Introduction
As a tissue that possesses a poor regenerative
capacity, articular cartilage tends to be structurally broken down
or degenerated under the shadow of disease, aging or trauma,
finally evolving into osteoarthritis (OA), where the prognosis is
poor (1,2). Over the course of OA development,
catabolic factors, including proinflammatory cytokines, are
activated, which induces the gradual self-destruction of cartilage
coupled with the curb of chondrogenic differentiation (3–5).
Accompanied with this process is the impact of
non-cartilage-specific extracellular matrix (ECM) with inferior
mechanical properties, which is produced by dedifferentiated
chondrocytes and can prevent chondroprogenitors from remodeling
cartilage defects through migration (6,7). These
factors lead to the acceleration and deterioration of OA.
As a traditional anti-inflammatory agent,
non-steroidal anti-inflammatory drugs have been administered to
patients with OA to attenuate associated symptoms, similar to other
therapies, including aminoglucose and sodium hyaluronate (HA)
(8). Tissue engineering, where cells
can be encapsulated in a carrier matrix, is a promising alternative
resolution although arthroplasty is time consuming and
microfracture suitable for minor lesions (9). Sustained phenotypic maintenance of
chondrocytes during expansion in vitro, and the synthesis of
mature ECM combined with anti-inflammation following
transplantation in vivo, are necessary for the successful
treatment of OA with tissue engineering (10,11),
which remains a serious challenge and requires further
research.
With marked antioxidation, anti-inflammatory and
antitumor properties, polyphenolic compounds isolated from plants,
such as green tea, and catechins, are attracting increasing
attention. Evidence suggests that polyphenols aid the
differentiation and phenotypic survival of numerous stomatocytes
(12). Among these, protocatechuic
acid (PCA) has been reported to possess analgesic and
anti-inflammatory activity in Freund's adjuvant arthritis (13). In addition, PCA has been shown to
stimulate the apoptosis of tumor cells (14) and resist the HA degradation of
experimental arthritis (15). An
additional study demonstrated that PCA benefited cell
differentiation and the maintenance of cellular phenotypes for
neural stem and progenitor cells in vitro (16). These findings indicated that as a
potent anti-inflammatory agent, PCA may exert an effect on
chondrocyte differentiation, which is of significance for the
treatment of long-term arthritis, via the maturation of ECM
secretion or the induction of chondrocyte expansion in cartilage
tissue engineering.
Therefore, PCA was hypothesized to be a potential
chondro-protective agent that may be applied to induce chondrocyte
in vitro expansion in cartilage tissue engineering for OA.
In the present study, the effect of PCA on the biological functions
of rabbit articular chondrocytes in vitro were investigated
through the determination of cytotoxicity, proliferation and cell
morphology, coupled with glycosaminoglycan (GAG) synthesis and
cartilage-specific gene expression. This study may provide a
reference for the application of PCA in cartilage tissue
engineering and the treatment of OA.
Materials and methods
Isolation and culture of articular
chondrocytes
A total of five one-week-old New Zealand rabbits
were purchased from the Center of Experimental Animals of Guangxi
Medical University (Nanning, China) and the relative operations
were approval by the Ethics Committee of Guangxi Medical
University. First, one rabbit was anaesthetized using
pentobarbitone sodium (60mg/kg, intraperitoneally; Sigma-Aldrich,
St. Louis, MO, USA). After sedation had been successfully induced,
150 mg pentobarbitone sodium was slowly administered in addition to
the initial dose, until the rabbit was euthanized. Standard
preoperative preparation was performed and cartilage slices were
harvested from hip and knee joints on a clean bench (JB-CJ-2FX;
Suzhou Jiebao Purification Engineering Equipment Co., Ltd.,
Jiangsu, China). These slices were primarily dissociated with 0.25%
trypsin (Solarbio, Beijing, China) for 30 min, and then with 2
mg/ml collagenase type II (Sigma-Aldrich) in α-modified Eagle's
medium (α-MEM; Gibco Life Technologies, Carlsbad, CA, USA) for 3 h.
Chondrocytes were isolated through centrifugation (400 × g, 5 min,
37°C) and resuspended in α-MEM containing 20% (v/v) fetal bovine
serum (Gibco Life Technologies) and 1% (v/v) antibiotics
(penicillin 100 U/ml and streptomycin 100 U/ml; Solarbio). The
cultures were maintained in a 5% CO2 incubator (Thermo
Fisher Scientific, Glasgow, UK) at 37°C, with the culture medium
changed every three days. Cells were passaged after reaching 80–90%
confluence. Confluent chondrocytes in a logarithmic growth phase
were prepared for the further experiments.
Preparation and treatment of PCA
PCA was purchased from Chengdu Must Bio-technology
Co., Ltd. (Chengdu, China). Prior to the experiment, PCA was
dissolved in 75% alcohol to form a 100-mmol/l stock solution.
Following filtration with a 0.22-µm filter (EMD Millipore,
Billerica, MA, USA) for sterilization, the solution was stored at
−4°C. The stock solution of PCA was added to the cell culture to
provide various concentrations for the subsequent experiments.
Cytotoxicity assay
Articular chondrocytes were cultured in 96-well
microplates (Corning Incorporated, New York, NY, USA) pretreated
with various concentrations of PCA (0–1 mmol/l) for three days. MTT
(5 mg/ml; Gibco Life Technologies) was added to the cultures in
each well. Following incubation at 37°C for 4 h, the culture medium
was removed and dimethyl sulfoxide (Gibco Life Technologies) was
added (150 µl per well). The microplates were gently shaken for 10
min by MH-2 Mini Shaker (Kylin-Bell Lab Instruments Co., Ltd.,
Jiangsu, China) in order to obtain a completely dissolved purple
solution. The optical density was detected at 570 nm using a
Multiskan GO Microplate spectrophotometer (Thermo Fisher
Scientific, USA).
Cell proliferation analysis and
biochemical assay
In light of the results of the cytotoxicity assay,
three doses of PCA with evident positive effects were selected
(0.0625, 0.125 and 0.25 mmol/l), along with a control group (0
mmol/l PCA), for the cell proliferation analysis and biochemical
assay. Chondrocytes in the different groups were cultured for two,
four and six days. The cells were digested with 0.25% trypsin and
resuspended in phosphate-buffered saline (PBS) containing 60 µg/ml
proteinase K (Sigma-Aldrich) for 16 h at 60°C. Following the
application of Hoechst 33258 (Sigma-Aldrich), the proliferation of
cells was analyzed via DNA production using an ultraviolet Hitachi
F-4500 spectrofluorometer (Hitachi, Ltd., Tokyo, Japan) at 460 nm,
with the absorbance value of Hoechst 33258 dye used as the
baseline.
The total production of GAGs was measured through
absorbance with a 1,9-dimethylmethylene blue spectrophotometric
assay at 525 nm, with chondroitin sulfate (Sigma-Aldrich) as the
standard sample. The synthesis and secretion of GAGs was calculated
according to the standard curve and normalized against the total
DNA production, which revealed the biosynthetic activity of the
cells in diverse culture media.
Morphological examination
Following culture for six days, the cells were fixed
with 95% alcohol for 30 min and then rinsed three times in PBS,
once every 3 min. Subsequently, chondrocytes were stained by
hematoxylin-eosin (HE) using a commercial kit (Jiancheng Biotech
Co., Ltd., Nanjing, China) according to the manufacturer
instructions. An inverted phase contrast microscope (Zeiss
International, Oberkochen, Germany) was used to conduct the cell
morphological analysis.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
To further investigate the effect of PCA on the
expression of cartilage-specific genes, the mRNA expression levels
of collagen I, II and X, aggrecan and Sox9 were analyzed by
RT-qPCR. Total RNA was extracted from the articular chondrocytes
using an RNeasy RNA extraction kit (Tiangen Biotechnology, Beijing,
China), according to the manufacturer's instructions. Total RNA
(~300 ng) was used as a template and reverse transcribed into cDNA
with a reverse transcription kit (Fermentas; Thermo Fisher
Scientific). cDNA was amplified using a SYBR® Green Master Mix kit
(Roche Diagnostics GmbH, Mannheim, Germany). qPCR was performed
using a qPCR Detection System (Realplex 4; Eppendorf, Enfield, CT,
USA) with FastStart Universal SYBR® Green Master Mix at 95°C for 5
min for the initial denaturation, then 40 cycles of 95°C for 15 s
and 60°C for 1 min. The designed primers used for PCR are shown in
Table I. The primer specificity was
confirmed by analyzing the dissociation curve of each primer pair.
Relative gene expression levels were calculated using the
2−ΔΔCt method, relative to GAPDH gene expression. Each
gene was analyzed in triplicate to reduce randomization error.
 | Table I.Primer sequences used in the reverse
transcription-quantitative polymerase chain reaction
experiments. |
Table I.
Primer sequences used in the reverse
transcription-quantitative polymerase chain reaction
experiments.
| mRNA | Forward primer | Reverse primer |
|---|
| GAPDH |
5′-CTATAAATTGAGCCCGCAGC-3′ |
5′-ACCAAATCCGTTGACTCCG-3′ |
| Aggrecan |
5′-CTACACGCTACACCCTCGAC-3′ |
5′-ACGTCCTCACACCAGGAAAC-3′ |
| Type I
collagen |
5′-GTTCAGCTTTGTGGACCTCCG-3′ |
5′-GCAGTTCTTGGTCTCGTCAC-3′ |
| Type II
collagen |
5′-AAGCTGGTGAGAAGGGACTG-3′ |
5′-GGAAACCTCGTTCACCCCTG-3′ |
| Type X
collagen |
5′-CGCTGAACGATACCAAATGCC-3′ |
5′-TTCCCTACAGCTGATGGTCC-3′ |
| Sox9 |
5′-AAGCTCTGGAGACTTCTGAACG-3′ |
5′-CGTTCTTCACCGACTTCCTCC-3′ |
Statistical analysis
Statistical analyses were conducted using SPSS
software, version 17.0 (SPSS, Inc., Chicago, IL, USA). Results are
expressed as the mean ± standard deviation for quantitative data.
Statistical significance was determined using one way analysis of
variance followed by Dunnett's post hoc test. P<0.05 was
considered to indicate a statistically significant difference.
Results
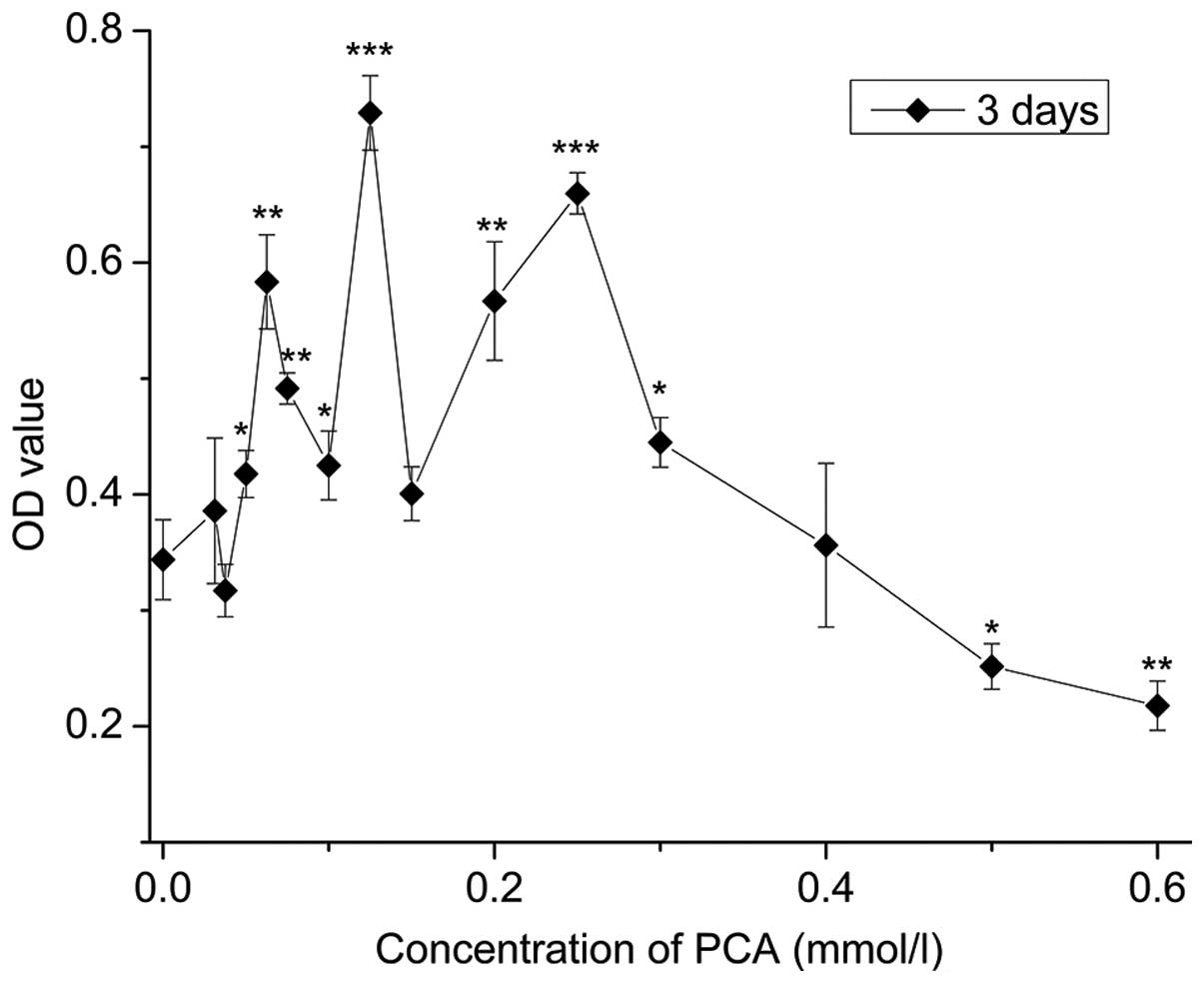
Cytotoxicity assay
As depicted in Fig.
1, compared with the control group (0 mmol/l), 0.03125–0.4
mmol/l PCA indicated OD values with no statistical significance or
higher that indicated low cytotoxicity; OD values of 0.0625–0.3
mmol/l PCA significantly increased (P<0.05), indicating
0.0625–0.3 mmol/l PCA accelerated cell growth (P<0.05), with the
most evident effect at a dose of 0.125 mmol/l. By contrast, at
concentrations ranging between 0.5 and 0.6 mmol/l PCA,
proliferation inhibition of the rabbit articular chondrocytes in
vitro was observed when compared with the control group.
Cell proliferation
Chondrocytes cultured with 0.0625, 0.125 and 0.25
mmol/l PCA grew faster than those in the control group (0 mmol/l
PCA), as indicated by the significantly higher DNA content
(P<0.05) in the same culture period (Fig. 2A). Among the three concentrations,
0.125 mmol/l PCA exhibited the strongest promoting effect on cell
growth at the same time-point of culture.
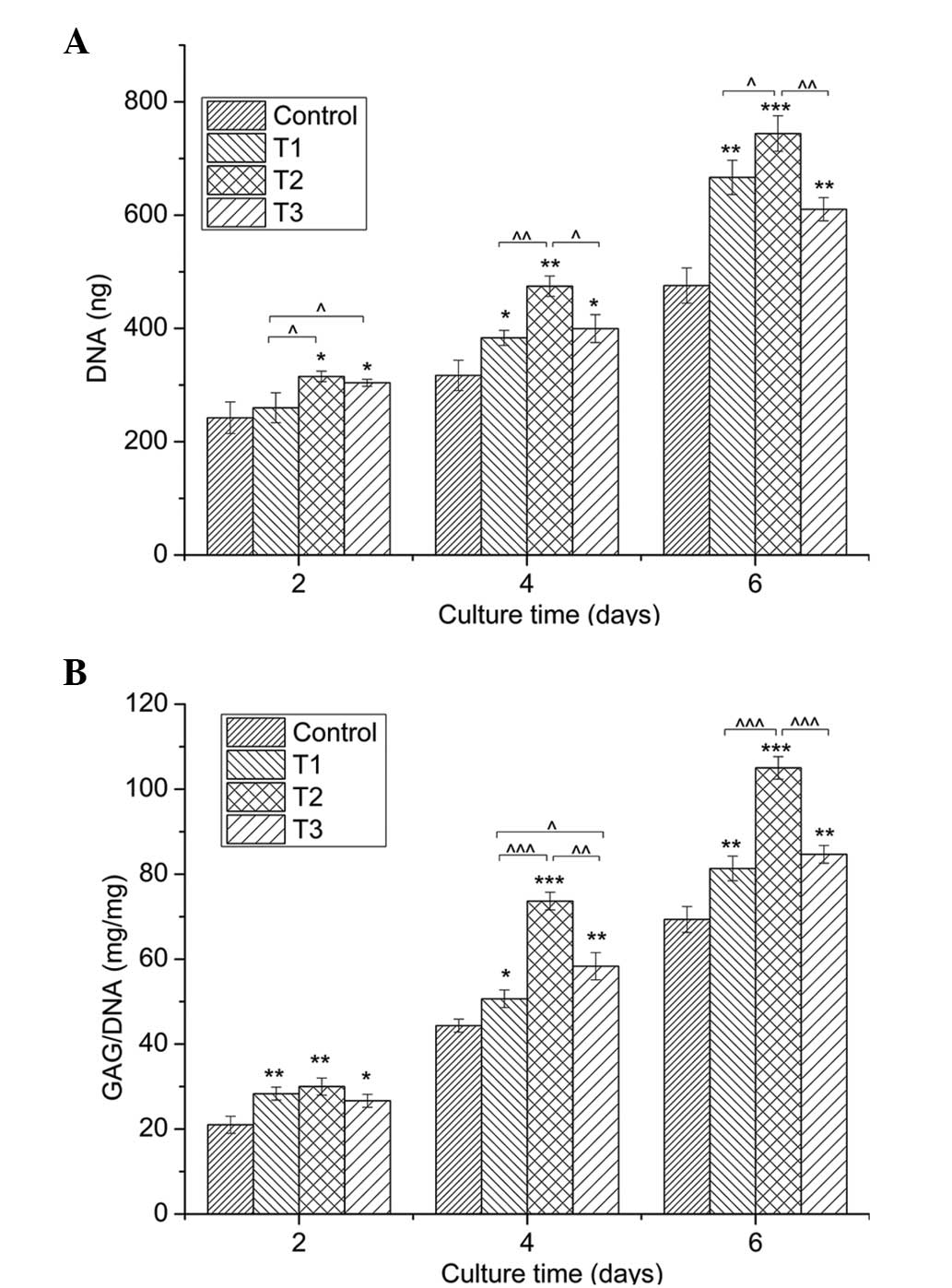 | Figure 2.Quantification of cell proliferation
by the detection of DNA content and matrix production by GAG
analysis. (A) Proliferation of chondrocytes cultured in
vitro with 0 (control), 0.0625 (T1), 0.125 (T2) and 0.25 mmol/l
(T3) protocatechuic acid for two, four and six days. (B) GAG
synthesis (mg) normalized against DNA content (mg). Data from three
independent experiments were evaluated, and the results are
presented as the mean ± standard deviation.
*,^P<0.05; **,^^P<0.01; and
***,^^^P<0.001. *, ** and *** vs. control.
^, ^^ and ^^^ vs. as indicated.
GAG, glycosaminoglycan. |
Secretion of GAGs
Fig. 2B demonstrates
an evident increase in the amount of GAGs, provided as a ratio of
GAG/DNA, in the PCA groups when compared with the control group for
the same culture period (P<0.05). In line with the cell
proliferation determined by DNA content, PCA at a dose of 0.125
mmol/l produced the best effect on GAG synthesis.
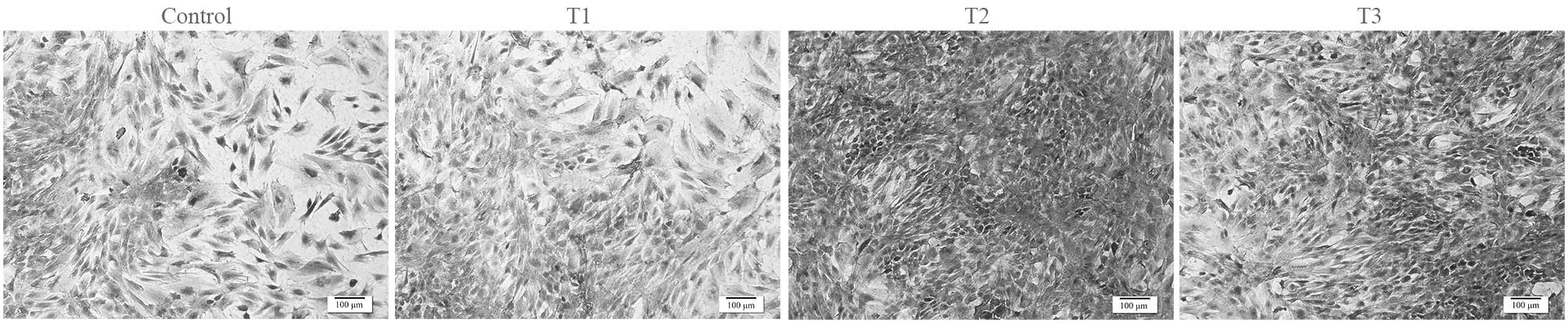
Cell morphology
Evaluation of chondrocyte morphology using HE
staining demonstrated that chondrocytes treated with PCA grew
better compared with the control group (Fig. 3). In the PCA groups, a higher number
of round cells were identified, which represented the typical
morphology of chondrocytes. In addition, PCA at the dose of 0.125
mmol/l was most effective at facilitating the proliferation of
rabbit articular chondrocytes in vitro.
Gene expression
The positive role of PCA on ECM synthesis was
further verified by examination of the expression of aggrecan, Sox9
and collagen I, II and X (Fig. 4).
The mRNA expression levels of the cartilage-specific genes,
aggrecan, collagen II and Sox9, were evidently promoted by PCA when
compared with the control group. In addition, the expression of the
collagen I gene, a marker of cell dedifferentiation, was
downregulated by PCA when compared with the control cells. The
expression of collagen X, an indicator of cell hypertrophy, was not
detected. Among the experimental groups, PCA at a concentration of
0.125 mmol/l exhibited the optimum performance with regard to the
upregulation of the expression levels of aggrecan, collagen II and
Sox9 genes, as well as the downregulation of collagen I gene
expression.
 | Figure 4.Quantitative comparison of
extracellular-matrix-related gene expression by reverse
transcription-quantitative polymerase chain reaction. Chondrocytes
were cultured with 0 (control), 0.0625 (T1), 0.125 (T2) and 0.25
mmol/l (T3) protocatechuic acid (PCA) for (A) two, (B) four and (C)
six days (n=3 for each group/time-point). Gene expression levels in
the PCA groups, relative to the control group, were analyzed by the
2−ΔΔCt method using GAPDH as the internal control. Data
are expressed as the mean ± standard deviation.
*,^P<0.05, **,^^P<0.01,
***,^^^ P<0.001. |
Discussion
PCA exists in numerous fruit and vegetables and is
readily available in daily life. The chemical composition of PCA is
similar to gallic acid, which are both categorized as a
polyphenolic compounds known to exert antioxidant,
anti-inflammatory and anticancer effects (17,18). PCA
and its analogs possess potent anti-inflammatory effects and have
been demonstrated to be effective in animal models of arthritis
(13). Based on the hypothesis that
PCA may serve as a potential chondro-protective agent, the impact
of PCA on the growth and phenotype maintenance of articular
chondrocytes in vitro was considered in the present study.
The results indicated that PCA was able to promote chondrocyte
proliferation and GAG deposition in chondrocytes (Figs. 2 and 3). Proteoglycans are crucial components of
the ECM (19). The ECM is
responsible for maintaining the cartilage load-bearing capacity
(20), and also plays a vital role
in chondrocyte phenotype (21).
Consistent with the increase in GAG production, PCA
was demonstrated to upregulate the gene expression of Sox9,
collagen II and aggrecan (Fig. 4).
The chondrogenic transcription factor, Sox9, is essential for
increasing the rate of chondrogenesis (22,23),
particularly when coexpressed with collagen II (24–26). In
addition, several gene therapy approaches, namely viral methods to
overexpress Sox9, have been shown to significantly improve the
synthesis of cartilaginous matrix produced by bone marrow-derived
stem cells and articular chondrocytes (27–29).
Evidence has also indicated that aggrecan production is
significantly upregulated by the Sox9 gene, as an early
chondrogenic marker (27,30). Therefore, a possible mechanism
underlying the promotive effect of PCA on chondrocyte growth and
matrix secretion may be the modulation of Sox9 expression.
Maintaining the chondrocyte phenotype is one of the
major challenges for cartilage tissue engineering and inhibition of
OA development (31,32). Dedifferentiation of articular
chondrocytes tends to appear as the culture time progresses in
tissue engineering or in the development of OA (11,33).
Simultaneously, non-cartilage-specific ECM is produced, which is
characteristic of a poor biomechanical response. PCA has been
demonstrated to enhance differentiation and benefit phenotypic
survival in neural-related cells (34). In the current study, upregulation of
collagen I gene expression, a marker of dedifferentiation, was not
detected in the PCA groups (Fig. 4),
indicating that PCA is beneficial to the phenotypic maintenance of
chondrocytes. Dedifferentiation occurs when the differentiated
phenotype of chondrocytes, consisting primarily of type II collagen
and cartilage-specific proteoglycan, is lost and replaced by a
complex collagen phenotype consisting predominately of type I
collagen and a low level of proteoglycan synthesis (35–37).
Hypertrophy is a predictor of secondary cell phenotype loss
(32,38), and the sequence of hypertrophy is
endochondral ossification (39). The
results of the present study revealed that the associated gene,
collagen X, was not detected in any of the groups (Fig. 4). Therefore, the reduced expression
of collagen I and the undetectable expression of collagen X
following PCA administration indicated that PCA promoted rabbit
articular chondrocytes in vitro to maintain their
phenotype.
As for the recommended concentration of PCA, the
proliferation of rabbit articular chondrocytes in vitro was
accelerated with PCA concentrations ranging between 0.05 and 0.3
mmol/l. In particular, at a dose of 0.125 mmol/l, PCA exhibited the
optimum performance with regard to cell growth and phenotype
maintenance. However, whether this treatment is suitable for
articular chondrocytes of other species, such as humans, is unable
to be confirmed. A lack of evidence also exists with regard to the
application of PCA in experiments in vivo.
In conclusion, PCA exerts a positive effect on the
proliferation and phenotypic maintenance of rabbit articular
chondrocytes in vitro, with the optimal concentration being
0.125 mmol/l. Therefore, PCA, a polyphenol compound widely found in
vegetable matter, may serve as a potential agent in the field of
cartilage tissue engineering and treatment of OA; however, further
studies are required.
Acknowledgements
This study was supported by the National Science
& Technology Pillar Program of China (no. 2012BAI42G00),
Guangxi Scientific Research and Technological Development
Foundation (no. Guikehe 14125008-2-14), the Guangxi Science Fund
for Distinguished Young Scholars (no. 2014GXNSFGA118006), the Key
Laboratory of Regenerative Medicine of Guangxi High School and the
Research Center for Regenerative Medicine and Collaborative
Innovation Center of Guangxi Biological Medicine.
Glossary
Abbreviations
Abbreviations:
|
PCA
|
protocatechuic acid
|
|
OA
|
osteoarthritis
|
|
ECM
|
extracellular matrix
|
|
GAG
|
glycosaminoglycan
|
|
HA
|
hyaluronate acid
|
|
HE
|
hematoxylin-eosin
|
|
RT-qPCR
|
reverse transcription-quantitative
polymerase chain reaction
|
References
|
1
|
van der Kraan PM: Age-related alterations
in TGF beta signaling as a causal factor of cartilage degeneration
in osteoarthritis. Biomed Mater Eng. 24:(Suppl). 75–80.
2014.PubMed/NCBI
|
|
2
|
Tetteh ES, Bajaj S and Ghodadra NS: Basic
science and surgical treatment options for articular cartilage
injuries of the knee. J Orthop Sports Phys Ther. 42:243–253. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Stone AV, Loeser RF, Vanderman KS, Long
DL, Clark SC and Ferguson CM: Pro-inflammatory stimulation of
meniscus cells increases production of matrix metalloproteinases
and additional catabolic factors involved in osteoarthritis
pathogenesis. Osteoarthritis Cartilage. 22:264–274. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ma B, Leijten JC, Wu L, et al: Gene
expression profiling of dedifferentiated human articular
chondrocytes in monolayer culture. Osteoarthritis Cartilage.
21:599–603. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Schulze-Tanzil G: Activation and
dedifferentiation of chondrocytes: implications in cartilage injury
and repair. Ann Anat. 191:325–338. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nicolini AP, Carvalho RT, Dragone B, Lenza
M, Cohen M and Ferretti M: Updates in biological therapies for knee
injuries: full thickness cartilage defect. Curr Rev Musculoskelet
Med. 7:256–262. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Drissi H, Zuscik M, Rosier R and O'Keefe
R: Transcriptional regulation of chondrocyte maturation: potential
involvement of transcription factors in OA pathogenesis. Mol
Aspects Med. 26:169–179. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bruyère O, Cooper C, Pelletier JP, et al:
An algorithm recommendation for the management of knee
osteoarthritis in Europe and internationally: A report from a task
force of the European Society for Clinical and Economic Aspects of
Osteoporosis and Osteoarthritis (ESCEO). Semin Arthritis Rheum.
44:253–263. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mollon B, Kandel R, Chahal J and
Theodoropoulos J: The clinical status of cartilage tissue
regeneration in humans. Osteoarthritis Cartilage. 21:1824–1833.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fahy N, Farrell E, Ritter T, Ryan AE and
Murphy JM: Immune modulation to improve tissue engineering outcomes
for cartilage repair in the osteoarthritic joint. Tissue Eng Part B
Rev. Aug 4–2014.(Epub ahead of print). PubMed/NCBI
|
|
11
|
Coates EE and Fisher JP: Phenotypic
variations in chondrocyte subpopulations and their response to in
vitro culture and external stimuli. Ann Biomed Eng. 38:3371–3388.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ignat I, Volf I and Popa VI: A critical
review of methods for characterisation of polyphenolic compounds in
fruits and vegetables. Food Chem. 126:1821–1835. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lende AB, Kshirsagar AD, Deshpande AD, et
al: Anti-inflammatory and analgesic activity of protocatechuic acid
in rats and mice. Inflammopharmacology. 19:255–263. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lo CW, Huang HP, Lin HM, Chien CT and Wang
CJ: Effect of Hibiscus anthocyanins-rich extract induces apoptosis
of proliferating smooth muscle cell via activation of P38 MAPK and
p53 pathway. Mol Nutr Food Res. 51:1452–1460. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yoon CH, Chung SJ, Lee SW, Park YB, Lee SK
and Park MC: Gallic acid, a natural polyphenolic acid, induces
apoptosis and inhibits proinflammatory gene expressions in
rheumatoid arthritis fibroblast-like synoviocytes. Joint Bone
Spine. 80:274–279. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Guan S, Zhang XL, Ge D, Liu TQ, Ma XH and
Cui ZF: Protocatechuic acid promotes the neuronal differentiation
and facilitates survival of phenotypes differentiated from cultured
neural stem and progenitor cells. Eur J Pharmacol. 670:471–478.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kakkar S and Bais S: A review on
protocatechuic acid and its pharmacological potential. ISRN
Pharmacol. 2014:9529432014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hsu CC, Hsu CL, Tsai SE, Fu TY and Yen GC:
Protective effect of Millettia reticulata Benth against
CCl(4)-induced hepatic damage and inflammatory action in rats. J
Med Food. 12:821–828. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Buschmann MD and Grodzinsky AJ: A
molecular model of proteoglycan-associated electrostatic forces in
cartilage mechanics. J Biomech Eng. 117:179–192. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Horkay F: Interactions of cartilage
extracellular matrix macromolecules. J Polym Sci B Polym Phys.
50:1699–1705. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Grogan SP, Chen X, Sovani S, et al:
Influence of cartilage extracellular matrix molecules on cell
phenotype and neocartilage formation. Tissue Eng Part A.
20:264–274. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Akiyama H: Transcriptional regulation in
chondrogenesis by Sox9. Clin Calcium. 21:845–851. 2011.(In
Japanese). PubMed/NCBI
|
|
23
|
Tew SR and Clegg PD: Analysis of post
transcriptional regulation of SOX9 mRNA during in vitro
chondrogenesis. Tissue Eng Part A. 17:1801–1807. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ng LJ, Wheatley S, Muscat GE, et al: SOX9
binds DNA, activates transcription, and coexpresses with type II
collagen during chondrogenesis in the mouse. Dev Biol. 183:108–121.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Marshall OJ and Harley VR: Molecular
mechanisms of SOX9 action. Mol Genet Metab. 71:455–462. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Davies SR, Chang LW, Patra D, et al:
Computational identification and functional validation of
regulatory motifs in cartilage-expressed genes. Genome Res.
17:1438–1447. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tew SR, Li Y, Pothacharoen P, Tweats LM,
Hawkins RE and Hardingham TE: Retroviral transduction with SOX9
enhances re-expression of the chondrocyte phenotype in passaged
osteoarthritic human articular chondrocytes. Osteoarthritis
Cartilage. 13:80–89. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Paul R, Haydon RC, Cheng H, et al:
Potential use of Sox9 gene therapy for intervertebral degenerative
disc disease. Spine. 28:755–763. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tsuchiya H, Kitoh H, Sugiura F and
Ishiguro N: Chondrogenesis enhanced by overexpression of sox9 gene
in mouse bone marrow-derived mesenchymal stem cells. Biochem
Biophys Res Commun. 301:338–343. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bi W, Deng JM, Zhang Z, Behringer RR and
de Crombrugghe B: Sox9 is required for cartilage formation. Nat
Genet. 22:85–89. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
31
|
Gan L and Kandel RA: In vitro cartilage
tissue formation by co-culture of primary and passaged
chondrocytes. Tissue Eng. 13:831–842. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Fosang AJ and Beier F: Emerging frontiers
in cartilage and chondrocyte biology. Best Pract Res Clin
Rheumatol. 25:751–766. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bailey AM: Balancing tissue and tumor
formation in regenerative medicine. Sci Transl Med. 4:147fs282012.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Guan S, Ge D, Liu TQ, Ma XH and Cui ZF:
Protocatechuic acid promotes cell proliferation and reduces basal
apoptosis in cultured neural stem cells. Toxicol In Vitro.
23:201–208. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Benya PD and Shaffer JD: Dedifferentiated
chondrocytes reexpress the differentiated collagen phenotype when
cultured in agarose gels. Cell. 30:215–224. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Schnabel M, Marlovits S, Eckhoff G, et al:
Dedifferentiation-associated changes in morphology and gene
expression in primary human articular chondrocytes in cell culture.
Osteoarthritis Cartilage. 10:62–70. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Karlsen TA, Shahdadfar A and Brinchmann
JE: Human primary articular chondrocytes, chondroblasts-like cells,
and dedifferentiated chondrocytes: differences in gene, microRNA,
and protein expression and phenotype. Tissue Eng Part C Methods.
17:219–227. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Grassel S and Ahmed N: Influence of
cellular microenvironment and paracrine signals on chondrogenic
differentiation. Front Biosci. 12:4946–4956. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Dreier R: Hypertrophic differentiation of
chondrocytes in osteoarthritis: the developmental aspect of
degenerative joint disorders. Arthritis Res Ther. 12:2162010.
View Article : Google Scholar : PubMed/NCBI
|