Introduction
Transplanted neural stem cells (NSCs) are involved
in the repair of central nervous system (CNS) disease,
predominantly by replacing damaged cells, secreting neurotrophic
factors and participating in immune regulation (1–3).
Therapeutic efficacy is associated with the quantity and quality of
grafted NSCs and is also largely dependent on the implantation
approach. Intracerebral transplantation can directly deliver
exogenous NSCs through the blood-brain barrier into the
immune-privileged brain. However, this approach requires a hole to
be drilled into skull, which is invasive and associated with risks.
Previous studies have demonstrated that several methods may be
utilized to reduce brain injuries during penetration of the skull,
in particular microtransplantation was found to improve the
function of grafts (4,5); however, the mechanism remains unclear.
Furthermore, inflammation secondary to brain injury during skull
penetration also influences the survival and function of NSCs
(4–6), although fewer studies focus on this
issue.
Previous studies have elucidated the relationship
between NSCs and immune cells, and identified that the immune
system has a complex role in CNS degeneration and reconstruction
(6,7). For example, while infiltrated immune
cells are considered to be pathological, some studies have
demonstrated that they have protective and healing properties in
the CNS (8,9). However, inflammation involving multiple
activated immune cells may aggravate brain injuries and constitute
a temporary risk to NSC survival.
In the present study, the effects of transplantation
approaches on brain injuries and secondary inflammation were
investigated. Two skull penetration models in adult mice were
constructed where mice received either syngeneic NSCs or PBS and
brain injuries and secondary inflammation from 1 to 28 days after
implantation was observed.
Materials and methods
Animals
A total of 252 healthy adult C57BL/6 (B6) mice (126
male and 126 female; age, 10–12 weeks), weighing 18–28 g (Vital
River Laboratories, Beijing, China) were housed in a temperature-
and humidity-controlled room (temperature at 21±2°C and relative
humidity at 50±5%) and maintained on a 12 h light/dark schedule
with free access to food and water. All experimental procedures
were approved by the Animal Care and Use Committee of Third
Military Medical University (Chongqing, China) and were in
compliance with the Guide for the Care and Use of Laboratory
Animals published by the National Institutes of Health.
Cells
B6 NSCs were isolated from the forebrain region of
13.5-day-old murine embryos and collected by several cycles of
0.05% trypsin-EDTA (Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) digestion. Cells were plated at a density of
105-106 cells/ml and cultured in neural stem
cell culture medium (NSCcm) which was composed of DMEM/F12 (1:1)
supplemented with 2% B27, 20 ng/ml basic fibroblast growth factor
(bFGF) and 20 ng/ml epidermal growth factor (EGF) (all Invitrogen;
Thermo Fisher Scientific, Inc.). For identification, NSCs were
digested with Accutase (Invitrogen; Thermo Fisher Scientific, Inc.)
and seeded on glass coverslips coated with poly-l-lysine
(Sigma-Aldrich; Merck Millipore, Darmstadt, Germany) in a 24-well
plate (1×105 cells per well) in NSCcm for 2–3 days. The
self-renewal potential of NSCs was determined by morphological
assessment and immunofluorescence staining with the NSC-specific
markers Nestin and Sox2. For transplantation, NSCs were digested
with accutase and washed with PBS. Cells were counted by trypan
blue (Sigma-Aldrich; Merck Millipore) exclusion in a haemocytometer
and the density of single cell suspension was adjusted accordingly.
Cells were maintained on ice.
Transplantation experiment
Mice were anesthetized with 3.6% chloral hydrate (10
ml/kg; intraperitoneal injection; Sigma-Aldrich; Merck Millipore)
and mounted using stereotaxic apparatus (Stoelting, Wood Dale, IL,
USA). After shaving and cleaning the skin, a midline scalp incision
was made using an 11 Swann scalpel and the parietal bone was
exposed.
For the drilling method, an appropriate puncture
point was selected on the left hemisphere, and the skull was
penetrated by drilling a hole with a sterile drill (Slite P-500-3A;
Ningbo Rising Trading Co., Ningbo, China) using a steel screw
(diameter, 1 mm) at the speed of 12,000 rpm. For skull penetration
with a syringe needle, an appropriate puncture point was selected
on the right hemisphere, and the skull was penetrated with a
sterile 25-µl 22S Hamilton syringe. The syringe was withdrawn once
through the skull.
Mice were randomly assigned to three groups
(n=84/group): Group A, receiving NSCs in the left hemisphere and
the same volume of PBS in the right hemisphere; group B, receiving
NSCs in the right hemisphere and PBS in the left hemisphere; and
group C, receiving the equal amount of NSCs in the two hemispheres.
Cell suspension or PBS was injected into the motor cortex (4.5 mm
anterior to the lambda suture, 1.0 mm lateral to the middle line
and 2.5 mm under the dura) using different sterile 25-µl 22S
Hamilton syringes. Each site received 5 µl of cell suspension
containing 5×105 cells or PBS at a speed of 0.5 µl/min.
Following injection for ~5 min, the syringe was slowly withdrawn.
At 1, 2, 3, 7, 14, 21 and 28 days after transplantation, mice were
sacrificed using a lethal dose of 10 ml/kg chloral hydrate and
their brain tissues were harvested for morphological and
molecular-biological analysis.
Morphological analysis
Brain tissues were postfixed in 4% paraformaldehyde
in 0.1 M PBS (pH 7.4) at 4°C overnight. Tissues were sectioned (8
µm) on a cryostat (Leica CM1950; Leica Microsystems, Inc., Buffalo
Grove, IL, USA) and mounted to an adhesion microscope slide.
Pathological features were assessed using hematoxylin and eosin
staining and viewed under a light microscope (DM2000; Leica
Microsystems, Inc.) at ×200 magnification.
Immunofluorescence analysis
Cultured cells and brain slices were blocked using
10% bovine serum albumin and 0.3% Triton X-100 for 1 h, and
subsequently incubated overnight at 4°C with primary antibodies
(Table I). Following washing in PBS,
cells and slices were incubated for 1–2 h at room temperature with
secondary antibodies (Table I).
Following several washes, the nuclei were stained using DAPI
Fluoromount-G (SouthernBiotech, Birmingham, AL, USA) and coverslips
were mounted onto glass slides and analyzed using a fluorescent
microscope (DM3000) or a confocal laser scanning microscope (TCS
SP5 II; both Leica Microsystems, Inc.). The number of positive
cells was manually counted at ×200 magnification directly under the
microscope and adjusted using image analysis software (Image Pro
Plus 5.0; Media Cybernetics, Inc., Rockville, MD, USA). The ratio
of positive cells was calculated as follows: Ratio of positive
cells=(number of positive cells/total number of cells) × 100%.
 | Table I.Antibodies used in the present
study. |
Table I.
Antibodies used in the present
study.
| Specificity | Host | Clone | Dilution | Detection | Catalogue no. |
|---|
| Nestina | Mouse | Rat-401 | 1:1,000 | NSCs | MAB353 |
| Sox2b | Goat | Y-17 | 1:200 | NSCs | sc-17320 |
| CD3c | Rabbit | Polyclonal | 1:200 | T cells | ab5690 |
| CD11bc | Rat | Monoclonal | 1:200 |
Microglia/Macrophages | ab8878 |
| CD19d | Rat | 1D3 | 1:200 | B cells | LS-C107165 |
| Mouse
IgGe | Donkey | Polyclonal | 1:1,000 | Nestin | A-21203 |
| Rat IgGe | Donkey | Polyclonal | 1:1,000 | CD11b/CD19 | A-21209 |
| Rabbit
IgGe | Donkey | Polyclonal | 1:1,000 | CD3 | A-21207 |
| Goat IgGe | Donkey | Polyclonal | 1:1,000 | Sox2 | A-11055 |
ELISA
Brain tissues were respectively isolated from the
two hemispheres and weighed. Protein was extracted from these brain
tissues using radioimmunoprecipitation assay buffer (Sigma-Aldrich;
Merck Millipore) supplemented with protease and phosphatase
inhibitors (Fermentas; Thermo Fisher Scientific, Inc.). Protein
concentration was determined using BCA assay (Thermo Fisher
Scientific, Inc.). Mouse brain-derived neurotrophic factor (BDNF)
glial cell line-derived neurotrophic factor (GDNF), neurotrophin-3
(NT-3), interleukin-6 (IL-6), interleukin (IL)-10 and tumor
necrosis factor alpha (TNF-α) were detected using ELISA kits. BDNF
ELISA kit was purchased from Promega Corp., (Madison, WI, USA).
GDNF ELISA kit was purchased from Santa Cruz Biotechnology Inc.,
(Dallas, TX, USA). NT-3 ELISA kit was purchased from Abcam,
(Cambridge, MA, USA) and IL-6, IL-10 and TNF-α ELISA kits were
purchased from Sigma-Aldrich (Merck Millipore). ELISA was carried
out in duplicate, according to the manufacturers' protocols.
Statistical analysis
The SPSS 17.0 statistical software (SPSS, Inc.,
Chicago, IL, USA) package was used for statistical analysis. Data
were presented as mean ± standard deviation. Student's t-test and
one-way analysis of variance were used to determine statistical
significance. P<0.05 was considered to indicate a statistically
significant difference.
Results
Identification of NSCs
B6 NSCs were isolated from the forebrain region of
13.5-day-old murine embryos and had self-renewal potential. These
cells formed neurospheres that grew in culture medium in a state of
suspension. The results of immunofluorescence staining indicated
that these cells were strongly positive for Nestin and Sox2 (data
not shown).
General information and morphological
analysis
Macroscopic inspection showed obvious injuries and
hemorrhage in the left hemisphere during the early stage of
post-implantation for all the groups (data not shown). Wounds
observed were irregular, unstable and varied within the drilling
method group; whereas in the right hemisphere, wounds were small
and exhibited a similar appearance to one another following syringe
needle penetration. Histological examination of all groups
indicated that a small amount of bleeding was observed in the first
two days in the right hemisphere, which underwent syringe
penetration. In contrast, the structure of the left hemisphere,
which underwent drill penetration, was broken and exhibited
extensive necrosis, infiltrated immune cells and hemorrhage. After
one week, scar tissue formation was observed in all groups, and
only a small number of neural cells were detected in the left
hemisphere, whereas the contralateral cells were well-arranged.
After 28 days, neural cells were rarely observed in the injured
area, which was filled with massive scar tissue in the left
hemisphere. In addition, brain injuries in the left hemisphere of
mice receiving PBS treatment were notably worse than those
receiving NSC transplantation.
Dynamic changes of infiltrated immune
cells
To measure the extent of inflammation secondary to
brain injury after implantation, the distribution of infiltrated
immune cells, including microglia/macrophages [cluster of
differentiation (CD)11b], T cells (CD3) and B cells (CD19) was
observed. The results of group C in the left and right hemispheres
are presented in Fig. 1A and B,
respectively. The number of immune cells was elevated in the left
hemisphere; however, these cells were rarely detectable in the
right hemisphere in all groups. As demonstrated in Fig. 1C-E, in group A, the three types of
immune cells (CD11b, CD3 and CD19, respectively) increased rapidly
in the first three days and remained at a high level for 2–3 weeks.
In group B, the difference between the two hemispheres was the most
obvious and the number of immune cells in the left hemisphere was
the highest of the three groups. Only a small number of infiltrated
immune cells were detected in group C, the difference between the
two hemispheres in this group was statistically significant
(P<0.05). In addition, the number of immune cells in the part of
the hemisphere receiving NSC transplantation was lower than that in
the same side receiving PBS treatment (P<0.05).
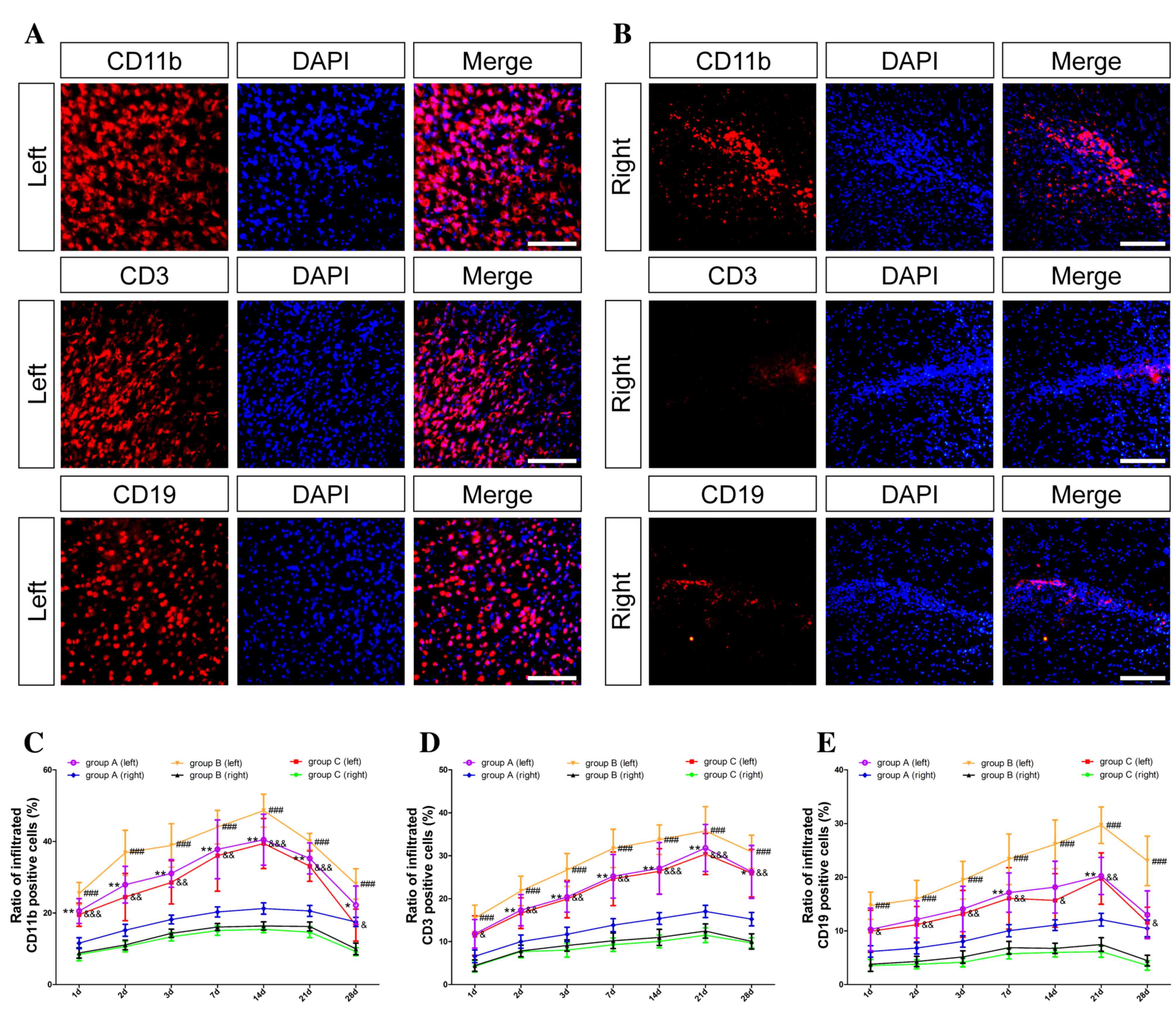 | Figure 1.Dynamic changes of infiltrated immune
cells. Inflammation secondary to brain injury during skull
penetration was determined by immunofluorescence staining. Nuclei
were stained with DAPI (blue); microglia/macrophages were
identified by anti-CD11b (red); T cells were identified by anti-CD3
(red); and B cells were identified by anti-CD19 (red). In group C,
(A) massive infiltrated immune cells, including CD11b-, CD3- and
CD19-positive cells, were observed in the left hemisphere; (B)
however, only a few immune cells were seen in the right hemisphere
at 3 days post-transplantation. (C-E) Statistical analysis of the
infiltrated CD11b-, CD3- and CD19-positive cells between the two
hemispheres among the three groups are presented, respectively.
Data are shown as mean ± standard deviation (n=6, *P<0.05,
**P<0.01 vs. group A (right); ###P<0.001 vs. group
B (right); &P<0.05,
&&P<0.01,
&&&P<0.001 vs. group C (right),
respectively). Scale bar, 100 µm. DAPI,
4′,6-diamidino-2-phenylindole; CD, cluster of differentiation 3;
group A, NSCs in the left hemisphere and PBS in the right; group B,
NSCs in the right hemisphere and PBS in the left; and group C,
equal NSCs in both hemispheres. |
Immune cells were predominantly distributed around
the broken structure of the skull and were positively correlated
with the degree of injury. An increase in the number of immune
cells observed suggested that inflammation secondary to skull
penetration using a drill was significantly more severe when
compared with skull penetration using a syringe needle (P<0.05).
The number of microglia and macrophages increased in the first week
but the peak remained only for a short time (Fig. 1C). T cells were noticeable at the
early stage following brain injury; however, the level of T cells
did not change significantly throughout the study period (Fig. 1D). Compared with the other immune
cells, the number of B cells was lower and the peak appeared later
in three weeks (P<0.001; Fig.
1E). In addition, the number of immune cells varied within the
left hemisphere treatment group among all the groups (groups
A-C).
Dynamic changes in levels of
neurotrophic and immunomodulatory factors
To investigate the effects of transplantation
approaches on hosts and grafts, the levels of neurotrophic and
immunomodulatory factors in the cerebral cortex were detected
(Fig. 2). ELISA results revealed
that BDNF, GDNF and NT-3 concentrations in the right hemisphere of
group B and C increased significantly (P<0.05; Fig. 2A-C, respectively), whereas there was
no significant difference in BDNF, GDNF and NT-3 concentrations
between the two hemispheres in group A. Levels of neurotrophic
factors in the NSC transplantation hemispheres were higher than
those in the PBS treatment side (P<0.05; Fig. 2A-C).
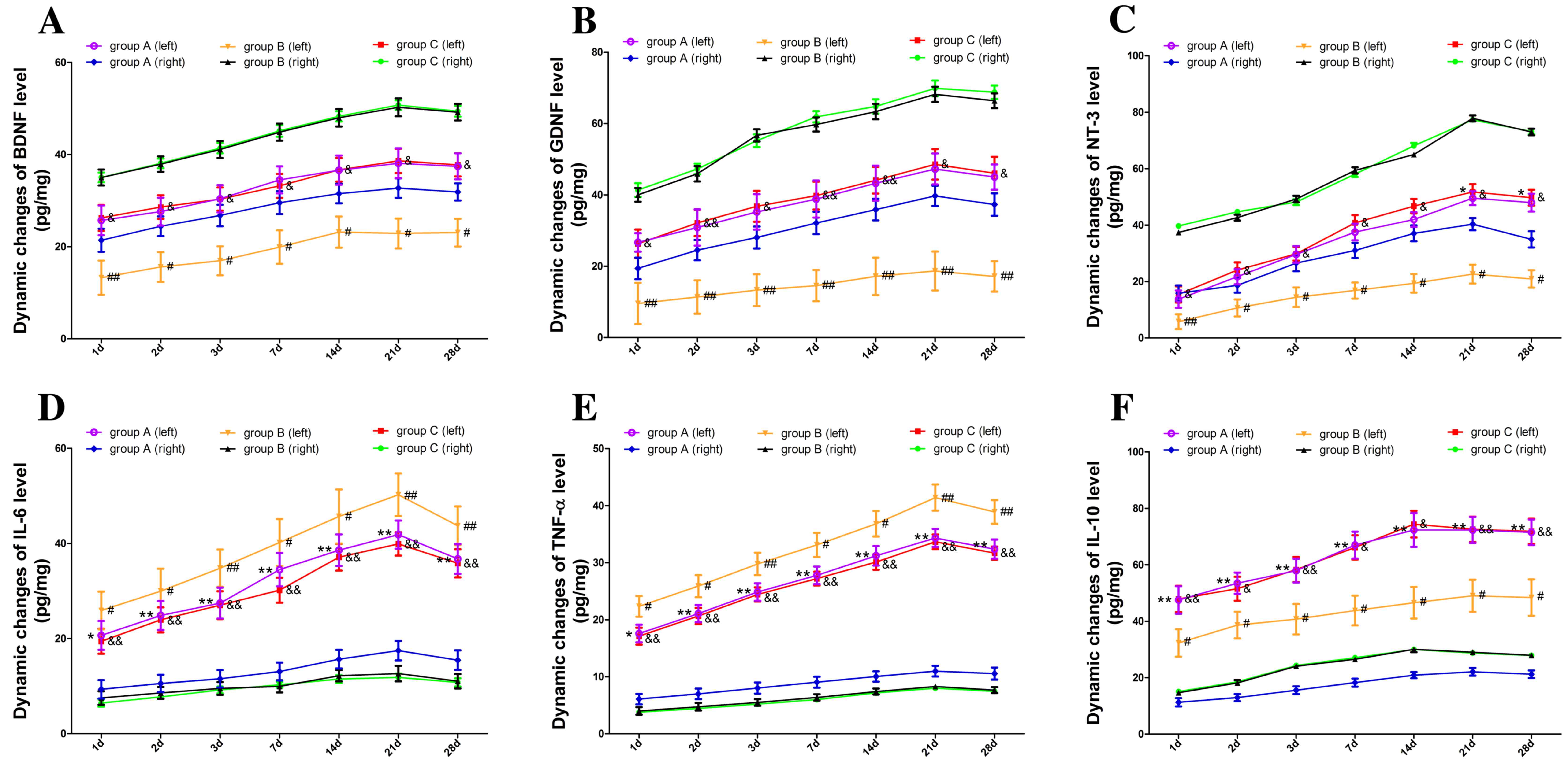 | Figure 2.Dynamic changes in levels of
neurotrophic and immunomodulatory factors. Levels of neurotrophic
and immunomodulatory factors were measured by ELISA assay. (A-C)
BDNF, GDNF and NT-3 concentrations in the right hemisphere of group
B and C increased significantly, whereas there was no significant
difference in BDNF, GDNF and NT-3 concentrations between the two
hemispheres in group A. (D and E) Levels of pro-inflammatory
cytokines (IL-6 and TNF-α) in the PBS-injected hemisphere were
higher than those in the NSC transplantation side. (F) Level of
IL-10 anti-inflammatory cytokine present in the PBS-injected
hemisphere was lower than the NSC transplantation hemisphere. Data
are shown as mean ± standard deviation from three independent
experiments (*P<0.05, **P<0.01 vs. group A (right);
#P<0.05, ##P<0.01 vs. group B (right);
&P<0.05, &&P<0.01 vs. group
C (right), respectively). ELISA, enzyme-linked immunosorbent assay;
NSC, neural stem cells; BDNF, brain-derived neurotrophic factor;
GDNF, glial cell line-derived neurotrophic factor; NT-3,
neurotrophin-3; IL-6, interleukin-6; IL-10, interleukin-10; TNF-α,
tumor necrosis factor alpha; group A, NSCs in the left hemisphere
and PBS in the right; group B, NSCs in the right hemisphere and PBS
in the left; and group C, equal NSCs in both hemispheres. |
Furthermore, immunomodulatory factors including pro-
and anti-inflammatory cytokines increased significantly in the left
hemisphere in response to the infiltrated immune cells among all
the groups (P<0.05). The levels of pro-inflammatory cytokines
(IL-6 and TNF-α) in the PBS injected hemisphere were significantly
higher than those in the NSC transplantation side (P<0.05;
Fig. 2D and E, respectively).
However, the levels of anti-inflammatory cytokine IL-10 in the PBS
injected hemisphere were significantly lower than that in the NSC
transplantation side (P<0.05; Fig.
2F).
Discussion
NSC implantation is a promising strategy for
neuronal functional recovery in the treatment of CNS diseases
(10,11). Although intracerebral transplantation
is useful for efficiently delivering NSCs directly, serious
limitations to this approach are evident as this form of
transplantation may induce brain injury and secondary inflammation.
Previous studies have reported that microtransplantation can
improve the function of grafts (4,5);
however, the mechanism of how this occurs remains to be elucidated.
Few studies have demonstrated that inflammation secondary to brain
injuries during skull penetration may also effect the survival and
function of NSCs (4–6). In the present study, the effects of two
types of intracerebral NSC transplantation approaches on recipients
and grafts were compared.
NSCs from B6 embryos that may avoid immune rejection
of adult syngeneic mice after transplantation were obtained to
ensure the immune cells expressed were predominantly due to
secondary inflammation. Microglia and macrophages were the key
contributors in the immune response observed, producing cytokines
to attract other immune cells and initiating a pro-inflammatory
response (12,13). T and B cell-mediated adaptive
immunity also has a complex role in the reconstruction of CNS
(6,14,15). The
dynamic changes exhibited by immune cells and cytokines revealed
that syringe needle skull penetration may reduce inflammation
secondary to brain injuries during skull penetration.
Notably, the present study identified that grafted
NSCs were able to decrease infiltrated immune cells when compared
with PBS-treated groups. The variability in the number of immune
cells observed within the group in the left hemisphere indicated
that the drill method was less stable. Furthermore, the levels of
IL-6, IL-10 and TNF-α in the brains treated with NSCs compared with
PBS-treated brains suggested that NSCs may modulate secondary
inflammation. In addition, the elevated levels of BDNF, GDNF and
NT-3 in the right hemisphere after NSC transplantation also
suggested that syringe needle skull penetration was conducive to
the grafts and the recipients when compared with drill
penetration.
Although NSC transplantation has been widely used in
clinical medicine, this method still exhibits limitations (3,16),
including brain injuries and secondary inflammation. Previous
studies have shown that the immune system is associated with the
effects of NSCs (6,17). Under mild inflammatory conditions,
immune cells may promote neurogenesis (6,18).
However, under severe inflammatory conditions, immune cells can
transform their polarization from anti- to pro-inflammatory, which
may result in detrimental effects on neural reconstruction
(19,20). Further research should address the
association between NSCs and immune cells.
In conclusion, the present study indicated that
syringe needle skull penetration is an improved method that results
in reduced brain injury and secondary inflammation for
intracerebral NSC transplantation, when compared with drill
penetration. Furthermore, findings from the present study suggest
that NSCs have the potential to modulate inflammation secondary to
brain injuries.
Acknowledgements
The present study was supported by funding from
Military Twelfth Five-Year Key Sci-Tech Research Projects (grant
no. BWS12J010).
References
|
1
|
Darsalia V, Allison SJ, Cusulin C, Monni
E, Kuzdas D, Kallur T, Lindvall O and Kokaia Z: Cell number and
timing of transplantation determine survival of human neural stem
cell grafts in stroke-damaged rat brain. J Cereb Blood Flow Metab.
31:235–242. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
English D, Sharma NK, Sharma K and Anand
A: Neural stem cells-trends and advances. J Cell Biochem.
114:764–772. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gage FH and Temple S: Neural stem cells:
Generating and regenerating the brain. Neuron. 80:588–601. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nikkhah G, Cunningham MG, Jödicke A,
Knappe U and Björklund A: Improved graft survival and striatal
reinnervation by microtransplantation of fetal nigral cell
suspensions in the rat Parkinson model. Brain Res. 633:133–143.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jiang W, Büchele F, Papazoglou A, Döbrössy
M and Nikkhah G: Multitract microtransplantation increases the
yield of DARPP-32-positive embryonic striatal cells in a rodent
model of Huntington's disease. Cell Transplant. 20:1515–1527. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kokaia Z, Martino G, Schwartz M and
Lindvall O: Cross-talk between neural stem cells and immune cells:
The key to better brain repair? Nat Neurosci. 15:1078–1087. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Capetian P, Döbrössy M, Winkler C, Prinz M
and Nikkhah G: To be or not to be accepted: The role of
immunogenicity of neural stem cells following transplantation into
the brain in animal and human studies. Semin Immunopathol.
33:619–626. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Schwartz M, Kipnis J, Rivest S and Prat A:
How do immune cells support and shape the brain in health, disease,
and aging? J Neurosci. 33:17587–17596. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Singec I, Jandial R, Crain A, Nikkhah G
and Snyder EY: The leading edge of stem cell therapeutics. Annu Rev
Med. 58:313–328. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lindvall O and Kokaia Z: Stem cells in
human neurodegenerative disorders-time for clinical translation? J
Clin Invest. 120:29–40. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Andres RH, Horie N, Slikker W, Keren-Gill
H, Zhan K, Sun G, Manley NC, Pereira MP, Sheikh LA, McMillan EL, et
al: Human neural stem cells enhance structural plasticity and
axonal transport in the ischemic brain. Brain. 134:1777–1789. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wynn TA, Chawla A and Pollard JW:
Macrophage biology in development, homeostasis and disease. Nature.
496:445–455. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Biswas SK and Mantovani A: Macrophage
plasticity and interaction with lymphocyte subsets: Cancer as a
paradigm. Nat Immunol. 11:889–896. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ekdahl CT, Claasen JH, Bonde S, Kokaia Z
and Lindvall O: Inflammation is detrimental for neurogenesis in
adult brain. Proc Nati Acad Sci USA. 100:13632–13637. 2003.
View Article : Google Scholar
|
|
15
|
Das S and Basu A: Inflammation: A new
candidate in modulating adult neurogenesis. J Neurosci Res.
86:1199–1208. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Delcroix GJ, Schiller PC, Benoit JP and
Montero-Menei CN: Adult cell therapy for brain neuronal damages and
the role of tissue engineering. Biomaterials. 31:2105–2120. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yirmiya R and Goshen I: Immune modulation
of learning, memory, neural plasticity and neurogenesis. Brain
Behav Immun. 25:181–213. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tang Y and Le W: Differential roles of M1
and M2 microglia in neurodegenerative diseases. Mol Neurobiol.
53:1181–1194. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Harry GJ: Microglia during development and
aging. Pharmacol Ther. 139:313–326. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hu X, Leak RK, Shi Y, Suenaga J, Gao Y,
Zheng P and Chen J: Microglial and macrophage polarization-new
prospects for brain repair. Nat Rev Neurol. 11:56–64. 2015.
View Article : Google Scholar : PubMed/NCBI
|














