Introduction
Tumor growth and metastasis are angiogenesis- and
lymph-angiogenesis-dependent events (1). By inducing endothelial cell
proliferation and migration, vascular endothelial growth factor
(VEGF) signaling plays an important role in promoting tumor
metastasis, and this pathway has become a promising target for
cancer therapy (2). The secreted
glycoprotein VEGF-A causes proliferation, sprouting, migration and
tube formation of endothelial cells, and has been demonstrated to
play a crucial role in tumor angiogenesis (3). VEGF-A binds to the transmembrane
receptor VEGFR-2 (4,5), leading to the activation of the
downstream signal transduction pathway and endothelial
proliferation-related genes (6).
Increased VEGF-A expression levels have been detected in numerous
types of human cancer, including lung, gastrointestinal tract,
kidney, thyroid, bladder, ovarian and cervical tumors (7). Animal models have confirmed that the
growth of a large variety of tumors is suppressed by the inhibition
of VEGF-A signaling. For example, our previous study showed that
blocking VEGF-A expression effectively inhibited bladder cancer
angiogenesis, as well as tumor growth and metastasis (8).
By enabling cancer cells to gain access to the
lymphatic system, lymph vessels in tumors provide another route for
metastasis. It was recently discovered that VEGF-A induces
lymphatic vessel growth in tumors and promotes metastasis to local
lymph nodes (5). VEGF-C and its
receptor VEGFR-3 also stimulate the proliferation and migration of
lymphatic endothelial cells, which in turn enhances the assembly of
new lymphatic vessels (9,10). The binding of VEGF-C to VEGFR-3
stimulates lymphatic growth in tumors (10), and patients with high intratumoral
levels of VEGF-C have poorer prognoses than those who express
little or no VEGF-C (11). Recent
studies have shown that overexpression of VEGF-C in several types
of human tumors, including bladder cancer, may greatly increase
intratumoral lymphangiogenesis, leading to significant increases in
regional lymph node metastasis (4,5).
These findings indicate that blocking both tumor angiogenesis and
lymphangiogenesis concurrently may provide an optimal strategy for
cancer therapy.
Small interfering RNAs (siRNAs) inhibit the
expression of specific target genes via the evolutionarily
conserved mechanism of RNA interference (RNAi) (6,12).
Transgenic RNAi is a powerful tool for silencing mammalian genes
with a high degree of specificity and efficacy, and is considered a
promising therapeutic approach for treating various diseases,
including cancer (13,14). In this study, we transfected siRNA
constructs to knock down the expression of VEGF-A, VEGF-C and
VEGFR-3 in green fluorescent protein (GFP)-expressing T739 mouse
bladder transitional carcinoma (BTT) cells, and investigated the
effects on tumor growth and metastasis. siRNA-mediated knockdown of
VEGF signaling was found to suppress tumor growth and reduce tumor
metastasis in vivo.
Materials and methods
siRNA synthesis
siRNA sequences were designed using the software
available at http://www.ambion.com/techlib/misc/siRNA_design.html
followed by a BLAST search to eliminate non-unique targeting
sequences. Three sequences targeting VEGF-A, four targeting VEGF-C
and six targeting VEGFR-3 were selected.
VEGF-A siRNA sequences containing
BamHI/HindIII sites were synthesized and cloned into
pSilencer puro H1 vector according to the manufacturer’s protocol
(Ambion). After transformation, the presence and orientation of the
VEGF-A siRNA inserts in plasmid DNA clones were confirmed by double
digestion with BamHI and HindIII enzymes (Takara).
The plasmid DNA (isolated with mini-prep kits; Qiagen) was also
sequenced to confirm the presence of the VEGF-A siRNAs. VEGF-C and
VEGFR-3 siRNAs were synthesized in vitro and purified using
Silencer® siRNA construction kits (Ambion) according to
the product manuals. The end product was a double-stranded 21-mer
siRNA with 3′ terminal uridine dimers that effectively reduce the
expression of target mRNA when transfected into mammalian cells.
The scrambled siRNA negative control had the same
construct-containing sequence and no homology with any genes.
siRNAs used in vivo were synthesized and modified by
Shanghai GenePharma Co., Ltd.
Cell cultures and transfections
BTT-T739-GFP mouse bladder cancer cells stably
expressing GFP were grown in RPMI-1640 medium (Invitrogen)
containing 10% FBS with penicillin/streptomycin antibiotics (100
units of penicillin and 100 μg of streptomycin per ml). VEGF-A
siRNA plasmid DNA, VEGF-C and VEGFR-3 siRNAs were transfected using
Lipofectamine 2000 (Invitrogen). Cells (8×104) were
seeded in 24-well plates the day before transfection. Plasmid DNA
(1 μg) or siRNA and (2 μl) Lipofectamine 2000 were applied
according to the protocol provided by the manufacturer. Each
condition was performed in triplicate. Cells transfected with a
scrambled siRNA construct were used as the negative control. Total
RNA was isolated from cells using TRIzol reagent (Invitrogen) 48 h
after transfection.
Real-time RT-PCR analysis
cDNA was amplified from total RNA using a reverse
transcription system (Promega). SYBR Green quantitative PCR
amplifications were performed in a MJ Opticon2 System (BioRad).
Reactions were carried out in 20-μl volumes containing 10 μl of 2X
SYBR Green PCR MasterMix (Takara). The cycle profile for real-time
PCR was 95°C for 2 min followed by 40 cycles of 95°C for 30 sec,
58°C for 30 sec and 72°C for 30 sec. Glyceraldehyde 3-phosphate
dehydrogenase (GAPDH) was used as the internal control to normalize
gene expression levels. Ct values were collected, and the average
ΔCt for each group was calculated using the following formula: ΔCt
= average target gene Ct − average GAPDH Ct. ΔΔCt was calculated by
the formula: ΔΔCt = ΔCt of control group − ΔCt of the siRNA-treated
group. The relative expression levels of target genes were
calculated using the 2−ΔΔCt method. Each
reaction was performed in triplicate.
ELISA and Western blotting
To measure VEGF-A concentration in the supernatant
from VEGF-A siRNA plasmid-transfected BTT-T739-GFP cells, a mouse
VEGF-A Quantikine Colorimetric Sandwich ELISA kit (R&D Systems)
was used. All tests were performed in triplicates. The
determination of VEGF-C protein expression in VEGF-C
siRNA-transfected BTT-T739-GFP cells was carried out using Western
blotting according to the standard method.
In vivo siRNA therapy
Animals in this research were maintained and studied
according to local and national regulations on animal welfare, and
the licensing committee approved the experiments. Twenty-two male
6- to 8-week-old T-739 mice were subcutaneously injected with
1×106 tumor cells/mouse. A week later, when the tumors
were palpable, the mice were randomly assigned into each of three
groups and anesthetized by an intraperitoneal injection of 50 mg/kg
pentobarbital. Each mouse received an intratumoral injection of 10
μg siRNA/mouse: scrambled siRNA (negative control group, NC; n=6),
siVEGF-C + siVEGFR-3 (n=8) or siVEGF-A (n=8). Following the
injections, the tumors were electroporated with 8 pulses of 900
V/cm for 100 μm at 1 Hz (15).
Injections and electroporations were repeated once every three days
a total of 5 times. Tumor length and width were measured with
calipers, and tumor volume was calculated with the formula: Volume
= 0.52 × (width × length2). Animals were sacrificed 1
week after the final injection, and livers, lungs and tumors were
excised for molecular biology and histological analysis. The
regional lymph nodes from axillary and femoral regions were
isolated, and lymph node volume was calculated as: Volume = 0.5 ×
(width × length × height).
Relevant gene expression level in tumor
tissues
VEGF-A, VEGF-C, VEGFR-2 and VEGFR-3 mRNA expression
levels in tumor tissues were detected to ascertain whether the
siRNAs effectively knock down the relevant gene expression in
vivo. The expression level of these genes also reflects the
state of angiogenesis and lymphogenesis in tumors. To accomplish
this, total RNA was extracted from tumor tissues after siRNA
treatment, and cDNA and real-time PCR was carried out according to
the the method described above. Each real-time PCR reaction was
carried out in triplicates.
Analysis of tumor metastasis
GFP-positive tumor cells in unfixed whole organ
samples were visualized using fluorescent microscopy. Livers and
lungs were then fixed in neutral formalin for histological
analysis.
Visualization of lymph nodes by Carnalym
lymphatic tracer
To visualize the regional lymph nodes, 30 μl of 50
mg/ml Carnalym lymphatic tracer (provided by the Chengdu Institute
of High-Technology Medicine) was injected into the tumors.
Twenty-four hours later, the animals were sacrificed, and the
regional lymph nodes from the axillary and femoral regions were
isolated for observation under a stereomicro-scope.
Statistical analysis
Statistical analyses were carried out using SPSS
software (version 11.0). Data were expressed as the means ± SEM and
analyzed by one-way ANOVA, the independent-samples t-test and the
least significant difference test. p<0.05 was considered
significant.
Results
siRNA-mediated silencing of VEGF-A,
VEGF-C and VEGFR-3 in mouse bladder cancer cells
The inhibitory effect of siRNAs targeting the VEGF
pathway was tested in BTT-T739-GFP cells, a mouse bladder cancer
cell line that expresses high levels of VEGF-A, VEGF-C and VEGFR-3
(16). VEGF-A expression was
measured by real-time RT-PCR following in vitro transfection
with one of three plasmids encoding unique VEGF-A siRNA sequences
(siVEGF-A #1–3). Of the three, siVEGF-A #3 greatly reduced VEGF-A
expression relative to the negative control. Specific siRNAs
against VEGF-C (siVEGF-C #3 and 4) and VEGFR-3 (siVEGFR-3 #3 and 5)
also significantly reduced target gene expression relative to the
negative controls (p<0.05; Fig.
1A). VEGF-A protein levels in the cell culture supernatant were
measured by ELISA, and siVEGF-A #3 showed the lowest level
(Fig. 1B). siVEGF-C #3 showed the
most inhibited level as noted by Western blotting (Fig. 1C). Thus, the most effective siRNA
sequences, siVEGF-A #3 (ATGTGAATGCAGACCAAAG), siVEGF-C #3
(AACAGAGGCCCAAGTCTGTGT) and siVEGFR-3 #5 (GAAGCCCAATCAATAACTG) were
used for subsequent in vivo experiments.
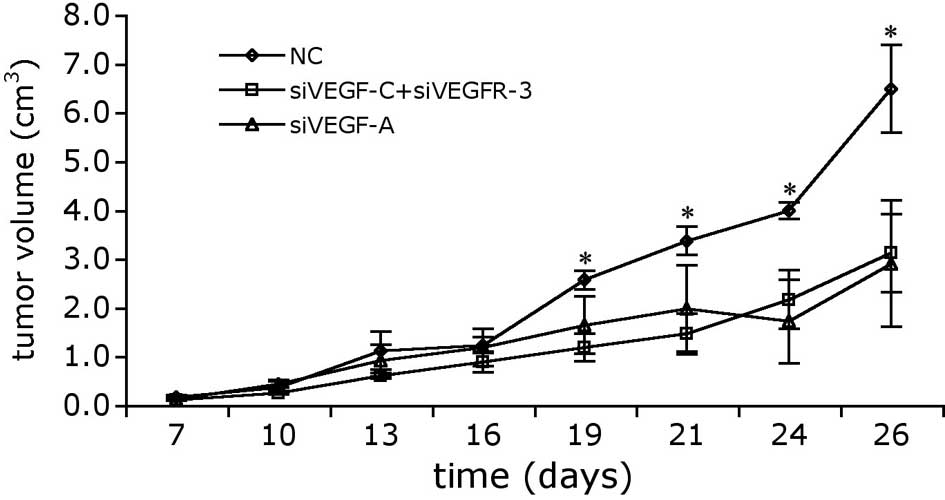
VEGF-A, VEGF-C and VEGFR-3 siRNAs inhibit
the growth of BTT-T739-GFP cell xenografts
To determine whether our siRNAs inhibited tumor
growth in vivo, BTT-T739-GFP cells were subcutaneously
injected into adult mice (1×106 cells/mouse). Tumors
were allowed to develop for 1 week and then a regimen of siRNA
therapy was administered. siRNAs were delivered directly into the
tumor by electroporation once every 3 days a total of 5 times, and
tumor growth was monitored over 26 days (7 days prior to and
during, and 7 days following treatment). Tumor volume was
calculated by measuring their width and length. As shown in
Fig. 2, delivering siVEGF-A alone
or siVEGF-C + siVEGFR-3 dramatically reduced the growth rate of
BTT-T739 tumors relative to the scramble siRNA-transfected negative
controls. Significant differences in tumor size were noted on Day
19, and the margin of difference widened over the remaining 7
observation days. By the end of the 26-day period, the mean tumor
size in the siVEGF-A group was 36.1% smaller than that in the
negative control group. Delivering siVEGFR-3 together with siVEGF-C
had an effect similar to that of siVEGF-A alone, exhibiting
significant effects by Day 19 and reducing tumor growth by 55.4% by
the end of the 26-day observation period.
mRNA expression of related genes was
down-regulated in tumor tissues after siRNA therapy
Real-time RT-PCR was applied to measure the mRNA
levels of VEGF-A, VEGFR-C, VEGFR-2 and VEGFR-3 in tumor tissues
after siRNA therapy. VEGF-A was significantly reduced in the
siVEGF-A group, which showed that siVEGF-A efficiently knocked down
the expression of VEGF-A in the tumors. VEGF-A receptor, VEGFR-2,
which partially represents vascular vessel density, was also
down-regulated in both siRNA therapy groups (Fig. 3). VEGF-C and VEGFR-3 expression was
decreased in the siRNA therapy groups, but without significance.
The results showed that angiogenesis and lymphangiogenesis in the
tumors were inhibited after siRNA therapy.
Effects of siRNA therapy on lymph node
volume
Lymph nodes play important roles during tumor
metastasis. Groin and axilla lymph nodes ipsilateral and
contralateral to the tumor were dissected and measured after the
26-day observation period. The volume of the lymph nodes was
calculated as Volume = 0.52 × (length × width)2. The
volume of the ipsilateral groin lymph nodes was much larger in both
treatment groups than in the controls (Fig. 4), likely due to the reactive
proliferation of the lymph nodes (22). On the other hand, the volume of the
contralateral groin lymph nodes and axilla lymph nodes was not
affected in either siRNA treatment group.
Reduced metastases following treatment
with siVEGF-A or siVEGF-C + siVEGFR-3
BTT-T739-GFP cells express GFP; thus, tumor
metastasis is directly observed in whole organs under a fluorescent
microscope. Whole lungs and livers were removed 26 days after tumor
cell injection, and the numbers of BTT-T739-GFP aggregates were
counted. In the siVEGF-C + siVEGFR-3 group, no obvious reduction in
lung metastasis and reduced liver metastasis were observed compared
to the negative control group. In the group treated with siVEGF-A
alone, metastases in the lung were reduced compared to the negative
control group, and no metastatic tumor cells were found in the
livers (Table I).
 | Table I.Grade of tumor metastasis in the liver
and lung. |
Table I.
Grade of tumor metastasis in the liver
and lung.
| | Lung (%)
| Liver (%)
|
|---|
| n | + | ++ | +++ | ND | + | ++ | +++ | ND |
|---|
| Negative control | 6 | 33.33 | 16.67 | 50.00 | 0.00 | 33.33 | 16.67 | 0.00 | 50.00 |
| siVEGF-C +
siVEGFR-3 | 8 | 37.50 | 12.50 | 50.00 | 0.00 | 25.00 | 0.00 | 0.00 | 75.00 |
| siVEGF-A | 8 | 25.00 | 37.50 | 0.00 | 37.50 | 0.00 | 0.00 | 0.00 | 100.00 |
Carnalym shows lymphatic drain from the
tumors
Regional lymph nodes were dissected and observed 24
h after the injection of Carnalym into the tumors. Blackened
regional lymph nodes or lymphatics were noted in the axillary and
femoral regions (Fig. 5)
suggesting drainage from the tumors.
Discussion
Previous studies have demonstrated that
VEGF-C-targeting RNAi effectively suppresses the lymphatic
metastasis of human breast cancer cells (17) and that, by inhibiting angiogenesis,
VEGF-targeted RNAi suppresses retinoblastoma and Ewing’s sarcoma
(18,19). In this study, we designed and
selected siRNAs targeting VEGF-A, VEGF-C and VEGFR-3, which
effectively reduced target gene expression in bladder cancer cells
in vitro. These siRNAs were then tested for their effects on
bladder cancer cell proliferation and metastasis in vivo.
Our results showed that tumor growth was significantly suppressed
after treating tumors with siVEGF-A as well as siVEGF-C + siVEGFR-3
relative to the negative control-treated tumors. VEGF-A, VEGF-C,
VEGFR-2 and VEGFR-3 expression was also reduced in the tumors after
treatment, showing that the siRNAs used efficiently interfered with
the relevant gene expression both in vitro and in
vivo and led to reduced angiogenesis and lymphangiogenesis in
the tumors. In addition, siVEGF-A therapy significantly reduced the
incidence of lung and liver metastases, and a combinational therapy
of siVEGF-C + siVEGFR-3 significantly reduced liver metastasis.
Notably, the injection of VEGF-A siRNAs into
BTT-T739 tumors resulted in the enlarged volume of ipsilateral
groin lymph nodes in comparison to the controls, which was
morphologically confirmed to contain a reactive proliferation.
These regional lymph nodes showed drainage from the tumors, which
was visualized by Carnalym lymphatic tracer. Another research group
also reported that lymphatic drainage near the tumor sites enlarged
lymph nodes without evidence of metastasis (20). These phenomena may represent
microenvironmental changes induced by the primary tumor prior to
metastasis and may be a future site for a secondary tumor (21). However, we deduce that these
phenomena may reflect a host antitumor reaction and prohibit
metastatic tumor cell growth.
The siRNAs used in this study were chemically
modified to allow broad biodistribution and effective cell entry
(22). In addition, in vivo
gene electrotransfer was applied as a means of delivering siRNAs to
the tumor cells. This technique offers some attractive advantages,
such as high efficiency of DNA transfer in vivo, ease of
manipulation and high gene accumulation through repeated
transfections, providing an efficient method of delivering genes
into tumors (23). Our results
demonstrated that decreasing VEGF gene expression in bladder cancer
cell tumors by this means suppressed tumor growth and metastasis.
Thus, siRNA-mediated inhibition of VEGF-A, VEGF-C or VEGFR-3
expression may be an option for arresting tumor growth and
metastasis and improving disease-free survival rates of patients
with bladder cancer.
Acknowledgements
This study was supported by grants
from the National Basic Research Program of China (973 project,
2004CB518804), the National Natural Science Foundation of China
(30325043), the Project for Talent Accomplishment of the Health
Bureau of Shanghai, China (99BR006), and the Rising-Star Program of
Shanghai, China (04QMX1417).
References
|
1.
|
Stacher SA, Achen MG, Jussila L, Baldwin
ME and Alitalo K: Lymphangiogenesis and cancer metastasis. Nat Rev
Cancer. 2:573–583. 2002. View
Article : Google Scholar
|
|
2.
|
Carmeliet P and Jain RK: Angiogenesis in
cancer and other diseases. Nature. 407:249–257. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Ferrara N, Gerber HP and LeCouter J: The
biology of VEGF and its receptors. Nat Med. 9:669–676. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Saban MR, Towner R, Smith N, et al:
Lymphatic vessel density and function in experimental bladder
cancer. BMC Cancer. 7:2192007. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Mylona E, Magkou C, Gorantonakis G, et al:
Evaluation of the vascular endothelial growth factor (VEGF)-C role
in urothelial carcinomas of the bladder. Anticancer Res.
26:3567–3571. 2006.PubMed/NCBI
|
|
6.
|
Fire A, Xu S, Montgomery MK, Kostas SA,
Driver SE and Mello CC: Potent and specific genetic interference by
double-stranded RNA in Caenorhabditis elegans. Nature.
391:806–811. 1998. View
Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Ferrara N: Molecular and biological
properties of vascular endothelial growth factor. J Mol Med.
77:527–543. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
8.
|
Wang F, Wu JH, Tian YH, et al: Role of
VEGF in the growth and metastasis of a murine bladder carcinoma.
Chin Sci Bull. 48:2404–2410. 2003.
|
|
9.
|
Joukov V, Pajusola K, Kaipainen A, et al:
A novel vascular endothelial growth factor. VEGF-C, is a ligand for
the Flt-4 (VEGFR-3) and KDR (VEGFR-2) receptor tyrosine kinases.
EMBO J. 15:290–298. 1996.
|
|
10.
|
Oh SJ, Jeltsch MM, Birkenhäger R, et al:
VEGF and VEGF-C: specific induction of angiogenesis and
lymphangiogenesis in the differentiated avian chorioallantoic
membrane. Dev Biol. 188:96–109. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
Zu X, Tang Z, Li Y, Gao N, Ding J and Qi
L: Vascular endothelial growth factor-C expression in bladder
transitional cell cancer and its relationship to lymph node
metastasis. BJU Int. 98:1090–1093. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Hammond SM, Caudy AA and Hannon GJ:
Post-transcriptional gene silencing by double-stranded RNA. Nat Rev
Genet. 2:110–119. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Devi GR: siRNA-based approaches in cancer
therapy. Cancer Gene Ther. 13:819–829. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Karagiannis TC and El-Osta A: RNA
interference and potential therapeutic applications of short
interfering RNAs. Cancer Gene Ther. 12:787–795. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Elbashir SM, Harborth J, Lendeckel W,
Yalcin A, Weber K and Tuschl T: Duplexes of 21-nucleotide RNAs
mediate RNA interference in cultured mammalian cells. Nature.
411:494–498. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Wang F, Tian YH, Chen XF and Huang Q:
Microarray analysis the role of VEGF signaling pathway for the
growth and metastasis of routine bladder earcinonla. Tumor.
27:847–885. 2007.
|
|
17.
|
Sun P, Gao J, Liu YL, Wei LW, Wu LP and
Liu ZY: RNA interference (RNAi)-mediated vascular endothelial
growth factor-C (VEGF-C) reduction interferes with
lymphangiogenesis and enhances epirubicin sensitivity of breast
cancer cells. Mol Cell Biochem. 308:161–168. 2008. View Article : Google Scholar
|
|
18.
|
Jia RB, Zhang P, Zhou YX, et al:
VEGF-targeted RNA interference suppresses angiogenesis and tumor
growth of retinoblastoma. Ophthalmic Res. 39:108–115. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Guan H, Zhou Z, Wang H, Jia SF, Liu W and
Kleinerman ES: A small interfering RNA targeting vascular
endothelial growth factor inhibits Ewing’s sarcoma growth in a
xenograft mouse model. Clin Cancer Res. 11:2662–2669.
2005.PubMed/NCBI
|
|
20.
|
Ioachim HLRH: Tumor-Reactive
Lymphadenopathy. 3rd edition. Lippincott Williams and Wilkins;
Philadelphia: pp. 254–258. 2002
|
|
21.
|
Qian CN, Berghuis B, Tsarfaty G, et al:
Preparing the ‘soil’: the primary tumor induces vasculature
reorganization in the sentinel lymph node before the arrival of
metastatic cancer cells. Cancer Res. 66:10365–10376. 2006.
|
|
22.
|
Manoharan M: RNA interference and
chemically modified small interfering RNAs. Curr Opin Chem Biol.
8:570–579. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Wang F, Chen XF, Tian YH, Wu JH, Li L, Li
CY and Huang Q: Target gene transfer mediated by electroporation
for cancer therapy in vivo. Prog Biochem Biophys. 29:734–740.
2002.
|