Introduction
Atherosclerosis is one of the most important causes
of cardiovascular and cerebrovascular diseases. Among the causes of
atherosclerosis, hyperlipidemia is a major independent risk factor
(1). The search for natural
compounds capable of reducing blood fat and the study of their
mechanism of action has been the focus of research in recent years
as such compounds have significant prospective applications and
social benefits. Apigenin is a flavonoid compound distributed
extensively in vegetables and fruits grown in the temperate and
tropical zones. Its content is particularly high in celery
(2). Its content is also high in
some medicinal plants such as Semen Plantaginis and Chinese
Star Jasmine Stem. It is also distributed in some plant-derived
beverages such as tea and alcohol, and in condiments (3,4).
Research in recent years has indicated that apigenin
has many active physiological functions: i) It exerts significant
anticancer effects during the three major stages of cancer:
Initiation, promotion, and progression. Furthermore, it has
positive therapeutic effects against breast cancer, colon cancer,
and melanoma (5–7); ii) it exerts spasmolytic,
antihypertensive, and vasodilator effects (8,9); iii) it
exerts anti-inflammatory effects by inhibiting the high expression
of nitric oxide (NO) synthase by macrophages (10); iv) it exerts sedative and anxiolytic
effects by enhancing γ-aminobutyric acid neurotransmission
(11,12); and v) it exerts inhibitory effects
against multiple gram-negative bacteria (13,14).
In terms of cholesterol regulation, previous studies
have shown that the flavonoid constituent has no significant effect
on blood lipid metabolism, while apigenin plays a significant role
in regulating blood fat. It can reduce the body weight of animals
and decrease the levels of total cholesterol (TC), triglyceride
(TG) and low-density lipoprotein cholesterol (LDL-C) in the serum
of the high-fat model mouse. Therefore, three key proteins,
HMG-CoAR, LDL-R, and CYP7A1, involved in cholesterol synthesis,
absorption, and metabolism have been studied (15,16) by
measuring their expression at the level of mRNA using RT-PCR to
determine the effect of apigenin on cholesterol metabolism. In
terms of blood pressure reduction and vasodilation, research has
indicated that apigenin can dilate the contracted aortic ring of
rats caused by phenylephrine in vitro. Experiments have
shown that apigenin can inhibit the proliferation of vascular
smooth muscle cells in a dose-dependent manner within a certain
concentration range (17),
suggesting that apigenin is important in preventing cardiovascular
diseases such as hypertension and atherosclerosis.
Animal models of hyperlipidemia have been
established by means of a high-fat diet and have been used to study
intervention with apigenin to observe its effect and mechanism of
action in regulating blood fat and to understand the effective
constituents capable of regulating cholesterol metabolism with
apigenin. In terms of protective effect on endothelial cells, the
present study aimed to observe the antagonistic effect of apigenin
on hydrogen peroxide (H2O2)-induced injury
and to understand the effect of apigenin on improving the functions
of vascular endothelial cells (EA.hy926 cells). The results
indicated that the antioxidant effect of apigenin and its role in
protecting vascular endothelial cells occurred by increasing the
activity of intracellular superoxide dismutase (SOD) and the
secretion of NO. In addition, apigenin can inhibit the
proliferation of vascular smooth muscle cells in a dose-dependent
manner and may have the potential to inhibit thickening of vascular
walls (17).
The present study involved the establishment of a
mouse model of hyperlipidemia, and verification of the efficacy of
apigenin in improving hyperlipidemia, and analysis of the mechanism
of action of apigenin in reducing cholesterol content.
Additionally, the roles of apigenin in reverse cholesterol
transport, inhibition of the formation of foam cells, and resisting
oxidization and protecting vascular endothelial cells were studied.
The study provides further evidence for the role of apigenin in
regulating cholesterol metabolism, suggesting it may be used for
clinical treatment of cardiovascular and cerebrovascular
diseases.
Materials and methods
Animal source and feed
Sixty female ICR mice (class II) weighing 18–20 g
were purchased from the Laboratory Animal Center of Shandong
University. The mice were raised in a clean animal room [no. SYXK
(Inner Mongolia) 2005-0036]. Experiments began after 1 week of
adaptive feeding. The animals were given access to food and water
ad libitum during experiments. All the feed and potable
water were disinfected.
Grouping and modeling
The mice were divided into six groups: Normal
control group, high-fat animal model group, apigenin (Research
Institution of Medicinal Plants of Chinese Academy of Medical
Sciences, Beijing, China) low-dosage group, apigenin
moderate-dosage group, apigenin high-dosage group, and simvastatin
(R&D Systems, Wiesbaden, Germany) group, with 12 mice each. The
daily dosage of the positive control drug, simvastatin, was 12
mg/kg. Sodium methyl cellulose (Sigma-Aldrich, St. Louis, MO, USA)
was used as the solvent for preparation of drug suspensions. The
mice were administered apigenin or simvastatin by intragastric
injection once per day for 28 consecutive days. Mice in the control
group and model group were administered intragastric injections of
equal volumes of solvent. The method for establishing the
hyperlipidemia mouse model was by feeding mice a high-fat diet
(Biosino Bio-Technology and Science, Inc., Beijing, China). The
animals in the normal group were fed with basal feed and the
modeling continued for 28 days.
Cell culture
THP-1 cells were cultured with RPMI-1640 medium
containing 10% fetal bovine serum (FBS) (Gibco BRL, Life
Technologies Inc., Grand Island, NY, USA) containing 100 U/ml
penicillin and 100 U/ml streptomycin at 37°C in an environment
containing 5% CO2. Media were changed every 2–3 days.
Human umbilical venous endothelial cells (EA.hy926) and vascular
smooth muscle cells (A10) were cultured in low-sugar Dulbecco's
modified Eagle's medium (DMEM) containing 10% FBS, 100 U/ml
penicillin and 100 U/ml streptomycin in a constant-temperature
incubator containing 5% CO2 at 37°C.
Establishment of macrophage-derived
foam cell model
Macrophage-derived foam cells were prepared as
previously described (18). THP-1
cells were cultured in RPMI-1640 containing 10% FBS. The cells were
centrifuged at 8,000 × g for 5 min, re-suspended, counted, and
seeded in 6-well microplates at a density of 1.5×105
cells/well. The cells were divided into five groups: Normal group,
model group, apigenin high-dosage group, apigenin moderate-dosage
group, apigenin low-dosage group, and simvastatin group. Each
condition was tested in triplicate. The cells were differentiated
into macrophages by PMA induction (Sigma-Aldrich) for 48 h. The
liquid in each well was discarded. The cells were washed three
times with PBS. Serum-free RPMI-1640 culture medium was then added
to each well. The appropriate solutions were added to each group
for pre-treatment. The cells were then incubated for 24 h. The
solution was changed every 24 h. The cells were cultured for an
additional 48 h.
Determination of TC, HDL-C, and
glucose content in serum
Total serum cholesterol was measured with a
biochemical analyzer (Hitachi, Ltd., Tokyo, Japan).
Determination of blood fat and hepatic
lipid content
The content of TC, TG, HDL-C, and LDL-C in serum of
the mice in the various groups were determined in accordance with
the instructions of the blood fat assay kit (Biosino Bio-Technology
and Science, Inc.). The formula for calculating the atherosclerosis
(AS) index was: AS index = [(TC-HDL-C)/HDL-C]. Determination of the
hepatic lipid content: 0.4 g of liver tissue was weighed, placed in
a mechanical homogenizer, cut into pieces with scissors, and ground
after addition of 4 ml Folch solution. Homogenates were placed at
4°C for 24 h and occasionally subjected to vibration. Then, 2 ml of
the extracting solution was used to wash the test tube after it was
filtered through a funnel. Samples were poured onto filter paper
and 2 ml of normal saline was added. The samples were mixed well
and allowed to stand for 1.5–2 h. The samples were then centrifuged
for 10 min at 1,500 × g following stratification. The solutions at
the upper and middle levels were pipetted and discarded. Samples
were then steamed in a water bath at 75°C until the presence of a
thick, yellow jelly-like lipid appeared. Dried test tubes were
cooled down at room temperature. Solutions were then mixed
following the addition of 0.4 ml of ethanol and centrifuged for 10
min at 1,500 × g. A volume of 0.2 ml of supernatant ethanol
solution was then taken. It was agitated and mixed well.
Determination was performed following 6 min in a water-bath.
Detection of cholesterol in the
cellular supernatant and intracellular [3H]
A volume of 1 ml of cell culture medium was
aspirated from each well. Then, 7 ml of methylbenzene-Triton
scintillation solution (Millipore, Billerica, MA, USA) was added.
Plates were mixed and placed in an LS 6500 liquid scintillation
analyzer (Thermo Fisher Scientific, Waltham, MA, USA). The CPM was
calculated in the supernatant of the various groups of cells. Then,
1 ml of 0.1 mol/l NaOH was added to each well. The cells were
agitated occasionally to cause cell lysis until all adherent cells
were floating as observed under a microscope (Thermo Fisher
Scientific). The formula to calculate cholesterol outflow rate (%)
was: The cholesterol outflow rate (%) = supernatant CPM
(supernatant CPM + cell lysis buffer CPM) × 100%.
Detection of NO content in the cell
culture medium
Samples were mixed well, allowed to stand for 10 min
and centrifuged at 2,500 × g for 10 min. The clarified supernatant
(ml) and color developing agent (ml) (Sigma-Aldrich, Munich,
Germany) were mixed. The optical density (OD) value was measured at
15 min (Thermo Fisher Scientific). The 550 nm colorimetric NO
(mol/l) = (OD value of the test tube - OD value of the blank
tube)/(OD value of the standard tube - OD value of the blank tube)
× standard tube concentration (20 µmol/l) × dilution ratio before
sample testing.
Determination of activity of
intracellular SOD
Cell disruption solution (100 µl) was taken from
each well. The activity of intracellular SOD was determined
according to the instructions of the SOD kit (Nanjing Jiancheng
Bioengineering Institute, Nanjing, China). The formula was: SOD
activity (U/mg protein) in tissue = absorbance of the control tube
- absorbance of the test tube, absorbance of the control tube/50% ×
dilution ratio of the reaction system/protein content in the tissue
(mg/ml).
Expression of mRNA related to hepatic
cholesterol
All of the following reagents were from Qiagen, Inc.
(Valencia, CA, USA). A mass of 0.5 g of liver tissue was treated
with 1 ml TRIzol. The samples were vortexed and allowed to stand
for 5 min at room temperature. After addition of roughly 1/5 volume
of chloroform, samples were inverted, mixed for 1 min, allowed to
stand for 5 min at room temperature, and centrifuged (15 min, 6,000
× g, 4°C). After addition of an equivalent volume of isopropanol,
samples were inverted and mixed, allowed to stand for 10 min at
room temperature, and centrifuged (10 min, 6,000 × g, 4°C). The
supernatant was discarded. A volume of 1 ml of 75% ethanol was
added. An appropriate volume of DEPC water was added for intensive
dissolution and precipitation. The total volume of the reaction
system was 25 µl including: 20 µl fluorescent RT-PCR reaction
solution, 1 µl DNA polymerase, 0.35 µl reverse transcriptase, and 5
µl template RNA. The reaction mixtures were mixed well, and
centrifuged at 3,000 × g for 10 sec. The thermal profile was as
follows: Reverse transcription for 30 min at 50°C; pre-denaturation
for 3 min at 95°C; denaturation for 15 sec at 95°C; annealing for
30 sec at 50°C, extension for 30 min at 72°C, for five cycles in
total; denaturation for 10 sec at 95°C; and annealing for 40 sec at
55°C, for 40 cycles in total. Primer sequences are shown in
Table I.
 | Table I.Primer sequences. |
Table I.
Primer sequences.
| Gene name | Sequences |
|---|
| GAPDH | U:
5′-ATTCATGTGATCGACCATCC-3′ |
|
| D:
5′-CTCATCGCTGATCGGGTGAA-3′ |
| HMG-CoAR | U:
5′-CAATGTGCGTCACAGAATGT-3′ |
|
| D:
5′-CTTGGCGTACTCCTTGAACA-3′ |
| LDL-R | U:
5′-TGATCGATGTCGATGCGTGTACG-3′ |
|
| D:
5′-ATATGTGTCGATGCTCAGTCTTCC-3′ |
| CYP7A1 | U:
5′-GGATGCTACGTACGTGTGCTAGC-3′ |
|
| D:
5′-GGGTCTAGTCGTACGCTTGCGTC-3′ |
Statistical analysis
Data were analyzed with SPSS 16.0 statistical
software (SPSS, Inc., Chicago, IL, USA). One-way ANOVA test was
used for comparisons between groups. P<0.05 was taken as
statistically significant. P<0.01 was taken as highly
statistically significant.
Results
Effect of apigenin on parameters of
the high-fat model
There were no statistically significant differences
in body weight among the various groups before the experiments
(P>0.05). Body weight, hepatic coefficient, and AS index in the
model group were significantly higher than in the control group
(P<0.05). Body weight, hepatic coefficient, and AS index in the
apigenin treatment group decreased with increasing dose, indicating
that apigenin significantly reduced body weight and prevented
development of atherosclerosis. The total serum cholesterol of mice
in the model group was significantly higher than that in the normal
control group (P<0.01). Apigenin and simvastatin decreased the
levels of blood fat in mice. Serum TC of the mice in the apigenin
group decreased with increasing dose (P<0.01). The
concentrations of TG and LDL-C in serum of mice in the model group
increased significantly compared with the normal control group
(P<0.01). The concentrations of TG and LDL-C in sera of mice in
the model group were 1.34- and 2.56-fold those in the normal group.
The content of TC and TG in the liver of mice in the model group
increased significantly and the activity of SOD in serum decreased
significantly (P<0.01) compared with the normal control group.
This indicated that the high-fat diet raised the levels of hepatic
lipid in mice, abnormally increased lipid content, and inhibited
the activity of SOD in vivo. SOD activity in serum of mice
in the apigenin group increased by 20.95% compared with the model
group (P<0.01) (Table II).
| Table II.Effect of apigenin on body weight,
liver weight, AS index, TC, LDL-C, and SOD (mean ± SD) of the
high-fat model mice. |
Table II.
Effect of apigenin on body weight,
liver weight, AS index, TC, LDL-C, and SOD (mean ± SD) of the
high-fat model mice.
| Groups | Initial body
weight | Final body
weight | Liver weight | AS index | Initial TC | Final TC | TG | LDL-C | SOD |
|---|
| Control | 22.05±1.40 | 34.05±1.58 | 3.49±0.45 | 0.58±0.29 | 2.13±0.25 | 2.68±0.53 | 1.90±0.34 | 0.57±0.26 | 150.18±9.98 |
| Model | 22.68±2.01 |
36.94±2.07a |
4.08±0.40a |
1.04±0.58b | 2.30±0.46 |
4.53±1.19a |
2.56±0.78a |
1.78±0.78a |
113.56±21.09a |
| Low-dose
apigenin | 22.8±1.33 | 36.51±2.25 | 3.9±0.42 | 1.01±0.65 | 2.31±0.44 | 4.07±0.98 | 2.41±0.45 | 1.45±0.89 | 129.65±20.44 |
| Moderate-dose
apigenin | 23.78±1.81 | 36.91±1.74 | 3.9±0.4 | 1.05±0.67 | 2.35±0.24 | 4.34±0.81 | 2.56±0.87 |
1.50±0.73c | 125.34±18.65 |
| High-dose
apigenin | 22.58±2.00 |
34.97±2.03d | 3.79±0.36 |
0.67±0.33d | 2.39±0.34 |
3.29±0.89c |
2.14±0.54d |
0.80±9.54d |
127.38±20.90c |
| Simvastatin | 23.06±1.61 | 35.88±2.21 | 3.87±0.39 |
0.66±0.22d | 2.30±0.42 |
4.01±1.12c |
2.16±0.54d |
1.08±0.50c |
136.76±19.98d |
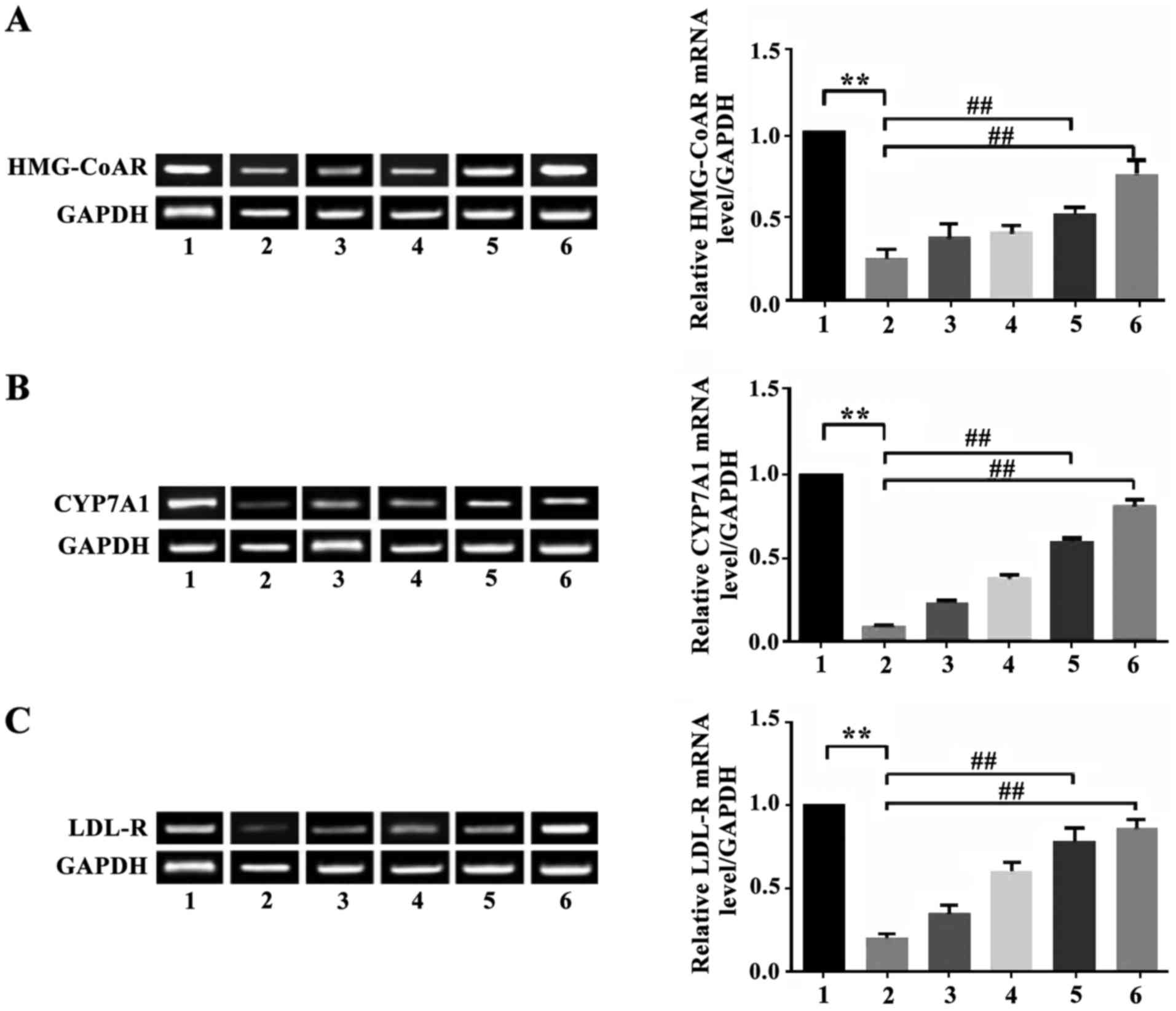
Effect of apigenin on expression of
HMG-CoAR, CYP7A1 and LDL-R mRNA in the liver of mice with
hyperlipidemia
The expression of HMG-CoAR mRNA in hepatic tissue of
mice in the model group decreased significantly (P<0.01)
compared with the normal control group. Apigenin was able to
upregulate the inhibited expression of HMG-CoAR mRNA. The
expression of HMG-CoAR mRNA in the liver of animals in the apigenin
high-dosage group and the simvastatin group increased significantly
compared with the model group (P<0.01). The expression of CYP7A1
mRNA in hepatic tissue of mice in the model group was significantly
lower than in the normal control group (P<0.01). Apigenin was
able to reverse the decreased expression level of CYP7A1 mRNA. The
expression of CYP7A1 mRNA in the liver of the apigenin high-dosage
group was significantly higher than in the model group (P<0.01).
The expression of LDL-R mRNA in the liver of mice of the apigenin
high-dosage group and the low-dosage group increased significantly
compared with the model group (P<0.01 and P<0.05). There were
no statistical differences in expression of LDL-R mRNA in the liver
between mice in the apigenin treatment group and the model group
(P>0.05) (Fig. 1).
Effect of apigenin on efflux of
cholesterol [3H] and cholesteryl ester/TC ratio
As shown in Table
III, 24-h incubation of THP-1 cells with apigenin significantly
promoted the efflux rate of [3H] cholesterol from THP-1
cells (P<0.01) in a dose-dependent manner. The efflux rate of
[3H] cholesterol in the cells of the apigenin
high-dosage group increased by 51.68% compared with the normal
group. The efflux rate of [3H] cholesterol in cells of
the apigenin moderate-dosage group increased by 31.88%. This
indicated that apigenin promoted the efflux of cholesterol in
macrophages and reduced the content of intracellular cholesterol.
This also indicated that apigenin was able to regulate cholesterol
metabolism in macrophages, increase the flow of cholesterol from
the inside to the outside of macrophages, and reduce the
accumulation of intracellular cholesterol. The content of
cholesteryl ester and the ratio of cholesteryl ester to TC
increased by 4.05-fold and 1.58-fold compared with the normal
group.
| Table III.Effect of apigenin on the efflux of
[3H] cholesterol in macrophages and the ratio of
cholesteryl ester/TC. |
Table III.
Effect of apigenin on the efflux of
[3H] cholesterol in macrophages and the ratio of
cholesteryl ester/TC.
| Groups | Dose (mol/l) | Cholesterol efflux
rate (%) | TC (mg/g) | Cholesteryl ester
(mg/g) | Cholesteryl
ester/TC (%) |
|---|
| Control | – |
8.76±1.34 | 11.58±1.03 |
4.08±0.76 | 32.56±2.34 |
| Model | – |
7.58±1.43 | 30.58±1.58 | 15.46±0.65 | 60.45±5.54 |
| High-dose
apigenin |
3×10−5 |
14.58±1.76b |
18.96±1.67b |
8.04±1.45 | 43.67±5.67 |
| Moderate-dose
apigenin |
3×10−6 |
11.23±1.34a |
22.34±1.56a |
10.76±0.54a |
50.87±3.45a |
| Low-dose
apigenin |
3×10−7 | 10.67±1.23 | 29.67±1.08 |
13.45±0.87b |
51.23±5.67b |
Effect of apigenin on NO and SOD
secretion by EA.hy926 cells injured by
H2O2
H2O2 oxidative damage
significantly decreased the levels of NO generated by EA.hy926
cells. The levels of NO secreted by the injured cells only reached
60.52% of the NO concentration of the normal control group
(P<0.01). Apigenin reversed the effect of
H2O2-injury in a dose-dependent manner and
raised the level of NO secreted by
H2O2-injured EA.hy926 cells. A concentration
of 1×10−5 mol/l apigenin caused the levels of NO
secreted by injured EA.hy926 cells to rise by 40.86% (P<0.01).
Based on comparison with the control group, incubation of EA.hy926
cells with H2O2 lead to a significant
decrease of 37.62% of intracellular SOD activity. Apigenin weakly
inhibited the proliferation of A10 vascular smooth muscle cells,
and there was a dose-dependent trend. However, there were no
significant differences in the inhibitory effects among the three
doses of apigenin (P>0.05). High-dose apigenin significantly
inhibited the proliferation of A10 cells (P<0.01) (Table IV) and the inhibitory effect was
dose-dependent.
| Table IV.Effect of apigenin on the activity of
NO and SOD secreted by H2O2-injured EA.hy926
cells. |
Table IV.
Effect of apigenin on the activity of
NO and SOD secreted by H2O2-injured EA.hy926
cells.
| Groups | Dose | Activity (%) | NO (µmol/l) | SOD (U/mg) |
|---|
| Control | – | – | 40.67±2.87 | 56.34±7.45 |
|
H2O2 | 75 µmol/l | – |
24.56±1.76a |
33.25±5.28a |
| High-dose
apigenin | 1×10−5
mol/l | 80.45c |
36.57±2.48c |
44.67±4.56b |
| Moderate-dose
apigenin | 1×10−6
mol/l | 40.19c | 22.56±2.16 | 40.15±3.40 |
| Low-dose
apigenin | 1×10−7
mol/l | 9.89 | 19.45±1.45 | 33.67±1.34 |
Discussion
In the present study, the high-fat diet mouse model
of hyperlipidemia was reproduced. The levels of TC, TG, and LDL-C
in serum of the model mice increased significantly at 4 weeks
exposure to a high-fat diet, indicating successful establishment of
the model of hyperlipidemia. The results indicated that apigenin
reduced the content of TC, TG, and LDL-C in mice in a
dose-dependent manner. Administration of apigenin significantly
decreased body weight; concentrations of TC, TG, and LDL-C in
serum; and the content of TC and TG, suggesting that apigenin
effectively inhibited rising levels of blood lipid, improved
disorder of cholesterol metabolism, mitigated accumulation of
hepatic lipid, and recovered the balance of lipid metabolism. The
data suggest that apigenin can help prevent and treat
atherosclerosis and decrease the incidence of cardiovascular and
cerebrovascular diseases.
To understand the mechanism of action of apigenin in
reducing blood fat, the mRNA levels corresponding to three key
proteins involved in cholesterol metabolism (HMG-CoAR, CYP7A1, and
LDL-R) were examined by RT-PCR. HMG-CoAR is a rate-limiting enzyme
for cholesterol synthesis in the liver. CYP7A1 is a key enzyme
which transforms cholesterol into bile acid in liver (19). LDL-R is an important receptor that
mediates liver uptake of LDL-C (20). It was shown in the present study that
the mRNA levels of HMG-CoAR, CYP7A1, and LDL-R in the liver of mice
of the model group decreased significantly following exposure to a
high-fat diet compared with those in the normal control group. The
results are consistent with other studies in the literature. The
results indicate that administration of apigenin can cause the
expression levels of these genes to approximate normal levels
(21).
During the development of atherosclerosis, oxidative
stress induces the expression of vascular adhesion molecules,
promotes local inflammatory reactions, and induces cell
proliferation (22). Hyperlipidemia
can increase the amount of oxygen free radicals generated in
vivo (23), increase oxidative
stress (24) and is involved in the
occurrence and progression of atherosclerosis. It was shown in the
present study that SOD activity of mice in the model group
decreased and the antagonistic effect against oxidative stress was
inhibited. Apigenin can increase SOD activity, increase
anti-oxidative capacity in vivo, reduce the amount of
oxidative low-density lipoprotein produced, and inhibits
development of atherosclerosis.
The above results indicated that apigenin
significantly decreased the levels of TC, TG, and LDL-C in serum of
mice with hyperlipidemia and reduced the accumulation of TC and TG
in the liver. Apigenin may decrease the content of cholesterol by
promoting hepatic LDL-C absorption and increasing the
transformation of hepatic cholesterol into bile acid. The present
study also showed that apigenin can improve the dysregulated lipid
balance and can potentially be used to treat diseases such as
atherosclerosis and fatty liver. The results of the present study
provide a foundation for further research on the mechanism of
action of apigenin in regulating cholesterol metabolism.
References
|
1
|
Liu-Smith F and Meyskens FL: Molecular
mechanisms of flavonoids in melanin synthesis and the potential for
the prevention and treatment of melanoma. Mol Nutr Food Res.
60:1264–1274. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yarla NS, Bishayee A, Sethi G, Reddanna P,
Kalle AM, Dhananjaya BL, Dowluru KS, Chintala R and Duddukuri GR:
Targeting arachidonic acid pathway by natural products for cancer
prevention and therapy. Semin Cancer Biol. Feb 4–2016.(Epub ahead
of print). doi: 10.1016/j.semcancer.2016.02.001. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Moosavi F, Hosseini R, Saso L and Firuzi
O: Modulation of neurotrophic signaling pathways by polyphenols.
Drug Des Devel Ther. 10:23–42. 2015.PubMed/NCBI
|
|
4
|
Venigalla M, Sonego S, Gyengesi E, Sharman
MJ and Münch G: Novel promising therapeutics against chronic
neuroinflammation and neurodegeneration in Alzheimer's disease.
Neurochem Int. 95:63–74. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Venigalla M, Gyengesi E and Münch G:
Curcumin and apigenin - novel and promising therapeutics against
chronic neuroinflammation in Alzheimer's disease. Neural Regen Res.
10:1181–1185. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shakeri F and Boskabady MH: A review of
the relaxant effect of various medicinal plants on tracheal smooth
muscle, their possible mechanism(s) and potency. J Ethnopharmacol.
175:528–548. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sak K: Cytotoxicity of dietary flavonoids
on different human cancer types. Pharmacogn Rev. 8:122–146. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Maurya SK, Kushwaha AK and Seth A:
Ethnomedicinal review of Usnakantaka (Echinops echinatus
Roxb.). Pharmacogn Rev. 9:149–154. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Armstrong CM and Gao AC: Drug resistance
in castration resistant prostate cancer: resistance mechanisms and
emerging treatment strategies. Am J Clin Exp Urol. 3:64–76.
2015.PubMed/NCBI
|
|
10
|
Shay J, Elbaz HA, Lee I, Zielske SP, Malek
MH and Hüttemann M: Molecular mechanisms and therapeutic effects of
(−)-epicatechin and other polyphenols in cancer, inflammation,
diabetes, and neurodegeneration. Oxid Med Cell Longev.
2015:1812602015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Johnston GA: Flavonoid nutraceuticals and
ionotropic receptors for the inhibitory neurotransmitter GABA.
Neurochem Int. 89:120–125. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Guedj F, Bianchi DW and Delabar JM:
Prenatal treatment of Down syndrome: a reality? Curr Opin Obstet
Gynecol. 26:92–103. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fernando W, Rupasinghe HP and Hoskin DW:
Regulation of hypoxia-inducible factor-1α and vascular endothelial
growth factor signaling by plant flavonoids. Mini Rev Med Chem.
15:479–489. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li A, Sun A, Liu R, Zhang Y and Cui J: An
efficient preparative procedure for main flavonoids from the peel
of Trichosanthes kirilowii Maxim. using polyamide resin
followed by semi-preparative high performance liquid
chromatography. J Chromatogr B Analyt Technol Biomed Life Sci.
965:150–157. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nabavi SM, Habtemariam S, Daglia M and
Nabavi SF: Apigenin and Breast Cancers: from Chemistry to Medicine.
Anticancer Agents Med Chem. 15:728–735. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gupta SC, Tyagi AK, Deshmukh-Taskar P,
Hinojosa M, Prasad S and Aggarwal BB: Downregulation of tumor
necrosis factor and other proinflammatory biomarkers by
polyphenols. Arch Biochem Biophys. 559:91–99. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cochran DB, Gray LN, Anderson KW and
Dziubla TD: Degradable poly(apigenin) polymer inhibits tumor cell
adhesion to vascular endothelial cells. J Biomed Mater Res B Appl
Biomater. Aug 6–2015.(Epub ahead of print). doi:
10.1002/jbm.b.33486. PubMed/NCBI
|
|
18
|
Mastantuono T, Battiloro L, Sabatino L,
Chiurazzi M, Di Maro M, Muscariello E, Colantuoni A and Lapi D:
Effects of citrus flavonoids against microvascular damage induced
by hypoperfusion and reperfusion in rat pial circulation.
Microcirculation. 22:378–390. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Alig SK, Stampnik Y, Pircher J, Rotter R,
Gaitzsch E, Ribeiro A, Wörnle M, Krötz F and Mannell H: The
tyrosine phosphatase SHP-1 regulates hypoxia inducible factor-1α
(HIF-1α) protein levels in endothelial cells under hypoxia. PLoS
One. 10:e01211132015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zargaran A, Borhani-Haghighi A, Faridi P,
Daneshamouz S, Kordafshari G and Mohagheghzadeh A: Potential effect
and mechanism of action of topical chamomile (Matricaria
chammomila L.) oil on migraine headache: a medical hypothesis.
Med Hypotheses. 83:566–569. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen X, Jin J, Chen Y, Peng L, Zhong G, Li
J, Bi H, Cai Y and Huang M: Effect of scutellarin on the metabolism
and pharmacokinetics of clopidogrel in rats. Biopharm Drug Dispos.
36:64–68. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Seo HS, Ku JM, Choi HS, Woo JK, Jang BH,
Go H, Shin YC and Ko SG: Apigenin induces caspase-dependent
apoptosis by inhibiting signal transducer and activator of
transcription 3 signaling in HER2-overexpressing SKBR3 breast
cancer cells. Mol Med Rep. 12:2977–2984. 2015.PubMed/NCBI
|
|
23
|
Chen YJ, Wang L, Zhou GY, Yu XL, Zhang YH,
Hu N, Li QQ, Chen C, Qing C, Liu YT, et al: Scutellarin attenuates
endothelium-dependent vasodilation impairment induced by hypoxia
reoxygenation, through regulating the PKG signaling pathway in rat
coronary artery. Chin J Nat Med. 13:264–273. 2015.PubMed/NCBI
|
|
24
|
Lee W, Yoon EK, Kim KM, Park DH and Bae
JS: Antiseptic effect of vicenin-2 and scolymoside from Cyclopia
subternata (honeybush) in response to HMGB1 as a late sepsis
mediator in vitro and in vivo. Can J Physiol Pharmacol. 93:709–720.
2015. View Article : Google Scholar : PubMed/NCBI
|