Introduction
Non-alcoholic fatty liver disease (NAFLD) is a
clinicopathological disease characterized by elevated accumulation
of triglycerides in the liver. In more than 10% of patients with
NAFLD, liver disease progresses to non-alcoholic steatohepatitis
(NASH), which is characterized by hepatic inflammatory cell
infiltration and ballooning of hepatocytes. Liver cirrhosis and
hepatocellular carcinoma occur in certain patients with NASH
(1).
Many studies have attempted to investigate the
mechanisms involved in the progression of hepatic steatosis to
control lipid content in NAFLD. Steatosis occurs when the rate of
hepatic fatty acid uptake from plasma and de novo fatty acid
synthesis is greater than the rate of fatty acid oxidation and
excretion as very low-density lipoprotein (VLDL) (2). Insulin resistance (IR), which is
frequently accompanied by obesity and T2DM, is known to promote the
progression of hepatic steatosis (3). Of note, insulin activity to suppress
triglyceride hydrolysis in adipose tissue is diminished in IR,
eventually causing fatty acid accumulation in the liver (4). Moreover, enhanced hepatic lipid
production is induced by IR. Hyperglycemia induces the
transactivation of transcriptional factors, carbohydrate responsive
element binding protein and Liver X receptor, which are known to
activate de novo hepatic lipid synthesis (5,6).
Furthermore, triglyceride hydrolysis in the liver is also modified
by IR. Hepatic mRNA expression levels of lipolytic enzymes,
hormone-sensitive lipase and adipose tissue triglyceride lipase
were found to be suppressed in NAFLD with IR (7). These observations suggest that IR may
adversely affect multiple mechanisms regulating hepatic lipid
contents and promote the progression of NAFLD.
Triglyceride release as VLDL requires coordinated
functions of apolipoprotein B (apoB) and microsomal triglyceride
transfer protein (MTP) (8). ApoB
is an essential protein required for assembly and secretion of VLDL
from the liver and chylomicron from the small intestine (9). MTP transfers free cholesterol,
phospholipids, triglyceride and cholesterol esters to apoB during
translation, allowing apoB to attain a pre-VLDL conformation, which
supports the subsequent fusion of apoB with MTP-stabilized
triglyceride droplets and the formation of mature VLDL (10). Genetic analyses confirm the crucial
roles of MTP in lipid metabolism; liver-specific Mttp-knockout
induced striking reductions in VLDL triglyceride and caused hepatic
steatosis (11). In addition,
liver fatty-acid binding protein (L-FABP) is known to facilitate
intracellular trafficking of long-chain fatty acids (LCFAs)
(12). This property of L-FABP
modulates diverse cellular functions, including fatty acid uptake
and intracellular lipid contents and metabolism.
To understand the effects of IR on the mechanism of
triglyceride release from the liver, we conducted this study to
estimate hepatic mRNA expression levels of apoB, MTP and L-FABP in
patients with NAFLD using RT-PCR. We examined the effects of
metabolic factors, including IR, and the extent of obesity and
hepatic lipid accumulation on the expression of the genes. We found
that the expression levels of all genes examined in the NAFLD liver
were significantly increased compared to those of the healthy
controls. Furthermore, IR or advanced hepatic steatosis
significantly reduced the expression levels of MTP in the NAFLD
liver. These observations may be helpful to understand the effects
of IR on the progression of NAFLD.
Materials and methods
Patients and samples
Liver tissue samples were obtained by biopsy from 22
patients with histologically diagnosed NAFLD who were admitted to
the Kyushu University Hospital between 2004 and 2006. To avoid the
effects of fibrosis on metabolic parameters, patients
histologically diagnosed with NASH were not included. Liver tissue
samples were obtained from living donors during liver
transplantation and used as healthy controls. Written consent was
obtained from all of the patients. Characteristics of the enrolled
subjects, including gender, age, body mass index (BMI), serum
levels of aspartate aminotransferase (AST), alanine
aminotransferase (ALT), γ-glutamyl transpeptidase (γGTP), lactate
dehydrogenase (LDH), total cholesterol, triglyceride, fasting
plasma glucose, C-reactive protein (CRP), platelet count and
prothrombin time, were documented.
RT-PCR
Total RNA was prepared with TRIzol reagent
(Invitrogen, Carlsbad, CA, USA), and cDNA was synthesized with
GeneAmp™ RNA PCR (Applied Biosystems. Branchburg, NJ, USA).
Real-time PCR was performed using LightCycler-FastStart DNA Master
SYBR Green 1 (Roche, Basel, Switzerland) according to the
manufacturer’s instructions. To control for variations in the
reactions, all PCR data were normalized against β-actin expression.
PCR primers for apoB were forward 5′-GCCGAGTTTGCCTTGCTCA-3′ and
reverse 5′-TCCGGAGGCTCACCAGTTTC-3′; the primers for MTP were
forward 5′-GGCTCCTGCCTACAGCTTCT-3′ and reverse
5′-CAGCCAGTGGATCACCACA-3′; the primers for L-FABP were forward
5′-CAGCTGCGGGATGAGATTGA-3′ and reverse
5′-AAACGCTGGTGTGTGATGGGTA-3′.
Statistical analyses
The results were expressed as the means ± standard
deviation. Significant differences between two groups were assessed
using unpaired two-tailed t-tests. A value of p<0.05 was
considered significant.
Results
Subject characteristics
We studied 22 NAFLD patients (12 males and 10
females) and 10 control subjects (6 males and 4 females) (Table I). Since control samples were
obtained from healthy donors for transplantation, the age of this
group was younger compared to the NAFLD group, but the difference
was not significant. BMI was significantly higher in the NAFLD
group than in the control group. AST, ALT and γGTP were higher in
the NAFLD group than in the control group, but the differences were
not significant. Nutritional parameters, including total
cholesterol, triglyceride and fasting plasma glucose, were higher
in the NAFLD group than in the control group, but the differences
were not statistically significant.
 | Table I.Clinical characteristics of the
patients with NAFLD and control subjects. |
Table I.
Clinical characteristics of the
patients with NAFLD and control subjects.
| Control (n=10) | NAFLD (n=22) | p-value |
|---|
| Male/female | 6/4 | 12/10 | 0.77000 |
| Age (years) | 32.6±7.06 | 38.0±14.1 | 0.06600 |
| BMI
(kg/m2) | 21.3±2.58 | 25.3±6.94 | 0.00261a |
| AST (U/l) | 15.0±3.35 | 39.9±47.1 | 0.15100 |
| ALT (U/l) | 12.3±4.09 | 54.2±69.6 | 0.10300 |
| γGTP (U/l) | 17.9±3.18 | 61.3±82.0 | 0.12900 |
| LDH (U/l) | 138±27.6 | 217±83.2 | 0.01450a |
| Total cholesterol
(mg/dl) | 164±26.7 | 191±45.1 | 0.12600 |
| Triglyceride
(mg/dl) | 77.0±22.0 | 130±112 | 0.20200 |
| Fasting plasma
glucose (mg/dl) | 87.6±4.90 | 108±32.2 | 0.09690 |
| Platelets
(×104/mm3) | 24.2±4.39 | 21.6±3.77 | 0.13600 |
| Prothrombin time
(%) | 88.8±6.65 | 94.3±9.32 | 0.13700 |
| CRP (mg/dl) | 0.062±0.13 | 0.081±0.1 | 0.69000 |
Up-regulation of genes involved in VLDL
assembly and fatty acid trafficking in the NAFLD liver
mRNA expression levels of apoB and MTP were
significantly higher, 3-fold and 2-fold, respectively, in the NAFLD
than in the control liver (Fig. 1A and
B). L-FABP was also significantly higher in the NAFLD liver
than in the control (Fig. 1C).
Since all three genes were up-regulated in the NAFLD liver in a
similar manner, we evaluated the correlations among the expression
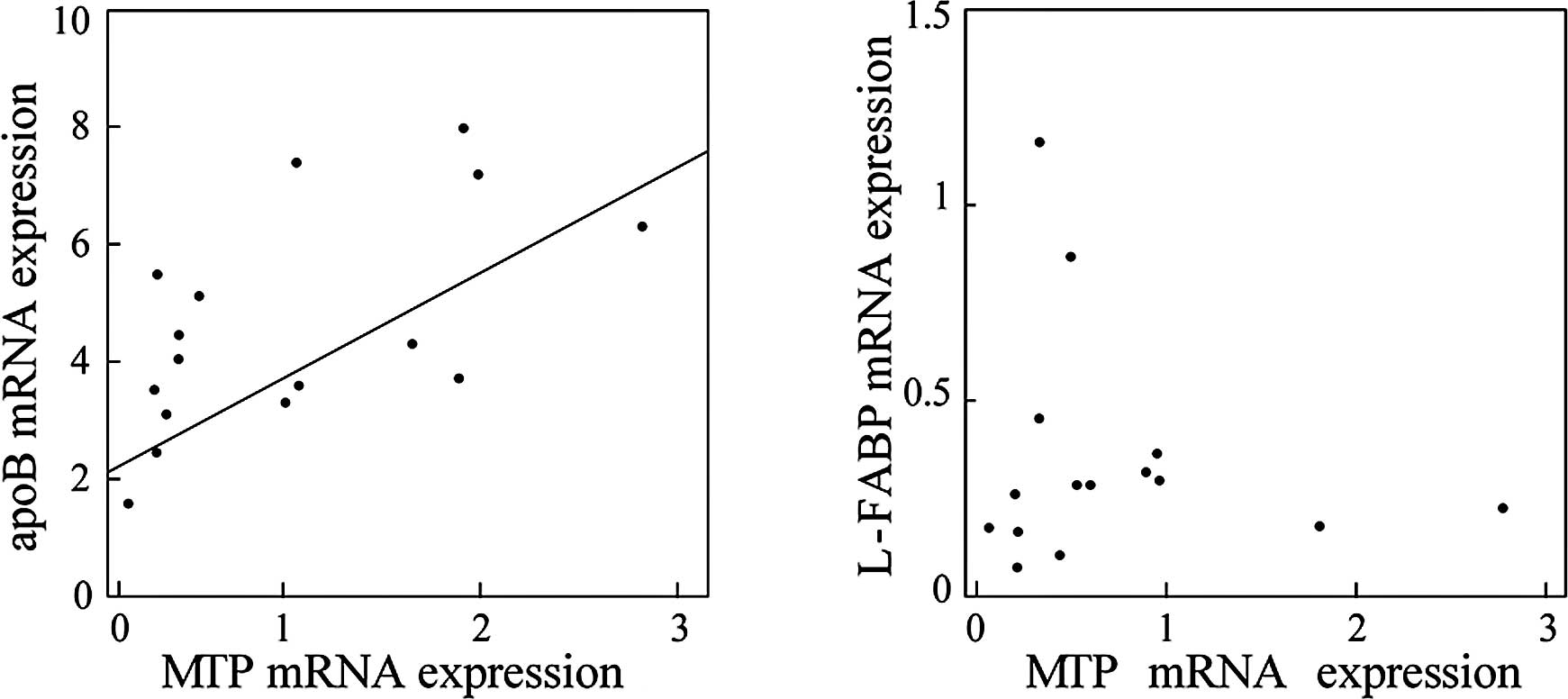
levels of these genes. As shown in Fig. 2, the expression levels of MTP were
significantly correlated with those of apoB (r=0.614, p=0.0006;
Fig. 2A), but not with those of
L-FABP (Fig. 2B), suggesting that
a common mechanism may be involved in the transcriptional
regulation of MTP and apoB in the NAFLD liver. Moreover, enhanced
expression of these genes suggests that a compensatory mechanism to
release excess lipids is activated in the NAFLD liver.
Effects of IR on the expression levels of
apoB, MTP and L-FABP in the NAFLD liver
We analyzed whether IR modifies the expression
levels of these genes in NAFLD. In 22 patients with NAFLD, 12 had
homeostasis model assessment of insulin resistance (HOMA-IR) values
>2.5 and were considered to have IR. Expression levels of all of
the genes measured were reduced in patients with IR. Of note, the
expression levels of MTP in patients with HOMA-IR >2.5 were
significantly reduced compared to those with HOMA-IR <2.5
(Fig. 3). To evaluate the effects
of obesity, we compared the expression levels of genes between
patients with BMI <30 kg/m2 (n=9) and those with BMI
>30 kg/m2 (n=13). However, significant differences
were not observed (Fig. 4).
Effects of the extent of lipid
accumulation on the expression levels of apoB, MTP and L-FABP in
the NAFLD liver
The numbers of lipid-laden hepatocytes in the NAFLD
liver were histologically counted on biopsy sections. mRNA
expression levels of genes were then compared between liver samples
with mild steatosis (<30% of hepatocytes showed marked lipid
accumulation; n=12) and advanced steatosis (>30% of hepatocytes
showed marked lipid accumulation; n=10). The expression levels of
MTP in livers with advanced steatosis were significantly reduced
compared to those with mild steatosis (Fig. 5). A similar tendency was observed
in the expression levels of apoB, but the difference was not
significant. These results suggest that the reduced expression of
MTP in the liver with advanced steatosis is involved in the
mechanism to exacerbate hepatic lipid accumulation in the NAFLD
liver, through the reduction of lipid excretion as VLDL.
Discussion
In this study, the hepatic mRNA expression levels of
apoB, MTP and L-FABP were increased in the NAFLD liver compared to
those in the control livers, suggesting that a compensatory
mechanism to release excess lipid as VLDL is promoted in the
steatotic liver. However, in patients with IR or advanced
steatosis, this compensatory mechanism seems to be disrupted via
the down-regulation of MTP. This process to attenuate the
compensatory mechanism may be involved in the progression of
NAFLD.
Transcriptional regulations of genes examined in
this study are known to be mediated by transcriptional factors,
including hepatocyte nuclear factor (HNF)4α and peroxisome
proliferator-activated receptor (PPAR) α. HNF4α controls the
expression of genes involved in lipoprotein and lipid metabolism,
including MTP and apoB (13–15).
HNF4α binds long chain fatty acyl-CoA and this binding stimulates
its transcriptional activity (16,17).
PPARα, which regulates the expression of a number of genes critical
for lipid and lipoprotein metabolism, is known to transactivate MTP
expression in rat liver and cultured hepatocytes (18,19).
PPARα is also responsible for the induction of L-FABP in
hepatocytes in response to HMG-CoA reductase inhibitor (20). Since our observations showed a
positive correlation between the expression levels of apoB and MTP,
but not between those of L-FABP and MTP, HNF4α may be more likely
to be responsible for the simultaneous and correlated
transactivation of apoB and MTP.
In this study, the expression levels of MTP were
reduced in NAFLD with IR; however, this observation is not
consistent with the known effects of insulin on the transcriptional
regulation of MTP. Hepatic expression of MTP is reported to be
down-regulated by insulin (21).
In detail, MTP expression is induced by transcriptional factor
forkhead box O1 (FoxO1), whose activation is down-regulated by
exposure to insulin (10,22). In response to insulin, FoxO1 is
phosphorylated by activated Akt, resulting in a reduction in
nuclear FoxO1 and time-dependent decreases in MTP transcription.
These findings suggest that MTP expression would not be
down-regulated in IR. Currently, we cannot explain why the opposite
results were observed in this study. Small sample size may be the
reason for this unexpected MTP expression. However, we hypothesize
that deteriorated cellular circumstances in the NAFLD liver may be
responsible. Of note, oxidative stress, which is induced by
mitochondrial dysfunction in steatotic cells, has a strong
potential to damage various cellular functions, including
homeostasis of endoplasmic reaction, control of cell death and
transcriptional regulation of various genes (23–26).
Furthermore, hepatic iron load in the NAFLD liver would be a factor
to enhance cellular damages. Since iron has a strong potential to
produce oxidative stress, hepatic iron load is known to promote the
progression of NASH (27,28). In the NAFLD liver, reduction in MTP
expression levels and increased iron score were simultaneously
observed during the progression of liver disease (29). Taken together, reduced MTP
expression in NAFLD liver with IR or advanced steatosis may be
induced by enhanced oxidative stress in these cellular
circumstances.
In this study, the expression of genes involved in
VLDL assembly and fatty acid trafficking was enhanced in the liver
with NAFLD. However, the reduced expression of MTP in livers with
IR or advanced steatosis may be factors to accelerate further
steatosis. Further studies are necessary to determine the effects
of IR on the progression of NAFLD.
References
|
1
|
Angulo P: Nonalcoholic fatty liver
disease. N Engl J Med. 346:1221–1231. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fabbrini E, Sullivan S and Klein S:
Obesity and nonalcoholic fatty liver disease: biochemical,
metabolic, and clinical implications. Hepatology. 51:679–689. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tiniakos DG, Vos MB and Brunt EM:
Nonalcoholic fatty liver disease: pathology and pathogenesis. Annu
Rev Pathol. 5:145–171. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jaworski K, Sarkadi-Nagy E, Duncan RE,
Ahmadian M and Sul HS: Regulation of triglyceride metabolism. IV.
Hormonal regulation of lipolysis in adipose tissue. Am J Physiol
Gastrointest Liver Physiol. 293:G1–G4. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Iizuka K and Horikawa Y: ChREBP: a
glucose-activated transcription factor involved in the development
of metabolic syndrome. Endocr J. 55:617–624. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Higuchi N, Kato M, Shundo Y, Tajiri H,
Tanaka M, Yamashita N, Kohjima M, Kotoh K, Nakamuta M, Takayanagi R
and Enjoji M: Liver X receptor in cooperation with SREBP-1c is a
major lipid synthesis regulator in nonalcoholic fatty liver
disease. Hepatol Res. 38:1122–1129. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kato M, Higuchi N and Enjoji M: Reduced
hepatic expression of adipose tissue triglyceride lipase and CGI-58
may contribute to the development of non-alcoholic fatty liver
disease in patients with insulin resistance. Scand J Gastroenterol.
43:1018–1019. 2008. View Article : Google Scholar
|
|
8
|
Meshkani R and Adeli K: Hepatic insulin
resistance, metabolic syndrome and cardiovascular disease. Clin
Biochem. 42:1331–1346. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shelness GS and Sellers JA:
Very-low-density lipoprotein assembly and secretion. Curr Opin
Lipidol. 12:151–157. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sparks JD and Sparks CE: Overindulgence
and metabolic syndrome: is FoxO1 a missing link? J Clin Invest.
118:2012–2015. 2008.PubMed/NCBI
|
|
11
|
Raabe M, Veniant MM, Sullivan MA, Zlot CH,
Bjorkegren J, Nielsen LB, Wong JS, Hamilton RL and Young SG:
Analysis of the role of microsomal triglyceride transfer protein in
the liver of tissue-specific knockout mice. J Clin Invest.
103:1287–1298. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Binas B and Erol E: FABPs as determinants
of myocellular and hepatic fuel metabolism. Mol Cell Biochem.
299:75–84. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sladek FM: Orphan receptor HNF-4 and
liver-specific gene expression. Receptor. 3:223–232.
1993.PubMed/NCBI
|
|
14
|
Sheena V, Hertz R, Nousbeck J, Berman I,
Magenheim J and Bar-Tana J: Transcriptional regulation of human
microsomal triglyceride transfer protein by hepatocyte nuclear
factor-4alpha. J Lipid Res. 46:328–341. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ladias JA, Hadzopoulou-Cladaras M,
Kardassis D, Cardot P, Cheng J, Zannis V and Cladaras C:
Transcriptional regulation of human apolipoprotein genes ApoB,
ApoCIII, and ApoAII by members of the steroid hormone receptor
superfamily HNF-4, ARP-1, EAR-2, and EAR-3. J Biol Chem.
267:15849–15860. 1992.
|
|
16
|
Hertz R, Magenheim J, Berman I and
Bar-Tana J: Fatty acyl-CoA thioesters are ligands of hepatic
nuclear factor-4alpha. Nature. 392:512–516. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jump DB: Dietary polyunsaturated fatty
acids and regulation of gene transcription. Curr Opin Lipidol.
13:155–164. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yoon M: The role of PPARalpha in lipid
metabolism and obesity: focusing on the effects of estrogen on
PPARalpha actions. Pharmacol Res. 60:151–159. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ameen C, Edvardsson U, Ljungberg A, Asp L,
Akerblad P, Tuneld A, Olofsson SO, Linden D and Oscarsson J:
Activation of peroxisome proliferator-activated receptor alpha
increases the expression and activity of microsomal triglyceride
transfer protein in the liver. J Biol Chem. 280:1224–1229. 2005.
View Article : Google Scholar
|
|
20
|
Landrier JF, Thomas C, Grober J, Duez H,
Percevault F, Souidi M, Linard C, Staels B and Besnard P: Statin
induction of liver fatty acid-binding protein (L-FABP) gene
expression is peroxisome proliferator-activated
receptor-alpha-dependent. J Biol Chem. 279:45512–45518. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Au WS, Kung HF and Lin MC: Regulation of
microsomal triglyceride transfer protein gene by insulin in HepG2
cells: roles of MAPKerk and MAPKp38. Diabetes. 52:1073–1080. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kamagate A, Qu S, Perdomo G, Su D, Kim DH,
Slusher S, Meseck M and Dong HH: FoxO1 mediates insulin-dependent
regulation of hepatic VLDL production in mice. J Clin Invest.
118:2347–2364. 2008.PubMed/NCBI
|
|
23
|
Wei Y, Wang D, Topczewski F and
Pagliassotti MJ: Saturated fatty acids induce endoplasmic reticulum
stress and apoptosis independently of ceramide in liver cells. Am J
Physiol Endocrinol Metab. 291:E275–E281. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Roebuck KA: Oxidant stress regulation of
IL-8 and ICAM-1 gene expression: differential activation and
binding of the transcription factors AP-1 and NF-κB (Review). Int J
Mol Med. 4:223–230. 1999.PubMed/NCBI
|
|
25
|
Gius D, Botero A, Shah S and Curry HA:
Intracellular oxidation/reduction status in the regulation of
transcription factors NF-kappaB and AP-1. Toxicol Lett. 106:93–106.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu H, Colavitti R, Rovira II and Finkel
T: Redox-dependent transcriptional regulation. Circ Res.
97:967–974. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
George DK, Goldwurm S, MacDonald GA,
Cowley LL, Walker NI, Ward PJ, Jazwinska EC and Powell LW:
Increased hepatic iron concentration in nonalcoholic
steatohepatitis is associated with increased fibrosis.
Gastroenterology. 114:311–318. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mantena SK, King AL, Andringa KK,
Eccleston HB and Bailey SM: Mitochondrial dysfunction and oxidative
stress in the pathogenesis of alcohol- and obesity-induced fatty
liver diseases. Free Radic Biol Med. 44:1259–1272. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mitsuyoshi H, Yasui K, Harano Y, Endo M,
Tsuji K, Minami M, Itoh Y, Okanoue T and Yoshikawa T: Analysis of
hepatic genes involved in the metabolism of fatty acids and iron in
nonalcoholic fatty liver disease. Hepatol Res. 39:366–373. 2009.
View Article : Google Scholar : PubMed/NCBI
|