Introduction
The longstanding theory that water passes through
the intestinal epithelia using simple diffusion was reassessed
following the identification of aquaporins (AQPs) (1). The current, established hypothesis
posits that water can cross the epithelia either by a paracellular
pathway across the tight junctions or by transcellular pathways,
involving 3 mechanisms as follows: Simple diffusion across the
lipid bilayer; cotransport with ions and solutes; and diffusion
across AQPs (1–3). AQPs are a family of homologous, water
channel proteins that provide a major route for osmotically-driven
water movement across the plasma membrane in various cell types.
Currently, 13 distinct subtypes of AQPs (AQPs 0–12) have been
identified, which are functionally subdivided into orthodox AQPs
(AQPs 1, 2, 4, 5 and 8, which are primarily water selective),
aquaglyceroporins (AQPs 3, 7, 9 and 10, which are permeable to
neutral solutes including glycerol and other small solutes, in
addition to water) and unorthodox AQPs (AQPs 6, 11 and 12, for
which localization and functions remain undetermined) (4–7).
Specific physiological roles for a number of the
AQPs have been established through phenotype analysis of
AQP-knockout mice. Compared with other organs, the role of AQPs in
the kidney has been extensively studied, in part as the kidney is
the most important organ involved in the homeostasis of water in
the body. Previous studies have reported the expression of AQPs at
various sites throughout the kidney, and these are suggested to be
important in the urinary concentrating mechanism (8). The quantity of fluid transported in the
intestinal tract is second only to that in the kidney, but the role
of AQPs in intestinal physiology is comparatively unexplored.
At least 9 AQP subtypes (AQPs 1, 3, 4, 5, 7, 8, 9,
10 and 11) have been localized to the mammalian intestine (9–14), as
follows: AQP1 in the endothelia and lacteals (9); AQP3 at the basolateral membrane of the
epithelial cells lining the villus tip in the small intestine
(10); AQP4 at the basolateral
membrane of the crypt epithelium; AQP5 in the apical membrane of
secretory cells in the duodenal glands; AQP7 and AQP8 at the apical
membrane of the small intestine and colon (11); AQP9 in goblet cells in the small
intestine (10); AQP10 in the
duodenum and jejunum (12,13); and AQP11 in the human duodenum and
colon (14). Based on the multiple
expression sites of AQPs in the intestinal tract, it is reasonable
to presume that AQPs may effect an important functional role in
water transport; however, in previous studies, phenotypic analyses
of mice lacking AQPs did not support this hypothesis. For example,
the water content in cecal stool from AQP4-null mice was similar to
that from wild-type mice (15), and
only minor phenotypic differences between wild-type and
AQP8-knockout mice have been reported with regard to intestinal
fluid absorption and secretion (16), leaving the physiological role of AQPs
in the intestine unresolved. However, evidence of AQP tissue
expression without demonstrable physiological function does not
rule out possible functional roles of AQPs in the intestine under
stressed or diseased conditions, as reported in other tissues
(17–19). To test this hypothesis, the present
study investigated the effects of hypertonic stress and
ischemia/reperfusion injury on the expression level of AQP mRNA and
protein in cultured intestinal epithelial cells, using
semi-quantitative reverse transcription polymerase chain reaction
(RT-PCR) and western blot analyses, respectively.
Materials and methods
Chemicals
All chemicals were purchased from Sigma Chemical
Company (St. Louis, MO, USA).
Cell culture and treatment
Rat intestinal epithelial cells (IEC-6; Bioresource
Collection and Research Center, Hsinchu, Taiwan) were cultivated in
Dulbecco's modified Eagle's medium (DMEM; Sigma-Aldrich, St. Louis,
MO, USA) with high glucose (4,500 mg/l), containing 5% fetal bovine
serum (FBS; Hyclone; GE Healthcare Life Sciences, Logan, UT, USA)
and penicillin-streptomycin (1:100; Sigma-Aldrich). To study the
effects of hypertonicity on AQP expression in IEC-6 cells, the
cells were seeded in 10-cm dishes and grown to confluence. Culture
medium was then replaced with hypertonic medium and the cells were
incubated for 16 h. Hypertonic medium was made by addition of 300
mOsmol mannitol in isosmolar DMEM. For ischemic treatment, the
cells were seeded in 10-cm dishes and grown to confluence in 5%
FBS-supplemented DMEM. The cells were then washed with
phosphate-buffered saline (PBS) prior to ischemic treatment. In
this method, the cultures were treated with FBS- and glucose-free
DMEM and placed into a modular incubator chamber
(Billups-Rothenberg, Del Mar, CA, USA) that was sealed and flushed
with 95% N2/5% CO2 for 30 min at a rate of 2
l/min. The hypoxic chamber was then sealed tightly and placed in a
37°C, humidified, 5% CO2/95% air incubator for 8 h.
Using this method, the O2 concentration of the culture
media was measured as 0.8–1.0% using a Dissolved Oxygen Meter,
Model 5509 (Lutron, Taipei, Taiwan); this was significantly lower
than that of the normoxic cultures (5–6%). At the completion of
ischemic exposure, half of the cultures were removed from the
chamber and processed for analysis as described later. For the
simulated ‘ischemia/reperfusion’ treatment, the other half of the
cultures were removed from the chamber, the ischemic medium was
replaced with normal culture medium (DMEM with FBS and glucose) and
the culture was maintained in the 95% N2/5%
CO2 incubator for the 2-h reperfusion period. This study
was approved by the Animal Ethics Committee of Kaohsiung Armed
Forces General Hospital Zuoying Branch (ZAFGH-101-09; Kaohsiung,
Taiwan).
MTT reduction assay
Cell viability at the end of each experiment was
analyzed by a MTT reduction assay. This assay is based upon the
capacity of mitochondrial enzymes to transform MTT to MTT formazan.
Briefly, MTT stock solution (5 mg/ml in PBS) was added to each well
of the multi-well plate containing cells at a final concentration
of 0.5 mg/ml. Following incubation at 37°C for 2 h, the medium was
aspirated and an equal volume of dimethyl sulfoxide was added to
each well in order to dissolve the reduced MTT formazan crystals.
The absorbance of the color product at 570 nm against the 630-nm
reference was measured using a microplate reader (Sunrise; Tecan
Systems, Inc., San Jose, CA, USA). All experiments were performed
in triplicate.
RT-PCR
Total RNA was isolated using TRIzol reagent
(Invitrogen, Carlsbad, CA, USA) according to the manufacturer's
instructions. Total RNA (5 µg) was reverse transcribed with random
primers obtained from a High Capacity cDNA Reverse Transcription
Kit (Applied Biosystems, Foster City, CA, USA). The cDNA was then
amplified using polymerase chain reaction with primers specific
against various AQPs (Table I). The
PCR products were electrophoresed in 1% agarose gels and the bands
were visualized by ethidium bromide staining. Densitometric
analysis was performed using Quantity One software (Bio-Rad
Laboratories, Inc., Hercules, CA, USA) and corrected for loading
using the β-actin gene.
 | Table I.Primers for AQP expression. |
Table I.
Primers for AQP expression.
| mRNA | Primer sequence
(5′-3′) |
|---|
| AQP1 |
|
| F |
CTGTGGTGGCTGAGTTCCTG |
| R |
ATTTCGGCCAAGTGAGTTCTC |
| AQP3 |
|
| F |
CTGTGGTGGCTGAGTTCCTG |
| R |
ATTTCGGCCAAGTGAGTTCTC |
| AQP4 |
|
| F |
TTGGACCAATCATAGGCGC |
| R |
GTCAATGTCGATCACATGC |
| AQP7 |
|
| F |
GCTTCGTGGATGAGGTATT |
| R |
ACTTATGGGTAGGGTAGGTTT |
| AQP8 |
|
| F |
GGTGGACACTTCAACCCTGC |
| R |
CCCAGCCAGTAGATCCAATG |
| AQP9 |
|
| F |
GATGGACTCATGGCCTTTGCTG |
| R |
CAATCATAGGACCCACGACAGG |
| AQP11 |
|
| F |
ATCCCTAGCGGTGAGGGAAC |
| R |
CACTGACTTCGTTTGAGTCTTTGG |
| β-actin |
|
| F |
ACAATGAGCTGCGTGTGGCC |
| R |
GGAACCGCTCATTGCCGATAG |
Western blot analysis
The cells were harvested in ice-cold lysis buffer
[50 mM Tris (pH 8.0), 150 mM NaCl, 0.1% SDS, 1% NP-40 and 1 mM
phenylmethylsulfonyl fluoride] containing protease inhibitor
cocktail and then ultrasonicated. The homogenate was centrifuged at
13,000 × g for 20 min and the supernatant collected. Protein
concentrations of supernatants were determined. Samples of
supernatants containing 100 µg protein were separated by 12%
SDS-polyacrylamide gel electrophoresis and transferred to Hybond-P
polyvinylidene difluoride membranes (EMD Millipore, Billerica, MA,
USA) by electroelution. After being blocked for 3 h with
Tris-buffered saline containing 5% skimmed milk powder and 0.1%
Tween-20, the membranes were incubated overnight with rabbit
polyclonal antibodies against AQPs 1, 3, 4, 7, 8, 9 and 11 (2 µg/ml
for each AQP; Alpha Diagnostic, TX, USA) or β-actin
(Sigma-Aldrich), diluted in the blocking solution. The membranes
were washed and incubated with horseradish peroxidase-conjugated
secondary antibodies for 1 h. Finally, the signals were detected by
enhanced chemiluminescence detection kit (GE Healthcare Life
Sciences, Shanghai, China). The chemiluminescent signal was
captured by a BioSpectrum 500 imaging system (UVP, Inc., Upland,
CA, USA).
Data analysis
All data are expressed as the mean ± standard error
of the mean. Statistical evaluations were made by Student's t-test,
using SigmaStat Software (Jandel Scientific, San Rafael, CA, USA).
P<0.05 was considered to indicate a statistically significant
difference.
Results
Cell viability and AQP expression in
response to hyperosmotic stress
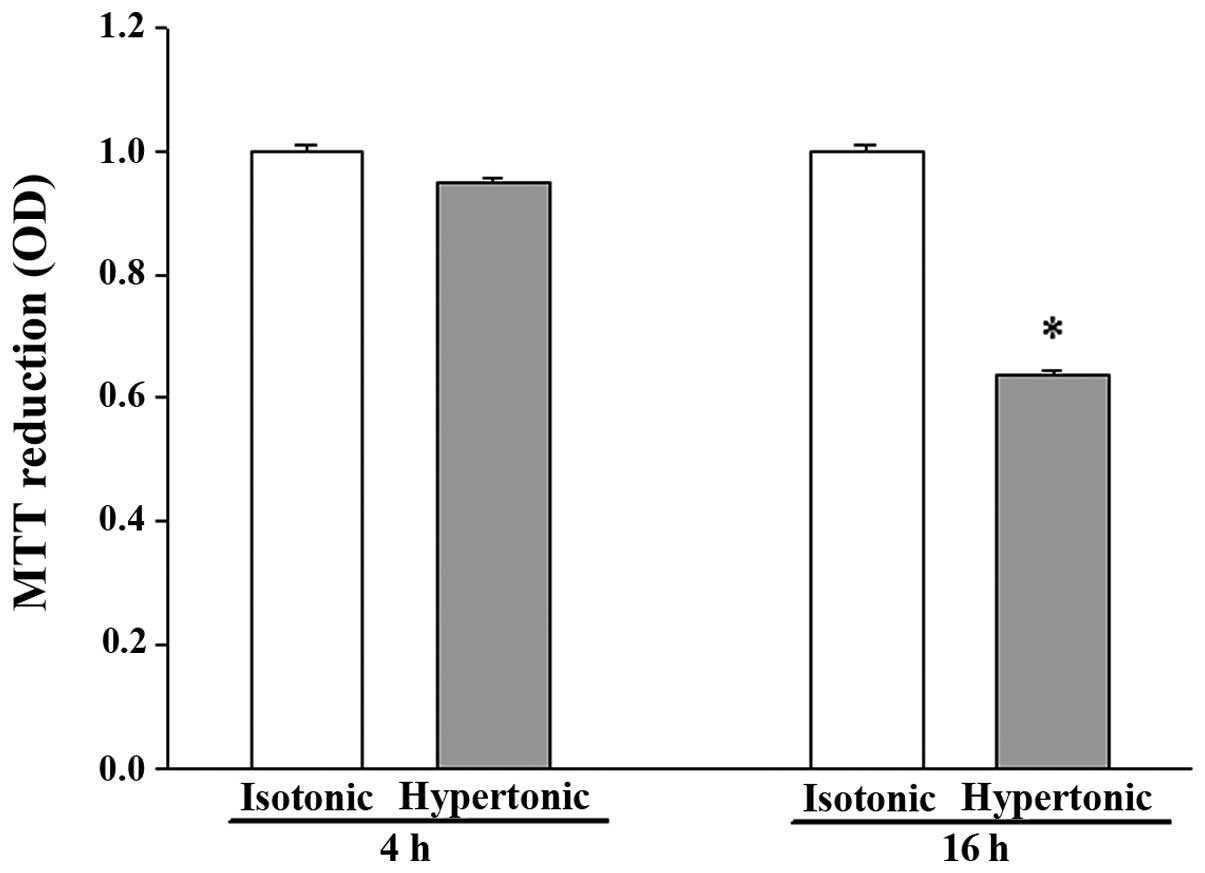
Exposure to hypertonicity for 4 h did not
significantly alter the viability of IEC-6 cells, while 16 h of
treatment resulted in apparent cell shrinkage and reduced viability
(Fig. 1).
There was no detectable difference in AQP1 mRNA
abundance after 4 h in hypertonic media when compared to its
abundance in isotonic (control) conditions (Fig. 2). Despite the moderate expression of
AQP1 mRNA in isotonic conditions (Fig.
2), an extremely low level of AQP1 protein was noted (Fig. 3). After 16 h of hypertonic exposure,
AQP1 mRNA expression was significantly enhanced (Fig. 2) and protein expression was
concomitantly elevated, undergoing a 37-fold increase over the
expression levels observed in isotonic conditions (Fig. 3). The mRNA and protein expression
level of AQPs 3, 4, 7 and 8 in response to 4 or 16 h of osmotic
stress were similar to their corresponding control levels (data not
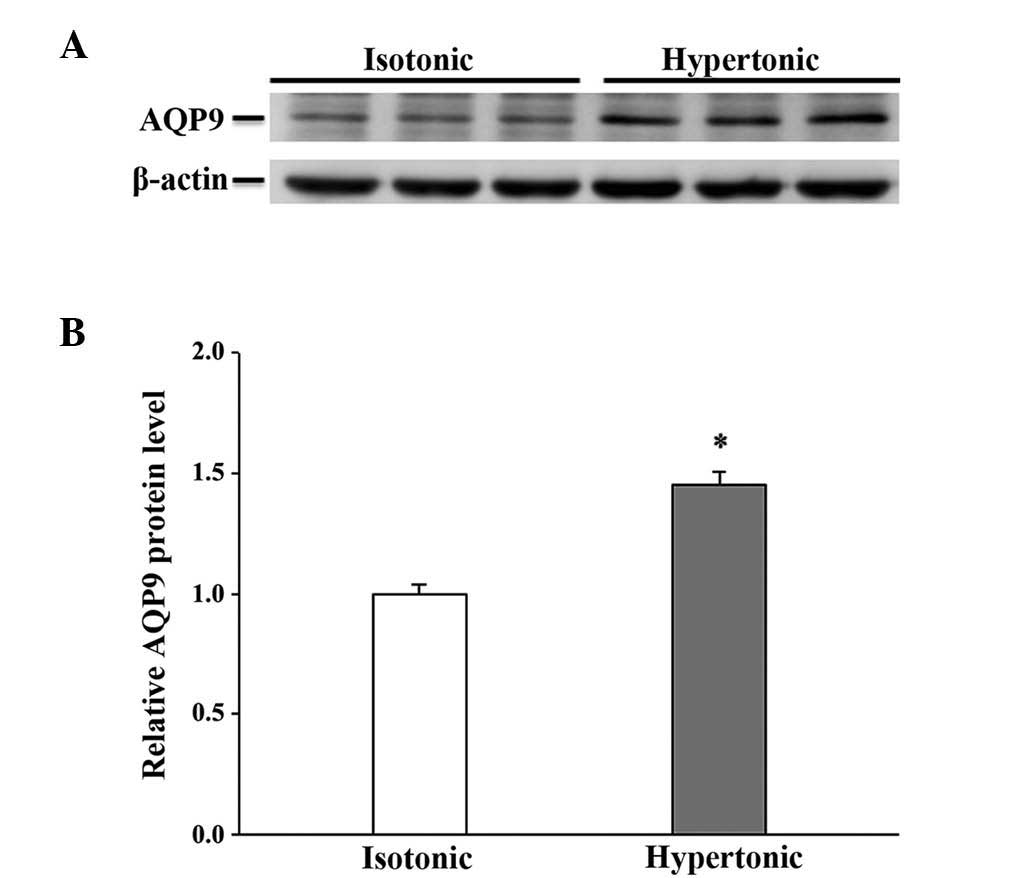
shown). Osmotic stress did not affect the mRNA abundance of AQPs 9
and 11 (data not shown), but a significant increase in the
expression level of the 2 proteins was observed in the cells
exposed to hypertonic medium for 16 h (Figs. 4 and 5), which was not observed following a 4-h
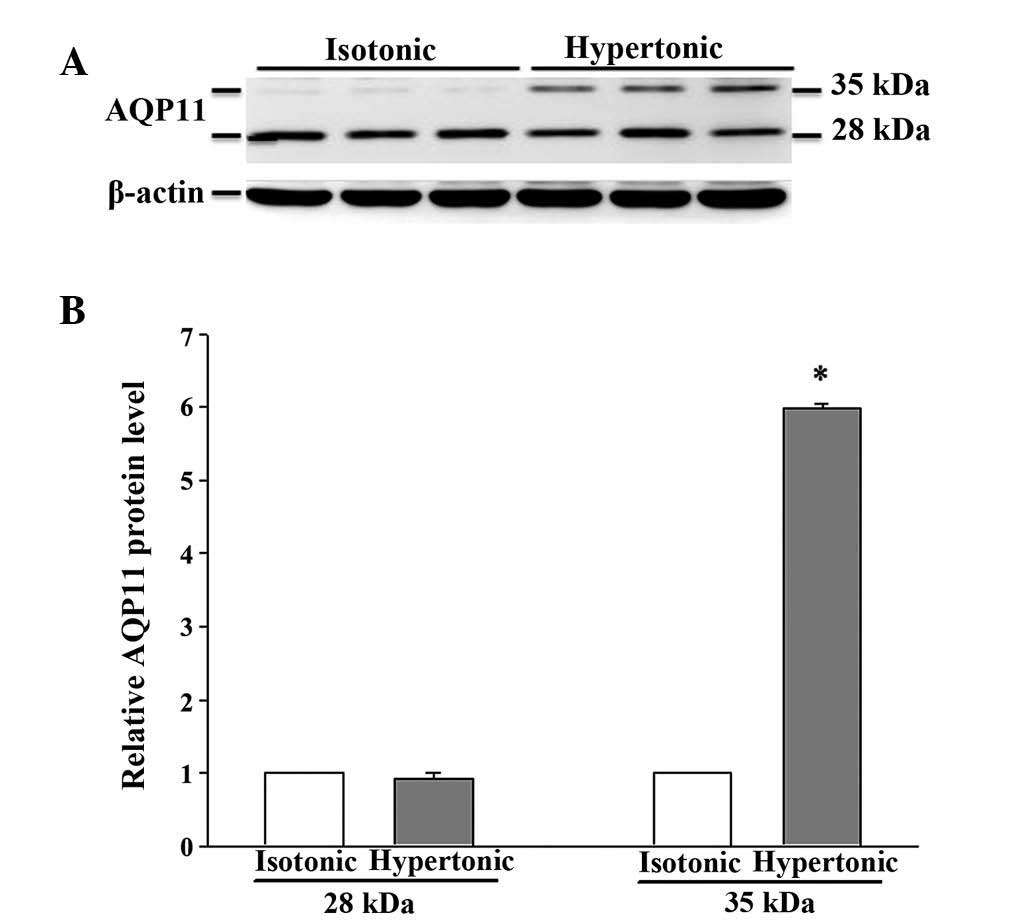
exposure (data not shown). Notably, the anti-AQP11 polyclonal
antibody used in this study recognized 2 protein bands on
immunoblots: One corresponded to the non-glycosylated form (~28
kDa), whereas the other was presumed to represent the glycosylated
form (~35 kDa) (20). Under isotonic
conditions, the non-glycosylated AQP11 was markedly expressed;
however, the glycosylated form was barely detectable (Fig. 5). After 16 h of osmotic stress, the
expression of the 28-kDa AQP11 was unchanged, yet the glycosylated
protein was markedly expressed. No significant alterations in the
mRNA expression levels of AQP9 and AQP11 were observed (data not
shown).
Cell viability and AQP expression in
response to ischemia-reperfusion
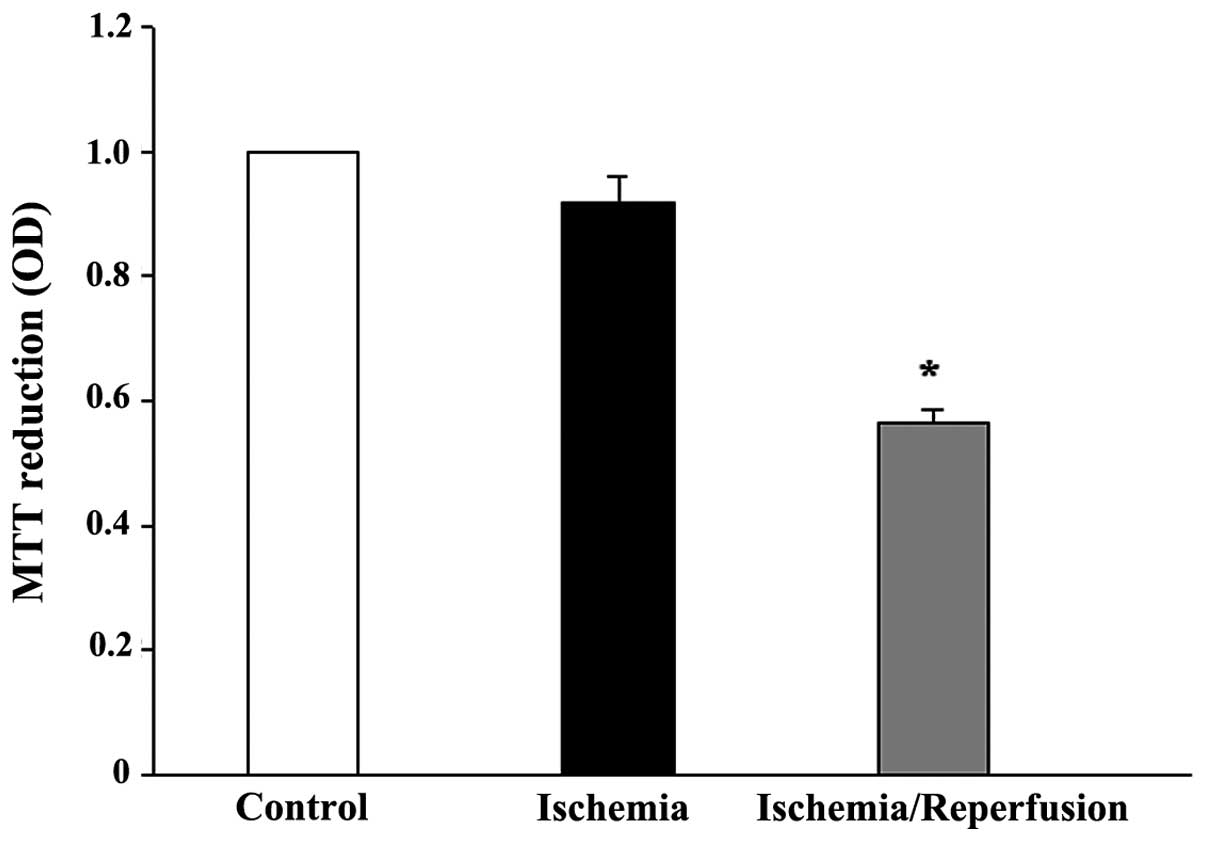
While 8 h of ischemia exposure did not affect the
cell viability (Fig. 6), it did
induce a cell morphological change from a cuboidal shape to a
smaller, thinner and loose cell-cell boundary appearance (data not
shown). Subsequent reperfusion markedly reduced the cell viability,
but restored the cell morphology. The mRNA expression levels of all
the tested AQPs (AQP1, 3, 4, 7, 8, 9 and 11) were unaffected by
ischemia alone or ischemia/reperfusion (data not shown). However,
the protein expression of AQP8 and AQP11 was significantly altered
(P<0.05 vs. control). AQP8 was downregulated by ischemia, but
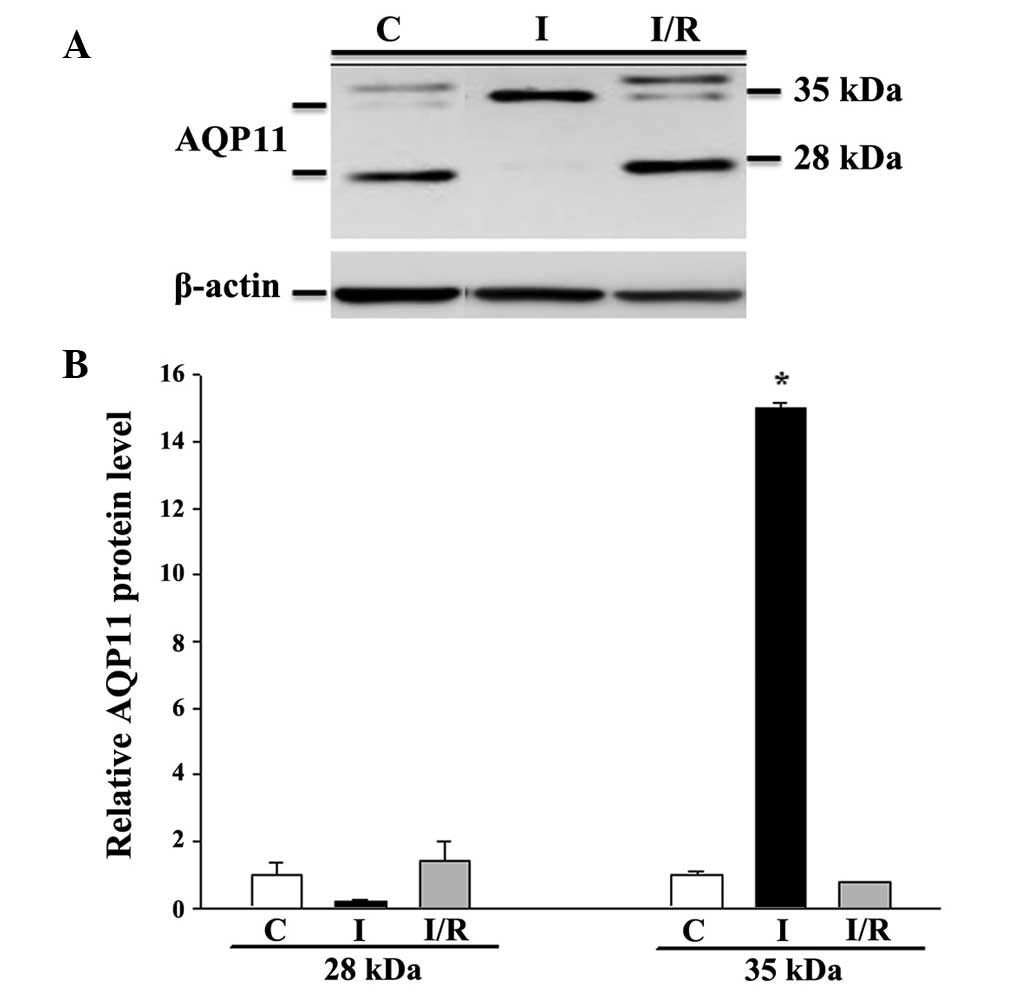
returned to the control level following reperfusion (Fig. 7). Similarly, moderate expression of
non-glycosylated AQP11 (28 kDa) and faint expression of the
glycosylated protein (35 kDa) were observed under control
conditions (Fig. 8). Ischemia alone
eradicated the expression of the 28-kDa AQP11 protein, but induced
the pronounced expression of the 35-kDa form. Following
reperfusion, this ischemia-altered expression was restored to the
normal profile of AQP11 expression.
Discussion
To the best of our knowledge, the present study
demonstrated for the first time the differential AQP expression
profiles in intestinal epithelial cells that were exposed to
environmental stress.
Intestinal epithelial cells, and similarly renal
epithelial cells, are exposed to extreme osmolar environments.
Osmotic stress may exert detrimental effects on cells, triggering
adaptation mechanisms to re-establish homeostasis through changes
to cell structure and function (21,22). In
response to osmotic stress, a limited number of genes, including
the AQP genes, are upregulated, despite an overall decrease in DNA
synthesis, RNA transcription and protein synthesis (23–25).
Previous studies have documented the enhanced expression of AQPs 1,
2, 3, 4, 5 and 9 in various, osmotically-disturbed cell types
(26). In the present study, the
protein expression of AQP1, 9 and 11 was increased after 16 h, but
not 4 h, of hypertonic exposure, suggesting that the elevated
protein expression is a delayed response. Altered protein
expression may be correlated with the observed cell shrinkage and
reduced cell viability after 16 h of osmotic challenge, although
the mechanisms involved are currently unknown.
In the intestinal tract, AQP1 is expressed in
endothelia and lacteals (9), but has
not previously been reported to exist in epithelial cells. In the
current study, hypertonic stress increased AQP1 mRNA and AQP1
protein abundance, suggesting that activation occurs at the
transcriptional and translational levels. Furthermore, upregulated
AQP1 protein may be partially caused by the hypertonicity-induced
increase in protein stability; a previous study has demonstrated
that hypertonic stress decreases AQP1 ubiquitination and increases
protein stability, thereby contributing to overall protein
induction (27). An increase in
protein stability may also contribute to the increased AQP9 and 11
protein expression, considering their unchanged mRNA level in
response to hyperosmotic stress. This increased protein expression
may also be due to a hypertonicity-induced increase in mRNA
stability.
AQP9, in addition to its function as a water
channel, is also permeable to lactate and a wide variety of neutral
solutes, including β-hydroxybutyrate, glycerol, carbamides,
purines, pyrimidines, urea, mannitol and sorbitol (28). In the present study, the upregulation
of AQP9 protein in cells maintained in mannitol-containing medium
for 16 h may suggest an involvement of AQP9 in the transmembrane
transport of mannitol. However, the transport of mannitol was not
measured in this experiment.
Glycosylated proteins are an established focus of
study, and have been suggested to be involved in protein folding
and stability, the transport of proteins through secretory
pathways, molecular recognition processes and cell-cell interaction
(29). A previous study reported
that glycosylation increases the thermostability of human AQP10
(30); the functional significance
for the expression of glycosylated AQP11 (35 kDa) after 16 h of
osmotic challenge thus requires greater investigation.
The intestine is the most sensitive to
ischemia/reperfusion injury amongst the internal organs (31). Despite the reduced viability in
ischemia/reperfusion-treated cells, the mRNA expression of all the
tested AQPs was unaffected by ischemia alone or
ischemia/reperfusion, and only the expression of AQP8 and AQP11
proteins was altered in the present study. AQP8 was substantially
downregulated in an ischemic state and returned to the control
level following reperfusion, indicating that this is a reversible
response. Similarly, ischemia alone induced the pronounced
expression of glycosylated AQP11 protein, revealed alongside a
comparative absence of the non-glycosylated form; this expression
pattern was completely restored following reperfusion, indicating
that this is also a reversible response.
In conclusion, although the functional significance
for the altered expression of AQPs is currently unresolved, the
observations presented in the current study may indicate a role of
AQPs in the adjustment of the intestinal epithelia in response to
stress.
Acknowledgements
The present study was supported by a grant from
Kaohsiung Armed Forces General Hospital Zuoying Branch, Taiwan (no.
ZAFGH 101-09).
References
|
1
|
Loo DD, Zeuthen T, Chandy G and Wright EM:
Cotransport of water by the Na+/glucose cotransporter.
Proc Natl Acad Sci USA. 93:13367–13370. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ma T and Verkman AS: Aquaporin water
channels in gastrointestinal physiology. J Physiol. 517:317–326.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Masyuk AI, Marinelli RA and LaRusso NF:
Water transport by epithelia of the digestive tract.
Gastroenterology. 122:545–562. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Agre P, King LS, Yasui M, Guggino WB,
Ottersen OP, Fujiyoshi Y, Engel A and Nielsen S: Aquaporin water
channels - from atomic structure to clinical medicine. J Physiol.
542:3–16. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nielsen J, Kwon TH, Christensen BM,
Frøkiaer J and Nielsen S: Dysregulation of renal aquaporins and
epithelial sodium channel in lithium-induced nephrogenic diabetes
insipidus. Semin Nephrol. 28:227–244. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nielsen S, Frøkiaer J, Marples D, Kwon TH,
Agre P and Knepper MA: Aquaporins in the kidney: From molecules to
medicine. Physiol Rev. 82:205–244. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rojek A, Praetorius J, Frøkiaer J, Nielsen
S and Fenton RA: A current view of the mammalian aquaglyceroporins.
Annu Rev Physiol. 70:301–327. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kwon TH, Nielsen J, Møller HB, Fenton RA,
Nielsen S and Frøkiær J: Aquaporins in the kidney. Aquaporins.
Handbook of Experimental Pharmacology. Beitz E: 190:(Berlin,
Heidelberg). Springer-Verlag. 95–132. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Marinelli RA, Tietz PS, Pham LD, Rueckert
L, Agre P and LaRusso NF: Secretin induces the apical insertion of
aquaporin-1 water channels in rat cholangiocytes. Am J Physiol.
276:G280–G286. 1999.PubMed/NCBI
|
|
10
|
Matsuzaki T, Tajika Y, Ablimit A, Aoki T,
Hagiwara H and Takata K: Aquaporins in the digestive system. Med
Electron Microsc. 37:71–80. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Laforenza U, Gastaldi G, Grazioli M, Cova
E, Tritto S, Faelli A, Calamita G and Ventura U: Expression and
immunolocalization of aquaporin-7 in rat gastrointestinal tract.
Biol Cell. 97:605–613. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hatakeyama S, Yoshida Y, Tani T, Koyama Y,
Nihei K, Ohshiro K, Kamiie JI, Yaoita E, Suda T, Hatakeyama K and
Yamamoto T: Cloning of a new aquaporin (AQP10) abundantly expressed
in duodenum and jejunum. Biochem Biophys Res Commun. 287:814–819.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ishibashi K, Morinaga T, Kuwahara M,
Sasaki S and Imai M: Cloning and identification of a new member of
water channel (AQP10) as an aquaglyceroporin. Biochim Biophys Acta.
1576:335–340. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Laforenza U: Water channel proteins in the
gastrointestinal tract. Mol Aspects Med. 33:642–650. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang KS, Ma T, Filiz F, Verkman AS and
Bastidas JA: Colon water transport in transgenic mice lacking
aquaporin-4 water channels. Am J Physiol Gastrointest Liver
Physiol. 279:G463–G470. 2000.PubMed/NCBI
|
|
16
|
Yang B, Song Y, Zhao D and Verkman AS:
Phenotype analysis of aquaporin-8 null mice. Am J Physiol Cell
Physiol. 288:C1161–C1170. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kwon TH, Hager H, Nejsum LN, Andersen ML,
Frøkiaer J and Nielsen S: Physiology and pathophysiology of renal
aquaporins. Semin Nephrol. 21:231–238. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hara-Chikuma M and Verkman AS: Roles of
aquaporin-3 in the epidermis. J Invest Dermatol. 128:2145–2151.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Umenishi F and Schrier RW: Identification
and characterization of a novel hypertonicity-responsive element in
the human aquaporin-1 gene. Biochem Biophys Res Commun.
292:771–775. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
García F, Kierbel A, Larocca MC, Gradilone
SA, Splinter P, LaRusso NF and Marinelli RA: The water channel
aquaporin-8 is mainly intracellular in rat hepatocytes, and its
plasma membrane insertion is stimulated by cyclic AMP. J Biol Chem.
276:12147–12152. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kültz D and Burg M: Evolution of osmotic
stress signaling via MAP kinase cascades. J Exp Biol.
201:3015–3021. 1998.PubMed/NCBI
|
|
22
|
Schwartz LM and Osborne BA: Programmed
cell death, apoptosis and killer genes. Immunol Today. 14:582–590.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cohen DM, Wasserman JC and Gullans SR:
Immediate early gene and HSP70 expression in hyperosmotic stress in
MDCK cells. Am J Physiol. 261:C594–C601. 1991.PubMed/NCBI
|
|
24
|
Burg MB, Kwon ED and Kültz D: Osmotic
regulation of gene expression. FASEB J. 10:1598–1606.
1996.PubMed/NCBI
|
|
25
|
Blomberg A: Osmoresponsive proteins and
functional assessment strategies in Saccharomyces cerevisiae.
Electrophoresis. 18:1429–1440. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Burg MB, Ferraris JD and Dmitrieva NI:
Cellular response to hyperosmotic stresses. Physiol Rev.
87:1441–1474. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Leitch V, Agre P and King LS: Altered
ubiquitination and stability of aquaporin-1 in hypertonic stress.
Proc Natl Acad Sci USA. 98:2894–2898. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tsukaguchi H, Shayakul C, Berger UV,
Mackenzie B, Devidas S, Guggino WB, van Hoek AN and Hediger MA:
Molecular characterization of a broad selectivity neutral solute
channel. J Biol Chem. 273:24737–24743. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sato Y and Endo T: Alteration of brain
glycoproteins during aging. Geriatr Gerontol Int. 10(Suppl 1):
S32–S40. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Öberg F, Sjöhamn J, Fischer G, Moberg A,
Pedersen A, Neutze R and Hedfalk K: Glycosylation increases the
thermostability of human aquaporin 10 protein. J Biol Chem.
286:31915–31923. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sasaki M and Joh T: Oxidative stress and
ischemia-reperfusion injury in gastrointestinal tract and
antioxidant, protective agents. J Clin Biochem Nutr. 40:1–12. 2007.
View Article : Google Scholar : PubMed/NCBI
|