Introduction
Osteoblasts and osteoclasts maintain normal bone
metabolism balance, and are involved in bone metabolism and the
immune activity of the body (1).
Receptor activator of nuclear factor-κB ligand (RANKL) expression,
which can be found on the surface of osteoblasts, is activated by
stimulation with endocrine hormones and local cytokines, and can be
combined with RANK receptors on the surface of osteoclasts to
induce osteoclast proliferation and activation (2).
At present, the known mechanism of osteoclast
formation is the RANK-RANKL-osteoprotegerin (OPG) shaft (3–5).
RANK lies on the surface of osteoclast precursors. Osteoblasts and
bone marrow stromal cells express RANKL to promote osteoclast
differentiation, enhance the viability of mature osteoclasts and
prevent osteoclast apoptosis. Certain cells, including bone marrow
stromal cells and osteoblasts, express OPG, which inhibits the
differentiation and maturation of osteoclasts to decrease bone
resorption activity and induce apoptosis. Regulators, including
serum osteoprotegerin, serve a function in the homeostasis of bone
resorption and reconstruction, and the whole process is regulated
by osteoblasts (6,7).
Bones and joints are the most commonly affected
organs in extrapulmonary tuberculosis. The use of anti-tuberculosis
drugs has allowed the control of tuberculosis in the past (8); however, due to human immunodeficiency
virus infection/acquired immune deficiency syndrome, drugs,
immunosuppressants, alcoholism and poverty, the incidence of
tuberculosis is once more on the increase, particularly in
developing countries (9). Bone
absorption and vertebral body structure damage are pathological
changes caused by bone tuberculosis. Pott’s disease (spinal
tuberculosis) can cause bone destruction, spinal cord compression
and nerve dysfunction. In vitro, recombinant
Mycobacterium tuberculosis (r-Mt) 10-kDa co-chaperonin
(cpn10) causes accelerated bone resorption and fragility (10–12),
increases osteoclast proliferation and activity, and stimulates
bone resorption, thus inhibiting osteoblast proliferation (13); however, whether the effect of r-Mt
cpn10 in stimulating bone resorption is mediated by any receptors
expressed in osteoblasts remains unclear. Furthermore, it is not
well understood how r-Mt cpn10 affects bone tissue at the molecular
level. In the present study, the effect of r-Mt cpn10 on the
expression levels of OPG and RANKL in third cultured osteoblasts
was investigated.
Materials and methods
Reagents
Cell culture media, penicillin (100 U/ml),
streptomycin (100 U/ml), L-glutamine, phosphate-buffered saline,
trypsin-EDTA, super fetal bovine serum and r-Mt cpn10 were
purchased from Shanghai Biological Engineering Co., Ltd. (batch no.
120628; Shanghai, China). Freeze-dried r-Mt cpn10 protein was
stored under nitrogen flow and dissolved in double-distilled water
prior to use.
Cell culture
The isolation and culture of the third-generation
osteoblasts was performed according to the methods used in a
previous study (14). The original
osteoblast-like cultures were isolated from bone fragments taken
from a total of 12 male patients undergoing surgery, none of which
had metabolic bone disease. Written informed consent was obtained
from all patients prior to the study. Briefly, the trabecular bone
was cut into sections (3–4-mm), thoroughly rinsed and vortexed five
times in phosphate-buffered saline at 1,200 × g for 15 min with a
Low-Speed Desktop Centrifuge (Lu Xiangyi Centrifuge Instrument Co.,
Ltd., Shanghai, China). Cells were incubated in a humidified
CO2 incubator at 37°C and the medium was changed twice a
week until confluence was achieved. Cells were cultured in
Dulbecco’s Modified Eagle’s Medium supplemented with 10% super
fetal bovine serum, L-glutamine (2 mM) and antibiotics. The
osteoblastic phenotype of the cells in the third generation
osteoblast-like cultures was verified by biochemical markers as
previously described (14). Prior
to stimulation with r-Mt cpn10, cells were incubated in serum-free
medium for 24 h. r-Mt cpn10 was added into fresh serum-free medium,
reaching final concentrations of 0.01–10 μg/ml. The controls were
incubated with fresh serum-free medium alone. For the measurement
of OPG secretion, 500,000 third-generation human osteoblasts were
seeded in triplicate into 25-cm2 dishes. After 24 h,
aliquots of the medium were collected and the number of cells in
each well was manually counted. For RNA isolation, cells were
seeded in 25-cm2 flasks at a density of 150,000
cells/flask. Cultures were harvested 2, 4, 8 and 24 h after
stimulation with r-Mt cpn10. For protein isolation, cells were
seeded in 25-cm2 flasks at a density of 600,000
cells/flask. Cultures were harvested 2, 4, 8 and 24 h after
stimulation with r-Mt cpn10. The present study was approved by the
Ethics Committee of the People’s Hospital of Xinjiang Autonomous
Region (registration no. 2012084; Urumqi, China).
Enzyme-linked immunosorbent assay
(ELISA)
The levels of OPG were determined using ELISA as
described previously (15).
Briefly, a MaxiSorp™ microtiter plate (Nalge Nunc International,
Penfield, NY, USA) was coated with mouse anti-human OPG monoclonal
antibody (cat. no. ab2147; dilution, 1:50; Abcam, Cambridge, MA,
USA). Samples or standard protein recombinant human OPG were added
and incubated for 2 h at room temperature. The bound proteins were
detected with biotinylated goat anti-human OPG antibody (batch no.
40849; Shanghai YongYe Biotechnology Co., Ltd., Shanghai, China).
Following development, the plate was read at 450 nm using a
microplate reader (MF Benchmark Plus reader; Bio-Rad, Hercules, CA,
USA). The detection limit was 35 pg/ml, and the intra- and
inter-assay variations were 6.2 and 21%, respectively. Protein
concentrations were normalized to the number of cells and expressed
as pg/ml/106 cells.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
RT-qPCR analysis was performed in triplicate for the
determination of the levels of RANKL and OPG in the
third-generation osteoblasts. To isolate the total RNA,
TRIzol® (Invitrogen Life Technologies, Carlsbad, CA,
USA) and SYBR® Select Master Mix (Applied
Biosystems®; Invitrogen Life Technologies) were used
according to the manufacturers’ instructions. RNA quality was
analyzed using a 2100 series Bioanalyzer Instrument (Agilent
Technologies, Santa Clara, CA, USA). RNA concentration and purity
were measured by determining the 260/280 nm ratio. All ratios were
>1.8. RNA (1 μg/sample) were reverse-transcribed to first-strand
cDNA with a RevertAid™ First Strand cDNA Synthesis kit (Thermo
Fisher Scientific, Inc., Waltham, MA, USA) and used in RT-qPCR
analysis in an ABI® 7500 Real-Time PCR system (Applied
Biosystems; Invitrogen Life Technologies). All RNA samples from the
same experiment were transcribed at the same time and each cDNA was
analyzed in duplicate. Fluorescent fluorescein amidite-labeled
probes and gene-specific primers spanning over the exon-intron
boundary for RANKL and OPG were used (Shanghai Biological
Engineering Co., Ltd.). A
4,5-dichloro-2,7-dimethoxy-fluorescein-labeled reference gene,
GAPDH, was used as an endogenous control. Standard curves for
various genes were achieved through a
1×10−2-1×10−8 dilution series of a verified
PCR product with a concentration of 10 ng/μl. The amplification and
analysis of cDNA fragments were performed on a thermocycler
(iCycler; Bio-Rad, Munich, Germany). The primers used in this
section were as follows: GAPDH forward, 5′-TGTTGCCATCAATGACCCCTT-3′
and reverse, 3′-GCGACTCATGCAGCACCTC-5; OPG forward,
5′-CCTCTGTGAAAACAGCGTGC-3′ and reverse, 3′-TTTACCGCTGGTTCTGTGGA-5′;
RANKL forward, 5′-GGAGTTGGCCGCAGACAAGA-3′ and reverse, 3′-TCG
CAGCGGGACAAGAAGAT-5′.
Western blotting
To determine the levels of OPG, RANKL and GAPDH,
equal quantities of proteins (40 μg) were separated by 10% sodium
dodecyl sulfate-polyacrylamide gel electrophoresis and blotted with
polyvinylidene fluoride membranes (Millipore, Billerica, MA, USA).
Following blocking with 5% skimmed milk and washing with 1X
Tris-buffered saline with Tween-20 (20 mM Tris-HCL, pH 7.5; 150 mM
NaCl; 0.1% Tween-20; all purchased from Sigma, St. Louis, MO, USA),
the membranes were probed with rabbit polyclonal anti-RANKL
(dilution, 1:800; cat. no. bs-0747R; Bioss, Beijing, China), rabbit
polyclonal anti-OPG (dilution, 1:800; cat. no. bs-0431R; Bioss) at
room temperature for 1 h and then incubated with anti-mouse or
anti-rabbit immunoglobulin G conjugated with horseradish peroxidase
(Zhongshan Jinqiao Biological Technology Co., Ltd., Beijing,
China.). Following final treatment with Amersham™ ECL™ Western
Blotting Detection reagents (Amersham; GE Healthcare, Little
Chalfont, UK), the samples were exposed to X-ray film and relevant
protein bands were recorded.
Statistical analysis
Statistical analysis was performed using SPSS 17.0
software (SPSS, Inc., Chicago, IL, USA). The mRNA levels are
expressed as mean arbitrary units ± intra-assay variation of
duplicate analyses. Each experiment was repeated for the assessment
of inter-assay variation and differed by <10%. For protein
production measured with ELISA and western blotting, the
statistical significance was calculated from three separate
experiments and evaluated by the Student’s t-test. P<0.05 was
considered to indicate a statistically significant difference.
Results
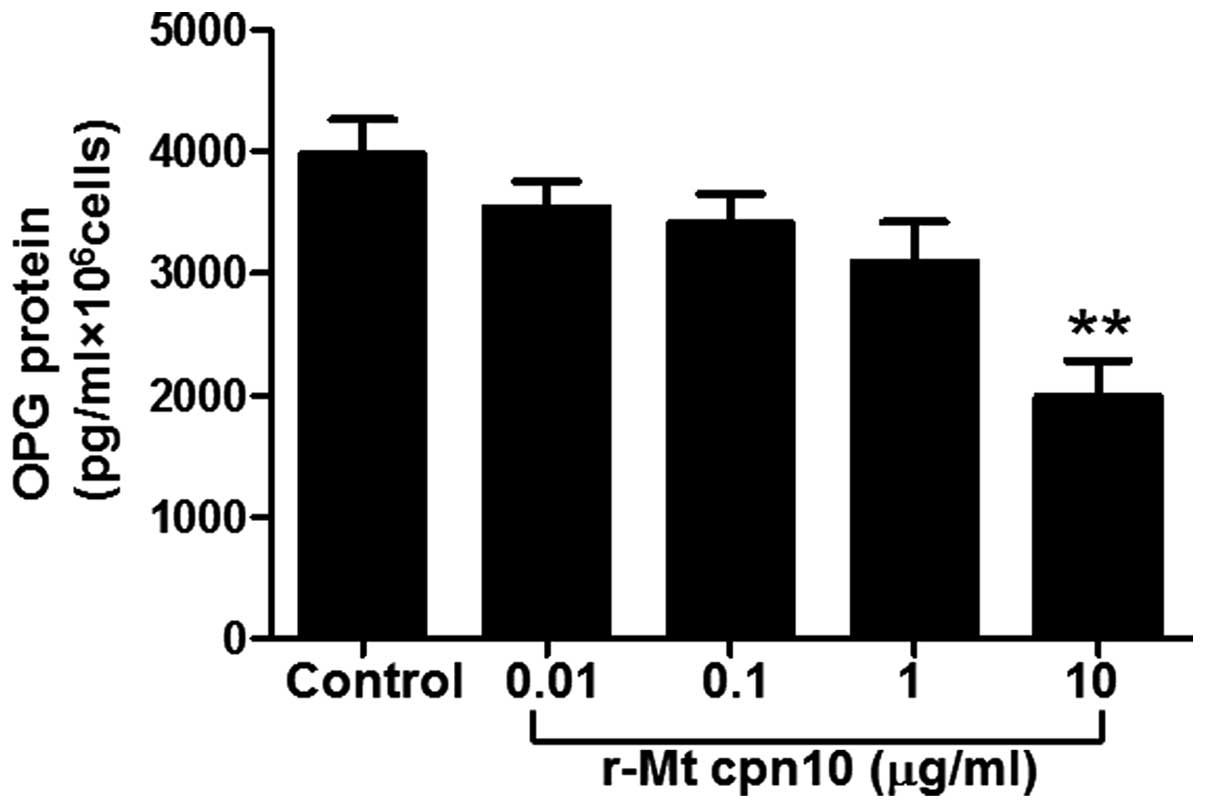
r-Mt cpn10 downregulates OPG protein
secretion
To assess the effect of r-Mt cpn10 on OPG protein
secretion, the third-generation cultured osteoblasts were treated
with r-Mt cpn10 at doses ranging from 0.01 to 10 μg/ml in
serum-free media. The results revealed that lower doses (0.01, 0.1
and 1 μg/ml) of r-Mt cpn10 failed to significantly decrease OPG
protein secretion; however, 10 μg/ml r-Mt cpn10 significantly
decreased OPG protein secretion by 50% compared with the control
(P<0.01) (Fig. 1). This result
suggested that r-Mt cpn10 downregulated OPG protein secretion at 10
μg/ml.
r-Mt cpn10 inhibits OPG mRNA and protein
expression in a time-dependent manner
RT-qPCR and western blotting were performed to
investigate the time-course effect of r-Mt cpn10 on the levels of
OPG. The RT-qPCR results for the third-generation cultured
osteoblasts demonstrated that r-Mt cpn10 (10 μg/ml) treatment
decreased OPG mRNA expression to 27% of that of the control after 4
h, and to 14.5% of that of the control after 8 h; however, it
increased OPG mRNA expression to 200% of that of the control after
24 h (Fig. 2A). Western blot
analysis revealed a similar effect of r-Mt cpn10 (10 μg/ml) on OPG
protein expression, with the maximal inhibitory effect occurring
after 8 h (Fig. 2B and C). These
results indicated that r-Mt cpn10 inhibited OPG mRNA and protein
expression in a time-dependent manner.
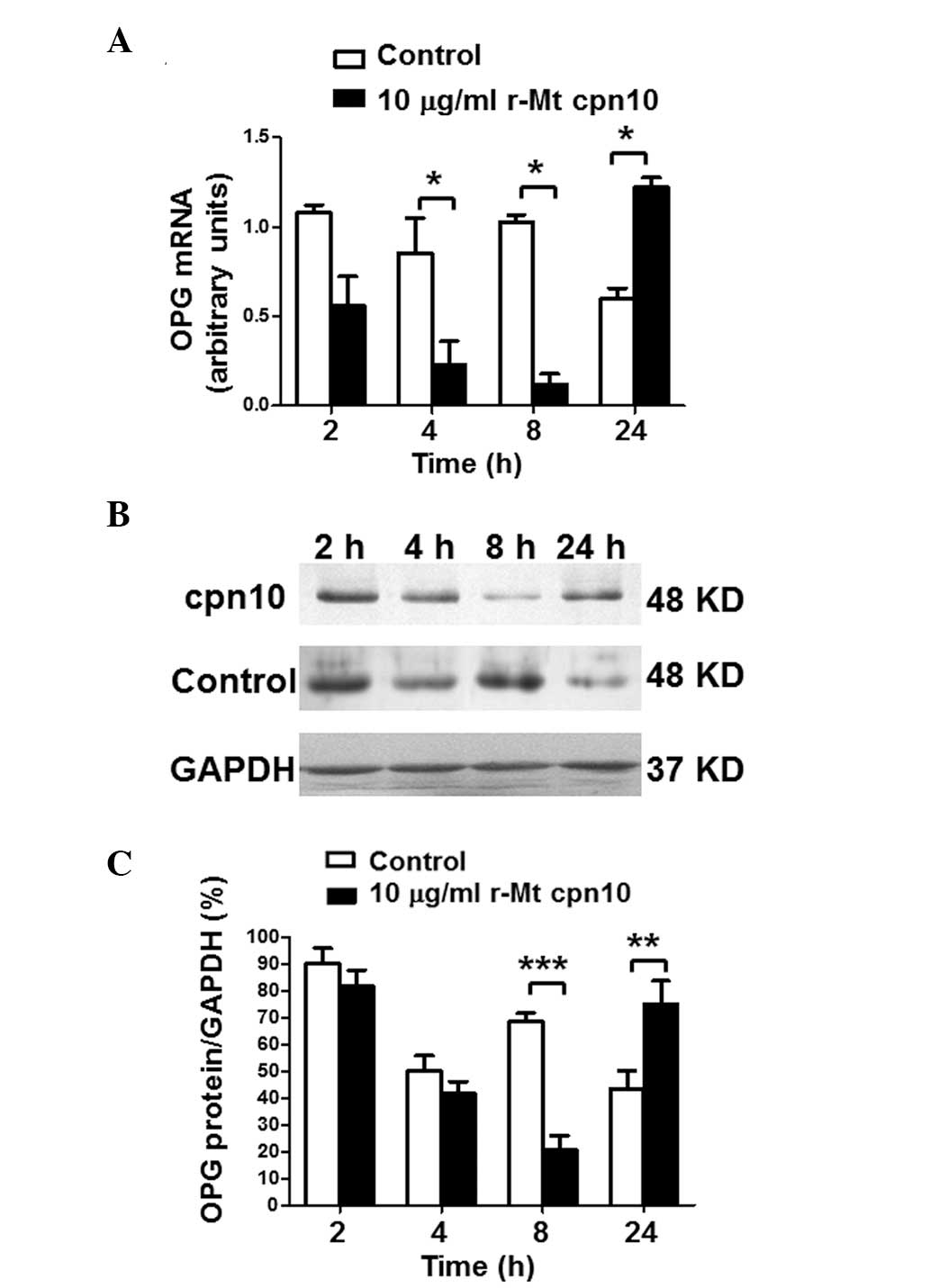 | Figure 2Levels of OPG in third-generation
osteoblasts following treatment with r-Mt cpn10 (10 μg/ml) for 2,
4, 8 and 24 h. (A) Levels of OPG mRNA following stimulation with
control or r-Mt cpn10 (10 μg/ml) for 2, 4, 8 and 24 h. Total RNA
was isolated from the harvested cells, transcribed to cDNA and
quantified by reverse transcription-quantitative polymerase chain
reaction analysis. The amount of OPG mRNA was compared with the
amount of the reference gene (GAPDH) mRNA. Values are expressed as
the mean ± intra-assay variation of duplicate analyses. (B) Western
blotting of OPG protein expression following treatment with the
control or r-Mt cpn10 (10 μg/ml) for 2, 4, 8 and 24 h. (C)
Quantification of the level of OPG protein following treatment with
the control or r-Mt cpn10 (10 μg/ml) for 2, 4, 8 and 24 h. Results
are expressed as the mean ± standard deviation.
*P<0.05, **P<0.01 and
***P<0.001. r-Mt, recombinant Mycobacterium
tuberculosis; cpn10, 10-kDa co-chaperonin; OPG,
osteoprotegerin. |
r-Mt cpn10 inhibits OPG mRNA and protein
expression in a dose-dependent manner
RT-qPCR and western blotting were performed to
investigate the effect of different doses of r-Mt cpn10 on the
levels of OPG. Data from the RT-qPCR revealed that the level of OPG
mRNA decreased following treatment with r-Mt cpn10 (0.01, 0.1, 1
and 10 μg/ml) for 8 h. The higher the dose of r-Mt cpn10, the lower
the OPG mRNA level (Fig. 3A). The
level of OPG mRNA was the lowest (9.9% of that of the control) when
the concentration of r-Mt cpn10 was increased to 10 μg/ml (Fig. 3A). Western blot analysis
demonstrated a similar trend for the level of OPG protein following
treatment with r-Mt cpn10 (0.01, 0.1, 1 and 10 μg/ml) for 8 h, with
the level of OPG protein at its lowest when the concentration of
r-Mt cpn10 was increased to 10 μg/ml (Fig. 3B and C). These results demonstrated
that r-Mt cpn10 inhibited OPG mRNA and protein expression in a
dose-dependent manner.
r-Mt cpn10 upregulates the levels of
RANKL mRNA and protein in a time-dependent manner
RT-qPCR and western blotting were performed to
investigate the time-course effect of r-Mt cpn10 on the level of
RANKL. The RT-qPCR results for the third-generation cultured
osteoblasts demonstrated that RANKL mRNA expression was induced
following treatment with r-Mt cpn10 (10 μg/ml), with a maximal
effect of a 2.6-fold induction compared with the control observed
after 8 h (Fig. 4A). Similarly,
western blot analysis revealed that treatment with r-Mt cpn10 (10
μg/ml) for 8 h increased the level of RANKL protein by 100%
compared with the control (Fig. 4B and
C). These results suggested that r-Mt cpn10 upregulated the
levels of RANKL mRNA and protein in a time-dependent manner.
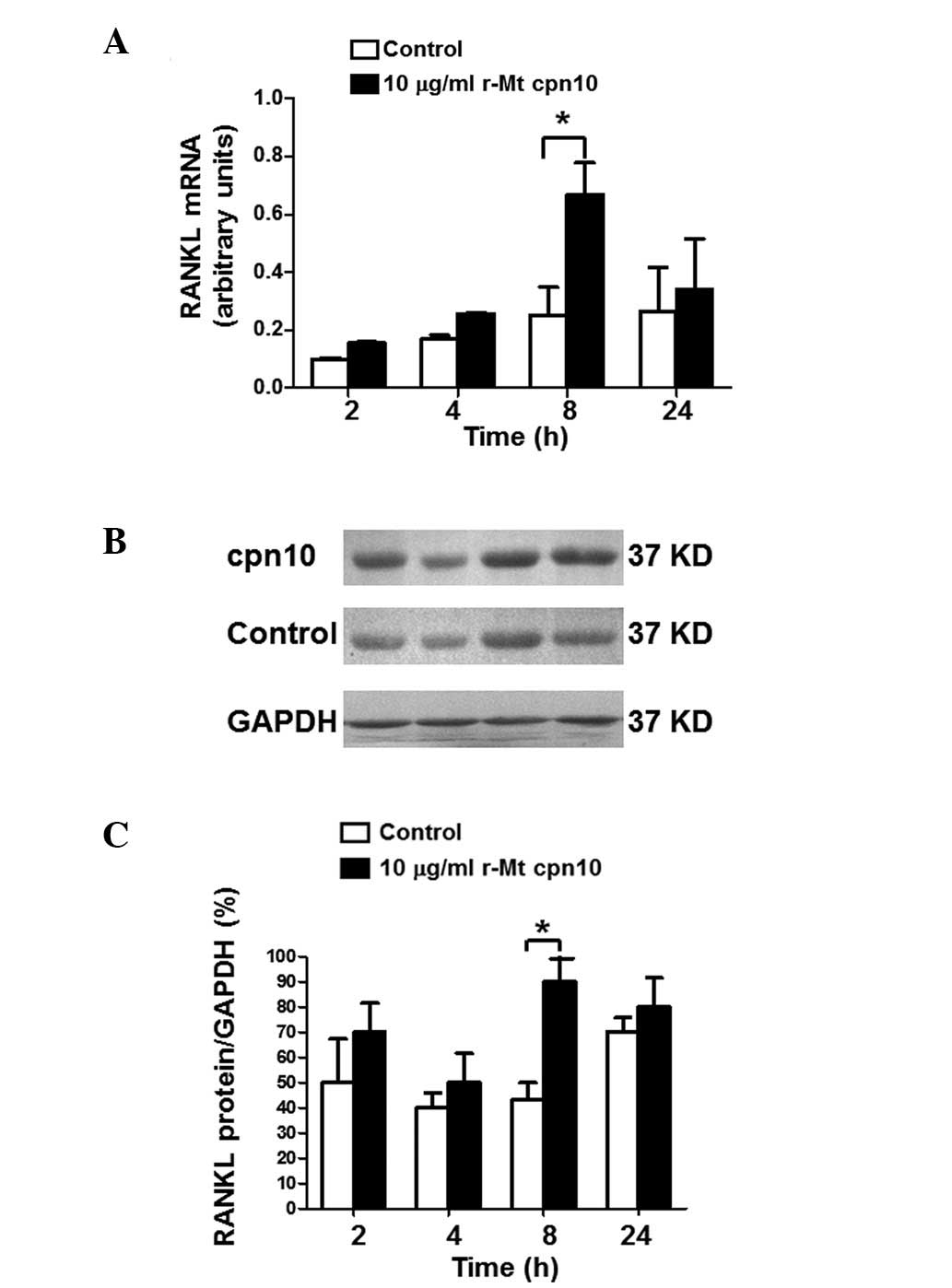 | Figure 4Levels of RANKL in third-generation
osteoblasts following treatment with r-Mt cpn10 (10 μg/ml) for 2,
4, 8 and 24 h. (A) Levels of RANKL mRNA following stimulation with
control or r-Mt cpn10 (10 μg/ml) for 2, 4, 8 and 24 h. Total RNA
was isolated from the harvested cells, transcribed to cDNA and
quantified by reverse transcription-quantitative polymerase chain
reaction analysis. The amount of RANKL mRNA was compared with the
amount of the reference gene (GAPDH) mRNA. Values are expressed as
the mean ± intra-assay variation of duplicate analyses. (B) Western
blot analysis of RANKL protein expression following treatment with
the control or r-Mt cpn10 (10 μg/ml) for 2, 4, 8 and 24 h. (C)
Quantification of the level of RANKL protein following treatment
with the control or r-Mt cpn10 (10 μg/ml) for 2, 4, 8 and 24 h.
Results are expressed as the mean ± standard devation.
*P<0.05. RANKL, receptor activator of nuclear
factor-κB ligand; r-Mt, recombinant Mycobacterium
tuberculosis; cpn10, 10-kDa co-chaperonin. |
Treatment with r-Mt cpn10 increases the
RANKL/OPG ratio
To evaluate the net effect of RANKL and OPG in the
in vitro system, the RANKL/OPG ratio in the third-generation
cultured osteoblasts was calculated following treatment with r-Mt
cpn10 (10 μg/ml). The results revealed that the RANKL/OPG mRNA
ratio was nine-fold higher than that in the control cells. However,
after 24 h, the ratio returned almost to its initial value
(Fig. 5). This calculation
indicated that treatment with r-Mt cpn10 (10 μg/ml) for 8 h had the
strongest effect in increasing the RANKL/OPG ratio.
Discussion
The present study demonstrated that OPG was
downregulated and RANKL was upregulated in osteoblasts by r-Mt
cpn10. It is well understood that OPG and RANKL are important
regulators of osteoclastogenesis, and RANKL has been found to be
involved in the differentiation and development of osteoclasts
(16). In a previous study,
RANKL-knockout (RANKL −/−) mice exhibited extensive bone sclerosis
and a lack of mature osteoclasts; this was alleviated following
RANKL treatment. Furthermore, the application of recombinant RANKL
led to severe osteoporosis and hypercalcemia in mice (17). Transgenic OPG mice (+/+) exhibited
bone sclerosis, while OPG-knockout mice (OPG −/−) showed
progressive aggravation of osteoporosis, with a reduction in the
cortical and trabecular bone volume, thickness and number of cells
(18). In vitro studies
showed that 10 ng/ml recombinant OPG completely inhibited the
generation of osteoblasts and osteoclasts in the co-culture system.
The addition of RANKL formed fully functional osteoclasts, which
were completely inhibited by the addition of OPG (19,20).
Thus, the differential regulation of the two proteins may be an
important mechanism by which r-Mt cpn10 induces bone
resorption.
The relative ratio of RANKL/OPG in the bone marrow
microenvironment is the principal factor that defines the effect of
cytokines on osteoclast formation and activation. Notably, it has
been shown that the serum RANKL/OPG ratio has prognostic
significance in multiple myeloma (21). A higher risk of osteolytic lesions
and mortality is associated with an increased RANKL/OPG ratio. In
previous studies, an increased RANKL/OPG ratio has been observed in
patients with severe osteolysis (22–25).
In the present study, the levels of RANKL and OPG in osteoblasts
were antagonistically affected by r-Mt cpn10, leading to a markedly
increased RANKL/OPG ratio. This observation supports our hypothesis
that the RANK-RANKL-OPG system mediates r-Mt cpn10-induced bone
resorption.
The resorptive effect of r-Mt Cpn10 is partly caused
by its direct effect on bone tissue (10). Regions of cpn10 responsible for its
osteolytic and osteoblast-antiproliferative activities have been
identified to exist in the loop spanning residues 65–70 and the
mobile loop of the protein. It has been previously speculated that
the most likely signaling pathway occurred via cpn10 receptors
(26), with the sequence SGLVIPDT
of cpn10 directly binding to RANK. In the present study, the
effects of r-Mt cpn10 on OPG and RANKL expression were dose- and
time-dependent; however, whether the changes in the levels of mRNA
resulted from the regulation of transcription or changes in message
stability remains unknown. A reporter assay using RANKL and OPG
upstream regulatory sequences is required to confirm the
speculation that these are transcriptional effects mediated by
cpn10.
In the current study, the effects of r-Mt cpn10 in
inducing bone resorption were similar to those reported previously
(27–30). The maximal r-Mt cpn10-dependent
downregulation of OPG mRNA expression was observed after 4–8 h and
that of OPG protein expression after 8 h. The r-Mt cpn10-dependent
downregulation of OPG was the most significant at a concentration
of 10 μg/ml. The maximal r-Mt cpn10-dependent upregulation of RANKL
mRNA and protein expression was observed with 10 μg/ml r-Mt cpn10
after 8 h. Meghji et al (13) reported that the addition of Mt to
murine calvarial bone produced a dose-dependent stimulation of bone
resorption, which was measured from calcium release into the tissue
culture medium. The number of osteoclasts in the calvarial explants
was counted and the results showed a parallel increase. In the
present study, an r-Mt cpn10-dependent downregulation of OPG
expression and upregulation of RANKL expression was observed in the
presence of 10 μg/ml r-Mt cpn10. The relative ratio of RANKL/OPG
was highest at 8 h, and it indicated osteoclast formation and
activation.
During the development process, in which the
expression levels of OPG and RANKL are regulated, OPG is enhanced
during the differentiation of osteoblasts (31), while RANKL expression is reduced as
the differentiation progresses. In the osteoblastic cultures used
in the current study, OPG expression was higher than RANKL
expression. The results suggested that r-Mt cpn10 inhibited OPG
mRNA and protein expression in osteoblastic cells at all stages of
differentiation.
In conclusion, the present study demonstrated that
the effect of r-Mt cpn10 on osteoblastic cells was achieved via an
increase in the RANKL/OPG ratio. The paracrine mechanisms by which
r-Mt cpn10 induces bone resorption, and thus increases the risk of
fractures, may involve a local increase in the level of RANKL and
decrease in the level of OPG in the bone microenvironment.
Acknowledgements
This study was supported by the Xinjiang Natural
Science Foundation (no. 2011211A054).
References
|
1
|
Andersen TL, Sondergaard TE, Skorzynska
KE, et al: A physical mechanism for coupling bone resorption and
formation in adult human bone. Am J Pathol. 174:239–247. 2009.
View Article : Google Scholar :
|
|
2
|
Trouvin AP and Goëb V: Receptor activator
of nuclear factor-κB ligand and osteoprotegrin: maintaining the
balance to prevent bone loss. Clin Interv Aging. 5:345–354.
2010.
|
|
3
|
Dougall WC: Molecular pathways:
osteoclast-dependent and osteoclast-independent roles of the
RANKL/RANK/OPG pathway in tumorigenesis and metastasis. Clin Cancer
Res. 18:326–335. 2012. View Article : Google Scholar
|
|
4
|
Raju R, Balakrishnan L, Nanjappa V, et al:
A comprehensive manually curated reaction map of
RANKL/RANK-signalling pathway. Database (Oxford). 2011:bar0212011.
View Article : Google Scholar
|
|
5
|
Singh PP, van der Kraan AG, Xu J,
Gillespie MT and Quinn JM: Membrane-bound receptor activator of
NFκB ligand (RANKL) activity displayed by osteoblasts is
differentially regulated by osteolytic factors. Biochem Biophys Res
Commun. 422:48–53. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rachner TD, Khosla S and Hofbauer LC:
Osteoporosis: now and the future. Lancet. 377:1276–1287. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wu WT, Lee RP, Wang CH, et al: The
association of serum osteoprotegerin and osteoporosis in
postmenopausal hemodialysis patients: a pilot study. J Womens
Health (Larchmt). 19:785–790. 2010. View Article : Google Scholar
|
|
8
|
Reitmanova S and Gustafson DL: Coloring
the white plague: a syndemic approach to immigrant tuberculosis in
Canada. Ethn Health. 17:403–418. 2012. View Article : Google Scholar
|
|
9
|
Glaziou P, Falzon D, Floyd K and
Raviglione M: Global epidemiology of tuberculosis. Semin Respir
Crit Care Med. 34:3–16. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Corrado A, Neve A, Macchiarola A, et al:
RANKL/OPG ratio and DKK-1 expression in primary osteoblastic
cultures from osteoarthritic and osteoporotic subjects. J
Rheumatol. 40:684–694. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Henderson B, Lund PA and Coates AR:
Multiple moonlighting functions of mycobacterial molecular
chaperons. Tuberculosis (Edinb). 90:119–124. 2010. View Article : Google Scholar
|
|
12
|
Taneja B and Mande SC: Metal ions modulate
the plastic nature of Mycobacterium tuberculosis chaperonin-10.
Protein Eng. 14:391–395. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Meghji S, White PA, Nair SP, et al:
Mycobacterium tuberculosis chaperonin 10 stimulates bone
resorption: a potential contributory factor in Pott’s disease. J
Exp Med. 186:1241–1246. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jonsson KB, Frost A, Nilsson O, Ljunghall
S and Ljunggren O: Three isolation techniques for primary culture
of human osteoblastlike cells: a comparison. Acta Orthop Scand.
70:365–373. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Brändström H, Björkman T and Ljunggren O:
Regulation of osteoprotegerin secretion from primary cultures of
human bone marrow stromal cells. Biochem Biophys Res Commun.
280:831–835. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Suda T, Takahashi N, Udagawa N, et al:
Modulation of osteoclast differentiation and function by the new
members of the tumor necrosis factor receptor and ligand families.
Endocr Rev. 20:345–357. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Morony S, Tintut Y, Zhang Z, et al:
Osteoprotegerin inhibits vascular calcification without affecting
atherosclerosis in ldlr(−/−) mice. Circulation. 117:411–420. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jones DH, Nakashima T, Sanchez OH, et al:
Regulation of cancer cell migration and bone metastasis by RANKL.
Nature. 440:692–696. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang Y, Yang C, Xie WL, et al: Puerarin
concurrently stimulates osteoprotegerin and inhibits receptor
activator of NF-κB ligand (RANKL) and interleukin-6 production in
human osteoblastic MG-63 cells. Phytomedicine. 21:1032–1036. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xiong Q, Zhang LC, Zhang LH, et al:
Effects of recombinant human osteoprotegerin and recombinant RANK
protein on the differentiation of osteoclast precursors. Zhongguo
Gu Shang. 26:324–327. 2013.PubMed/NCBI
|
|
21
|
Goranova-Marinova V, Goranov S, Pavlov P
and Tzvetkova T: Serum levels of OPG, RANKL and RANKL/OPG ratio in
newly-diagnosed patients with multiple myeloma. Clinical
correlations. Haematologica. 92:1000–1001. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Samelson EJ, Broe KE, Demissie S, et al:
Increased plasma osteoprotegerin concentrations are associated with
indices of bone strength of the hip. J Clin Endocrinol Metab.
93:1789–1795. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kearns AE, Khosla S and Kostenuik PJ:
Receptor activator of nuclear factor kappaB ligand and
osteoprotegerin regulation of bone remodeling in health and
disease. Endocr Rev. 29:155–192. 2008. View Article : Google Scholar
|
|
24
|
Lamoureux F, Richard P, Wittrant Y, et al:
Therapeutic relevance of osteoprotegerin gene therapy in
osteosarcoma: blockade of the vicious cycle between tumor cell
proliferation and bone resorption. Cancer Res. 167:7308–7318. 2007.
View Article : Google Scholar
|
|
25
|
Joseph F, Chan BY, Durham BH, et al: The
circadian rhythm of osteoprotegerin and its association with
parathyroid hormone secretion. J Clin Endocrinol Metab.
92:3230–3238. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Roberts MM, Coker AR, Fossati G, et al:
Mycobacerium tuberculosis chaperonin 10 heptamers self-associate
through their biologically active loops. J Bacteriol.
185:4172–4185. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Qamra R, Mande SC, Coates AR and Henderson
B: The unusual chaperonins of Mycobacterium tuberculosis.
Tuberculosis (Edinb). 85:385–394. 2005. View Article : Google Scholar
|
|
28
|
Roberts MM, Coker AR, Fossati G, et al:
Mycobacterium tuberculosis chaperonin 10 heptamers self-associate
through their biologically active loops. J Bacteriol.
185:4172–4185. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Castanié-Cornet MP, Bruel N and Genevaux
P: Chaperone networking facilitates protein targeting to the
bacterial cytoplasmic membrane. Biochim Biophys Acta.
1843:1442–1456. 2014. View Article : Google Scholar
|
|
30
|
Henderson B, Lund PA and Coates AR:
Multiple moonlighting functions of mycobacterial molecular
chaperones. Tuberculosis (Edinb). 90:119–124. 2010. View Article : Google Scholar
|
|
31
|
Bai YD, Yang FS, Xuan K, Bai YX and Wu BL:
Inhibition of RANK/RANKL signal transduction pathway: a promising
approach for osteoporosis treatment. Med Hypotheses. 71:256–258.
2008. View Article : Google Scholar : PubMed/NCBI
|