kistner M:
AP-1 subunits: quarrel and harmony among siblings. J Cell Sci.
117:5965–5973. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Karin M: The regulation of AP-1 activity
by mitogen-activated protein kinases. J Biol Chem. 270:16483–16486.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Harrington EA, Fanidi A and Evan GI:
Oncogenes and cell death. Curr Opin Genet Dev. 4:120–129. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dang CV and Semenza GL: Oncogenic
alterations of metabolism. Trends Biochem Sci. 24:68–72. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xiong Y, Fang JH, Yun JP, et al: Effects
of micro RNA-29 on apoptosis, tumorigenicity, and prognosis of
hepatocellular carcinoma. Hepatology. 51:836–845. 2010.PubMed/NCBI
|
|
25
|
Graveel CR, Jatkoe T, Madore SJ, Holt AL
and Farnham PJ: Expression profiling and identification of novel
genes in hepatocellular carcinomas. Oncogene. 20:2704–2712. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Frevel MA, Bakheet T, Silva AM, Hissong
JG, Khabar KS and Williams BR: p 38 Mitogen-activated protein
kinase-dependent and -independent signaling of mRNA stability of
AU-rich element-containing transcripts. Mol Cell Biol. 23:425–436.
2003. View Article : Google Scholar : PubMed/NCBI
|