Introduction
Pleckstrin homology-like domain, family A, member 1
(PHLDA1; also known as PHRIP, TDAG51, DT1P1B11 and
MGC131738) was first identified as a potential transcription factor
required for Fas expression and activation-induced apoptosis in
mouse T cell hybridomas (1). It was
later mapped to human chromosome 12q15 by radiation hybrid mapping
(2). The PHLDA family is
evolutionarily conserved and comprises three members that are
located on different chromosomes: PHLDA1 (12q21.2),
PHLDA2 (11p15.4) and PHLDA3 (1q32.1). All three share
similar gene organization [i.e., two exons (one coding exon) and a
small intron] and encode proteins with a PHL domain (3). During the last decade, PHLDA1 has
received increasing attention, although its functional role remains
to be elucidated.
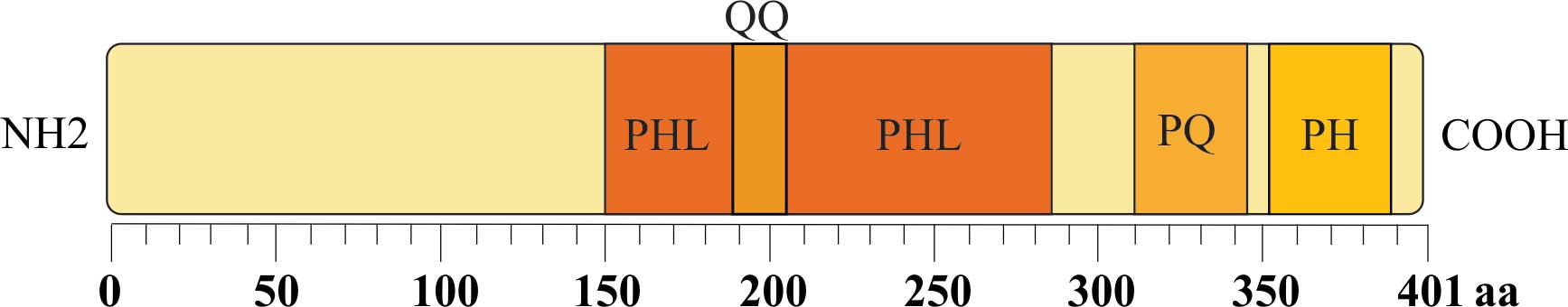
PHLDA1 encodes a protein that is 401 amino
acids (aa) in length. This protein is rich in polyglutamine (QQ),
proline-glutamine (PQ) and proline-histidine tracts with a PHL
domain (1–3) (Fig. 1).
Sequencing comparisons have shown that mouse, human and rat PHLDA1
cDNA and protein exhibit high levels of sequence identity (2,4).
PHL domains are evolutionarily conserved and are
found in proteins from prokaryotes and eukaryotes (5,6). These
domains are composed of 100–120 aa residues, and their properties
include binding to phosphatidylinositol lipids (5). Numerous proteins containing PHL domains
(such as insulin receptor substrate proteins) have been found to
interact with membrane components, eliciting a variety of cellular
responses and participating in cell signaling transduction,
vesicular trafficking and cytoskeletal rearrangement (7,8). QQ and PQ
stretches are well-conserved motifs in eukaryotic proteins, and
they are present predominantly in transcription factors that are
involved in transcriptional activation and protein-protein
interactions (9,10). In addition, a number of variations in
tandem repeats of the trinucleotide sequence CAG, which encodes
homopolymeric stretches of QQ, have been associated with several
inherited neurodegenerative diseases (10,11). Due to
the presence of QQ stretches that function as transcriptional
activation domains, PHLDA1 could act as a transcription factor
required for Fas expression (1).
PHLDA1 expression and cell death
PHLDA1 mRNA and protein are expressed in several
mammalian tissues, suggesting that it may have important cellular
functions (1,4,12). In
addition to being widely expressed in different tissues, PHLDA1
subcellular localization varies. PHLDA1 is found predominantly in
the cytoplasm (13–16); however, perinuclear, nucleolar and
nuclear localization have also been reported (12,13,16,17). PHLDA1
expression is induced by different stimuli, such as estrogens,
growth factors, differentiation and endoplasmic reticulum
(ER)-stress agents, and it is associated with different biological
processes, such as cell apoptosis, cell proliferation and
differentiation (Fig. 2).
 | Figure 2.Modulators and biological functions
of Pleckstrin homology-like domain, family A, member 1 (PHLDA1).
PHLDA1 expression is modulated by a variety of external stimuli in
different cell types. As a consequence, diverse biological
processes are modulated. ER-stress agents, endoplasmic
reticulum-stress agents; ERBB2, receptor tyrosine kinase 2; P,
phosphorylation; FGF, fibroblast growth factor; IGF1, insulin-like
growth factor 1; RAS, rat sarcoma virus oncogene; E2,
17β-estradiol; ER, estrogen receptor; NF-κB, nuclear factor of κ
light polypeptide gene enhancer in B cells; p38 MAPK, p38
mitogen-activated protein kinase; PI3K, phosphatidylinositol
3-kinase; TNF-α, tumor necrosis factor-α; miR-181, microRNA
181. |
Evasion of apoptosis or programmed cell death is a
hallmark of cancer, and the development of resistance to apoptosis
is associated with tumor pathogenesis and affects chemo- and
radioresistance (18). Several lines
of experimental evidence have implicated PHLDA1 in intrinsic and
extrinsic apoptotic pathways; however, the mechanisms involved in
its action as a proapoptotic or antiapoptotic agent remain to be
elucidated.
The role of PHLDA1 in activation-induced cell death
(AICD) was first reported by Park et al (1) in 1996, who used a T cell hybridoma model
to determine that PHLDA1 expression is induced by T cell receptor
activation in the presence of phorbol myristate acetate and
ionomycin (1). The study also reported
that PHLDA1 is required for Fas (CD95) expression, which has an
important role in T cell receptor-induced apoptosis. These data
point to an essential role for PHLDA1 in the induction of apoptosis
in a T cell hybridoma model. However, the study by Rho et al
(19) investigated the assocation
between PHLDA1 and Fas expression in vivo and demonstrated
that PHLDA1−/− mice express normal levels of Fas
and have normal T cell apoptosis, indicating that PHLDA1 is not
essential for Fas expression. Furthermore, Oberg et al
(20) demonstrated that in human T
cells and pancreatic tumor cells, induction of PHLDA1 is inhibited
by mitogen-activated protein kinase (MAPK) and protein kinase C
(PKC) inhibitors, indicating that these two pathways may be
involved in the regulation of PHLDA1 expression. However, no
association was found between PHLDA1 expression and AICD or cell
proliferation. By contrast, increased Fas expression was shown to
be dependent on PKC activation (1,21), which
induces Fas expression only in cells expressing wild-type PHLDA1
(21). Sequence analysis and
experimental studies revealed that PHLDA1 contains putative casein
kinase II and PKC phosphorylation sites (4,21). These
data suggest that PHLDA1 transcriptional activation of the target
gene FAS may be dependent on post-transcriptional modification upon
PKC activation, which raises the possibility of transcriptional
activity of PHLDA1 in a broader context.
In agreement with the proapoptotic role executed by
PHLDA1 in rat neuronal cells, PHLDA1 mRNA and protein were induced
by fibroblast growth factor and phorbol dibutyrate, which is a
differentiation agent that activates PKC (4). In addition, Gomes et al (4) reported that microinjection of
differentiating H19-7 hippocampal cells with the neutralizing
antibody anti-PHLDA1 increases cell survival. Furthermore,
transient PHLDA1 expression decreases cell survival in
differentiating and non-differentiating H19-7 cells, indicating
that PHLDA1 acts as a mediator of differentiation-associated
apoptosis in H19-7 cells (4). PHLDA1
expression is induced by the phorbol ester and ionomycin in
different transformed T cell lines and was also found to interact
with two mRNA-binding proteins, eIF3-p66 and PABP, in 293T cells.
Additionally, PHLDA1 cotransfection with the luciferase gene leads
to protein biosynthesis inhibition, a process that is important for
the execution of apoptosis (13).
ER stress-inducing agents, such as homocysteine,
tunicamicyn and farnesol (FOH), have been reported to induce PHLDA1
expression in different cell types (12,22,23). Hossain et al (12) demonstrated that induction of PHLDA1 by
homocysteine involves ER stress and is dependent on eIF2α
phosphorylation. In addition, transient overexpression of PHLDA1
leads to changes in cell morphology, decreases cell adhesion, and
promotes detachment-mediated cell death in endothelial cells
(12). To improve the understand how
FOH induces apoptosis, Joo et al (22) performed cDNA microarray analysis and
compared the gene expression profiles between vehicle-treated and
FOH-treated H460 human lung adenocarcinoma cells. An induction of a
number of genes associated with ER stress-response signaling,
including PHLDA1, was observed. The study also reported that
FOH-induced apoptosis in human lung carcinoma cells is coupled to
the activation of an ER-stress response that includes activation of
the sensors IRE1 and PERK, dependent on the activation of the
MEK-ERK signaling pathway.
Carlisle et al (23) found that treatment of HK-2 cells (an
immortalized proximal tubule epithelial cell line) with the
ER-stress agents thapsigargin and cyclosporine A increases the
cytosolic Ca2+ concentration and induces PHLDA1
expression, leading to epithelial-mesenchymal transition (EMT). In
addition, overexpression of PHLDA1 in HK-2 cells leads to shape
changes, β-catenin disruption and increased apoptosis, indicating
that PHLDA1 may cause dissolution of adherens junctions, β-catenin
nuclear localization and EMT induction (23). EMT is an important biological process
that involves downregulation of epithelial markers and upregulation
of mesenchymal markers, in which epithelial cells lose cell-cell
adhesion and cell polarity, and gain the migratory and invasive
properties that are characteristic of mesenchymal cells (24,25). As EMT
has a key role in tumor progression and may influence therapy
response, the potential role of PHLDA1 as an EMT regulator requires
further investigation.
In support of the proapoptotic function exhibited by
PHLDA1, Neef et al (14)
reported that constitutive PHLDA1 expression in different
melanoma-derived cell lines leads to reduced cell growth, reduced
colony formation and increased basal apoptosis, which is associated
with increased cleaved caspase 9 and cleaved poly adenosine
diphosphate ribose polymerase. Upregulation of PHLDA1 has been
identified in autophagy and apoptosis induced by rapamycin in T47D
breast cancer cells (26). In
addition, stress-induced heat-shock proteins (Hsp; including Hsp70,
Hsp110 and Hsp40) bind directly to the pleckstrin domain of PHLDA1
and suppress its proapoptotic function in mouse embryonic
fibroblast (MEF) cells (27). The
promoter region of PHLDA1 has heat-shock element binding
sites, where heat-shock transcription factor 1 (HSF1) binds and
modulates PHLDA1 expression. In addition, HSF1 is able to
directly bind to and activate PHLDA1 in response to heat
shock in male germ cells (27).
Furthermore, during a time-course induction of cryptorchidism in
rat testes, HSF1 and PHLDA1 were strongly expressed, suggesting
that the HSF1/PHLDA1 pathways have an important role in the
apoptosis of primary spermatocytes in cryptorchid testes (28).
In addition to its association with apoptosis
induction, PHLDA1 has also been shown to act as an antiapoptotic
agent. PHLDA1 mRNA and protein are induced by insulin-like growth
factor 1 (IGF-1) through IGF-1R via the p38 MAPK pathway in NIH-3T3
(NWTb3) cells and via the phosphatidylinositol 3-kinase pathway in
normal skin fibroblasts (29,30). Knockdown of PHLDA1 expression by
small interfering RNA (siRNA) abolishes the protective effects of
IGF-1 against apoptosis induction in NIH-3T3 (NWTb3) cells in
response to serum starvation (30).
Toyoshima et al (29) also
identified that PHLDA1 downregulation increases apoptosis,
which is independent of IGF-1, suggesting that PHLDA1 may have
antiapoptotic effects in NWTb3 cells.
PHLDA1 is also a direct target of the oncogenic
fusion protein EWS/FLI1. Boro et al (31) demonstrated that EWS/FLI1 can bind to
the promoter of PHLDA1, in vivo and in vitro, leading
to its repression. In MEF cells derived from conditional EWS/FLI1
knockin embryos, EWS/FLI1 expression resulted in apoptosis and
EWS/FLI1 was able to directly bind the caspase 3 promoter in CHP100
cells with a concomitant increase in endogenous caspase 3
expression (32). In addition,
PHLDA1 knockdown by siRNA in Ca9-22 cells increases
activated caspase 3 expression, suggesting that PHLDA1 has a role
as an apoptosis suppressor (33).
Role of PHLDA1 in cancer
Several lines of evidence suggest that PHLDA1 has an
important role in cancer. Although the exact molecular and cellular
functions of PHLDA1 have not been fully elucidated, evidence from
experimental studies show that it is a potential transcriptional
activator that acts as a modulator of apoptosis and cell
proliferation, two biological processes that impact cell survival
and are considered hallmarks of cancer (34). In addition, altered expression of
PHLDA1 has been described in different types of tumor, such as
melanoma, breast and intestinal and gastric tumors (Table I).
 | Table I.Aberrant expression of PHLDA1 in
cancer. |
Table I.
Aberrant expression of PHLDA1 in
cancer.
| Type of cancer | Expression | Function | (Ref.) |
|---|
| Melanoma | Strongly expressed
in benign melanocytic nevi and progressively reduced in primary and
metastatic melanomas | Expression in
melanoma cell lines is associated with reduced cell growth and
colony formation, and increased basal apoptosis | (14) |
| Breast | Reduced expression,
a strong predictor of poor prognosis for breast cancer
patients |
| (15) |
|
| Reduced expression
associated with estrogen receptor expression in breast tumors | Regulatory feedback
loop between Aurora-A and PHLDA1 | (35) |
|
| Elevated expression
associated with a higher risk of distant metastasis in estrogen
receptor-positive breast cancer patients |
|
|
|
|
| PHLDA1
regulated by E2 and TNF-α at the | (40) |
|
|
| transcriptional and
post-transcriptional level in breast cancer cells | (41) |
|
|
| Silencing affects
mammosphere formation and growth | (41) |
|
|
| Reduced expression
during ras-mediated cellular transformation induces proliferation
under anchorage-independent conditions and increases ERK
activation | (42) |
|
|
| Upregulation found
in authophagy and apoptosis induced by rapamycin in breast cancer
cells | (26) |
|
| Expression reduced
in ERBB2-negative tumor samples and ERBB2-negative breast cancer
cell lines | Overexpression
inhibits cell proliferation, colony-forming ability, and migration
in SKBR3 cells | (36) |
| Oral | Reduced expression
is associated with good prognosis in oral cancer |
| (37) |
|
|
| Silencing leads to
apoptosis induction | (33) |
| Gastric | Reduced expression
associated with shorter overall survival in gastric cancer
patients |
| (16) |
| Basal cell
carcinoma | Reduced expression
in basal cell carcinoma |
| (44) |
| Colorectal | Overexpressed in
colorectal cancer | Silencing in colon
cancer cells inhibits | (38) |
|
|
|
anchorage-independent cell growth and
reduces cell migration | (17) |
| Intestinal | Overexpressed at
all stages of intestinal cancer development |
| (17) |
| Osteosarcoma |
| High expression
associated high metastatic potential in osteosarcoma cells, and
silencing delays overall survival in mouse xenograft models | (39) |
Reduced PHLDA1 expression has been described in
melanoma (14), breast carcinoma
(15,35,36), oral
carcinoma (37) and gastric
adenocarcinoma (16). This indicates
that PHLDA1 may act as a tumor suppressor gene. However, PHLDA1 was
reported to be upregulated in colon cancer and in human intestinal
adenoma and carcinoma, and PHLDA1 knockdown in colon cancer
cells inhibits anchorage-independent cell growth and reduces cell
migration (17,38). Furthermore, high expression of PHLDA1
is associated with high metastatic potential in osteosarcoma cells
and reduced PHLDA1 expression delays overall survival metastasis
progression in mouse xenograft models (39). These findings suggest that PHLDA1 has
tissue-specific functions and may have an oncogenic role in other
cancer types.
In melanoma, downregulation of PHLDA1 expression was
found in metastatic lesions but not in primary tumors, suggesting
that loss of PHLDA1 expression may contribute to the progression of
malignant melanoma (14). The effects
of PHLDA1 expression on the biological characteristics of different
melanoma-derived cell lines were also evaluated. PHLDA1 expression
leads to reduced cell growth and colony formation, increased basal
apoptosis, and increased sensitivity to apoptosis induced by
chemotherapeutic agents (14).
Reduced expression of PHLDA1 was observed in 60.7%
of oral squamous cell carcinomas (OSCC), particularly in
well-differentiated tumors. Positive PHLDA1 immunostaining was
associated with advanced clinical stages of the disease, suggesting
that PHLDA1 has a functional role in oral tumorigenesis. Overall
and disease-free survival rates were significantly improved in
patients with tumors that were negative for PHLDA1, and a
multivariate analysis suggested that PHLDA1 is an independent
prognostic factor in OSCC patients (37).
Downregulation of PHLDA1 mRNA and protein expression
is frequently observed in primary invasive breast tumors. In a
large series of primary breast tumors, Nagai et al (15) reported that downregulation of the
PHLDA1 protein is a strong predictor of poor prognosis for breast
cancer patients. This suggests that reduced PHLDA1 expression
contributes to breast cancer progression and may serve as a useful
prognostic biomarker of disease outcome. The study also found that
the subgroup of breast tumors that were negative for estrogen
receptor and expressed PHLDA1 had a more favorable outcome. PHLDA1
downregulation has also been associated with estrogen receptor
expression in breast tumors (35). In
addition, high PHLDA1 mRNA expression is associated with a
high risk of distant metastasis in estrogen receptor-positive
breast tumors. This suggests that in breast cancer cells, PHLDA1
may have different functions depending on the estrogen receptor
status, which affects the clinical behavior of tumors.
In an investigation of the effects of E2 and the
antiestrogen agent ICI 182780 on transcriptional regulation of
PHLDA1 in MCF7 breast cancer cells, Marchiori et al
(40) demonstrated that PHLDA1 is
upregulated by E2 via estrogen receptor in a time-dependent manner.
More recently, PHLDA1 was shown to be regulated by crosstalk
between ER and inflammatory nuclear factor-κB (NF-κB) (41). PHLDA1 is regulated by E2 and tumor
necrosis factor-α in MCF7 breast cancer cells through estrogen
receptor and NF-κB activation. Kastrati et al (41) also found that estrogen receptor and the
NF-κB pathway activation upregulates PHLDA1 directly by
transcriptional activation and by indirectly repressing the
expression of miR-181a and b, which directly target PHLDA1.
Reduction of PHLDA1 expression in HME16C mammary
epithelial cells during ras-mediated cellular transformation
induces proliferation under anchorage-independent conditions and
results in increased ERK activation, which suggests that PHLDA1
opposes ERK-mediated transformation in breast cancer cells
(42). In a profiling study with SKBR3
breast cancer cells treated with lapatinib, Li et al
(36) identified PHLDA1 as a novel
downstream target in the Erb-B2 receptor tyrosine kinase 2
(ERBB2)/epidermal growth factor receptor pathway and demonstrated
that PHLDA1 expression is reduced in ERBB2-negative breast tumors
and breast cancer cell lines. In addition, overexpression of PHLDA1
was found to inhibit proliferation, colony-formation ability and
migration in SKBR3 breast cancer cells (36).
A regulatory loop between Aurora-A and PHLDA1 was
also reported in breast cancer cells (35). In MDA-MB-231 breast cancer cells,
Aurora-A predominantly phosphorylates PHLDA1, leading to its
degradation. PHLDA1 overexpression results in decreased levels of
Aurora-A, suggesting that PHLDA1 acts as a negative regulator of
Aurora-A-mediated breast oncogenesis (35).
In an expression profiling study, Ohyama et
al (43) determined that PHLDA1 is
highly expressed and precisely localized in the bulge, which is the
multipotent compartment of the hair follicle. This indicates that
PHLDA1 may be an epithelial stem cell marker. The identification of
PHLDA1 as a potential epithelial stem cell marker has attracted the
attention of several investigators who are researching the possible
role of PHLDA1 in the tumorigenic process; however, this area of
investigation is in the early stages.
In an investigation of the expression patterns of
PHLDA1 in 16 desmoplastic trichoepitheliomas and 14 morphoeic basal
cell carcinomas (BCCs), the study by Sellheyer and Krahl (44) reported PHLDA1-positive immunostaining
in >80% of the trichoepitheliomas and PHLDA1-negative
immunostaining in all the BCCs examined, indicating that expression
of the hair follicle bulge marker PHLDA1 can be used to
differentiate between trichoepitheliomas and BCCs. Thus, PHLDA1
could be used as a biomarker in the differential diagnosis of
basaloid tumors.
Aberrant PHLDA1 expression has been described in
intestinal and colorectal cancers (17,38).
Upregulation of PHLDA1 mRNA expression was associated with
the presence of anemia in patients with colon cancer (38), and increased nuclear expression of
PHLDA1 was observed in the invasive margins of human colorectal
carcinomas (17). siRNA knockdown of
PHLDA1 expression in HCT116 and SW480 colon cancer cells
under anchorage-dependent growth leads to inhibition of cell growth
and colony-formation ability, and PHLDA1 downregulation also
inhibits cell migration in these colon cancer cells (17). In addition, strong PHLDA1-positive
immunostaining has been found in undifferentiated crypt base cells
that exhibit a similar pattern of LGR5 expression. LGR5 is a
putative epithelial stem cell marker for human small and large
intestine (17). In agreement with
this finding, Guezguez et al (45) determined that PHLDA1 is part of a
primordial stem-like cell signature in normal human intestinal
epithelial crypt cells treated with noggin, an antagonist to bone
morphogenetic protein signaling.
In an evaluation of a large number of gastric
carcinomas and normal gastric mucosa by immunohistochemisty, Zhao
et al (16) observed that
PHLDA1 is expressed diffusely in the cytoplasm and nucleus in crypt
base cells. PHLDA1 expression appeared to mark putative epithelial
stem cells in all the normal gastric mucosa analyzed. The study by
Zhao et al (16) identified
negative PHLDA1 expression in 57% of the gastric carcinomas and the
most poorly differentiated cases were negative for PHLDA1. Strong
nuclear staining was found in invasive margins. Furthermore,
reduced PHLDA1 expression was associated with shorter overall
survival in gastric cancer patients (16).
In breast cancer, PHLDA1 was identified among the
gene set of oncogenic ras-pathway and interferon-response
signatures in selected small populations of cluster of
differentiation (CD) 24−/low/CD44+ stem cells
from several breast cancer cell lines (46). More recently, high PHLDA1 expression
was shown to contribute to the survival and growth of breast cancer
stem cells (41). Kastrati et
al (41) also found upregulation
of PHLDA1 mRNA and protein expression in a panel of estrogen
receptor-positive breast cancer cells that were growing in
mammosphere cultures compared to cells growing in adherent
monolayer cultures. The study also observed that PHLDA1
knockdown affects mammosphere formation and growth, leading to a
reduction in the number and size of the mammospheres.
Conclusions
Over the last few years, PHLDA1 has attracted
increasing attention. However, the data reviewed herein show that
the precise molecular and cellular functions of PHLDA1 remain to be
fully elucidated. Although several lines of evidence have
implicated PHLDA1 as a potential transcriptional activator that
acts as a proapoptotic and antiproliferative factor, the exact
mechanisms by which PHLDA1 mediates cell survival are still under
investigation. PHLDA1 expression can be modulated by various
stimuli, resulting in pleiotropic effects, with upregulation and
downregulation of different biological processes, such as
proliferation, apoptosis, migration and invasion, which are
considered key hallmarks of carcinogenesis. The pleiotropic effects
of PHLDA1 indicate tissue-specific activities that depend on the
cellular context. PHLDA1 is expressed in different tissues with
variable subcellular localization and function, and its expression
pattern is altered in different types of tumors. Data gathered thus
far indicate that PHLDA1 has an important role in the tumorigenic
process; however, new results from clinical and experimental
studies are necessary to improve the definition of its role as a
tumor suppressor gene or oncogene and its contribution to tumor
progression. Further, well-designed clinical and experimental
studies, in vitro and in vivo, that use informative
and testable animal models are required to improve the
understanding of the PHLDA1 mechanisms of action in normal and
cancer cells. In addition, the identification of PHLDA1 as a
potential stem cell marker and evidence of its possible roles in
the EMT indicate that there are new perspectives and research
fields to be explored to evaluate the actual contributions of
PHLDA1 to the tumorigenic process and its role not only as
prognostic factor, but also as a potential therapeutic target.
Glossary
Abbreviations
Abbreviations:
|
aa
|
amino acid
|
|
AICD
|
activation-induced cell death
|
|
BCC
|
basal cell carcinoma
|
|
EMT
|
epithelial-mesenchymal transition
|
|
ER
|
endoplasmic reticulum
|
|
FGF
|
fibroblast growth factor
|
|
IGF-1
|
insulin-like growth factor 1
|
|
NF-κB
|
nuclear factor-κB
|
|
OSCC
|
oral squamous cell carcinoma
|
|
PH
|
proline-histidine
|
|
PHL
|
pleckstrin homology-like
|
|
PHLDA1
|
pleckstrin homology-like domain,
family A, member 1
|
|
PKC
|
protein kinase C
|
|
PQ
|
proline-glutamine
|
|
QQ
|
polyglutamine
|
References
|
1
|
Park CG, Lee SY, Kandala G, Lee SY and
Choi Y: A novel gene product that couples TCR signaling to
Fas(CD95) expression in activation-induced cell death. Immunity.
4:583–591. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kuske MD and Johnson JP: Assignment of the
human PHLDA1 gene to chromosome 12q15 by radiation hybrid mapping.
Cytogenet Cell Genet. 89:12000. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Frank D, Mendelsohn CL, Ciccone E,
Svensson K, Ohlsson R and Tycko B: A novel pleckstrin
homology-related gene family defined by Ipl/Tssc3, TDAG51, and
Tih1: Tissue-specific expression, chromosomal location, and
parental imprinting. Mamm Genome. 10:1150–1159. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gomes I, Xiong W, Miki T and Rosner MR: A
proline- and glutamine-rich protein promotes apoptosis in neuronal
cells. J Neurochem. 73:612–622. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lemmon MA, Ferguson KM and Abrams CS:
Pleckstrin homology domains and the cytoskeleton. FEBS Lett.
513:71–76. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Xu Q, Bateman A, Finn RD, Abdubek P,
Astakhova T, Axelrod HL, Bakolitsa C, Carlton D, Chen C, Chiu HJ,
et al: Bacterial pleckstrin homology domains: A prokaryotic origin
for the PH domain. J Mol Biol. 396:31–46. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lemmon MA and Ferguson KM:
Signal-dependent membrane targeting by pleckstrin homology (PH)
domains. Biochem J. 350:1–18. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Scheffzek K and Welti S: Pleckstrin
homology (PH) like domains - versatile modules in protein-protein
interaction platforms. FEBS Lett. 586:2662–2673. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Williamson MP: The structure and function
of proline-rich regions in proteins. Biochem J. 297:249–260. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Butland SL, Devon RS, Huang Y, Mead CL,
Meynert AM, Neal SJ, Lee SS, Wilkinson A, Yang GS, Yuen MM, et al:
CAG-encoded polyglutamine length polymorphism in the human genome.
BMC Genomics. 8:1262007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu CR and Cheng TH: Allele-selective
suppression of mutant genes in polyglutamine diseases. J
Neurogenet. 29:41–49. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hossain GS, van Thienen JV, Werstuck GH,
Zhou J, Sood SK, Dickhout JG, de Koning AB, Tang D, Wu D, Falk E,
et al: TDAG51 is induced by homocysteine, promotes
detachment-mediated programmed cell death, and contributes to the
cevelopment of atherosclerosis in hyperhomocysteinemia. J Biol
Chem. 278:30317–30327. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hinz T, Flindt S, Marx A, Janssen O and
Kabelitz D: Inhibition of protein synthesis by the T cell
receptor-inducible human TDAG51 gene product. Cell Signal.
13:345–352. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Neef R, Kuske MA, Pröls E and Johnson JP:
Identification of the human PHLDA1/TDAG51 gene: Down-regulation in
metastatic melanoma contributes to apoptosis resistance and growth
deregulation. Cancer Res. 62:5920–5929. 2002.PubMed/NCBI
|
|
15
|
Nagai MA, Fregnani JH, Netto MM, Brentani
MM and Soares FA: Down-regulation of PHLDA1 gene expression is
associated with breast cancer progression. Breast Cancer Res Treat.
106:49–56. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhao P, Lu Y and Liu L: Correlation of
decreased expression of PHLDA1 protein with malignant phenotype of
gastric adenocarcinoma. Int J Clin Exp Pathol. 8:5230–5235.
2015.PubMed/NCBI
|
|
17
|
Sakthianandeswaren A, Christie M,
D'Andreti C, Tsui C, Jorissen RN, Li S, Fleming NI, Gibbs P, Lipton
L, Malaterre J, et al: PHLDA1 expression marks the putative
epithelial stem cells and contributes to intestinal tumorigenesis.
Cancer Res. 71:3709–3719. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Mohammad RM, Muqbil I, Lowe L, Yedjou C,
Hsu HY, Lin LT, Siegelin MD, Fimognari C, Kumar NB, Dou QP, et al:
Broad targeting of resistance to apoptosis in cancer. Semin Cancer
Biol. 35:S78–S103. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rho J, Gong S, Kim N and Choi Y: TDAG51 is
not essential for Fas/CD95 regulation and apoptosis in vivo. Mol
Cell Biol. 21:8365–8370. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Oberg HH, Sipos B, Kalthoff H, Janssen O
and Kabelitz D: Regulation of T-cell death-associated gene 51
(TDAG51) expression in human T-cells. Cell Death Differ.
11:674–684. 2004.PubMed/NCBI
|
|
21
|
Wang R, Zhang L, Yin D, Mufson RA and Shi
Y: Protein kinase C regulates Fas (CD95/APO-1) expression. J
Immunol. 161:2201–2207. 1998.PubMed/NCBI
|
|
22
|
Joo JH, Liao G, Collins JB, Grissom SF and
Jetten AM: Farnesol-induced apoptosis in human lung carcinoma cells
is coupled to the endoplasmic reticulum stress response. Cancer
Res. 67:7929–7936. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Carlisle RE, Heffernan A, Brimble E, Liu
L, Jerome D, Collins CA, Mohammed-Ali Z, Margetts PJ, Austin RC and
Dickhout JG: TDAG51 mediates epithelial-to-mesenchymal transition
in human proximal tubular epithelium. Am J Physiol Renal Physiol.
303:F467–F481. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shang Y, Cai X and Fan D: Roles of
epithelial-mesenchymal transition in cancer drug resistance. Curr
Cancer Drug Targets. 13:915–929. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Pasquier J, Abu-Kaoud N, Al Thani H and
Rafii A: Epithelial to Mesenchymal Transition in a Clinical
Perspective. J Oncol. 2015:7921822015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Moad AI, Muhammad TS, Oon CE and Tan ML:
Rapamycin induces apoptosis when autophagy is inhibited in T-47D
mammary cells and both processes are regulated by Phlda1. Cell
Biochem Biophys. 66:567–587. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hayashida N, Inouye S, Fujimoto M, Tanaka
Y, Izu H, Takaki E, Ichikawa H, Rho J and Nakai A: A novel
HSF1-mediated death pathway that is suppressed by heat shock
proteins. EMBO J. 25:4773–4783. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liu F, Xu ZL, Qian XJ, Qiu WY and Huang H:
Expression of Hsf1, Hsf2, and Phlda1 in cells undergoing
cryptorchid-induced apoptosis in rat testes. Mol Reprod Dev.
78:283–291. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Toyoshima Y, Karas M, Yakar S, Dupont J,
Lee Helman and Le Roith D: TDAG51 mediates the effects of
insulin-like growth factor I (IGF-I) on cell survival. J Biol Chem.
279:25898–25904. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wu S, Walenkamp MJ, Lankester A,
Bidlingmaier M, Wit JM and De Luca F: Growth hormone and
insulin-like growth factor I insensitivity of fibroblasts isolated
from a patient with an IκBα mutation. J Clin Endocrinol Metab.
95:1220–1228. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Boro A, Prêtre K, Rechfeld F, Thalhammer
V, Oesch S, Wachtel M, Schäfer BW and Niggli FK: Small-molecule
screen identifies modulators of EWS/FLI1 target gene expression and
cell survival in Ewing's sarcoma. Int J Cancer. 131:2153–2164.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sohn EJ, Li H, Reidy K, Beers LF,
Christensen BL and Lee SB: EWS/FLI1 oncogene activates caspase 3
transcription and triggers apoptosis in vivo. Cancer Res.
70:1154–1163. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Murata T, Sato T, Kamoda T, Moriyama H,
Kumazawa Y and Hanada N: Differential susceptibility to hydrogen
sulfide-induced apoptosis between PHLDA1-overexpressing oral cancer
cell lines and oral keratinocytes: Role of PHLDA1 as an apoptosis
suppressor. Exp Cell Res. 320:247–257. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Johnson EO, Chang KH, de Pablo Y, Ghosh S,
Mehta R, Badve S and Shah K: PHLDA1 is a crucial negative regulator
and effector of Aurora A kinase in breast cancer. J Cell Sci.
124:2711–2722. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li G, Wang X, Hibshoosh H, Jin C and
Halmos B: Modulation of ErbB2 blockade in ErbB2-positive cancers:
The role of ErbB2 Mutations and PHLDA1. PLoS One. 9:e1063492014.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Coutinho-Camillo CM, Lourenço SV, Nonogaki
S, Vartanian JG, Nagai MA, Kowalski LP and Soares FA: Expression of
PAR-4 and PHLDA1 is prognostic for overall and disease-free
survival in oral squamous cell carcinomas. Virchows Arch.
463:31–39. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chiu ST, Hsieh FJ, Chen SW, Chen CL, Shu
HF and Li H: Clinicopathologic correlation of up-regulated genes
identified using cDNA microarray and real-time reverse
transcription-PCR in human colorectal cancer. Cancer Epidemiol
Biomarkers Prev. 14:437–443. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ren L, Mendoza A, Zhu J, Briggs JW, Halsey
C, Hong ES, Burkett SS, Morrow J, Lizardo MM, Osborne T, et al:
Characterization of the metastatic phenotype of a panel of
established osteosarcoma cells. Oncotarget. 6:29469–29481.
2015.PubMed/NCBI
|
|
40
|
Marchiori AC, Casolari DA and Nagai MA:
Transcriptional up-regulation of PHLDA1 by 17beta-estradiol in
MCF-7 breast cancer cells. Braz J Med Biol Res. 41:579–582. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kastrati I, Canestrari E and Frasor J:
PHLDA1 expression is controlled by an estrogen
receptor-NFκB-miR-181 regulatory loop and is essential for
formation of ER+ mammospheres. Oncogene. 34:2309–2316. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Oberst MD, Beberman SJ, Zhao L, Yin JJ,
Ward Y and Kelly K: TDAG51 is an ERK signaling target that opposes
ERK-mediated HME16C mammary epithelial cell transformation. BMC
Cancer. 8:189–204. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ohyama M, Terunuma A, Tock CL, Radonovich
MF, Pise-Masison CA, Hopping SB, Brady JN, Udey MC and Vogel JC:
Characterization and isolation of stem cell-enriched human hair
follicle bulge cells. J Clin Invest. 116:249–260. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Sellheyer K and Krahl D: PHLDA1 (TDAG51)
is a follicular stem cell marker and differentiates between
morphoeic basal cell carcinoma and desmoplastic trichoepithelioma.
Br J Dermatol. 164:141–147. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Guezguez A, Paré F, Benoit YD, Basora N
and Beaulieu JF: Modulation of stemness in a human normal
intestinal epithelial crypt cell line by activation of the WNT
signaling pathway. Exp Cell Res. 322:355–364. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Murohashi M, Hinohara K, Kuroda M, Isagawa
T, Tsuji S, Kobayashi S, Umezawa K, Tojo A, Aburatani H and Gotoh
N: Gene set enrichment analysis provides insight into novel
signalling pathways in breast cancer stem cells. Br J Cancer.
102:206–212. 2010. View Article : Google Scholar : PubMed/NCBI
|