Introduction
Type 2 diabetes mellitus is a metabolic syndrome
with a complex pathogenesis, which is currently exhibiting an
increasing morbidity rate each year. However, there is no effective
therapeutic drug (1). Type 2
diabetes mellitus is predominantly caused by abnormal sugar and
lipid metabolism, due to the relative deficiency of insulin
(insufficient insulin secretion and insulin resistance), or
excessive glucagon levels. Clinically, patients present with
hyperglycemia and hyperlipemia (2). While having a large impact on patient
quality of life, type 2 diabetes mellitus also becomes a burden for
the development of a modern economic society. The American Diabetes
Association and the European Association for the Study of Diabetes
proposed that the identification of a glycosylated hemoglobin level
of >7% among patients may be used as an indication of undesired
blood glucose control (3). The
existing oral antidiabetics demonstrate severe side-effects, and
treatment with insulin increases the risk of weight gain and
hypoglycemia. Therefore, the development of novel and effective
methods for the treatment of type 2 diabetes mellitus, or the
identification of novel small molecular drugs, is urgently required
(4). Peroxisome
proliferator-activated receptor-γ (PPARγ) is a member of the
nuclear receptor super family, which plays an important role in
regulating glucolipid metabolism (5). PPARγ is a ligand-dependent nuclear
receptor (6). Following the
integration of ligand binding and activation, the receptor forms a
heterodimer with the retinoid X receptor-α (RXRα), and regulates
the expression levels of associated genes on the peroxisome
proliferator response element (PPRE) (7).
Among the clinical drugs used in the treatment of
type 2 diabetes mellitus, thiazolidinediones, such as rosiglitazone
(Ros) and pioglitazone, can be used as PPARγ agonists to
significantly reduce hypoglycemia and improve the sensitivity to
the insulin pathway. However, these drugs have strong side-effects
concurrently, including weight gain or increased risks of
cardiovascular diseases, which constrain their use in clinical
practice (8). However, in recent
years, studies have demonstrated that a PPARγ gene knockout or
intermediate depression of PPARγ activity (9) can relieve the insulin resistance
induced by a high fat diet (10).
This treatment method also avoids the side-effects that are
observed with a PPARγ full agonist (11). Therefore, the screening of small
molecular compounds based on PPARγ antagonists is a key strategy
for the identification of novel compounds for the treatment of type
2 diabetes mellitus (12).
Currently, there are a limited number of studies investigating
PPARγ antagonists. Therefore, the development of novel drugs for
the treatment of type 2 diabetes mellitus based on PPARγ
antagonists is of great importance (13).
In the present study, transcriptional activation and
a mammalian one-hybrid system were applied to screen for the PPARγ
antagonist N-((1H-benzo[d]imidazol-2-yl)methyl) aniline, referred
to as ‘Compound Q’. Furthermore, the effects of this antagonist on
the expression levels of PPARγ-regulated lipid metabolism genes and
the differentiation of adipocytes were studied in order to develop
novel reagents with the potential to ameliorate obesity and type 2
diabetes.
Materials and methods
Cell culture of the 293T and 3T3-L1
preadipocyte cell lines
A 293T cell line and 3T3-L1 preadipocyte cell line
(American Type Culture Collection, Manassas, VA, USA) were cultured
in Dulbecco’s modified Eagle’s medium (DMEM; Gibco Life
Technologies, Grand Island, NY, USA), containing 10% fetal bovine
serum (Gibco Life Technologies) and 1% penicillin-streptomycin, at
37°C in saturated humidity with 5% CO2. The culture
solution of the cells was exchanged on a daily basis and
trypsinization was processed every two days. Cells at a logarithmic
growth phase were seeded for the experiment.
Luciferase activity test
Effects of Compound Q (J&K Chemical Co., Ltd.,
Shanghai, China) on the coactivator recruitment of PPARγ and the
transcriptional activity of the RXRα-PPARγ heterodimer were
analyzed. At a logarithmic phase of cell growth, the 293T cells
were seeded in a 24-well cell culture plate. When the cells reached
a density of 50–70%, the medium was replaced with serum-free DMEM.
Plasmids were transfected into the cells using a Calcium Phosphate
Transfection kit (Nanjing Jiancheng Bioengineering Institute,
Nanjing, China). To assess the ability of the compound for PPARγ
coactivator recruitment, PPARγ-LBD, UAS-TK-Luc reporter and an
internal reference plasmid, pRL-SV40 (all from Promega Corporation,
Madison, WI, USA), were transfected in the 293T cells. When
assaying the effects of the compound on the transcriptional
activity of the RXRα-PPARγ heterodimer, the full-length plasmids of
RXRα and PPARγ, PPRE-Luc and the internal reference plasmid,
pRL-SV40, were transfected into the 293T cells. After 6 h of
transfection, the medium was replaced with complete medium
containing 10% fetal bovine serum. In addition, Compound Q was
added to the medium for 18 h of culturing. The medium from the
culture dish was removed and the plates were washed with
phosphate-buffered saline (PBS). To each well, 100 μl cell lysis
buffer was added and incubated at 37°C for 20 min to enable cell
lysis. Next, with reference to the instructions in the luciferase
kit (Dual-Luciferase® Reporter Assay system; Promega
Corporation), the activity of the firefly luciferase and internal
reference, fluorescein, was analyzed.
3T3-L1 adipocyte differentiation
experiment
A 3T3-L1 preadipocyte differentiation assay was
conducted according to the reported classic procedure (14). Briefly, 3T3-L1 preadipocytes at a
logarithmic growth phase were seeded into six-well cell culture
plates. The cell density was allowed to reach 100%, and this time
point was set as day 0. On day 4, the complete medium containing
0.115 mg/l methylisobutylxanthine (MIX), 0.39 mg/l dexamethasone
(DEX) and 1 mg/l insulin was cultured for three days. The solution
was further stimulated through the removal of old medium and the
addition of fresh medium with 1 mg/l insulin, which was cultured
for an additional three days. Following these stimulations, the
differentiated adipocytes were cultured in normal medium (DMEM
containing 10% FBS) for 1–2 days prior to carrying out further
experiments.
To observe the effects of Compound Q on adipocyte
differentiation, a negative control [dimethyl sulfoxide (DMSO)],
positive control (Ros) and Compound Q at various concentrations (1,
10 and 20 μM) were added to the differentiation medium. After
adipocyte stimulation for 6 days, the differentiation medium was
removed and the cells were washed three times with PBS. The
solution was stained using a Oil Red O kit (Nanjing Jiancheng
Bioengineering Institute) according to manufacturer’s protocol, to
investigate the lipid accumulation in adipocytes. Images were
photographed for observation using a BX 50 microscope (Olympus
Corporation, Tokyo, Japan) (15).
Quantitative polymerase chain reaction
(PCR)
Mature 3T3-L1 adipocytes that had undergone
differentiation were cultured in six-well plates and processed with
different compounds, including a negative control (DMSO), positive
control (Ros) and Compound Q at various concentrations (1, 10 and
20 μM), that were added to the complete medium for 24 h. Total RNA
was extracted using TRIzol reagent (Invitrogen Life Technologies,
Carlsbad, CA, USA), and reversed transcribed to cDNA using a
PrimeScriptTM RT kit (Takara Biotechnology, Co., Ltd.,
Dalian, China). SYBR Green Real Time PCR Master Mix (Toyobo Co.,
Ltd., Tokyo, Japan) was used for PCR, which was performed in a DNA
Engine Opticon 2 device (Bio-Rad Laboratories, Inc., Hercules, CA,
USA). The primers used in the experiment were as follows: GAPDH
forward, 5′-GTATGACTCCACTCACGGCAAA-3′ and reverse,
5′-GGTCTCGCTCCTGGAAGATG-3′; fatty acid synthase (FAS) forward,
5′-CTGAGATCCCAGCACTTCTTGA-3′ and reverse,
5′-GCCTCCGAAGCCAAATGAG-3′; CCAAT/enhancer-binding protein-α
(C/EBPα) forward, 5′-CTGAGATCCCAGCACTTCTTGA-3′ and reverse,
5′-CACGGCTCAGCTGTTCCA-3′; adipocyte fatty acid binding protein 2
(aP2) forward, 5′-CATGGCCAAGCC CAACAT-3′ and reverse,
5′-CGCCCAGTTTGAAGG AAATC-3′; HMG-CoA forward, 5′-CATGCAGATTCT
GGCAGTCAGT-3′ and reverse, 5′-CGGCTTCACAAACCA CAGTCT-3′. These
genes were selected as they are PPARγ-regulated and the main genes
involved in lipid metabolism. All the samples were assayed
according to the manufacturer’s instructions.
Transcriptional activation and mammalian
one-hybrid system
A mammalian one-hybrid system was carried out to
screen compounds which were able to directly bind and influence the
recruitment of co-activators to PPARγ-LBD. In the mammalian
one-hybrid system, the Gal4/UAS system was used to investigate the
activity regulation of PPARγ, whether the co-transfection of
Gal4-PPARγ-LBD and UAS-TK-Luc into 293T cells was able to express
the Gal4-PPARγ-LBD protein, and whether the potential compound was
able to activate or inhibit PPARγ-LBD and its ability to influence
the expression of TK-Luc. A transcriptional activation system was
also carried out to investigate the transcription activity of
PPARγ. As the co-transfection of pGL3-PPRE-Luc and pcDNA3.1-PPARγ
into 293T cells results in PPARγ expression, whether the potential
compound was able to activate or inhibit PPARγ activity, and
increase or decrease the transcription of PPRE-Luc was
investigated. In the mammalian one-hybrid and transcriptional
activation systems, SV40 was used as the control for transfection
efficiency.
Statistical analysis
Data are shown as mean ± standard error of the mean.
A student’s t-test was performed for the comparison of two groups
and one-way analysis of variance was carried out for the comparison
of >2 groups by GraphPad Prism 5 software (GraphPad, San Diego,
CA, USA). P<0.05 was considered to indicate a statistically
significant difference.
Results
Compound Q as a PPARγ antagonist
As shown in Fig. 1,
a mammalian one-hybrid method using a luciferase-reporter system
was used for the experiment. Compound Q (Fig. 1A) was shown to depress the
coactivator recruitment of PPARγ in a concentration-dependent
manner (Fig 1B). In addition, with
regard to transcriptional activation, Compound Q was revealed to
reverse the transcriptional activity of the Ros-activated
RXRα-PPARγ heterodimer in a concentration-dependent manner
(Fig. 1C). The observations
indicated that as a PPARγ antagonist, Compound Q is able to reduced
the transcriptional activity of the RXRα-PPARγ heterodimer.
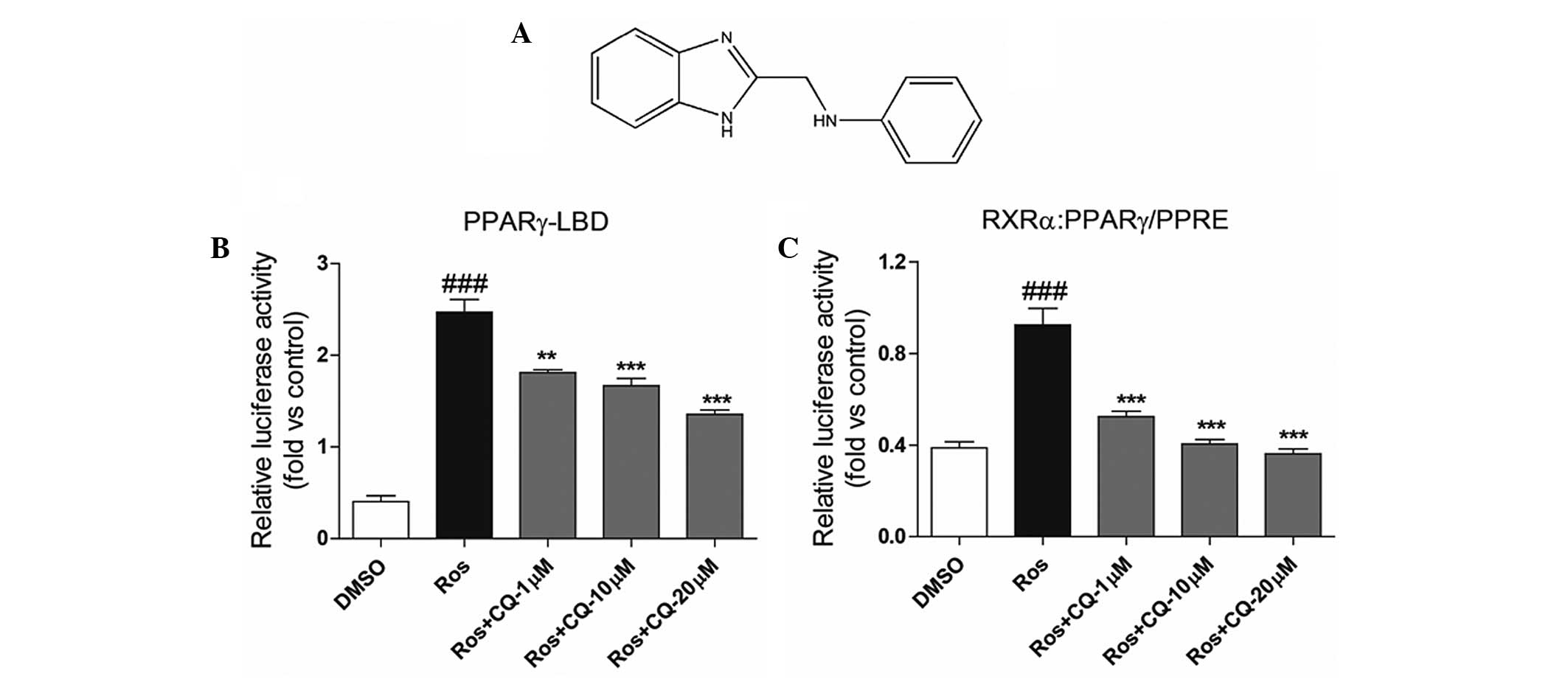 | Figure 1(A) Structural formula of Compound Q.
Compound Q was shown to depress the activity of Ros-activated (B)
PPARγ-LBD-Luc and (C) RXRα:PPARγ-PPRE-Luc in a
concentration-dependent manner. Ros, a known PPARγ agonist, is able
to activate PPARγ-LBD and increase the transcription of PPRE-Luc.
As compared with Ros treatment, the combination of Compund Q and
Ros decreased Ros-induced luciferase activity in the mammal
one-hybrid and transcriptional activation systems, suggesting that
Compund Q is a PPARγ antagonist and can inhibit the transcriptional
activity of PPARγ. ###P<0.001 compared to DMSO group;
**P<0.01 and ***P<0.001, respectively,
compared to Ros group. Ros, rosiglitazone; DMSO, dimethyl
sulfoxide; PPARγ, peroxisome proliferator-activated receptor-γ;
Luc, luciferase; PPRE, peroxisome proliferator response element;
RXRα, retinoid X receptor-α; CQ, Compound Q. |
Compound Q depresses 3T3-L1 preadipocyte
differentiation in a concentration-dependent manner
As PPARγ is a key moderator of lipid metabolism and
a vital gene involved in adipocyte differentiation, a previous
study reported that PPARγ agonists, such as Ros, are able to
significantly promote adipocyte differentiation (14). Thus, the present study further
investigated the effects of Compound Q on adipocyte
differentiation.
Compound Q at various concentrations, or the
negative or positive control, were added to the differentiation
medium. At the end of the differentiation assay, staining with Oil
Red O demonstrated that the positive control, Ros, was able to
significantly enhance adipocyte differentiation, while Compound Q
decreased the formation of grease drops in adipocytes and the
formation of adipocytes in a concentration-dependent manner
(Fig. 2). This indicates that
Compound Q has the potential to inhibit triglyceride accumulation
in adipose tissue and ameliorate obesity in obese patients.
Compound Q decreases the expression
levels of genes associated with lipid metabolism
Adipocyte differentiation is a complex process
involving the regulation of multiple genes. In order to study the
mechanism underlying the Compound Q-induced depression of adipocyte
differentiation, quantitative PCR was used to analyze the effects
of Compound Q on the expression levels of key genes involved in
lipid metabolism. As shown in Fig.
3, following the culture of adipocytes for 24 h with Compound Q
at various concentrations, the positive control (Ros) or the
negative control (DMSO), the total RNA of the different cells was
extracted. Following quantitative PCR, Ros was shown to enhance the
expression levels of FAS, C/EBPα, aP2 and HMG-CoA. By contrast,
treatment with Compound Q decreased the expression levels of the
genes associated with lipid metabolism in a concentration-dependent
manner.
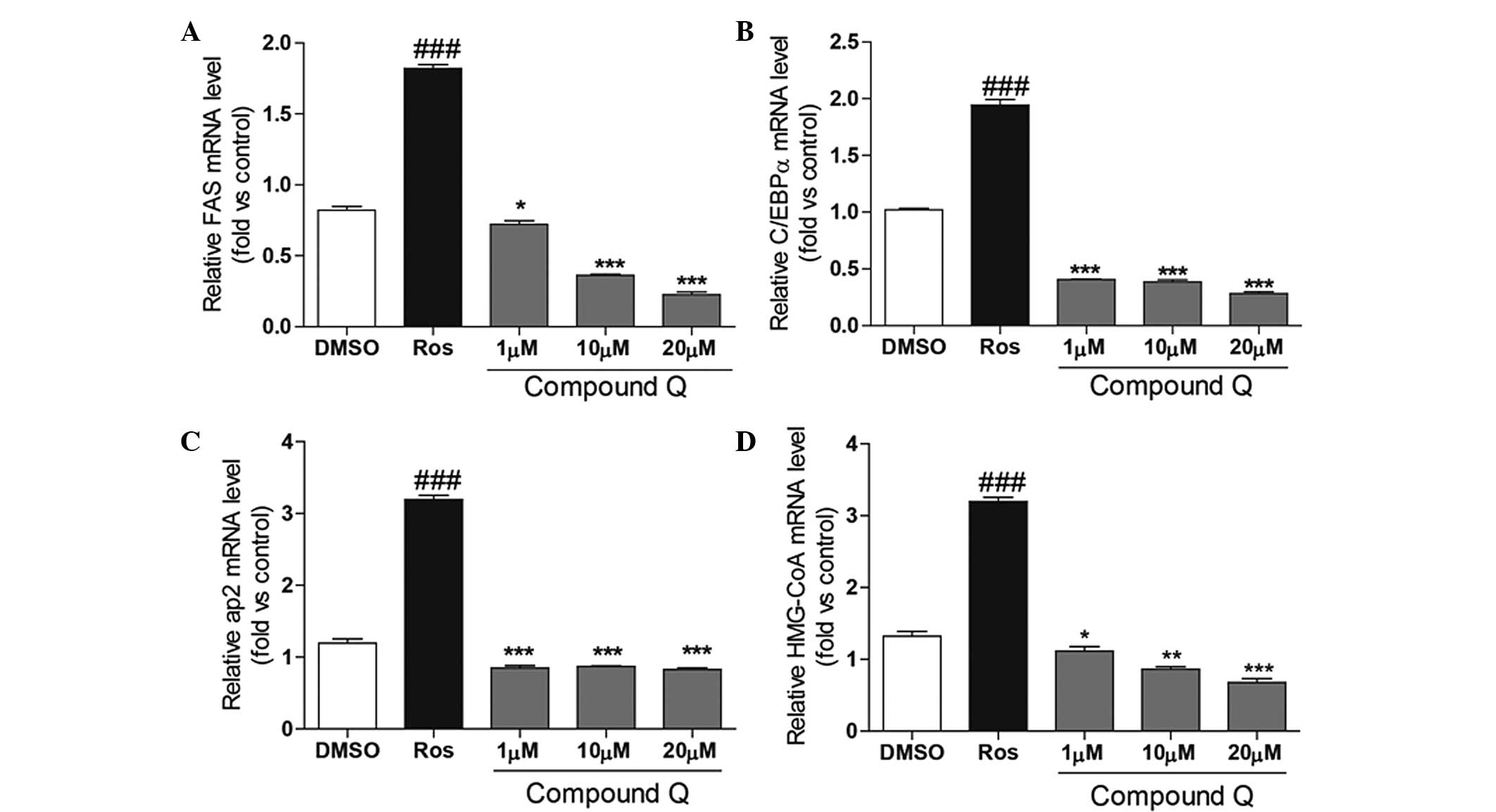 | Figure 3Differentiated mature adipocytes were
cultured for 24 h in a medium containing Compound Q at various
concentrations, a positive control (Ros) or a negative control
(DMSO). Compound Q was shown to decrease the expression levels of
(A) FAS, (B) C/EBPα, (C) aP2 and (D) HMG-CoA in a
concentration-dependent manner, while Ros was found to increase the
expression levels. ###P<0.001, *P<0.05,
**P<0.01 and ***P<0.001. Ros,
rosiglitazone; DMSO, dimethyl sulfoxide; FAS, fatty acid synthase;
C/EBPα, CCAAT/enhancer-binding protein-α; aP2, adipocyte fatty acid
binding protein 2. |
Discussion
Type 2 diabetes mellitus has become a common disease
with a high morbidity rate in modern society. Patients mainly
present with hyperglycemia and hyperlipemia, as well as a range of
complications induced by hyperglycemia in the later stages, such as
diabetic nephropathy and diabetic foot (16). Hyperglycemia is primarily caused by
the failure of pancreatic β cells to sufficiently secrete insulin
and compensate for the insulin resistance of tissues. This results
in reduced glucose absorption and increased lipidolysis (17). Hyperglycemia may also be the result
of excessive glucagon secretion by pancreatic α cells, which leads
to an increase in gluconeogenesis. The condition ultimately results
in abnormal glucolipid metabolism of the organism (18). Clinically, oral antidiabetics have
diverse side-effects, and treatment with insulin has revealed risks
of weight gain and hypoglycemia (19,20).
Currently, oral antidiabetics, including Ros and
pioglitazone of the thiazolidinedione class of medications, perform
hypoglycemic effects and increase insulin sensitivity by activating
the nuclear receptor, PPARγ (21).
However, these drugs have severe side-effects, including weight
gain and increased risks of cardiovascular diseases (22). In recent years, research has found
that decreasing the activity of PPARγ by constructing PPARγ gene
knockout models or inducing mutations at PPARγ activity sites can
relieve the insulin resistance induced by a high fat diet (23).
Based on the aforementioned observations, the
present study adopted a mammalian one-hybrid method with
transcriptional activation to identify that Compound Q, as a PPARγ
antagonist, is able to reduce PPARγ coactivator recruitment and
reverse Ros-activated RXRα-PPARγ transcriptional activity in a
concentration-dependent manner. In the cell activity assay,
Compound Q was found to reduce the formation of grease drops in
adipocytes and the formation of adipocytes in a
concentration-dependent manner. With further investigation into the
underlying mechanism, Compound Q was revealed to decrease the rate
of differentiation by decreasing the expression levels of key genes
involved in lipid metabolism. Therefore, the effects of Compound Q
in depressing adipocyte differentiation and regulating the
expression levels of genes associated with lipid metabolism
indicate the potential of the compound for weight loss and lipid
metabolism regulation therapy.
In conclusion, the present study investigated a
PPARγ antagonist, and the results provide a new understanding and
research basis for the future investigation into novel small
molecular compounds with fewer side-effects for the treatment of
type 2 diabetes mellitus. Compound Q is a small molecular compound
that was demonstrated to be a significant active compound. However,
the biological activity of Compound Q in vivo requires
further evaluation.
Acknowledgements
The study was supported by a grant from the Natural
Science Foundation of Shanghai Science Technology Commission (nos.
12ZR1425600; 14411965200 and 14411970900).
References
|
1
|
Samuel VT and Shulman GI: Mechanisms for
insulin resistance: common threads and missing links. Cell.
148:852–871. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kahn SE, Hull RL and Utzschneider KM:
Mechanisms linking obesity to insulin resistance and type 2
diabetes. Nature. 444:840–846. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kumar PR, Bhansali A, Ravikiran M, et al:
Utility of glycated hemoglobin in diagnosing type 2 diabetes
mellitus: a community-based study. J Clin Endocrinol Metab.
95:2832–2835. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Halperin F and Goldfine AB: Metabolic
surgery for type 2 diabetes: efficacy and risks. Curr Opin
Endocrinol Diabetes Obes. 20:98–105. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Houseknecht KL, Cole BM and Steele PJ:
Peroxisome proliferator-activated receptor gamma (PPAR gamma) and
its ligands: a review. Domest Anim Endocrinol. 22:1–23. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bajaj M, Suraamornkul S, Hardies LJ, Glass
L, Musi N and DeFronzo RA: Effects of peroxisome
proliferator-activated receptor (PPAR)-alpha and PPAR-gamma
agonists on glucose and lipid metabolism in patients with type 2
diabetes mellitus. Diabetologia. 50:1723–1731. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sharma AM and Staels B: Review: Peroxisome
proliferator-activated receptor gamma and adipose tissue -
understanding obesity-related changes in regulation of lipid and
glucose metabolism. J Clin Endocrinol Metab. 92:386–395. 2007.
View Article : Google Scholar
|
|
8
|
Shearer BG and Billin AN: The next
generation of PPAR drugs: do we have the tools to find them?
Biochim Biophys Acta. 1771:1082–1093. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Clement K, Hercberg S, Passinge B, et al:
The Pro115Gln and Pro12Ala PPAR gamma gene mutations in obesity and
type 2 diabetes. Int J Obes Relat Metab Disord. 24:391–393. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kadowaki T: PPAR gamma agonist and
antagonist. Nihon Yakurigaku Zasshi. 118:321–326. 2001.(In
Japanese). View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zinman B: PPAR gamma agonists in type 2
diabetes: how far have we come in ‘preventing the inevitable’? A
review of the metabolic effects of rosiglitazone. Diabetes Obes
Metab. 3(Suppl 1): S34–S43. 2001. View Article : Google Scholar
|
|
12
|
Wojtowicz AK, Szychowski KA and Kajta M:
PPAR-γ agonist GW1929 but not antagonist GW9662 reduces
TBBPA-induced neurotoxicity in primary neocortical cells. Neurotox
Res. 25:311–322. 2014. View Article : Google Scholar
|
|
13
|
de Boer RA, Martens FM, Kuipers I, Boomsma
F and Visseren FL: The effects of the PPAR-gamma agonist
pioglitazone on plasma concentrations of circulating vasoactive
factors in type II diabetes mellitus. J Hum Hypertens. 24:74–76.
2010. View Article : Google Scholar
|
|
14
|
Chawla A, Schwarz EJ, Dimaculangan DD and
Lazar MA: Peroxisome proliferator-activated receptor (PPAR) gamma:
adipose-predominant expression and induction early in adipocyte
differentiation. Endocrinology. 135:798–800. 1994.PubMed/NCBI
|
|
15
|
Huang C, Zhang Y, Gong Z, Sheng X, Li Z,
Zhang W and Qin Y: Berberine inhibits 3T3-L1 adipocyte
differentiation through the PPARgamma pathway. Biochem Biophys Res
Commun. 348:571–578. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sheu W, Jeng CY, Fuh M, Chen YD and Reaven
GM: Resistance to insulin-mediated glucose disposal in patients
with noninsulin-dependent diabetes mellitus in the absence of
obesity or microalbuminuria - a Clinical Research Center study. J
Clin Endocrinol Metab. 81:1156–1159. 1996.PubMed/NCBI
|
|
17
|
Chatterjee PK: Hepatic inflammation and
insulin resistance in pre-diabetes - further evidence for the
beneficial actions of PPAR-gamma agonists and a role for SOCS-3
modulation. Br J Pharmacol. 160:1889–1891. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Khan A, Safdar M, Ali Khan MM, Khattak KN
and Anderson RA: Cinnamon improves glucose and lipids of people
with type 2 diabetes. Diabetes Care. 26:3215–3218. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bøg-Hansen E, Lindblad U, Rånstam J,
Melander A and Rastam L: Impaired glucose metabolism and obesity in
Swedish patients with borderline isolated systolic hypertension:
Skaraborg Hypertension and Diabetes Project. Diabetes Obes Metab.
3:25–31. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hanas R and Ludvigsson J: Side effects and
indwelling times of subcutaneous catheters for insulin injections:
a new device for injecting insulin with a minimum of pain in the
treatment of insulin-dependent diabetes mellitus. Diabetes Res Clin
Pract. 10:73–83. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chiang CK, Ho TI, Peng YS, et al:
Rosiglitazone in diabetes control in hemodialysis patients with and
without viral hepatitis infection: effectiveness and side effects.
Diabetes Care. 30:3–7. 2007. View Article : Google Scholar
|
|
22
|
Zhang F, Lavan BE and Gregoire FM:
Selective modulators of PPAR-gamma activity: Molecular aspects
related to obesity and side-effects. PPAR Res. 2007:326962007.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Evans RM, Barish GD and Wang YX: PPARs and
the complex journey to obesity. Nat Med. 10:355–361. 2004.
View Article : Google Scholar : PubMed/NCBI
|















